Abstract
The occurrence of heavy metals in aquatic ecosystems is a serious environmental hazard, and their effective removal is imperative. In this regard, the feasibility of living microalga Chlorella vulgaris (C. vulgaris) to remove heavy metals (Ni, Pb, Zn, Cd, and Cu) is investigated by using 1, 5, and 10 ppm concentrations of single- and multiple-metal-treated (MT) cultures. Experiments were performed in controlled laboratory conditions, and metal removal analysis was performed through atomic absorption spectroscopy (AAS). The cultures were also examined by means of optical microscopy, UV-Vis spectrophotometry, and Fourier transform infrared (FTIR) spectroscopy to follow the cultures’ pigment content, cell population, and functional group changes during cultivation. The removal efficiency results of both single and multiple MT cultures were evaluated using the Langmuir isotherm model. The results indicate that C. vulgaris presents potential for heavy metal bioremediation, even towards multi-MT conditions, despite the influence of a competitive uptake in multi-MT cultures. In mono-MT cultures, the removal efficiency of C. vulgaris presents values of 65–99% on Day 3 and 72–99% on Day 7 of cultivation, while the results for the multi-MT cultures are 49–99% and 62–99% for Days 3 and 7 of cultivation, respectively. The research illustrates the potential for C. vulgaris as a promising biosorbent for heavy metal remediation along with its post-treatment use in applications supporting the green circular economy.
1. Introduction
Although heavy metals are frequently associated with pollution, they are naturally occurring components in the environment. However, their buildup in excessive amounts has remarkable effects on environmental and health-related risks. While trace quantities of some heavy metals are important for certain biological functions, human activities have magnified their presence rather significantly, to toxic levels in some cases [1,2]. Activities in the industrial sector, such as mining and metal production, and in electronic production industries, along with agriculture using high amounts of fertilizers and pesticides, are some of the major causes of heavy metal contamination in inland waters [1,2,3,4,5]. They persist in the environment, bioaccumulate in organisms, and biomagnify up the food chain, especially in aquatic life. Thus, they cause various acute health disorders, including neurological ones from lead, kidney damage from cadmium, and carcinogenic effects from nickel [1,6,7]. To mitigate these risks, regulatory bodies such as the European Parliament and the Council have established limits for heavy metals in drinking water, surface water, and wastewater to safeguard human health and the environment [8].
Apart from their bioaccumulation, which may cause serious health problems, aquatic ecosystems and biodiversity are threatened too [9]. Although steps have been taken to limit heavy metal contamination, existing remediation technologies often face challenges due to high costs and limited efficiency [10,11]. To find a practical solution to this problem, there is a pressing necessity for innovative, sustainable, and economical solutions [11,12]. To this direction, microalgae have gained increasing attention as biosorbents for heavy metal removal due to their efficiency, as well as their affordability, availability, and ability to function effectively in both living and non-living states [2,12,13,14,15].
Microalgae have shown significant potential as biosorbents for removing heavy metals from contaminated environments due to their ability to bind and immobilize metal ions through various mechanisms [2,16,17,18,19]. This ability provides a sustainable and eco-friendly alternative to traditional methods, which are typically costly and generate secondary waste [20]. Among the various microalgal species, Chlorella vulgaris (C. vulgaris), Spirulina (Arthrospira) platensis, Scenedesmus obliquus, and Nannochloropsis oculata are particularly notable for their high heavy metal uptake efficiency. For example, C. vulgaris is widely recognized for its ability to adsorb, among others, cadmium, lead, and copper [14,21,22,23,24,25,26,27]. These microalga species, used both as dried biomass for adsorption and also alive for in vivo bioremediation, exhibit high metal-binding capacities, making them ideal candidates for wastewater treatment and environmental restoration. Their versatility, combined with advancements in bioremediation technologies, highlights their effectiveness as cost-efficient and sustainable solutions for heavy metal pollution [2,14,21].
C. vulgaris is a single-cell microalga valued for its high content in proteins (45–60%), lipids (10–20%), carbohydrates (10–20%), vitamins, and essential minerals, making it widely used in the food, pharmaceutical, biodiesel, and cosmetic industries [28,29]. It should also be noted that its cultivation significantly contributes to mitigating greenhouse gas emissions by assimilating CO2 from the atmosphere [17,30,31,32,33].
C. vulgaris stands out among microalgae for heavy metal removal due to its fast growth, high biomass yield, and ability to increase in diverse conditions, including its tolerance of high metal concentrations. Unlike many other species, it combines passive adsorption on the cell surface with active intracellular uptake, enhancing its metal removal efficiency. All the above factors, along with its added value as a product, make it a cost-effective and sustainable option for bioremediation [34,35].
Heavy metals accumulation in C. vulgaris is achieved through passive and active processes, thus allowing for the efficient binding and sequestration of toxic metal ions from contaminated environments [2,21,36]. The key mechanisms involved in the uptake and detoxification process of metals include biosorption, bioaccumulation, intracellular sequestration, and biotransformation. The passive biosorption processes of C. vulgaris on its cell surface are accomplished through the interaction of functional groups such as carboxyl, hydroxyl, sulfate, and amine through ion exchange and complexation in an electrostatic mode of attraction with the aforementioned heavy metal ions [16,29]. Because of the high affinities between C. vulgaris’s cell wall component and metal ions, the microalga functions as a good biosorbent [37]. Bioaccumulation is the active process of transporting metal ions across the cell membrane, where they accumulate in the cytoplasm. Essentially, this process involves the use of metal transporters and intracellular binding proteins to help regulate metal ion concentrations inside cells to non-toxic levels [14,17,27,38,39]. Within the cell, heavy metals are generally detoxified through their binding to intracellular molecules such as metallothioneins and phytochelatins, which constitute stable complexes with the metal ions. In this way, cytotoxic damage is avoided, thus granting the microalga with the capacity to bear and survive in a metal-contaminated environment. In addition to that, some metals are also compartmentalized within the vacuoles, further reducing their toxicity [40]. C. vulgaris can also convert toxic metal ions into less harmful or insoluble forms by utilizing various mechanisms for the detoxification of heavy metals. Enzymatic reduction is one such mechanism, whereby toxic metal ions are reduced to less toxic ions by specific enzymes. For instance, copper ions (Cu2⁺) can be reduced to Cu⁺, which is less toxic and easier for the algal cells to handle. Another vital mechanism involves precipitation, where metals like Cd or Cu precipitate as insoluble compounds, i.e., CdS or CuS, that become trapped in the cells or are released, thereby reducing their toxic effect on cellular structures [41,42]. These processes aid in the long-term sequestration of heavy metals in aquatic and terrestrial environments [43].
Several studies have examined C. vulgaris removal efficiency for mono-metal-treated (MT) cultures [24,25,26,44,45,46,47]. In contrast, although mixtures of metals better reflect real-world scenarios in ecosystems and wastewater, studies on multi-MT cultures remain limited, not only in the case of C. vulgaris [3,14,17] but also regarding other microalgae, such as Spirulina platensis [14,48,49,50] and Desmodesmus multivariabilis [51]. Mono-MT cultures cannot always be used for the prediction of multi-MT cultures because of the simultaneous synergistic or antagonistic effects that may occur [37]. Most natural ecosystems have a complex mixture of different metals, creating rivalries for binding sites and hence complicating the removal process.
The present study investigates the heavy metal bioremediation efficiency of C. vulgaris in multi-MT cultures under controlled laboratory conditions, as the microalga’s uptake ability during multi-metal presence remains less explored. This covers both (a) the use of live C. vulgaris cultures and (b) the efficiency of C. vulgaris as a sorbent for metal mixtures. The hypothesis tested concerns the fact that the simultaneous presence of multiple metals could influence their removal due to antagonistic or synergistic effects, thus leading to different removal efficiencies between mono- and multi-MT conditions. Atomic absorption spectroscopy (AAS), optical microscopy, UV-Vis spectrophotometry, and Fourier transform infrared (FTIR) spectroscopy were the methods performed to check the metal removal efficiency of C. vulgaris, the effect of the induced heavy metal stress, and the changes induced in the microalga cultures to assess the possibility of using them for other applications after bioremediation in a circular economy framework [52].
2. Materials and Methods
2.1. Microalga Cultivation
C. vulgaris (strain NIVA-CHL 108) was purchased by NORCCA (Oslo, Norway) and cultivated using Bold’s Basal Medium (BBM) [53], as described in [54]. In brief, BBM is composed of ten (10) stock solutions (SLs) (Table 1) that are mixed with distilled water for a final volume of 1 L. All the BBM ingredients were procured from Sigma-Aldrich, Carl Roth, Honeywell/Fluka, Merck, and PanReac Applichem. BBM presented pH (pH-GL21 m, Crison Instruments, Barcelona, Spain) and electrical conductivity (Orion Star™ A212, Thermo Fisher Scientific, Waltham, MA, USA) values of 6.7 ± 0.1 and 1.35 ± 0.05 mS·cm−1, respectively, in accordance with the literature [54].

Table 1.
Bold’s Basal Medium (BBM) composition [53,54].
The microalga, as an initial stock culture, was axenically cultivated using the produced BBM at 20 ± 0.5 °C using a water bath, with the illumination of ~160 μmol photons m−2s−1 of photosynthetically active radiation (PAR) over successive 12 h light/12 h dark periods (full spectrum LED array ATTIS 200 W, LUMATEK, London United Kingdom). Finally, the culture was treated with a continuous aeration of ~1 L/min.
2.2. Single- and Multi-Metal Treatment of C. vulgaris
Solutions of heavy metals were prepared in distilled water using the following nitrate salts of each metal: Pb(NO3)2, Carl Roth; Cu(NO3)2·3H2O, Sigma-Aldrich; Zn(NO3)2·4H2O, Chem-Lab NV; Ni(NO3)2·6H2O, Sigma-Aldrich; and Cd(NO3)2·4H2O, Thermo Scientific.
For the single- and multi-metal in vivo treatment of C. vulgaris, 250 mL of the initial microalga stock culture, with a cell concentration of (12.0 ± 0.5) × 105 cells/mL, was introduced to 19 Erlenmeyer flasks of 500 mL, and 50 mL of BBM was added.
