
Soil Texture’s Hidden Influence: Decoding Plant Diversity Patterns in Arid Ecosystems


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
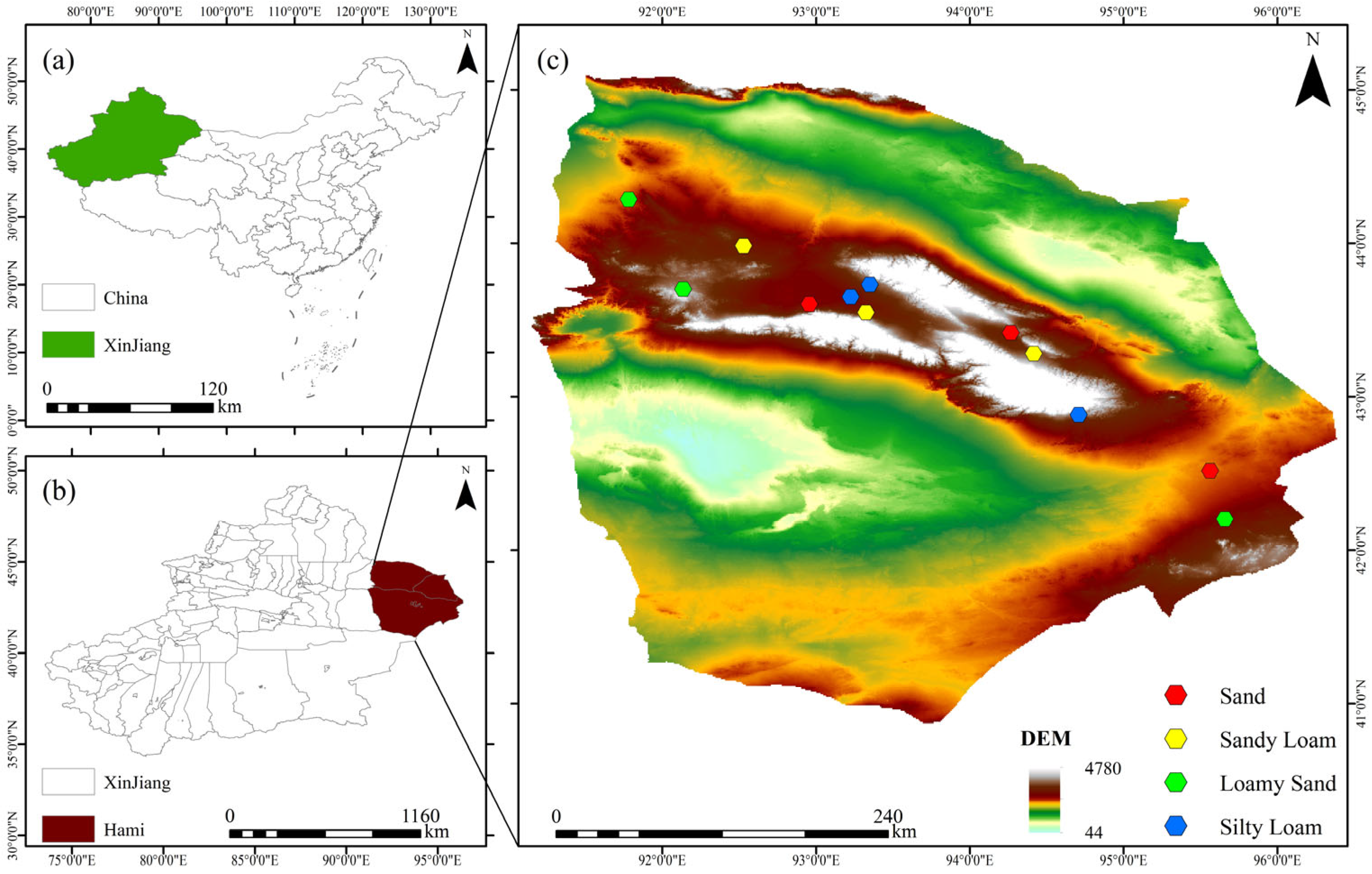
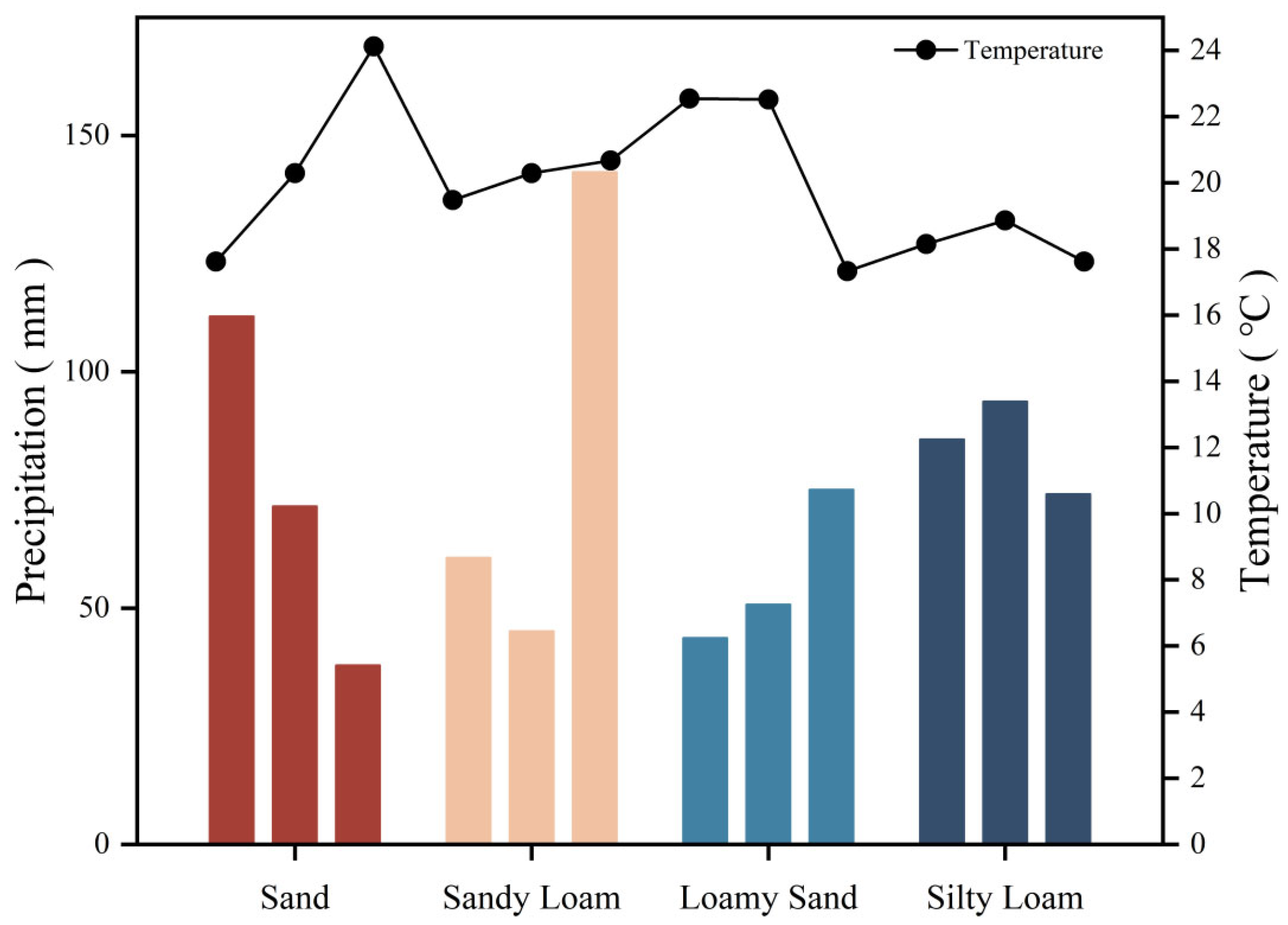
2.1. Study Site
2.2. Vegetation Survey
2.3. Soil Sampling and Experiment
2.4. Soil Texture Classification
2.5. Subsection
2.6. Data Processing
3. Results
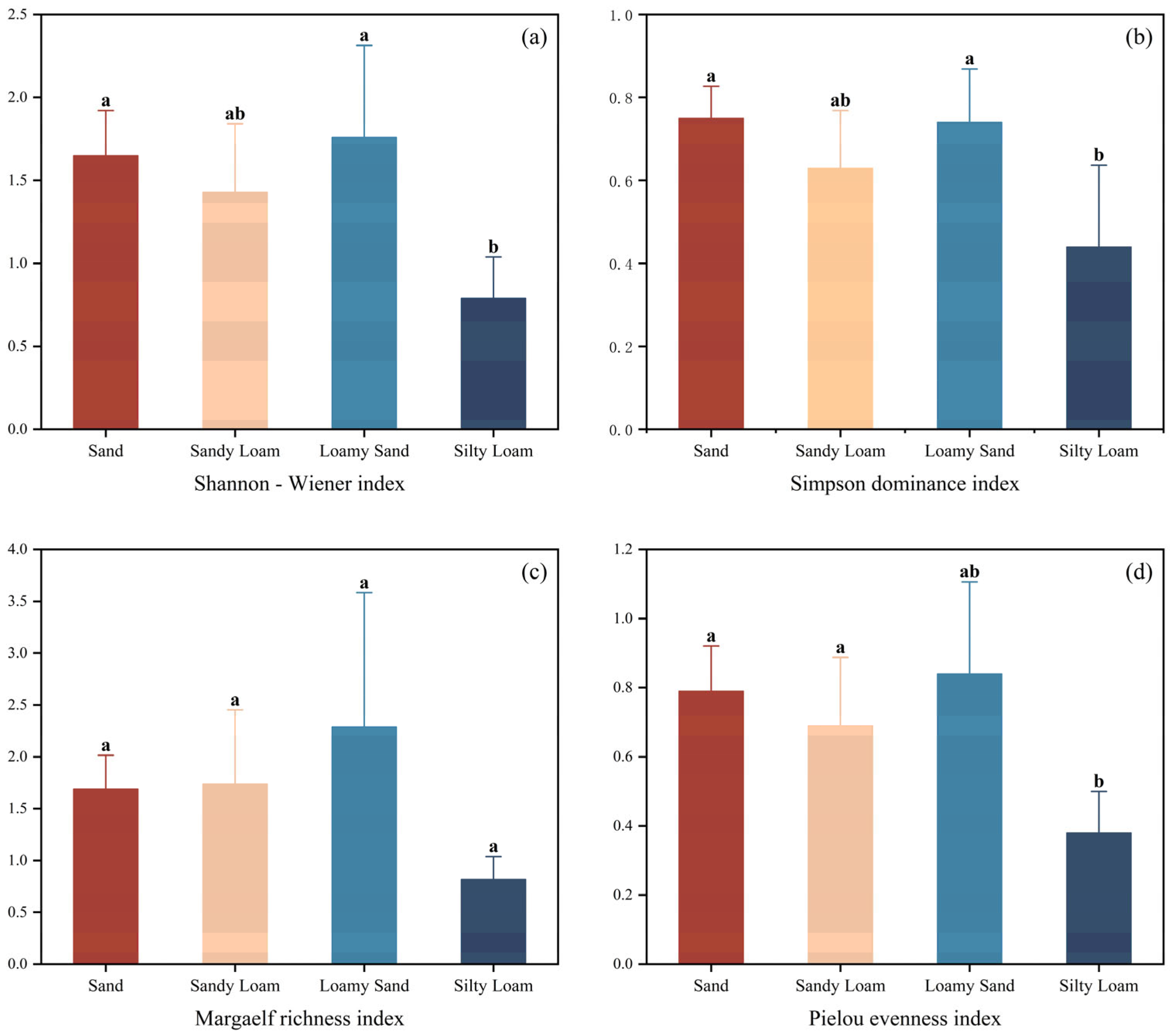
3.1. Plant Species Diversity
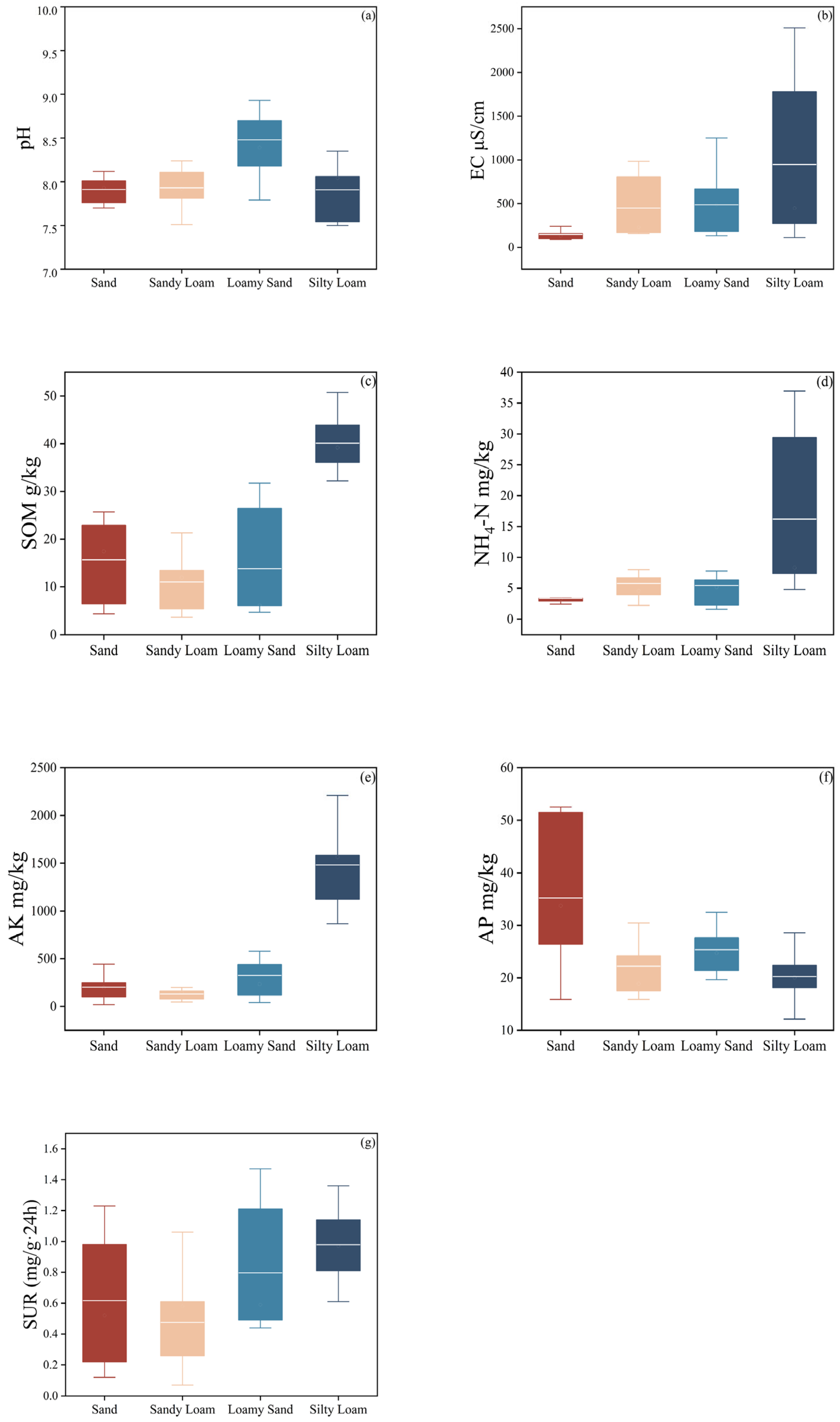
3.2. Soil Physicochemical Properties and Urease Activity
3.3. Factors Influencing Plant Diversity
4. Discussion
4.1. Effects of Soil Texture on Plant Species Diversity
4.2. Relationship Between Soil Texture and Physicochemical Properties and Urease Activity
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cao, Y.; Nie, M.; Shen, Y.; Hu, Y.; Ma, D.; Li, D.; Hou, T.; Fang, P.; Wang, X. Changes in vegetation and soil characteristics and their correlations in grasslands at different stages of degradation on the desert steppe in an arid wind-sandy area of Ningxia. Acta Pratacult. Sin. 2024, 33, 1–14. [Google Scholar]
- Zhang, F.; Zhang, L.; Wang, W.; Wang, X. Relationships Between Characteristic of Soil Nutrients and Soil Texture and Moisture in Wind-Water Erosion Crisscross Region. J. Soil Water Conserv. 2012, 26, 99–104. [Google Scholar]
- Miao, J.; Li, S.; Xu, X.; Liu, G.; Wang, H.; Fan, J.; Akhmadi, K. Benefits and ecological restoration implications of hanging grass fences in Mongolian desert steppe. J. Arid Land 2024, 16, 1541–1561. [Google Scholar] [CrossRef]
- Yan, Y.; Xin, X.; Xu, X.; Wang, X.; Yang, G.; Yan, R.; Chen, B. Quantitative effects of wind erosion on the soil texture and soil nutrients under different vegetation coverage in a semiarid steppe of northern China. Plant Soil 2013, 369, 585–598. [Google Scholar] [CrossRef]
