
Improvement of Salinity Tolerance in Rice Seedlings by Exogenous Magnesium Sulfate Application

 ,
,  ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Results
2.1. Phenotypic Performances
2.2. Chemical Performance
3. Discussion
4. Materials and Methods
4.1. Materials and Treatments
4.2. Phenotypic and Chemical Measurements
4.3. Identification and Quantification of Phenolics and Momilactones A and B
4.4. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Jaiswal, S.; Gautam, R.K.; Singh, R.K.; Krishnamurthy, S.L.; Ali, S.; Sakthivel, K.; Iquebal, M.A.; Rai, A.; Kumar, D. Harmonizing technological advances in phenomics and genomics for enhanced salt tolerance in rice from a practical perspective. Rice 2019, 12, 89. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Guo, Y. Unraveling salt stress signaling in plants. J. Integr. Plant Biol. 2018, 60, 796–804. [Google Scholar] [CrossRef] [PubMed]
- Tester, M.; Davenport, R. Na+ tolerance and Na+ transport in higher plants. Ann. Bot. 2003, 91, 503–527. [Google Scholar] [CrossRef] [PubMed]
- Munns, R. Plant adaptations to salt and water stress: Differences and commonalities. Adv. Bot. Res. 2011, 57, 1–32. [Google Scholar]
- Eynard, A.; Lal, R.; Wiebe, K. Crop response in salt-affected soils. J. Sustain. Agric. 2005, 27, 5–50. [Google Scholar] [CrossRef]
- Roy, S.J.; Negrao, S.; Tester, M. Salt resistant crop plants. Curr. Opin. Biotechnol 2014, 26, 115–124. [Google Scholar] [CrossRef]
- Huong, C.T.; Anh, T.T.T.; Tran, H.D.; Duong, V.X.; Trung, N.T.; Khanh, T.D.; Xuan, T.D. Assessing salinity tolerance in rice mutants by phenotypic evaluation alongside simple sequence repeat analysis. Agriculture 2020, 10, 191. [Google Scholar] [CrossRef]
- Hussain, S.; Bai, Z.; Huang, J.; Cao, X.; Zhu, L.; Zhu, C.; Khaskheli, M.A.; Zhong, C.; Zhang, J. 1-Methylcyclopropene modulates physiological, biochemical, and antioxidant responses of rice to different salt stress levels. Front. Plant Sci. 2019, 10, 124. [Google Scholar] [CrossRef]
- Bhusan, D.; Das, K.D.; Hossain, M.; Murata, Y.; Hoque, M.A. Improvement of salt tolerance in rice (Oryza sativa L.) by increasing antioxidant defence systems using exogenous application of proline. Aust. J. Crop Sci. 2016, 10, 50–56. [Google Scholar]
- Ashraf, M.; Athar, H.R.; Harris, J.C.; Kwon, T.R. Some prospective strategies for improving crop salt tolerance. Adv. Agron. 2008, 97, 45–110. [Google Scholar]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.M.; Roychowdhury, R.; Fujita, M. Physiological, biochemical, and molecular mechanisms of heat stress tolerance in plants. Int. J. Mol. Sci. 2013, 14, 9643–9684. [Google Scholar] [CrossRef] [PubMed]