The heavy metal treatment followed the same concentrations as in Malletzidou et al. [50]. In particular, 1, 5, and 10 ppm of each metal were used for the mono-MT cultures, while for the multi-MT cultures, three flasks contained all metals in concentrations of 1 ppm (Mix 1), 5 ppm (Mix 5), and 10 ppm (Mix 10) of each metal. Additionally, one of the flasks was prepared without metal treatment, acting as the Control culture.
Finally, after the introduction of the initial culture, the BBM, and the heavy metal solution, each flask was filled with distilled water up to a final volume of 0.5 L.
All C. vulgaris cultures (metal-treated and Control) were cultivated with the same temperature, aeration, and illumination conditions for 7 days, as described in Section 2.1 [3,27]. All measurements were performed twice, and the results are presented as their corresponding averages.
2.3. Estimation of Culture Biomass
The estimation of the microalga cell concentration and the daily examining of the cells’ health state, in terms of morphology, were performed in an improved Neubauer counting chamber using a microscope equipped with a digital camera (CX43RF microscope and EP50 camera, Olympus Corporation, Tokyo, Japan). Samples (1 mL) were collected and diluted with distilled water (1:10) before the measurements, where 10 μL of the diluted cultures were placed on an improved Neubauer hemocytometer after applying a cover glass [53]. Images were collected using 10× and 60× objective lenses. The C. vulgaris cell counts were performed with the online counting tool at http://chaos.athenarc.gr:31304/index (accessed on 10 October 2024), which uses the microalga datasets created by Pistolas et al. [53].
2.4. Pigment Content Estimation
C. vulgaris contains chlorophylls-a and -b as its major photosynthetic pigments [55], together with carotenoids [56]. The pigment content of the microalga was estimated using a UV-Vis spectrophotometer (Perkin Elmer Lambda 25, Waltham, MA, USA) by applying 60 nm/min scan speed at 400–800 nm. Culture samples of 4 mL were gathered on Days 0, 3, and 7 of cultivation. They were centrifuged for 5 min at 4000 rpm (servo spin NECT centrifuge); their supernatant was removed, and 10 mL of methanol was added. They remained in a vortex for 1 min, which was followed by 10 min in an ultrasound bath at 90 °C. After a final centrifugation for 5 min at 4000 rpm, the supernatant was measured using a QG 10 mm cuvette (3 mL). Methanol, which was placed in a similar cuvette, was used as the blank.
When methanol is used as the extraction solvent, chlorophyll-a and -b present their absorption maxima at 665.2 and 652.4 nm, respectively [57], and the equations for determining the contents of chlorophyll-a (Chl.a), chlorophyll-b (Chl.b), total chlorophylls (Chl.tot), and total carotenoids (Chl.x) in μg/mL are [57]:
where A represents the measured absorbance values at different wavelengths, after the subtraction of the reading at 750 nm to eliminate turbidity [58].
2.5. Fourier Transform Infrared Spectroscopy
Fourier transform infrared (FTIR) spectroscopy in attenuated total reflectance (ATR) mode was applied to the dried microalga biomass to assess any functional group differentiations induced in cells by the heavy metals. For this reason, a diamond ATR accessory (45° angle of incidence), model GladiATR, PIKE Technologies (Madison, WI, USA), attached to a Cary 670 FTIR spectroscope (Agilent Technologies Inc., Palo Alto, CA, USA) was used, and the measurements were applied with 32 accumulative scans at a resolution of 4 cm−1 in the spectral area from 4000 to 400 cm−1. Microalga culture volumes (50 mL) were collected on Day 7 of cultivation, centrifuged (5 min at 5000 rpm) with Multifuge X1 Pro-MD (Thermo Fisher Scientific, Waltham, MA, USA), and left to dry in a desiccant before the measurements.
2.6. Atomic Absorption Spectroscopy (AAS)
The microalga’s removal efficiency on Days 3 and 7 of cultivation was evaluated from the metals’ residual concentration in each culture medium using atomic absorption spectroscopy (AAS) (AAnalyst 800, Perkin–Elmer, Waltham, MA, USA). For this reason, 10 mL from each culture was centrifuged (5 min at 5000 rpm); the supernatant was used for the AAS measurements. The removal was estimated in terms of removal efficiency (R):
where C0 is the initial metal concentration at Day 0 and Ct is the metal concentration at time t during cultivation.
The spectroscope was configured to analyze the heavy metals under study by selecting the appropriate wavelength for each one (213.9 nm for zinc, 283.3 nm for lead, 324.8 nm for copper, 232.0 nm for nickel, and 228.8 nm for cadmium). Finally, the flame (air–acetylene) was fine-tuned to optimize the atomization of the sample, ensuring precise detection.
2.7. Application of the Langmuir Isotherm Model to Mono- and Multi-MT Cultures
The Langmuir isotherm two-parameter model was applied to the mono- and multi-MT cultures to describe the metal removal process by C. Vulgaris. Although the adsorption isotherms can be approached by various models, such as those introduced by Langmuir, Freundlich, and Temkin [18,51,59,60,61,62], the Langmuir isotherm model was used in the present study as an attempt to describe the metal removal process by C. Vulgaris; since it is a simple model, it is frequently used as a basis for microalgae sorption capacity definition [18,60]. According to the Langmuir isotherm model for single components, it is assumed that the biosorbent is homogenous and has sites with equal affinity for the sorbate, and a monolayer is created upon saturation [14,26,60,63,64]. According to the model, the equation that describes the metal biosorption capacity and concentration at equilibrium is
where qe is the amount of the adsorbed metal at equilibrium (mg/g), qm is the maximum adsorbed metal capacity (mg/g), KL is the Langmuir adsorption constant (L/mg), and Ce is the metal concentration in the culture (mg/L) [65]. In order to express the microalga biomass in terms of cells instead of mass, the population was used (N, cells/L). Following this, the amount of the adsorbed metal (q′e) is now expressed as
The modified Langmuir equation is now expressed as
where q′e and q′m are the amount of the absorbed metal at equilibrium and the maximum metal uptake, respectively, now expressed in mg/cell. Following this, the modified Langmuir equation has the linearized form of
By plotting 1/q′e versus 1/Ce and applying linear regression, the slope results in the calculation of KL and q’m for every metal in the mono-MT cultures.
To assess the multiple MT cultures, the multi-component non-modified Langmuir isotherm was applied [14,66]. To calculate the sorption capacity of every sorbate/metal qe,i (mg/cell) at equilibrium, the following equation was applied:
This multi-component model uses the q′m,i and KL,i parameters, as derived from the single Langmuir isotherm model for every metal, as all cultures/experiments were performed under the same conditions.
3. Results and Discussion
3.1. Cultures’ Monitoring During Cultivation
Figure 1 presents the normalized population growth curves of the mono- and multi-MT cultures of C. vulgaris, depicting the concentration toxicity profile depending on the specific metal and its concentration.
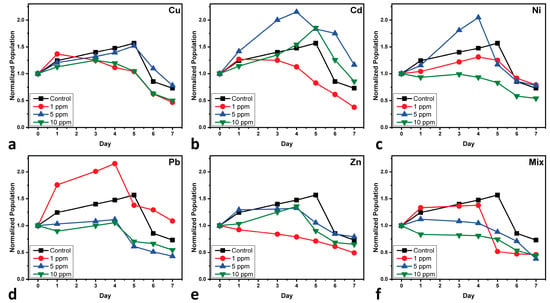
Figure 1.
Normalized population growth curves of C. vulgaris cultures after the treatment of (a) copper-MT, (b) cadmium-MT, (c) nickel-MT, (d) lead-MT, (e) zinc-MT, and (f) multi-MT. Normalization is performed at Day 0 of cultivation for minor differences in the initially used biomass. Each value represents the mean of two separate measurements, with each one having an uncertainty of less than 2.5%.
In the case of Cu (Figure 1a), the population of C. vulgaris exhibits the most significant decline at 1 ppm, indicating a higher toxicity effect compared to 5 and 10 ppm. Interestingly, the population at 10 ppm remains relatively low, while the 5 ppm condition shows results similar to the Control culture. This suggests a potential hormetic response, where low concentrations of a stressor can have more detrimental effects than moderate levels. Copper can create oxidative stress in cells by inducing reactive oxygen species (ROS), which damage cellular components such as lipids, proteins, and DNA [21]. While C. vulgaris can trigger antioxidant defenses, like superoxide dismutase (SOD) and catalase (CAT) [67], high concentrations of Cu overwhelm these protective mechanisms, leading to reduced cell viability. Copper also disrupts the photosynthetic machinery by replacing essential metal cofactors in enzymes, further inhibiting cell growth [68].
Cd (Figure 1b) concentrations of 5 and 10 ppm follow similar behavior with the Control sample; the exponential phase lasts up to Days 4–5, after which the decline phase follows. On the other hand, 1 ppm presents a very short exponential phase, and its decline seems to start between Days 2–3. This behavior of high toxicity effects of low doses could be attributed to the self-protective mechanisms of the microalga against high doses, where it adapts against damage; in this scenario, low doses do not trigger such mechanisms [69]. The observed increase in C. vulgaris population at 5 and 10 ppm of Cd can be attributed to the hormesis effect. Hormesis refers to a biphasic dose–response where low to moderate exposure to a stressor can stimulate growth, while higher concentrations become inhibitory [70]. This phenomenon has been previously documented in algae exposed to Cd, where sublethal concentrations enhance growth by inducing antioxidant responses, activating stress-related enzymes, and promoting nutrient uptake. Furthermore, C. vulgaris has effective detoxification mechanisms, including the binding of Cd to phytochelatins and metallothioneins, which mitigate the toxic effects of the metal [41]. This adaptive response of C. vulgaris at higher Cd concentrations contrasts with its reaction to other metals, where no such growth enhancement is observed at similar levels. This response difference can be attributed to specific metal uptake, sequestration, and detoxification pathways, as previously reported in microalgae under heavy metal stress. A study on the toxicity of cadmium to C. vulgaris found that high concentrations of cadmium could stimulate certain photosynthetic parameters, while lower concentrations led to inhibition, indicating a complex dose–response relationship [71].