- Broecher, M.; Meyer, S.T.; Eisenhauer, N.; Ebeling, A. The positive plant diversity/consumer relationship is independent of grassland age. BASIC Appl. Ecol. 2024, 76, 58–68. [Google Scholar] [CrossRef]
- Liu, Q.; Shi, J.; Ma, Y.; Wang, Y.; Wang, X.; Lyu, L.; He, M.; Cai, Z. Relationship between plant diversity and community stability in alpine mining areas. J. Mt. Sci. 2025, 22, 901–912. [Google Scholar] [CrossRef]
- Dietrich, P.; Ebeling, A.; Meyer, S.T.; Asato, A.E.B.; Broecher, M.; Gleixner, G.; Huang, Y.; Roscher, C.; Schmid, B.; Vogel, A.; et al. Plant diversity and community age stabilize ecosystem multifunctionality. Glob. Change Biol. 2024, 30, e17225. [Google Scholar] [CrossRef] [PubMed]
- Cui, Z.; Sun, J.; Wu, G.-L. Plant diversity increases spatial stability of aboveground productivity in alpine grasslands. Oecologia 2024, 205, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Zhu, Y.; Yang, X.; Shan, D.; Wang, D.; Tu, Y.; Shi, Z.; Indree, T. Effects of plant diversity and community structure on ecosystem multifunctionality under different grazing potentials in the eastern Eurasian steppe. Sci. Total Environ. 2024, 934, 173128. [Google Scholar] [CrossRef] [PubMed]
- English, N.B.; Weltzin, J.F.; Fravolini, A.; Thomas, L.; Williams, D.G. The influence of soil texture and vegetation on soil moisture under rainout shelters in a semi-desert grassland. J. Arid Environ. 2005, 63, 324–343. [Google Scholar] [CrossRef]
- Liu, X.; Chen, X.; Wang, L.; Zhang, Y.; Cheng, Q. Effect of vapour transport on soil evaporation under different soil textures and water table depths in an arid area of Northwest China. Hydrol. Process. 2023, 37, e14821. [Google Scholar] [CrossRef]