- Mishra, P.; Bhoomika, K.; Dubey, R.S. Differential responses of antioxidative defense system to prolonged salinity stress in salt-tolerant and salt-sensitive Indica rice (Oryza sativa L.) seedlings. Protoplasma 2013, 250, 3–19. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, N.; Umar, S.; Khan, N.A. Nitrogen availability regulates proline and ethylene production and alleviates salinity stress in mustard (Brassica juncea). J. Plant Physiol. 2015, 178, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Bistgani, Z.E.; Hashemi, M.; Dacosta, M.; Craker, L.; Maggi, F.; Morshedloo, M.R. Effect of salinity stress on the physiological characteristics, phenolic compounds and antioxidant activity of Thymus vulgaris L. and Thymus daenensis Celak. Ind. Crop Prod. 2019, 135, 311–320. [Google Scholar] [CrossRef]
- Mahdavi, V.; Farimani, M.M.; Fathi, F.; Ghassempour, A. A targeted metabolomics approach toward understanding metabolic variations in rice under pesticide stress. Anal. Biochem. 2015, 478, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Thakur, S.; Kumar, V.; Kanwar, M.K.; Kesavan, A.K.; Thukral, A.K.; Bhardwaj, R.; Alam, P.; Ahmad, P. Pre-sowing seed treatment with 24-epibrassinolide ameliorates pesticide stress in Brassica juncea L. through the modulation of stress markers. Front. Plant Sci. 2016, 7, 1569. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Yuan, H.; Kumar, V.; Ramakrishnan, M.; Kohli, S.K.; Kaur, R.; Thukral, A.K.; Bhardwaj, R.; Zheng, B. Cas-tasterone attenuates insecticide induced phytotoxicity in mustard. Ecotoxicol. Environ. Saf. 2019, 179, 50–61. [Google Scholar] [CrossRef]
- Rayee, R.; Tran, H.D.; Xuan, T.D.; Khanh, T.D. Imposed water deficit after anthesis for the improvement of macronutrients, quality, phytochemicals, and antioxidants in rice grain. Sustainability 2018, 10, 4843. [Google Scholar] [CrossRef]
- Hartwig, A. Role of magnesium in genomic stability. Mutat. Res. 2001, 18, 113–121. [Google Scholar] [CrossRef]
- Soetan, K.O.; Olaiya, C.O.; Oyewole, O.E. The importance of mineral elements for humans, domestic animals and plants: A review. Afr. J. Food Sci. 2010, 4, 200–222. [Google Scholar]
- Ali, H.; Sarwar, N.; Muhammad, S.; Farooq, O.; Rehman, A.; Wasaya, A.; Yasir, T.A.; Mubeen, K.; Akhtar, M.N. Foliar application of magnesium at critical stages improved the productivity of rice crop grown under different cultivation systems. Sustainability 2021, 13, 4962. [Google Scholar] [CrossRef]
- Farhangi-Abriz, S.; Ghassemi-Golezani, K. Changes in soil properties and salt tolerance of safflower in response to biochar-based metal oxide nanocomposites of magnesium and manganese. Ecotoxicol. Environ. Saf. 2021, 211, 111904. [Google Scholar] [CrossRef] [PubMed]
- Rivelli, A.R.; De Maria, S.; Pizza, S.; Gherbin, P. Growth and physiological response of hydroponically-grown sunflower as affected by salinity and magnesium levels. J. Plant Nutr. 2010, 33, 1307–1323. [Google Scholar] [CrossRef]