The growth of C. vulgaris under varying Ni (Figure 1c) concentrations exhibits distinct patterns. At concentration of 5 ppm, the population experiences significant growth that surpasses the Control culture until Day 5, after which it declines sharply. At 1 ppm, growth remains below the level of the Control culture, though some development is observed. In contrast, at 10 ppm, the population is reduced from the outset. The toxicity induced by Ni can be attributed to its capacity to substitute essential metals in metalloenzymes, disrupting metabolic pathways. At specific concentrations, this disruption may initially stimulate certain physiological processes, leading to temporary growth enhancement, followed by a decline as toxicity effects accumulate [37].
The phenomenon of hormesis could be used to explain why low concentrations of Pb stimulate the growth of C. vulgaris (Figure 1d). It is a biphasic dose–response relationship wherein a toxic substance at a low concentration exerts a stimulatory effect but becomes inhibitory at higher levels. In microalgae, low doses of Pb may stimulate cellular metabolism by inducing antioxidant defenses, enhancing photosynthetic efficiency, or increasing enzyme activity responsible for nutrient uptake [72]. Some studies have suggested that at low levels, Pb may also induce stress responses that enhance algal adaptation mechanisms, leading to temporary growth stimulation [69]. However, with increasing Pb concentrations beyond the threshold level, the aforementioned benefits are outweighed by oxidative damage, enzyme inhibition, and membrane disruption, culminating in toxicity and growth inhibition [68]. This hormetic response clearly underlines the intricacy of metal–microalgae interaction and justifies the relevance of concentration-dependent toxicity assessments.
Regarding the effects of Zn (Figure 1e) on the growth of microalga at 1 ppm, a decline is observed from Day 1. At 10 ppm, growth increases until Day 4, followed by a sharp decrease. At 5 ppm, growth is closer to the Control culture, but all three Zn-MT cultures remain below the level of the Control culture. Low concentrations can cause immediate toxic effects, while moderate concentrations may initially stimulate growth due to zinc’s role as an essential micronutrient before toxicity effects dominate over time. Higher concentrations tend to be toxic from the outset, leading to reduced growth. Studies have shown that low zinc concentrations can enhance growth and metabolic activity in microalgae, whereas higher concentrations exhibit inhibitory effects [73].
The microalga exposed to a mixture of heavy metals during cultivation (Figure 1f) exhibits distinct patterns: at 1 ppm, there is an increase until Day 4, followed by a sharp decline to levels lower than all other treatments; at 5 ppm, a very slight increase is observed before a decrease; and at 10 ppm, a reduction occurs from the beginning of the cultivation. These responses could be ascribed to the combined effects of heavy metals, which can interact synergistically or antagonistically, influencing algal growth. Low concentrations might initially stimulate growth due to a hormetic effect, but as exposure continues, the cumulative toxicity leads to a sharp decline. Higher concentrations likely impose immediate stress, inhibiting growth from the outset. The toxicity of a specific metal is influenced not just by its chemical speciation, as other chemical compounds and biological interactions also have a significant impact in the final toxicity. The multi-MT cultures present the highest growth reduction, thus indicating synergistic toxicity, wherein the combined effects of multiple metals exceed their individual toxicities. This may be due to enhanced oxidative stress, competition for cellular uptake, and the cumulative disruption of essential biochemical pathways [74]. These observations also point out the complexity of heavy metal toxicity in microalgae, where low doses of some metals may give rise to hormetic effects, while others are potent growth inhibitors.
Figure 2 shows representative images captured using optical microscopy on Day 7 of cultivation, illustrating the qualitative effects of heavy metal exposure (Cu, Cd, Ni, Pb, and Zn mono- and multi-MT cultures) on C. vulgaris. The influence of heavy metals on the growth of the microalga is noticeable, showing a decrease in cell density, the formation of cell aggregates, and morphological changes compared to the Control culture. During the cultivation of the Control culture, the cells show normal morphology, intact cell membrane, tightly packed pigmentation, and typical green color from healthy photosynthesis. In the Zn-MT of 5 ppm, cell number slightly decreases, which may be due to the inhibitory effect of zinc on C. vulgaris metabolism, as it inhibits the photosynthesis progress and protein synthesis [75]. Also, some cells are more transparent, which may suggest cellular stress and structural damage, possibly due to increased membrane penetrability and the damage of intracellular components [68]. In the Pb-MT culture of 10 ppm, cell density decreases far more, indicating increased lead toxicity, which triggers oxidative stress, cell membrane damage, and photosynthesis progress inhibition. Regarding the multi-MT cultures, a dose-dependent decrease in cell density appears, suggesting synergistic toxicity of the metals to the cells. This effect results from mechanisms such as reduced production of ROS through photosynthesis, leading to oxidative stress, and the interference of cell division, which affects the growth rate [76]. The presence of more transparent cells in the multi-metal treatments supports the hypothesis of severe cellular damage. Heavy metal exposure promotes the growth of cell aggregates, a common stress response in microalgae, possible as a protection mechanism to reduce metal exposure [77]. Generally, the results show that C. vulgaris cells are sensitive to heavy metals. The observed effects include damage to key cellular structures such as the cell wall, chloroplasts, and mitochondria; an increase in cell aggregates; and the presence of transparent cells, verifying the negative effect of metals on the cells [7,69,75,78].

Figure 2.
Optical microscopy images of representative cultures on Day 7 of cultivation; images were captured with a 60× objective lens. The samples were diluted with distilled water before observation.
3.2. Pigments
The chlorophyll and carotenoid contents of microalga serve as key indicators of photosynthetic efficiency, plant health, and responses to environmental stress, as the induction of oxidative stress through the membrane inhibits damage to various photosynthetic pigments [79]. Changes in chlorophyll levels reflect the ability of microalgae to adapt to diverse environmental conditions, including exposure to heavy metals [80]. Heavy metals can disrupt chlorophyll synthesis, ultimately reducing biomass production [78]. Monitoring chlorophyll content helps assess the impact of pollutants on C. vulgaris cells [81]. Carotenoids, on the other hand, are included in the factors that prevent degenerative stress due to their photoprotective role against high ROS levels [79].
Figure 3 shows the photosynthetic pigment content of all cultures on Days 0 and 7 of cultivation, calculated using Equations (1)–(4). Specifically, the first column bar of each graph represents the pigment levels at Day 0, showing that all cultures begin with the same initial pigment level. The chlorophyll and carotenoid contents of C. vulgaris are significantly affected by exposure to heavy metals [77]. As observed, all calculated pigment concentrations exhibit comparable trends, indicating that any of them can be used to describe pigment behavior under metal exposure. Copper at lower concentrations appears to enhance all the photosynthetic pigment contents under examination due to its role as a cofactor in photosynthetic enzymes [82]. At higher Cu concentrations (5 and 10 ppm), pigment levels decrease, indicating damage to the photosynthesis process [67]. Similar trends, where increasing concentrations reduce the pigment content, are observed for cadmium and lead, with pigment degradation due to metal-induced stress [80]. Nickel enhances pigment synthesis, but higher concentrations are harmful, disrupting essential metabolic pathways [81]. Zinc treatments show a smaller effect on the pigment content due to its role in stabilizing photosynthetic proteins. The multi-MT cultures (Mix) result in a pronounced reduction in pigments, reflecting collective toxicity effects. These findings highlight the dual role of heavy metals as both essential micronutrients and potential toxicants, depending on the heavy metal concentration. The apparent discrepancy between population growth (Figure 1) and the photosynthetic pigment content (Figure 3) in the Ni-treated C. vulgaris cultures results in the varied physiological responses that these parameters represent. While Figure 1 represents total population growth, which includes cellular division and metabolic activity function, Figure 3 represents the content of photosynthetic pigments, which is an indicator of the algal cells’ ability to carry out photosynthesis. For Ni exposure, the high level of photosynthetic pigments in Figure 3 reflects an initial adaptive response where C. vulgaris increases its photosynthetic machinery to overcome nickel-stressed conditions. However, although there is this temporary increase in pigment synthesis, the overall growth of the population in Figure 1 is compromised since nickel still has an inhibitory effect on cell division and metabolic processes in the long term. This highlights the complexity of heavy metal stress in microalgae, with short-term physiological effects, such as increased pigment production, not converting into long-term population increase.
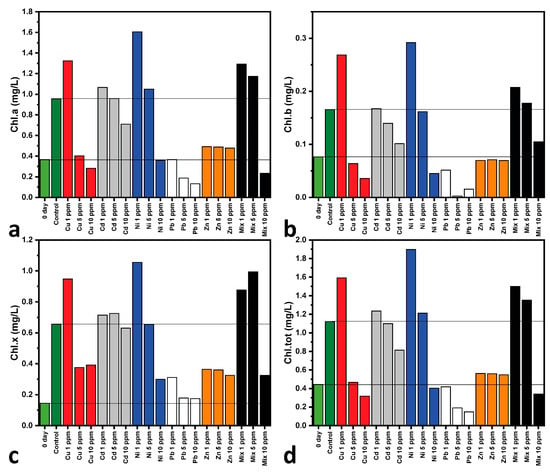
Figure 3.
Photosynthetic pigment content of mono- and multi-MT cultures on Days 0 (of all cultures) and 7 of cultivation; (a) Chlorophyll-a, (b) chlorophyll-b, (c) total chlorophyll, and (d) total carotenoid contents. Day 0 of cultivation of all cultures and the Control at Day 7 are highlighted for comparative reasons. Each value represents the mean of two separate measurements, with each one having an uncertainty of less than 5%.
3.3. Fourier Transform Infrared Analysis
ATR-FTIR analysis was performed on the dry microalga biomass that was collected on Day 7 of cultivation from all cultures to identify any spectral changes induced by heavy metal interactions. These changes would be associated with particular functional groups that are involved in metal bioremediation [26]. In Figure 4, the ATR-FTIR spectrum of the Control culture from the biomass collected on Day 7 of cultivation is presented. All characteristic bands are presented in Table 2, together with their attributions.