- Sager, C.; Airo, A.; Mangelsdorf, K.; Arens, F.L.; Karger, C.; Schulze-Makuch, D. Habitability of Polygonal Soils in the Hyper-Arid Atacama Desert After a Simulated Rain Experiment. J. Geophys. Res. Biogeosci. 2023, 128, e2022JG007328. [Google Scholar] [CrossRef]
- Li, D.; Wang, S.; Li, X.; Mao, X.; Cao, T. Experimental Study on One-dimensional Permeability of Sandy Clayey Purple Soil at Unsaturated Condition. Chin. J. Undergr. Space Eng. 2020, 16, 373–380. [Google Scholar]
- Dai, S.; Shin, H.; Santamarina, J.C. Formation and development of salt crusts on soil surfaces. ACTA Geotech. 2016, 11, 1103–1109. [Google Scholar] [CrossRef]
- Shokri-Kuehni, S.M.S.; Vetter, T.; Webb, C.; Shokri, N. New insights into saline water evaporation from porous media: Complex interaction between evaporation rates, precipitation, and surface temperature. Geophys. Res. Lett. 2017, 44, 5504–5510. [Google Scholar] [CrossRef]
- Sadeghi, M.; Babaeian, E.; Tuller, M.; Jones, S.B. Particle size effects on soil reflectance explained by an analytical radiative transfer model. Remote Sens. Environ. 2018, 210, 375–386. [Google Scholar] [CrossRef]
- Yuan, G.; Zhang, L.; Liu, Y. Impacts of soil moisture and atmospheric moisture transport on the precipitation in two typical regions of China. Atmos. Res. 2021, 247, 105151. [Google Scholar] [CrossRef]
- Su, J.; Zhao, X.; Li, G. Distribution pattern of plant community and its relationship with environmental factors in Qilian Mountains National Nature Reserve of Gansu Province. Guihaia 2022, 42, 450–459. [Google Scholar]
- Zhang, H.; Zou, W.; Chen, Z.; He, L.; Peng, X.; Wang, G.; Peng, P.; Li, J.; Shi, S. Distribution pattern of plant communities and its relationship with environmental factors in eastern Xizang, China. Chin. J. Appl. Environ. Biol. 2023, 29, 1289–1297. [Google Scholar] [CrossRef]