- de Brito Neto, J.F.; Beltrão, N.D.E.M.; Rigon, J.P.G.; Capuani, S. Effect of calcium and magnesium silicate on the growth of the castor oil plant subjected to salinity levels. Semin. Cienc. Agrar. 2012, 33, 2949–2958. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.C.; Yamaji, N.; Horie, T.; Che, J.; Li, J.; An, G.; Ma, J.F. A magnesium transporter OSMGT1 plays a critical role in salt tolerance in rice. Plant Physiol. 2017, 174, 1837–1849. [Google Scholar] [CrossRef] [PubMed]
- Rahman, A.; Nahar, K.; Mahmud, J.A.; Hasanuzzaman, M.; Hossain, S.; Fujita, M. Salt stress tolerance in rice: Emerging role of exogenous phytoprotectants. In Advances in International Rice Research; Li, J., Ed.; IntechOpen: London, UK, 2017; Volume 9, pp. 139–174. [Google Scholar]
- Šamec, D.; Karalija, E.; Šola, I.; Vujčić, B.V.; Salopek-Sondi, B. The role of polyphenols in abiotic stress response: The influence of molecular structure. Plants 2021, 10, 118. [Google Scholar] [CrossRef]
- Minh, L.T.; Khang, D.T.; Thu Ha, P.T.; Tuyen, P.T.; Minh, T.N.; Quan, N.V.; Xuan, T.D. Effects of salinity stress on growth and phenolics of rice (Oryza sativa L.). Int. Lett. Nat. Sci. 2016, 57, 1–10. [Google Scholar] [CrossRef]
- Gupta, P.; De, B. Differential responses of cell wall bound phenolic compounds in sensitive and tolerant varieties of rice in response to salinity. Plant Signal. Behav. 2017, 12, e1379643. [Google Scholar] [CrossRef]
- Wiszniewska, A.; Koźmińska, A.; Hanus-Fajerska, E.; Dziurka, K.; Dziurka, M. Insight into phytohormonal modulation of defense mechanisms to salt excess in a halophyte and a glycophyte from Asteraceae family. Plant Soil 2021, 463, 55–76. [Google Scholar] [CrossRef]
- Dempsey, D.A.; Klessig, D.F. How does the multifaceted plant hormone salicylic acid combat disease in plants and are similar mechanisms utilized in humans? BMC Biol. 2017, 15, 23. [Google Scholar] [CrossRef]
- Xuan, T.D.; Minh, T.N.; Anh, L.H.; Khanh, T.D. Allelopathic momilactones A and B are implied in rice drought and salinity tolerance, not weed resistance. Agro. Sust. Dev. 2016, 36, 52. [Google Scholar] [CrossRef]
- Kim, Y.H.; Khan, A.L.; Waqas, M.; Shim, J.K.; Kim, D.H.; Lee, K.Y.; Lee, I.J. Silicon application to rice root zone influenced the phytohormonal and antioxidant responses under salinity stress. J. Plant Growth Regul. 2014, 33, 137–149. [Google Scholar] [CrossRef]
- White, P.J.; Broadley, M.R. Biofortification of crops with seven mineral elements often lacking in human diets—Iron, zinc, copper, calcium, magnesium, selenium and iodine. New Phytol. 2009, 182, 49–84. [Google Scholar] [CrossRef] [PubMed]
- Cowan, J.A. Structural and catalytic chemistry of magnesium-dependent enzymes. Biometals 2002, 15, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Sreedhara, A.; Cowan, J.A. Structural and catalytic roles for divalent magnesium in nucleic acid biochemistry. Biometals 2002, 15, 211–223. [Google Scholar] [CrossRef]