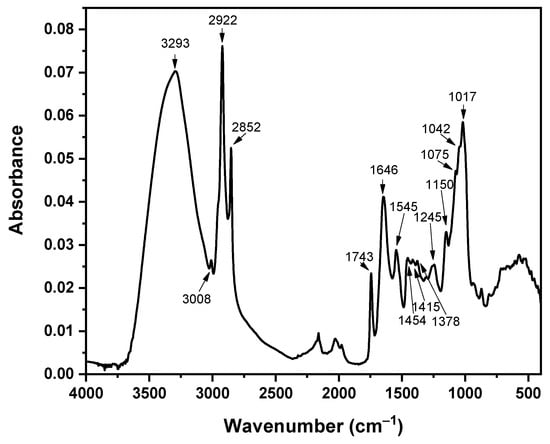
Figure 4.
ATR-FTIR spectrum of the C. vulgaris Control culture on Day 7 of cultivation. All characteristic bands are noted.
The band at 1743 cm−1 that is attributed to C=O groups is often absent, or of low intensity, in C. vulgaris cultures [24,83,84,85]. Its occurrence is associated with the presence of lipids in the cell wall of C. Vulgaris [25] and soluble polysaccharides that contain uronic acids (compounds derived from the oxidation of sugars CH2OH → COOH), sulfates and peptides [86], esters [87], and cellulose fatty acids [83]. As the thickness of the cell wall increases, the band at 1743 cm−1 also increases in intensity, which is always in relation to a simultaneous increase in the intensity of the band at ~3300 cm−1. In the latter band, apart from the -OH and -NH bands of proteins, the -OH band of the carboxyl groups of fatty acids also participates. The increase in the 1743 cm−1 band is also related to the increase in the bands at 3000–2800 cm−1 of aliphatic chains, as there are more aliphatic groups due to the elevated fatty acids [25]. The bands at 3000–2800 cm−1 are usually of low intensity in cultivations with low nutrient content [25,86,87]. Furthermore, the high intensity of the 1743 cm−1 band has also been related with cultivations of reduced nitrogen, where the cells produce more lipids and carbohydrates as an adaption for survival [25].
Table 2.
Characteristic FTIR bands of C. vulgaris and their attribution to functional groups.
Table 2.
Characteristic FTIR bands of C. vulgaris and their attribution to functional groups.
| Wavenumber (cm−1) | Attribution | References |
|---|---|---|
| 3293 | Hydroxyl and amino groups -OH, -NH, and -OH from carboxyl groups of lipid origin | [25] |
| 3008 | C-H stretching | [34] |
| 2922 and 2852 | C-H asymmetric and symmetric stretching | [25,34] |
| 1743 | C=O of esters | [25,83,86,87] |
| 1646 | C=O amide I | [25,88] |
| 1545 | C-N, N-H amide II | [25,88] |
| 1454 | CH3 asymmetric stretching or CH2 scissoring | [87,88] |
| 1378 | OH symmetric stretch of mono-, di-, and tri-glycerides, CH2 and CH3 proteins, the COO− of carboxylates, and the N(CH3)3 of lipids | [83,86,87,88] |
| 1245 | C-O stretch of COOH, PO, or OCO | [26,89] |
| 1150 | C-O-C of polysaccharides | |
| 1075 | C-O stretching and C-O-C, P=O, nucleic acid, or other phosphates and stretching of phosphodiesters | [83,86,87,88] |
| 1042 | C-O-C ether of polysaccharides | [25,26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54,55,56,57,58,59,60,61,62,63,64,65,66,67,68,69,70,71,72,73,74,75,76,77,78,79,80,81,82,83] |
| 1017 | C-H | [25] |
Figure 5 shows the collected ATR-FTIR spectra of C. vulgaris biomass on Day 7 of cultivation. In general, and in comparison to the Control spectrum, the mono- and multi-MT spectra present only slight differentiations and only regarding the intensity of bands, while no band shifting is observed. These differentiations are more obvious in the Mix 10 ppm spectrum, where a total decrease in the bands’ intensity is noted, while the relative intensity of the 1743 cm−1 band of lipids is the most decreased one, followed by the one at 1646 cm−1 of amide I. Other bands affected in terms of relative intensity decrease are those in the spectral area of 1100–950 cm−1, which are associated with the carbohydrate, phosphate, and CO groups; this is mostly observed in the spectra of Cu and Zn at 10 ppm and Pb and Mix at 5 and 10 ppm. Such differentiations in band intensities have also been observed in mono-MT cultures of Ni, Zn, Pb [26], and Cu [24] and in multi-MT cultures [14].
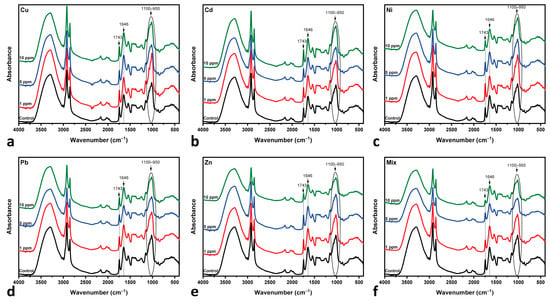
Figure 5.
ATR-FTIR spectra of the microalga. All measurements were performed on biomass collected on Day 7 of cultivation: (a) copper-MT; (b) cadmium-MT; (c) nickel-MT; (d) lead-MT; (e) zinc-MT; (f) multi-MT. The spectrum of the Control culture is included for comparison. The bands at 1743 (C=O of lipids), 1646 (C=O of amide I), and 1100–950 cm−1 are highlighted.
Since the spectra of all MT cultures show only slight variations, the I1646/I1743 ratio (amide I to lipid C=O band intensity) was calculated, as shown in Figure 6, to enhance spectral differences [26]. This ratio relies on the basis that lipids, i.e., the 1743 cm−1 band, remain stable in intensity, unaffected under heavy metal exposure [26]. Specifically, Rodrigues et al. [14] and Ferreira et al. [26] have observed that in mono-MT cultures, the 1743 cm−1 band remains intact, suggesting that the lipids are not significantly altered, while in multi-MT cultures, the authors have observed a decrease in the same band. By combining the results presented in Figure 5 and Figure 6, the following are observed:
- Cu-, Cd- and Pb-MT cultures: The amide I/lipid ratio increases at 5 ppm, followed by a slight decrease at the 10 ppm concentration. This behavior is indicative of lipid degradation or protein loss under metal-induced stress. Pb appears to have moderate changes in comparison to Cu and Cd.
- Zn-MT and Mix (multi-MT) cultures: The ratio decreases slightly up to the 5 ppm concentration. This is followed by an increase up to 10 ppm, while Mix increases sharper than that of the Zn-MT culture. Both cases present similar spectral behavior, as shown in Figure 6. The overall increase in the amide I/lipid ratio suggests a cumulative effect of both protein and lipid changes.
- Ni-MT culture: in this case, the steady upward trend of the amide I/lipid ratio over the metal concentration suggests that structural changes increase with the concentration of metals.

Figure 6.
The I1646/I1743 ratio, which represents the amide I-to-lipid ratio under the metal exposure of all cultures on Day 7 of cultivation.
Figure 6.
The I1646/I1743 ratio, which represents the amide I-to-lipid ratio under the metal exposure of all cultures on Day 7 of cultivation.

The calculation of the amide I/lipid ratio in the present study could be considered as inconclusive; although it has been previously supported that the C=O band at 1743 cm−1 [26] remains stable in mono-MT cultures, Figure 5 shows that both this and the amide I band at 1646 cm−1 present changes. In particular, the changes in the 1743 cm−1 and 1646 cm−1 bands show that both ester groups of lipids and -COO− of amide I are involved during the interaction of metals with the microalga, while the slight changes in the 1100–950 cm−1 bands suggest that the C-O, C-C, and P=O functional groups also participate [14,26].
3.4. Removal Efficiency of C. vulgaris
The results, as presented in Figure 7, highlight the complex interactions involved in using C. vulgaris to remove heavy metals, particularly the differences between single- and multi-MT cultures. In single-MT conditions, metals like Cu, Ni, and Zn tend to have higher removal rates because they can easily access functional groups on the algal cell wall. However, in multi-MT systems, competition between metals affects adsorption and biosorption, leading to variations in removal efficiency [14]. For Pb, lower removal rates in some cases are likely due to pH constraints. Pb adsorption is most effective at pH 6, but during cultivation, the pH often rises due to photosynthesis [50,85]. This effect is noticeable in single-MT systems at 1 ppm, where Pb removal increased from 71% on Day 3 to 92% by Day 7, indicating strong biosorption over time. In contrast, in multi-MT cultures at 1 ppm, removal was significantly lower (49% on Day 3, increasing to 62% on Day 7), likely due to competition with Cu, Zn, and Ni for binding sites, presenting an antagonistic behavior. At higher concentrations, Pb removal improves significantly (93% of 5 ppm culture and 96% for 10 ppm on Day 3), possibly due to the greater availability of binding sites at elevated metal levels [14].
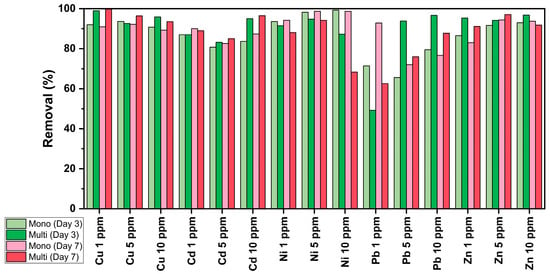
Figure 7.
Removal (%) efficiency of C. vulgaris during the cultivation of multi- and mono-MT cultures. Each presented value represents the mean of two separate measurements, with each one having an uncertainty of less than 5%.
The primary removal mechanism is biosorption, a passive process where metal ions bind to functional groups such as carboxyl, hydroxyl, amino, and phosphate through ion exchange, chelation, and complexation [2,90,91]. In ion exchange, metal ions like Pb or Cd replace naturally associated cations on the algal surface [17,92,93]. Chelation and complexation further stabilize metal attachment, particularly for Ni and Cu, which form coordination bonds with amino groups. Cu has the highest removal efficiency, staying above 89% across all concentrations and conditions, likely due to its strong affinity for adsorption sites and stable chelation. Even in multi-MT cultures, Cu removal remained consistently high: 99% at 1 ppm, 96% at 5 ppm, and 93% at 10 ppm on Day 7, demonstrating its competitive advantage in adsorption interactions [93].
Electrostatic interactions and van der Waals forces also contribute to metal adsorption, as metals bind to negatively charged sites on the algal surface. For example, in mono-MT cultures, Zn removal remains stable, indicating stronger biosorption [93].