- Wang, Z.; Pan, C. Spatial Distribution of Vegetation Community and Environmental Interpretations in Desert-oasis Ecotone in the Lower Reaches of Tarim River. Xinjiang Agric. Sci. 2009, 46, 449–458. [Google Scholar]
- Lund, E.D.; Wolcott, M.C.; Hanson, G.P. Applying nitrogen site-specifically using soil electrical conductivity maps and precision agriculture technology. Sci. World J. 2001, 1, 767–776. [Google Scholar] [CrossRef] [PubMed]
- Austin, A.T.; Yahdjian, L.; Stark, J.M.; Belnap, J.; Porporato, A.; Norton, U.; Ravetta, D.A.; Schaeffer, S.M. Water pulses and biogeochemical cycles in arid and semiarid ecosystems. Oecologia 2004, 141, 221–235. [Google Scholar] [CrossRef] [PubMed]
- Alaoui, A. A Simple Method to Assess Key Soil Hydraulic Properties. Water 2023, 15, 467. [Google Scholar] [CrossRef]
- Liu, X.; Wang, J.; Zhao, X. Effects of simulated nitrogen deposition on the soil enzyme activities in a Pinus tabulaeformis forest at the Taiyue Mountain. Acta Ecol. Sin 2015, 35, 4613–4624. [Google Scholar]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef]
- Simpson, E.H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Margalef, R. Diversidad de Especies en las Comunidades Naturales; Instituto de Biología Aplicada: Barcelona, Spain, 1951. [Google Scholar]
- Pielou, E.C. The measurement of diversity in different types of biological collections. J. Theor. Biol. 1966, 13, 131–144. [Google Scholar] [CrossRef]
- An, N.; Guo, B.; Zhang, D.; Yang, Q.; Luo, W. Desert vegetation composition and spatial distribution of soil nutrients in the middle section of Hexi Corridor. Arid Zone Res. 2024, 41, 432–443. [Google Scholar]
- Zhou, S.; Duan, Y.; Zhang, Y.; Guo, J. Vegetation dynamics of coal mining city in an arid desert region of Northwest China from 2000 to 2019. J. Arid Land 2021, 13, 534–547. [Google Scholar] [CrossRef]