- Marschner, H. Mineral Nutrition of Higher Plants, 3rd ed.; Academic Press: London, UK, 2012; pp. 277–284. [Google Scholar]
- Moomaw, A.S.; Maguire, M.E. The unique nature of Mg2+ channels. Physiology 2009, 23, 275–285. [Google Scholar] [CrossRef]
- Guo, W.; Nazim, H.; Liang, Z.; Yang, D. Magnesium deficiency in plants: An urgent problem. Crop J. 2016, 4, 83–91. [Google Scholar] [CrossRef]
- Gregorio, G.B.; Senadhira, D.; Mendoza, R.D. Screening rice for salinity tolerance. IRRI Discuss. Pap. Ser. 1997, 22, 1–30. [Google Scholar]
- IRRI. Phenotyping Protocol for Abiotic Stress Tolerance in Rice; International Rice Research Institute: Los Banos, Philippines, 2021; 65p. [Google Scholar]
- Govindarajan, R.; Rastogi, S.; Vijayakumar, M.; Shirwaikar, A.; Rawat, A.K.; Mehrotra, S.; Pushpangadan, P. Studies on the antioxidant activities of Desmodium gangeticum. Biol. Pharm. Bull. 2003, 26, 1424–1427. [Google Scholar] [CrossRef]
- Quan, N.V.; Xuan, T.D.; Tran, H.D.; Thuy, N.T.D.; Trang, L.T.; Huong, C.T.; Yusuf, A.; Tuyen, P.T. Antioxidant, α-amylase and α-glucosidase inhibitory activities and potential constituents of Canarium tramdenum Bark. Molecules 2019, 24, 605. [Google Scholar] [CrossRef]
- Quan, N.V.; Thien, D.D.; Khanh, T.D.; Tran, H.D.; Xuan, T.D. Momilactones A, B, and tricin in rice grain and by-products are potential skin aging inhibitors. Foods 2019, 8, 602. [Google Scholar] [CrossRef] [PubMed]
- Quan, N.V.; Hoang-Dung, T.; Xuan, T.D.; Ahmad, A.; Dat, T.D.; Khanh, T.D.; Teschke, R. Momilactones A and B are α-amylase and α-glucosidase inhibitors. Molecules 2019, 24, 482. [Google Scholar] [CrossRef] [PubMed]
- Quan, N.V.; Xuan, T.D.; Hoang-Dung, T.; Ahmad, A.; Khanh, T.D.; Dat, T.D. Contribution of momilactones A and B to diabetes inhibitory potential of rice bran: Evidence from in vitro assays. Saudi Pharm. J. 2019, 27, 643–649. [Google Scholar] [CrossRef] [PubMed]



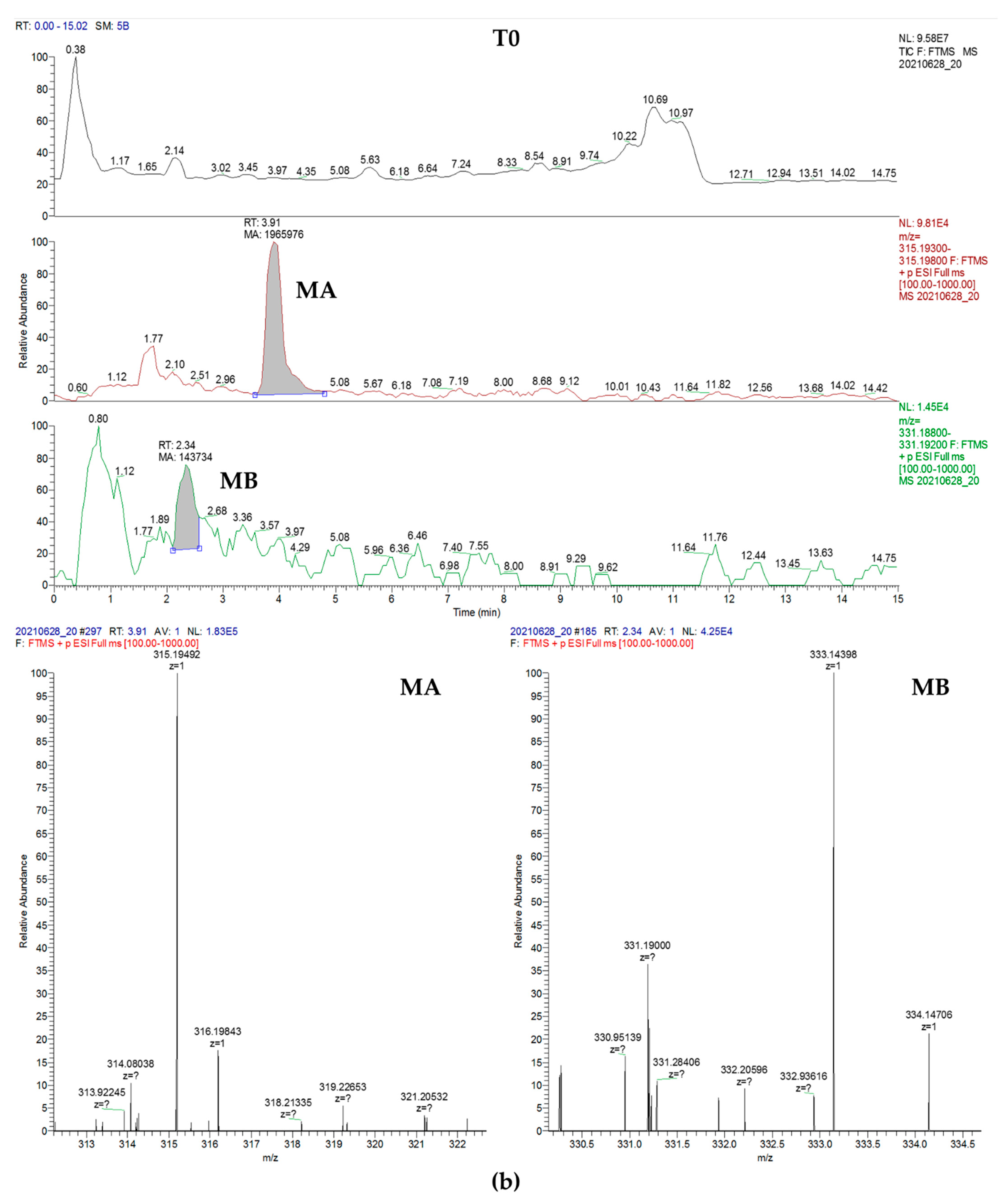

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Injury Score | Root Length (mm) | Plant Height (mm) | Fresh Weight (mg) | Dry Weight (mg) |
|---|---|---|---|---|---|
| T0 | 1.00 ± 0.00 e | 64.93 ± 0.55 ab | 190.67 ± 1.63 a | 97.57 ± 0.64 a | 41.43 ± 0.67 a |
| T1 | 3.43 ± 0.10 c | 65.37 ± 1.44 a | 170.83 ± 2.26 b | 79.67 ± 0.58 c | 36.00 ± 1.00 bc |
| T2 | 3.00 ± 0.00 d | 65.30 ± 0.38 a | 175.10 ± 2.70 ab | 90.93 ± 0.41 b | 38.97 ± 2.37 b |
| S0 | 1.00 ± 0.00 e | 59.80 ± 2.17 bc | 187.43 ± 3.60 a | 96.56 ± 0.58 a | 39.67 ± 0.41 a |
| S1 | 5.23 ± 0.07 a | 58.03 ± 0.70 c | 145.63 ± 0.20 c | 70.47 ± 0.58 d | 26.67 ± 0.88 d |
| S2 | 4.73 ± 0.03 b | 60.00 ± 0.56 abc | 166.97 ± 2.32 bc | 80.60 ± 0.90 c | 28.57 ± 1.33 c |