Unlike biosorption, bioaccumulation is an active process where living C. vulgaris cells transport metals into the cytoplasm and organelles, storing them in vacuoles or binding them to intracellular proteins like metallothioneins [1,94,95]. This mechanism could be particularly effective for Cd, which presents high removal rates under all conditions. In multi-MT cultures at 10 ppm, Cd removal reaches 95% on Day 3 and 96% on Day 7, demonstrating strong intracellular retention and lower desorption compared to Zn and Ni.
Metal competition plays a major role in removal efficiency, with Cu and Pb often dominating due to their strong affinity for the amino and carboxyl groups. This leads to the displacement of Ni and Zn, reducing their removal efficiency [13,27]. For instance, Ni removal is high in mono-MT cultures (~98–99%) but drops significantly in multi-MT cultures, particularly at 10 ppm (68% on Day 7), likely because Cu forms stronger chelation bonds, thereby outcompeting Ni for binding sites. As C. vulgaris photosynthesizes, it consumes CO2, deprotonating functional groups, which enhances biosorption [4,17,96]. Interestingly, Cd remains stable even in multi-MT cultures at 10 ppm, likely due to its strong binding with the carboxyl and phosphate groups. Unlike Ni and Zn, Cd is less affected by competition from Cu or Pb, maintaining high removal efficiency even in complex multi-metal environments [2,16,17].
Overall, metals like Cu and Cd benefit from higher biosorption affinity, while Ni removal efficiency drops due to competition and solubility effects. These findings emphasize the importance of ion competition and multi-metal interactions in optimizing heavy metal removal using C. vulgaris. The removal efficiencies achieved in the present study, for both mono- and multi-MT cultures, are comparable to, or even exceed, the removal efficiencies of mono-MT systems, as reported in previous studies. The recorded removal efficiencies of Cu, Cd, Ni, Pb, and Zn are in the ranges of 39–95, 57–96, 32–99, 48–99, and 51–98%, respectively [14,17,27,97,98]. Nevertheless, it should be taken into consideration that these previous studies were conducted under varying conditions that may differ significantly in comparison with our work. Such variations include different cultivation conditions, varying metal concentrations and their combination (if multi-MT systems were applied), and the use of pretreated or dried microalga biomass; however, in one case, the removal of Cu from mine wastewater was examined.
3.5. Biosorption Analysis
Biosorption analysis using the Langmuir isotherm two-parameter model was used as an initial step to estimate the maximum adsorption ability (qm) and the metal affinity (expressed by the Langmuir constant KL) of C. vulgaris towards heavy metal bioremediation [63]. The model was applied to data collected on Day 3 of cultivation, which is considered an equilibrium state, as previous studies have shown that microalgae species reach equilibrium after less than 10 h of metal exposure [59]
Table 3 presents the calculated Langmuir isotherm model parameters of the mono-MT cultures, i.e., the maximum adsorption capacity (qm) and the Langmuir adsorption constant (KL) for each metal. For the mono-MT cultures, the cells present varying adsorption efficiencies of the metals under study. Pb was the one with the highest adsorption capacity. This shows that the cells were very efficient in adsorbing Pb ions. Cu was also efficient, with high adsorption capacity and moderate affinity, which shows the extent to which the metal ions are attracted to the adsorption sites on the cells. The high R2 values for Cu and Pb indicate the good fit of the Langmuir model; therefore, the adsorption process is well explained by the applied isotherm model. Cd presents lower adsorption capacity compared to Cu but higher affinity, which reflects the effective capture of Cd ions. Zn presents moderate adsorption capacity with high affinity, reflecting the effective capture of zinc ions by the microalga. Both Cd and Zn provided high R2 values, which reflects that their adsorption is also well described by the isotherm model. Ni, however, presents the highest affinity but the least adsorption capacity, which means that the cells had high attraction towards nickel ions but were not effective in capturing them. The low R2 value for nickel also indicates that there are other factors involved in its adsorption process, meaning that the Langmuir isotherm model is not adequate to explain the dynamics of nickel adsorption.
Table 3.
The derived Langmuir isotherm model parameters of C. vulgaris for mono-MT cultures.
The results of the multi-component non-modified Langmuir model, as applied to the multi-MT cultures (Table 4), indicate that Cu was absorbed mostly in the 5 ppm culture, as its qe,i is higher in comparison to the ones of the 1 ppm and 10 ppm cultures. Qe,Cd presents its peak absorption in the 5 ppm culture. The adsorption capacity of Ni has a consistent increase in absorption as the initial metal concentration increases. Pb uptake is the highest in the 1 ppm culture and decreases with increasing concentrations. Zn uptake is the highest in the 5 ppm culture. These findings suggest that the absorption profile of each metal varies with the concentration and possibly provide insights into the adsorption processes involved according to the Langmuir model.
Table 4.
Adsorption capacity of C. vulgaris at equilibrium (qe,i) for multi-MT cultures.
The difference in adsorption capacity between the mono- and the multi-MT systems suggests that the adsorption efficiency and properties of metals both vary significantly in terms of the availability and concentration of the rest of the metals that are present. In the mono-MT cultures, the highest adsorption capacity is observed in Cu and Pb, which suggests good adsorption, while Ni possesses the greatest affinity but the lowest adsorption capacity. The Langmuir isotherm model is well fitted for Cu, Cd, Pb, and Zn, which means that their adsorption is well-explained by the model. The model does not fit for Ni, which suggests that other mechanisms may be involved in its adsorption process [63]. Cu and Cd present good adsorption at moderate concentrations in the multi-MT systems, while Ni is more adsorbed at higher concentrations. These findings suggest that the adsorption curve of every metal is concentration-dependent, which explains the competitive adsorption processes involved, as implied by the AAS results in Section 3.4.
4. Conclusions
In the present study, the living microalga biomass of C. vulgaris was experimented towards its removal efficiency of mono- and multi-MT systems during its cultivation with BBM in laboratory-controlled conditions.
In both mono- and multi-MT cultures, significant removal was achieved, which exceeds 80%, apart from Pb and Ni, the latter in the case of the multi-MT culture, and only on Day 7 of cultivation. Lead presents lower removal efficiency in the case of the mono-MT culture and the multi-MT culture, with the latter occurring on Day 7 of cultivation. Regarding the other metals in the multi-MT cultures, the slight differentiations observed, as presented in Table 5, are attributed to synergistic or antagonistic interactions among them. Nevertheless, it should be emphasized that these differences are minimal. The effect of antagonistic and synergistic interactions is also supported by the cell population measurements, as well as the Langmuir isotherm model that was applied to both the mono- and multi-MT cultures. It should be noted that the model fitted well in the case of Cu, Cd, Pb, and Zn (qm between 1.55 and 7.93 × 10−8 mg/cell) for the mono-MT cultures, but Ni showed poor fitting, indicating that the Langmuir model is not the appropriate model to describe its adsorption. By comparing the qe,i values of the multi-MT cultures, Cu, Cd, and Zn present their highest adsorption capacity in the 5 ppm cultures, while Pb adsorption capacity decreases. This supports the presence of an antagonistic interaction of Pb to the other participating metals. The low qe,i values of all metals in the lowest multi-MT system concentration, the one at 1 ppm, suggest that the C. vulgaris cells are adapting to this low metal intoxication. The decrease of qe,i values of Cu, Cd, Pb, and Zn in the 10 ppm multi-MT cultures indicates that the metals compete between each other to cover the same binding sites. However, it should be noted that although the Langmuir isotherm model is widely accepted for describing biosorption capacity, future work should incorporate more experiments with a wider range of variables, leading to extended datasets that would allow for the implementation of other models, such as Freundlich and Temkin, for a more comprehensive approach of the adsorption processes, sites, and the interaction between metals in multi-MT systems.
Table 5.
Removal efficiency order of all metals for mono- and multi-MT cultures.
Chlorophyll-a, chlorophyll-b, total chlorophyl, and total carotenoid contents were calculated. It was displayed that all calculated pigment concentrations can be used to track the pigment content overall trend, as they follow the same pattern. The chlorophyll concentration indicated that certain metals affect the pigment content of the microalga, having a negative impact over photosynthesis and consequently over cell health. These findings agree with the collected optical microscopy images during cultivation, in terms of cell transparency and aggregate formation in affected and unhealthy cultures. FTIR spectroscopy, in general, showed slight spectral differentiations among the cultures. However, the analysis supports that certain cell functional groups participate in metal interaction.
The study supports that C. vulgaris can effectively be used for the removal of heavy metals from affected cultures, even in the case of multi-metal conditions, and for short treatment periods, i.e., 3 days. Since the lipid, protein, and carbohydrate contents of the microalga are not dramatically affected during bioremediation, the biomass of the intoxicated microalgal cells could be used for several applications, such as biofuel production, biogas, and catalysts, supporting a circular economy, while using CO2 and releasing oxygen through photosynthesis [20,47]. Future research should explore the possibility of such applications. Finally, while the present study demonstrates the efficiency of C. vulgaris in removing heavy metal mixtures during cultivation and under controlled laboratory conditions, scale-up experiments are required. Although our results are in accordance with previous studies over heavy metal bioremediation using microalgae and during their cultivation, most studies are focused on laboratory experiments. Pilot-scale and large-scale experiments and applications are scarce and require further research; key factors such as biomass production, wastewater treatment without additional nutrients, and microalgae viability in real conditions must be explored before industrial implementation. To investigate this, future research should be focused on scaling up the experiments by testing C. vulgaris and other microalgae in pilot-scale bioreactors and large-scale systems, such as open pond ones. This will determine the feasibility of this approach towards real-world perspectives, including economic and environmental aspects, such as cultivation yield and heavy metal removal capacity. Towards this, initial pilot-scale experiments using real wastewater and non-laboratory conditions are indeed in progress to further evaluate the large-scale feasibility of this approach.