- Karatassiou, M.; Parissi, Z.M.; Panajiotidis, S.; Stergiou, A. Impact of Grazing on Diversity of Semi-Arid Rangelands in Crete Island in the Context of Climatic Change. Plants 2022, 11, 982. [Google Scholar] [CrossRef] [PubMed]
- Hou, H.; Zhang, Y.; Zhou, J.; Guo, Y.; Liu, H.; Li, Y.; Gao, S. Aridity and Soil Properties Drive the Shrub–Herb Interactions Along Drought Gradient in Desert Grassland in Inner Mongolia. Agronomy 2024, 14, 2588. [Google Scholar] [CrossRef]
- Lundell, S.; Batbaatar, A.; Carlyle, C.N.; Lamb, E.G.; Otfinowski, R.; Schellenberg, M.P.; Bennett, J.A. Plant responses to soil biota depend on precipitation history, plant diversity, and productivity. Ecology 2022, 103, e3784. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhao, M.; Qi, L.; Zhao, C.; Zhang, W.; Zhang, Y.; Wen, W.; Yuan, J. Coupled Relationship between Soil Physicochemical Properties and Plant Diversity in the Process of Vegetation Restoration. Forests 2022, 13, 648. [Google Scholar] [CrossRef]
- Yao, K.; Wang, Y.; Wu, Y. Competition and Niche Differentiation of Water and Nutrients between Broussonetia papyrifera and Platycladus orientalis under Prolonged Drought Stress. Agronomy 2022, 12, 1489. [Google Scholar] [CrossRef]
- Klimek, S.; Marini, L.; Hofmann, M.; Isselstein, J. Additive partitioning of plant diversity with respect to grassland management regime, fertilisation and abiotic factors. Basic Appl. Ecol. 2008, 9, 626–634. [Google Scholar] [CrossRef]
- Du, H.; Fan, P.; Bi, Y.; Xie, S.; Liu, Y.; Liu, Y. Plant Community Characteristics and Their Relationship with Environmental Factors in Different Geomorphic Units in Arid Gravel Desert. Bull. Bot. Res. 2024, 44, 459–469. [Google Scholar] [CrossRef]
- Jarvis, N.J. A review of non-equilibrium water flow and solute transport in soil macropores: Principles, controlling factors and consequences for water quality. Eur. J. Soil Sci. 2020, 71, 279–302. [Google Scholar] [CrossRef]