| Samples | Caffeic Acid | ρ-Coumaric Acid | Salicylic Acid | Cinnamic Acid | Benzoic Acid | Ferulic Acid | Tricin | MA | MB |
|---|---|---|---|---|---|---|---|---|---|
| T0 | 0.05 ± 0.00 b | 0.12 ± 0.01 d | 0.18 ± 0.01 d | 0.04 ± 0.00 b | 0.65 ± 0.03 d | 0.11 ± 0.00 d | 0.03 ± 0.00 cd | 3.72 ± 0.07 a | 1.45 ± 0.11 a |
| T1 | 0.01 ± 0.00 c | 0.16 ± 0.00 c | 0.29 ± 0.00 d | 0.01 ± 0.00 d | 0.62 ± 0.02 de | 0.15 ± 0.01 c | 0.02 ± 0.00 e | - | - |
| T2 | 0.04 ± 0.00 b | 0.17 ± 0.00 bc | 0.49 ± 0.03 c | 0.02 ± 0.00 cd | 1.00 ± 0.04 c | 0.24 ± 0.01 b | 0.09 ± 0.00 a | - | 0.61 ± 0.01 b |
| S0 | 0.23 ± 0.01 a | 0.47 ± 0.01 a | 2.45 ± 0.05 a | 0.07 ± 0.01 a | 2.74 ± 0.10 a | 0.34 ± 0.01 a | 0.04 ± 0.00 bc | - | 0.68 ± 0.04 b |
| S1 | 0.02 ± 0.00 c | 0.09 ± 0.00 e | 0.45 ± 0.02 c | 0.02 ± 0.00 cd | 0.42 ± 0.02 e | 0.10 ± 0.00 d | 0.03 ± 0.00 de | - | - |
| S2 | 0.01 ± 0.00 c | 0.18 ± 0.01 b | 0.76 ± 0.01 b | 0.03 ± 0.00 bc | 1.35 ± 0.02 b | 0.16 ± 0.00 c | 0.05 ± 0.00 de | - | 0.53 ± 0.01 b |
| Treatment Code | Description |
|---|---|
| T0 | BC15-Control |
| T1 | BC15-Stress (100 mM NaCl) |
| T2 | BC15-Stress & Treatment (100 mM NaCl + 0.5 mM MgSO4) |
| S0 | DT84DB-Control |
| S1 | DT84DB-Stress (100 mM NaCl) |
| S2 | DT84DB-Stress snd Treatment (100 mM NaCl + 0.5 mM MgSO4) |
| Score | Observation | Tolerance |
|---|---|---|
| 1 | Normal growth, no leaf symptoms | Highly tolerant |
| 3 | Nearly normal growth, but leaf tips or a few leaves whitish and rolled | Tolerant |
| 5 | Growth severely retarded most leaves rolled, only a few are elongating | Moderately tolerant |
| 7 | Complete cessation of growth, most leaves dried, some plants are dying | Susceptible |
| 9 | Almost all plants dead or dying | Highly susceptible |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xuan, T.D.; Huong, C.T.; Quan, N.V.; Anh, L.H.; Khanh, T.D.; Rayee, R. Improvement of Salinity Tolerance in Rice Seedlings by Exogenous Magnesium Sulfate Application. Soil Syst. 2022, 6, 69. https://doi.org/10.3390/soilsystems6030069
Xuan TD, Huong CT, Quan NV, Anh LH, Khanh TD, Rayee R. Improvement of Salinity Tolerance in Rice Seedlings by Exogenous Magnesium Sulfate Application. Soil Systems. 2022; 6(3):69. https://doi.org/10.3390/soilsystems6030069
Chicago/Turabian StyleXuan, Tran Dang, Can Thu Huong, Nguyen Van Quan, La Hoang Anh, Tran Dang Khanh, and Ramin Rayee. 2022. "Improvement of Salinity Tolerance in Rice Seedlings by Exogenous Magnesium Sulfate Application" Soil Systems 6, no. 3: 69. https://doi.org/10.3390/soilsystems6030069
APA StyleXuan, T. D., Huong, C. T., Quan, N. V., Anh, L. H., Khanh, T. D., & Rayee, R. (2022). Improvement of Salinity Tolerance in Rice Seedlings by Exogenous Magnesium Sulfate Application. Soil Systems, 6(3), 69. https://doi.org/10.3390/soilsystems6030069