Author Contributions
Conceptualization, N.A.K.; methodology, E.K., N.K., L.M., E.N. and N.A.K.; software, N.A.K.; validation, E.K., N.K., L.M. and N.A.K.; formal analysis, E.K., N.K. and L.M.; investigation, E.K., N.K., L.M., E.N. and N.A.K.; resources, N.A.K.; data curation, E.K., N.K. and L.M.; writing—original draft preparation, E.K., N.K. and L.M.; writing—review and editing, E.K., N.K., L.M. and N.A.K.; visualization, L.M.; supervision, N.A.K.; project administration, N.A.K.; funding acquisition, N.A.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research is financially supported under the Research Project “ALGEBRA—Innovative waste bioremediation practice for the removal of toxic compounds with the use of microalgae in the context of a circular economy”, which is funded by the program “NATURAL ENVIRONMENT & INNOVATIVE ACTIONS 2022”, PRIORITY AXIS 3: RESEARCH & IMPLEMENTATION, total budget: 199,501.23 €, Green Fund, Athena—Research and Innovation Center in Information Communication and Knowledge Technologies.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in this study are included in the article. Further inquiries can be directed to the corresponding author.
Acknowledgments
The authors would like to thank G. Vourlias for providing access to the FTIR facilities of the Laboratory of Advanced Materials and Devices (AMDE Lab), School of Physics, Aristotle University of Thessaloniki, Greece.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| AAS | Atomic absorption spectroscopy |
| ATR | Attenuated total reflectance |
| BBM | Bold’s Basal Medium |
| CAT | Catalase |
| C. vulgaris | Chlorella vulgaris |
| DNA | Deoxyribonucleic acid |
| FTIR | Fourier transform infrared |
| MT | Metal treated |
| ROS | Reactive oxygen species |
| SL | Stock Solution(s) |
| SOD | Superoxide dismutase |
References
- Kalita, N.; Baruah, P.P. Cyanobacteria as a Potent Platform for Heavy Metals Biosorption: Uptake, Responses and Removal Mechanisms. J. Hazard. Mater. Adv. 2023, 11, 100349. [Google Scholar] [CrossRef]
- Suresh Kumar, K.; Dahms, H.U.; Won, E.J.; Lee, J.S.; Shin, K.H. Microalgae—A Promising Tool for Heavy Metal Remediation. Ecotoxicol. Environ. Saf. 2015, 113, 329–352. [Google Scholar] [CrossRef] [PubMed]
- Saavedra, R.; Muñoz, R.; Taboada, M.E.; Vega, M.; Bolado, S. Comparative Uptake Study of Arsenic, Boron, Copper, Manganese and Zinc from Water by Different Green Microalgae. Bioresour. Technol. 2018, 263, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Arunakumara, K.K.I.U.; Zhang, X. Heavy Metal Bioaccumulation and Toxicity with Special Reference to Microalgae. J. Ocean Univ. China 2007, 7, 60–64. [Google Scholar] [CrossRef]
- Alloway, B.J. (Ed.) Heavy Metals in Soils. Trace Metals and Metalloids in Soils and Their Bioavailability, 3rd ed.; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012; Volume 22, ISBN 9780128243152. [Google Scholar]
- Velarde, L.; Nabavi, M.S.; Escalera, E.; Antti, M.L.; Akhtar, F. Adsorption of Heavy Metals on Natural Zeolites: A Review. Chemosphere 2023, 328, 138508. [Google Scholar] [CrossRef]
- Abadin, H.; Ashizawa, A.; Stevens, Y.-W.; Llados, F.; Diamond, G.; Sage, G.; Citra, M.; Quinones, A.; Bosch, S.J.; Swarts, S.G. Toxicological Profile for Lead; Agency for Toxic Substances and Disease Registry (US): Atlanta, GA, USA, 2020; pp. 1–583. [Google Scholar]
- Dettori, M.; Arghittu, A.; Deiana, G.; Castiglia, P.; Azara, A. The Revised European Directive 2020/2184 on the Quality of Water Intended for Human Consumption. A Step Forward in Risk Assessment, Consumer Safety and Informative Communication. Environ. Res. 2022, 209, 112773. [Google Scholar] [CrossRef]
- Xu, K.; Du, M.; Yao, R.; Luo, J.; Chen, Z.; Li, C.; Lei, A.; Wang, J. Microalgae-Mediated Heavy Metal Removal in Wastewater Treatment: Mechanisms, Influencing Factors, and Novel Techniques. Algal Res. 2024, 82, 103645. [Google Scholar] [CrossRef]
- European Parliament and Council. European Pollutant Release and Transfer Register. Off. J. Eur. Union 2006, L33, 1–17. [Google Scholar]
- Li, H.; Watson, J.; Zhang, Y.; Lu, H.; Liu, Z. Environment-Enhancing Process for Algal Wastewater Treatment, Heavy Metal Control and Hydrothermal Biofuel Production: A Critical Review. Bioresour. Technol. 2020, 298, 122421. [Google Scholar] [CrossRef]
- Arora, R. Adsorption of Heavy Metals—A Review. Mater. Today Proc. 2019, 18, 4745–4750. [Google Scholar] [CrossRef]
- Anastopoulos, I.; Kyzas, G.Z. Progress in Batch Biosorption of Heavy Metals onto Algae. J. Mol. Liq. 2015, 209, 77–86. [Google Scholar] [CrossRef]
- Rodrigues, M.S.; Ferreira, L.S.; de Carvalho, J.C.M.; Lodi, A.; Finocchio, E.; Converti, A. Metal Biosorption onto Dry Biomass of Arthrospira (Spirulina) platensis and Chlorella Vulgaris: Multi-Metal Systems. J. Hazard. Mater. 2012, 217–218, 246–255. [Google Scholar] [CrossRef] [PubMed]
- Jayakumar, V.; Govindaradjane, S.; Rajamohan, N.; Rajasimman, M. Biosorption Potential of Brown Algae, Sargassum polycystum, for the Removal of Toxic Metals, Cadmium and Zinc. Environ. Sci. Pollut. Res. 2022, 29, 41909–41922. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Li, W.; Jin, M.; Zhang, L.; Qin, L.; Geng, W. Responses and Tolerance Mechanisms of Microalgae to Heavy Metal Stress: A Review. Mar. Environ. Res. 2023, 183, 105805. [Google Scholar] [CrossRef]
- Mehta, S.K.; Gaur, J.P. Use of Algae for Removing Heavy Metal Ions from Wastewater: Progress and Prospects. Crit. Rev. Biotechnol. 2005, 25, 113–152. [Google Scholar] [CrossRef]
- Rangabhashiyam, S.; Balasubramanian, P. Characteristics, Performances, Equilibrium and Kinetic Modeling Aspects of Heavy Metal Removal Using Algae. Bioresour. Technol. Rep. 2019, 5, 261–279. [Google Scholar] [CrossRef]
- Khan, Z.; Anees, A.; Khan, I.; Shahwar, D. Advancements in Microalgal Bioremediation of Heavy Metal-Contaminated Water. In Bio-Organic Amendments for Heavy Metal Remediation: Water, Soil and Plant Approaches and Technologies; Elsevier: Amsterdam, The Netherlands, 2024; pp. 33–55. ISBN 9780443216107. [Google Scholar]
- El-Sheekh, M.M.; El-Kassas, H.Y.; Ali, S.S. Microalgae-Based Bioremediation of Refractory Pollutants: An Approach towards Environmental Sustainability. Microb. Cell Factories 2025, 24, 19. [Google Scholar] [CrossRef]
- Edris, G.; Alhamed, Y.; Alzahrani, A. Biosorption of Cadmium and Lead from Aqueous Solutions by Chlorella vulgaris Biomass: Equilibrium and Kinetic Study. Arab. J. Sci. Eng. 2014, 39, 87–93. [Google Scholar] [CrossRef]
- Rachlin, J.W.; Grosso, A. The Effects of PH on the Growth of Chlorella vulgaris and Its Interactions with Cadmium Toxicity. Arch. Environ. Contam. Toxicol. 1991, 20, 505–508. [Google Scholar] [CrossRef]
- Jones, L.A.; Ogden, K.L.; Jia, F. Comparative Study of Biosorption of Copper(II) by Lipid Extracted and Non-Extracted Chlorella sorokiniana. CLEAN Soil Air Water 2015, 43, 73–78. [Google Scholar] [CrossRef]
- Zeng, Y.; Chen, X.; Zhu, J.; Long, D.; Jian, Y.; Tan, Q.; Wang, H. Effects of Cu (II) on the Growth of Chlorella vulgaris and Its Removal Efficiency of Pollutants in Synthetic Piggery Digestate. Toxics 2024, 12, 56. [Google Scholar] [CrossRef]
- Soto-Ramírez, R.; Lobos, M.G.; Córdova, O.; Poirrier, P.; Chamy, R. Effect of Growth Conditions on Cell Wall Composition and Cadmium Adsorption in Chlorella vulgaris: A New Approach to Biosorption Research. J. Hazard. Mater. 2021, 411, 125059. [Google Scholar] [CrossRef]
- Ferreira, L.S.; Rodrigues, M.S.; de Carvalho, J.C.M.; Lodi, A.; Finocchio, E.; Perego, P.; Converti, A. Adsorption of Ni2+, Zn2+ and Pb2+ onto Dry Biomass of Arthrospira (Spirulina) platensis and Chlorella vulgaris. I. Single Metal Systems. Chem. Eng. J. 2011, 173, 326–333. [Google Scholar] [CrossRef]
- Manzoor, F.; Karbassi, A.; Golzary, A. Removal of Heavy Metal Contaminants from Wastewater by Using Chlorella vulgaris Beijerinck: A Review. Curr. Environ. Manag. 2019, 6, 174–187. [Google Scholar] [CrossRef]