- Chen, X.; Duan, Z. Changes in soil physical and chemical properties during reversal of desertification in Yanchi County of Ningxia Hui autonomous region, China. Environ. Geol. 2009, 57, 975–985. [Google Scholar] [CrossRef]
- Eltarabily, M.G.; Bali, K.M.; Negm, A.M.; Yoshimura, C. Evaluation of Root Water Uptake and Urea Fertigation Distribution under Subsurface Drip Irrigation. Water 2019, 11, 1487. [Google Scholar] [CrossRef]
- Li, Y.; Jiang, J.; Song, C.; Li, Z. Relationship Between Soil Factors and Ephemeral Plant Species Diversity on Typical Sand Dunes in the Mosuowan Region. Arid Zone Res. 2018, 35, 912–919. [Google Scholar]
- Tan, Q.; Huo, S.; Wang, D.; Wen, M.; Xing, M.; Lu, M. Hysteresis of Water Retention Curve of the Capillarimetric Diapason in Saline Soils. Eurasian Soil Sci. 2024, 57, 1217–1230. [Google Scholar] [CrossRef]
- Ayari, M.; Charef, A.; Azouzi, R.; Khiari, N.; Ben Sassi, M. Effect of Zeolite, Femur and Lignite Application Rates on Soil Hydraulic Properties and Fertility of Sandy and Clay Soils at Semi-arid Climate. Commun. Soil Sci. Plant Anal. 2022, 53, 408–425. [Google Scholar] [CrossRef]
- Guo, C.; Yao, C.; Wu, J.; Qin, S.; Yang, H.; Li, H.; Mao, J. Field and numerical experiments of subsurface drainage systems in saline and low-permeability interlayered fields in arid regions. Agric. Water Manag. 2024, 300, 108898. [Google Scholar] [CrossRef]
- Ye, C.; Zheng, G.; Tao, Y.; Xu, Y.; Chu, G.; Xu, C.; Chen, S.; Liu, Y.; Zhang, X.; Wang, D. Effect of Soil Texture on Soil Nutrient Status and Rice Nutrient Absorption in Paddy Soils. Agronomy 2024, 14, 1339. [Google Scholar] [CrossRef]
- Yu, M.; Zhang, L.; Xu, X.; Feger, K.-H.; Wang, Y.; Liu, W.; Schwaerzel, K. Impact of land-use changes on soil hydraulic properties of Calcaric Regosols on the Loess Plateau, NW China. J. Plant Nutr. Soil Sci. 2015, 178, 486–498. [Google Scholar] [CrossRef]
- Wu, Y.; Zhang, H.; Li, J.; Wang, Z.; Jiang, Y. Adsorption of soil invertase to goethite, gibbsite and their organic complexes and the effects on enzyme catalytic performance. Colloids Surf. B Biointerfaces 2023, 222, 113073. [Google Scholar] [CrossRef] [PubMed]
- Khadem, A.; Raiesi, F. Influence of biochar on potential enzyme activities in two calcareous soils of contrasting texture. Geoderma 2017, 308, 149–158. [Google Scholar] [CrossRef]
- Song, W.; Han, F.; Bao, Z.; Chai, Y.; Wang, L.; Huang, C.; Cheng, H.; Chang, L. Mulching Practices Improve Soil Moisture and Enzyme Activity in Drylands, Increasing Potato Yield. Agronomy 2024, 14, 1077. [Google Scholar] [CrossRef]
- Wu, Y.; Zhao, Z.; Zhao, F.; Cheng, X.; Zhao, P.; Liu, S. Response of Pear Trees’ (Pyrus bretschneideri ‘Sinkiangensis’) Fine Roots to a Soil Water Regime of Regulated Deficit Irrigation. Agronomy 2021, 11, 2316. [Google Scholar] [CrossRef]
- Yu, Q.; Zhang, Z.; He, Y.; Hao, M.; Wang, G.; Dun, X.; Wu, Q.; Gao, P. Secondary shrubs promoted the priming effect by increasing soil particle organic carbon mineralization. Front. For. Glob. Change 2023, 6, 1288259. [Google Scholar] [CrossRef]
- Fischer, C.; Tischer, J.; Roscher, C.; Eisenhauer, N.; Ravenek, J.; Gleixner, G.; Attinger, S.; Jensen, B.; de Kroon, H.; Mommer, L.; et al. Plant species diversity affects infiltration capacity in an experimental grassland through changes in soil properties. Plant Soil 2015, 397, 1–16. [Google Scholar] [CrossRef]
- Tavanti, R.F.R.; Montanari, R.; Panosso, A.R.; Freddi, O.S.; Paz-Gonzalez, A. Pedotransfer function to estimate the soil structural “s” index and spatial variability in an oxisol within a livestock farming system. Eng. Agric. 2020, 40, 34–44. [Google Scholar] [CrossRef]