- El-Sayed, A.E.K.B.; Reda, M.M.; Almutairi, A.W.; Mavromatis, C. Biomass Production and Biochemical Composition of Chlorella vulgaris Grown in Net-House Photobioreactor (NHPBR) Using Sugarcane Press Mud Waste. J. Taibah Univ. Sci. 2023, 17, 2194843. [Google Scholar] [CrossRef]
- Spain, O.; Funk, C. Detailed Characterization of the Cell Wall Structure and Composition of Nordic Green Microalgae. J. Agric. Food Chem. 2022, 70, 9711–9721. [Google Scholar] [CrossRef]
- Spolaore, P.; Joannis-Cassan, C.; Duran, E.; Isambert, A. Commercial Applications of Microalgae. J. Biosci. Bioeng. 2006, 101, 87–96. [Google Scholar] [CrossRef]
- Lam, M.K.; Lee, K.T. Potential of Using Organic Fertilizer to Cultivate Chlorella vulgaris for Biodiesel Production. Appl. Energy 2012, 94, 303–308. [Google Scholar] [CrossRef]
- Ali, S.S.; Hassan, L.H.S. Microalgae-Mediated Bioremediation: Current Trends and Opportunities-a Review. Arch. Microbiol. 2024, 206, 343. [Google Scholar] [CrossRef]
- Chia, M.A.; Lombardi, A.T.; da Melão, M.G.G. Growth and Biochemical Composition of Chlorella vulgaris in Different Growth Media. An. Acad. Bras. Cienc. 2013, 85, 1427–1438. [Google Scholar] [CrossRef]
- Yang, J.S.; Cao, J.; Xing, G.L.; Yuan, H.L. Lipid Production Combined with Biosorption and Bioaccumulation of Cadmium, Copper, Manganese and Zinc by Oleaginous Microalgae Chlorella minutissima UTEX2341. Bioresour. Technol. 2015, 175, 537–544. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Qiu, H.; Chang, Z.; Jiang, Z.; Yin, W. The Effect of Cadmium on the Growth and Antioxidant Response for Freshwater Algae Chlorella vulgaris. SpringerPlus 2016, 5, 1290. [Google Scholar] [CrossRef] [PubMed]
- Qasem, N.A.A.; Mohammed, R.H.; Lawal, D.U. Removal of Heavy Metal Ions from Wastewater: A Comprehensive and Critical Review. npj Clean Water 2021, 4, 1–15. [Google Scholar] [CrossRef]
- Expósito, N.; Carafa, R.; Kumar, V.; Sierra, J.; Schuhmacher, M.; Papiol, G.G. Performance of Chlorella vulgaris Exposed to Heavy Metal Mixtures: Linking Measured Endpoints and Mechanisms. Int. J. Environ. Res. Public Health 2021, 18, 1037. [Google Scholar] [CrossRef]
- Liu, Z.W.; Zhang, L.S.; Feng, X.Y.; He, Z.X.; Sun, Y.; Tao, X.Y.; Yin, Q.; Yang, L.M.; Zhou, R.J.; He, X.Q.; et al. Near-Infrared Spectroscopy Bioprobe Estimation of Metabolites’ Responses to Pb2+ in Cladophora rupestris. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2024, 306, 123544. [Google Scholar] [CrossRef]
- Rinaldi, K.L.; Senhorinho, G.N.A.; Laamanen, C.A.; Scott, J.A. A Review of Extremophilic Microalgae: Impacts of Experimental Cultivation Conditions for the Production of Antimicrobials. Algal Res. 2024, 78, 103427. [Google Scholar] [CrossRef]
- Bajguz, A. Suppression of Chlorella vulgaris Growth by Cadmium, Lead, and Copper Stress and Its Restoration by Endogenous Brassinolide. Arch. Environ. Contam. Toxicol. 2011, 60, 406–416. [Google Scholar] [CrossRef]
- Hasanuzzaman, M. Approaches to the Remediation of Inorganic Pollutants; Springer: Hyderabad, India, 2021; ISBN 9789811562211. [Google Scholar]
- Tripathi, S.; Poluri, K.M. Heavy Metal Detoxification Mechanisms by Microalgae: Insights from Transcriptomics Analysis. Environ. Pollut. 2021, 285, 117443. [Google Scholar] [CrossRef]
- Liu, S.; Jiang, M.; Wu, J.; Li, X.; Zhu, J. Mechanism of Biological Transport and Transformation of Copper, Cadmium, and Zinc in Water by Chlorella. Water 2024, 16, 1906. [Google Scholar] [CrossRef]
- Santos, F.M.; Mazur, L.P.; Mayer, D.A.; Vilar, V.J.P.; Pires, J.C.M. Inhibition Effect of Zinc, Cadmium, and Nickel Ions in Microalgal Growth and Nutrient Uptake from Water: An Experimental Approach. Chem. Eng. J. 2019, 366, 358–367. [Google Scholar] [CrossRef]
- Geng, W.; Xiao, X.; Zhang, L.; Ni, W.; Li, N.; Li, Y. Response and Tolerance Ability of Chlorella vulgaris to Cadmium Pollution Stress. Environ. Technol. 2022, 43, 4391–4401. [Google Scholar] [CrossRef] [PubMed]
- Lalhmunsiama; Gupta, P.L.; Jung, H.; Tiwari, D.; Kong, S.H.; Lee, S.M. Insight into the Mechanism of Cd(II) and Pb(II) Removal by Sustainable Magnetic Biosorbent Precursor to Chlorella vulgaris. J. Taiwan Inst. Chem. Eng. 2017, 71, 206–213. [Google Scholar] [CrossRef]
- Kusuma, H.S.; Illiyanasafa, N.; Jaya, D.E.C.; Darmokoesoemo, H.; Putra, N.R. Utilization of the Microalga Chlorella vulgaris for Mercury Bioremediation from Wastewater and Biomass Production. Sustain. Chem. Pharm. 2024, 37, 101346. [Google Scholar] [CrossRef]
- Dmytryk, A.; Saeid, A.; Chojnacka, K. Biosorption of Microelements by Spirulina: Towards Technology of Mineral Feed Supplements. Sci. World J. 2014, 2014, 356328. [Google Scholar] [CrossRef]
- Zinicovscaia, I.; Safonov, A.; Tregubova, V.; Ilin, V.; Cepoi, L.; Chiriac, T.; Rudi, L.; Frontasyeva, M.V. Uptake of Metals from Single and Multi-Component Systems by Spirulina platensis Biomass. Ecol. Chem. Eng. S 2016, 23, 401–412. [Google Scholar] [CrossRef][Green Version]
- Malletzidou, L.; Kyratzopoulou, E.; Kyzaki, N.; Nerantzis, E.; Kazakis, N.A. Towards the Sustainable Removal of Heavy Metals from Wastewater Using Arthrospira platensis: A Laboratory-Scale Approach in the Context of a Green Circular Economy. Appl. Sci. 2025, 15, 791. [Google Scholar] [CrossRef]
- Birungi, Z.S.; Chirwa, E.M.N.; Botai, O.J. Competitive Adsorption in a Ternary System of Toxic Metals and Rare Earth Elements Using Desmodesmus multivariabilis: Empirical and Kinetic Modelling. J. Appl. Phycol. 2017, 29, 2899–2910. [Google Scholar] [CrossRef]
- Kazakis, N.A. Green Approaches to Heavy Metal Removal from Wastewater: Microalgae Solutions in a Circular Economy Framework. Soc. Impacts 2025, 5, 100103. [Google Scholar] [CrossRef]
- Bischoff, H.W. Phycological Studies IV. Some Soil Algae from Enchanted Rock and Related Algal Species; University of Texas Publication: Austin, TX, USA, 1963; No. 6318; pp. 1–95. [Google Scholar]
- BBM Medium. CCCryo Culture Collection of Cryophilic Algae, 06/2020. 2020. Available online: http://cccryo.fraunhofer.de/sources/files/medien/BBM.pdf (accessed on 28 January 2024).
- Challagulla, V.; Walsh, K.B.; Subedi, P. Biomass and Total Lipid Content Assessment of Microalgal Cultures Using Near and Short Wave Infrared Spectroscopy. Bioenergy Res. 2014, 7, 306–318. [Google Scholar] [CrossRef]
- Gille, A.; Trautmann, A.; Posten, C.; Briviba, K. Bioaccessibility of Carotenoids from Chlorella vulgaris and Chlamydomonas reinhardtii. Int. J. Food Sci. Nutr. 2016, 67, 507–513. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and Carotenoids: Pigments of Photosynthetic Biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar] [CrossRef]
- Parsons, T.R.; Strickland, J.D. Discussion of Spectrophotometric Determination of Marine-Plant Pigments, with Revised Equations for Ascertaining Chlorophylls and Carotenoids. J. Mar. Res. 1963, 21, 155–163. [Google Scholar] [CrossRef]
- Plöhn, M.; Escudero-Oñate, C.; Funk, C. Biosorption of Cd(II) by Nordic Microalgae: Tolerance, Kinetics and Equilibrium Studies. Algal Res. 2021, 59, 102471. [Google Scholar] [CrossRef]
- Rajfur, M.; Kłos, A. Sorption of Heavy Metals in the Biomass of Alga Palmaria palmata. Water Sci. Technol. 2013, 68, 1543–1549. [Google Scholar] [CrossRef]
- Omolabake Abiodun, O.-A.; Oluwaseun, O.; Kayode Oladayo, O.; Abayomi, O.; Arubi George, A.; Opatola, E.; Friday Orah, R.; Jeffery Isukuru, E.; Chiamaka Ede, I.; Temitayo Oluwayomi, O.; et al. Remediation of Heavy Metals Using Biomass-Based Adsorbents: Adsorption Kinetics and Isotherm Models. Clean Technol. 2023, 5, 934–960. [Google Scholar] [CrossRef]
- Abdulaziz, M.; Musayev, S. Multicomponent Biosorption of Heavy Metals from Aqueous Solutions: A Review. Pol. J. Environ. Stud. 2017, 26, 1433–1441. [Google Scholar] [CrossRef]
- Cheng, J.; Yin, W.; Chang, Z.; Lundholm, N.; Jiang, Z. Biosorption Capacity and Kinetics of Cadmium(II) on Live and Dead Chlorella vulgaris. J. Appl. Phycol. 2017, 29, 211–221. [Google Scholar] [CrossRef]
- Langmuir, I. The Adsorption of Gases on Plane Surfaces of Glass, Mica and Platinum. J. Am. Chem. Soc. 1918, 40, 1361–1403. [Google Scholar] [CrossRef]
- Rangsayatorn, N.; Upatham, E.S.; Kruatrachue, M.; Pokethitiyook, P.; Lanza, G.R. Phytoremediation Potential of Spirulina (Arthrospira) platensis: Biosorption and Toxicity Studies of Cadmium. Environ. Pollut. 2002, 119, 45–53. [Google Scholar] [CrossRef]
- Jiang, L.; Zhou, W.; Liu, D.; Liu, T.; Wang, Z. Biosorption Isotherm Study of Cd2 +, Pb2 + and Zn2 + Biosorption onto Marine Bacterium Pseudoalteromonas sp. SCSE709-6 in Multiple Systems. J. Mol. Liq. 2017, 247, 230–237. [Google Scholar] [CrossRef]
- Zhou, G.J.; Peng, F.Q.; Zhang, L.J.; Ying, G.G. Biosorption of Zinc and Copper from Aqueous Solutions by Two Freshwater Green Microalgae Chlorella pyrenoidosa and Scenedesmus obliquus. Environ. Sci. Pollut. Res. 2012, 19, 2918–2929. [Google Scholar] [CrossRef] [PubMed]
- Gupta, V.K.; Rastogi, A. Biosorption of Lead(II) from Aqueous Solutions by Non-Living Algal Biomass Oedogonium sp. and Nostoc sp.-A Comparative Study. Colloids Surf. B Biointerfaces 2008, 64, 170–178. [Google Scholar] [CrossRef] [PubMed]
- Nowicka, B. Heavy Metal–Induced Stress in Eukaryotic Algae—Mechanisms of Heavy Metal Toxicity and Tolerance with Particular Emphasis on Oxidative Stress in Exposed Cells and the Role of Antioxidant Response. Environ. Sci. Pollut. Res. 2022, 29, 16860–16911. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, E.J.; Baldwin, L.A. Hormesis: U-Shaped Dose Responses and Their Centrality in Toxicology. Trends Pharmacol. Sci. 2001, 22, 285–291. [Google Scholar] [CrossRef]
- Iqhrammullah, M.; Saudah, S.; Monalisa, M.; Fahrurrozi, F.; Akbar, S.A.; Lubis, S.S. Network Meta-Analysis of Cadmium Toxicity against Chlorella vulgaris and the Role of Growth Stimulants and Macronutrients. Glob. J. Environ. Sci. Manag. 2024, 10, 1561–1572. [Google Scholar] [CrossRef]
- Calabrese, E.J.; Baldwin, L.A. Applications of Hormesis in Toxicology, Risk Assessment and Chemotherapeutics. Trends Pharmacol. Sci. 2002, 23, 331–337. [Google Scholar] [CrossRef]
- Nagajyoti, P.C.; Lee, K.D.; Sreekanth, T.V.M. Heavy Metals, Occurrence and Toxicity for Plants: A Review. Environ. Chem. Lett. 2010, 8, 199–216. [Google Scholar] [CrossRef]
- Gikas, P. Single and Combined Effects of Nickel (Ni(II)) and Cobalt (Co(II)) Ions on Activated Sludge and on Other Aerobic Microorganisms: A Review. J. Hazard. Mater. 2008, 159, 187–203. [Google Scholar] [CrossRef]
- Monteiro, C.M.; Castro, P.M.L.; Malcata, F.X. Metal Uptake by Microalgae: Underlying Mechanisms and Practical Applications. Biotechnol. Prog. 2012, 28, 299–311. [Google Scholar] [CrossRef]
- Zeraatkar, A.K.; Ahmadzadeh, H.; Talebi, A.F.; Moheimani, N.R.; McHenry, M.P. Potential Use of Algae for Heavy Metal Bioremediation, a Critical Review. J. Environ. Manag. 2016, 181, 817–831. [Google Scholar] [CrossRef]
- Kondzior, P.; Butarewicz, A. Effect of Heavy Metals (Cu and Zn) on the Content of Photosynthetic Pigments in the Cells of Algae Chlorella vulgaris. J. Ecol. Eng. 2018, 19, 18–28. [Google Scholar] [CrossRef] [PubMed]
- Rocha, G.S.; Parrish, C.C.; Espíndola, E.L.G. Effects of Copper on Photosynthetic and Physiological Parameters of a Freshwater Microalga (Chlorophyceae). Algal Res. 2021, 54, 102223. [Google Scholar] [CrossRef]
- Sytar, O.; Kumar, A.; Latowski, D.; Kuczynska, P.; Strzałka, K.; Prasad, M.N.V. Heavy Metal-Induced Oxidative Damage, Defense Reactions, and Detoxification Mechanisms in Plants. Acta Physiol. Plant. 2013, 35, 985–999. [Google Scholar] [CrossRef]
- Wang, M.; Zou, J.; Duan, X.; Jiang, W.; Liu, D. Cadmium Accumulation and Its Effects on Metal Uptake in Maize (Zea Mays L.). Bioresour. Technol. 2007, 98, 82–88. [Google Scholar] [CrossRef]
- Ekmekçi, Y.; Tanyolaç, D.; Ayhan, B. Effects of Cadmium on Antioxidant Enzyme and Photosynthetic Activities in Leaves of Two Maize Cultivars. J. Plant Physiol. 2008, 165, 600–611. [Google Scholar] [CrossRef]
- Qian, H.; Li, J.; Sun, L.; Chen, W.; Sheng, G.D.; Liu, W.; Fu, Z. Combined Effect of Copper and Cadmium on Chlorella vulgaris Growth and Photosynthesis-Related Gene Transcription. Aquat. Toxicol. 2009, 94, 56–61. [Google Scholar] [CrossRef]
- Duygu, D.; Udoh, A.; Ozer, T.; Akbulut, A.; Erkaya, I.; Yildiz, K.; Guler, D. Fourier Transform Infrared (FTIR) Spectroscopy for Identification of Chlorella vulgaris Beijerinck 1890 and Scenedesmus obliquus (Turpin) Kützing 1833. Afr. J. Biotechnol. 2012, 11, 3817–3824. [Google Scholar] [CrossRef]
- Sukhikh, S.; Prosekov, A.; Ivanova, S.; Maslennikov, P.; Andreeva, A.; Budenkova, E.; Kashirskikh, E.; Tcibulnikova, A.; Zemliakova, E.; Samusev, I.; et al. Identification of Metabolites with Antibacterial Activities by Analyzing the FTIR Spectra of Microalgae. Life 2022, 12, 1395. [Google Scholar] [CrossRef]
- Dao, L.; Beardall, J.; Heraud, P. Characterisation of Pb-Induced Changes and Prediction of Pb Exposure in Microalgae Using Infrared Spectroscopy. Aquat. Toxicol. 2017, 188, 33–42. [Google Scholar] [CrossRef]
- El-Naggar, N.E.A.; Hussein, M.H.; Shaaban-Dessuuki, S.A.; Dalal, S.R. Production, Extraction and Characterization of Chlorella vulgaris Soluble Polysaccharides and Their Applications in AgNPs Biosynthesis and Biostimulation of Plant Growth. Sci. Rep. 2020, 10, 3011. [Google Scholar] [CrossRef]
- Baldev, E.; MubarakAli, D.; Sivasubramanian, V.; Pugazhendhi, A.; Thajuddin, N. Unveiling the Induced Lipid Production in Chlorella vulgaris under Pulsed Magnetic Field Treatment. Chemosphere 2021, 279, 130673. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Saxena, A.; Song, B.; Ward, B.B.; Beveridge, T.J.; Myneni, S.C.B. Elucidation of Functional Groups on Gram-Positive and Gram-Negative Bacterial Surfaces Using Infrared Spectroscopy. Langmuir 2004, 20, 11433–11442. [Google Scholar] [CrossRef] [PubMed]
- Yalçın, S.; Sezer, S.; Apak, R. Characterization and Lead(II), Cadmium(II), Nickel(II) Biosorption of Dried Marine Brown Macro Algae Cystoseira barbata. Environ. Sci. Pollut. Res. 2012, 19, 3118–3125. [Google Scholar] [CrossRef] [PubMed]
- Salama, E.S.; Roh, H.S.; Dev, S.; Khan, M.A.; Abou-Shanab, R.A.I.; Chang, S.W.; Jeon, B.H. Algae as a Green Technology for Heavy Metals Removal from Various Wastewater. World J. Microbiol. Biotechnol. 2019, 35, 75. [Google Scholar] [CrossRef]
- Fitri, W.E.; Putra, A.; Febria, F.A. Removal of Heavy Metals Using Chlorella vulgaris: A Review. J. Katalisator 2024, 9, 148–162. [Google Scholar] [CrossRef]
- Mitrogiannis, D.; Markou, G.; Çelekli, A.; Bozkurt, H. Biosorption of Methylene Blue onto Arthrospira platensis Biomass: Kinetic, Equilibrium and Thermodynamic Studies. J. Environ. Chem. Eng. 2015, 3, 670–680. [Google Scholar] [CrossRef]
- Hussain, M.K.; Khatoon, S.; Nizami, G.; Fatma, U.K.; Ali, M.; Singh, B.; Quraishi, A.; Assiri, M.A.; Ahamad, S.; Saquib, M. Unleashing the Power of Bio-Adsorbents: Efficient Heavy Metal Removal for Sustainable Water Purification. J. Water Process Eng. 2024, 64, 105705. [Google Scholar] [CrossRef]
- Kiran Marella, T.; Saxena, A.; Tiwari, A. Diatom Mediated Heavy Metal Remediation: A Review. Bioresour. Technol. 2020, 305, 123068. [Google Scholar] [CrossRef]
- Safi, C.; Zebib, B.; Merah, O.; Pontalier, P.Y.; Vaca-Garcia, C. Morphology, Composition, Production, Processing and Applications of Chlorella vulgaris: A Review. Renew. Sustain. Energy Rev. 2014, 35, 265–278. [Google Scholar] [CrossRef]
- Soeprobowati, T.R. Phycoremediation of Pb+2, Cd+2, Cu+2, and Cr+3 by Spirulina platensis (Gomont) Geitler. Am. J. Biosci. 2014, 2, 165–170. [Google Scholar] [CrossRef]
- Goswami, R.K.; Agrawal, K.; Shah, M.P.; Verma, P. Bioremediation of Heavy Metals from Wastewater: A Current Perspective on Microalgae-based Future. Lett. Appl. Microbiol. 2022, 75, 701–717. [Google Scholar] [CrossRef]
- Aung, W.L.; Hlaing, N.; Aye, K.N. Biosorption of Lead (Pb2+) by Using Chlorella vulgaris. Int. J. Chem. Environ. Biol. Sci. 2013, 1, 2320–4087. [Google Scholar]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).