- Lin, H.S.; McInnes, K.J.; Wilding, L.P.; Hallmark, C.T. Effects of soil morphology on hydraulic properties: II. Hydraulic pedotransfer functions. Soil Sci. Soc. Am. J. 1999, 63, 955–961. [Google Scholar] [CrossRef]
- Baveye, P. Comment on “Soil structure and management: A review” by C.J. Bronick and R. Lal. Geoderma 2006, 134, 231–232. [Google Scholar] [CrossRef]
- Binh Thanh, N.; Binh Trung, P.; Tong Xuan, N.; Vinh Ngoc, N.; Thanh Van, T.; Quang-Vu, B. Contrastive nutrient leaching from two differently textured paddy soils as influenced by biochar addition. J. Soils Sediments 2020, 20, 297–307. [Google Scholar] [CrossRef]
- Juice, S.M.; Schaberg, P.G.; Kosiba, A.M.; Waite, C.E.; Hawley, G.J.; Wang, D.; Perdrial, J.N.; Adair, E.C. Soil type modifies the impacts of warming and snow exclusion on leachate carbon and nutrient losses. Biogeochemistry 2022, 160, 199–217. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, S.; Wang, Y.; Li, Z.; Li, C. Soil Texture’s Hidden Influence: Decoding Plant Diversity Patterns in Arid Ecosystems. Soil Syst. 2025, 9, 84. https://doi.org/10.3390/soilsystems9030084
Wang S, Wang Y, Li Z, Li C. Soil Texture’s Hidden Influence: Decoding Plant Diversity Patterns in Arid Ecosystems. Soil Systems. 2025; 9(3):84. https://doi.org/10.3390/soilsystems9030084
Chicago/Turabian StyleWang, Shuaiyu, Younian Wang, Zhiwei Li, and Chengzhi Li. 2025. "Soil Texture’s Hidden Influence: Decoding Plant Diversity Patterns in Arid Ecosystems" Soil Systems 9, no. 3: 84. https://doi.org/10.3390/soilsystems9030084
APA StyleWang, S., Wang, Y., Li, Z., & Li, C. (2025). Soil Texture’s Hidden Influence: Decoding Plant Diversity Patterns in Arid Ecosystems. Soil Systems, 9(3), 84. https://doi.org/10.3390/soilsystems9030084