
Effects of Different Land Use Types and Soil Depth on Soil Nutrients and Soil Bacterial Communities in a Karst Area, Southwest China

 , , , ,
, , , ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site Description and Soil Sampling
2.2. Soil Nutrients Analyses
2.3. High-Throughput Absolute Abundance Quantification 16S-seq
2.4. 16S rRNA Gene Sequence Analysis
2.5. Statistical Analyses
3. Results
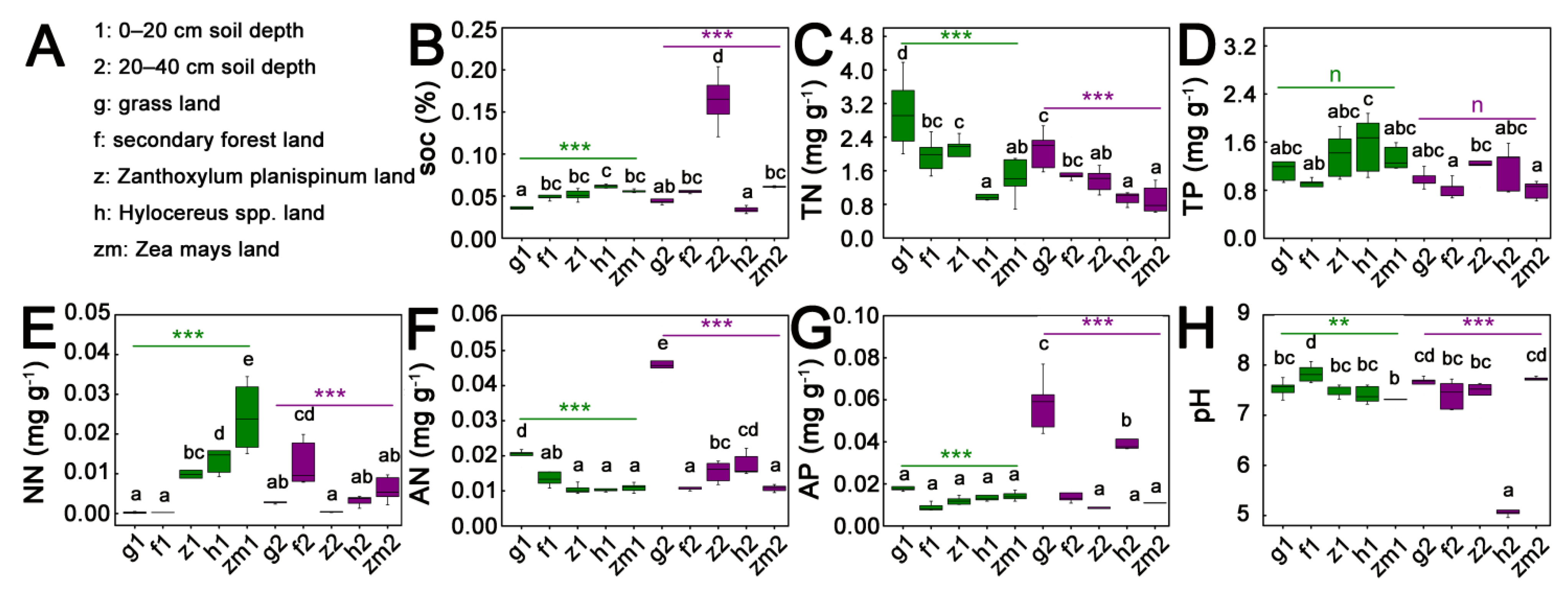
3.1. Soil Nutrient Status of the Different Depth under the Five Different Land Use Types
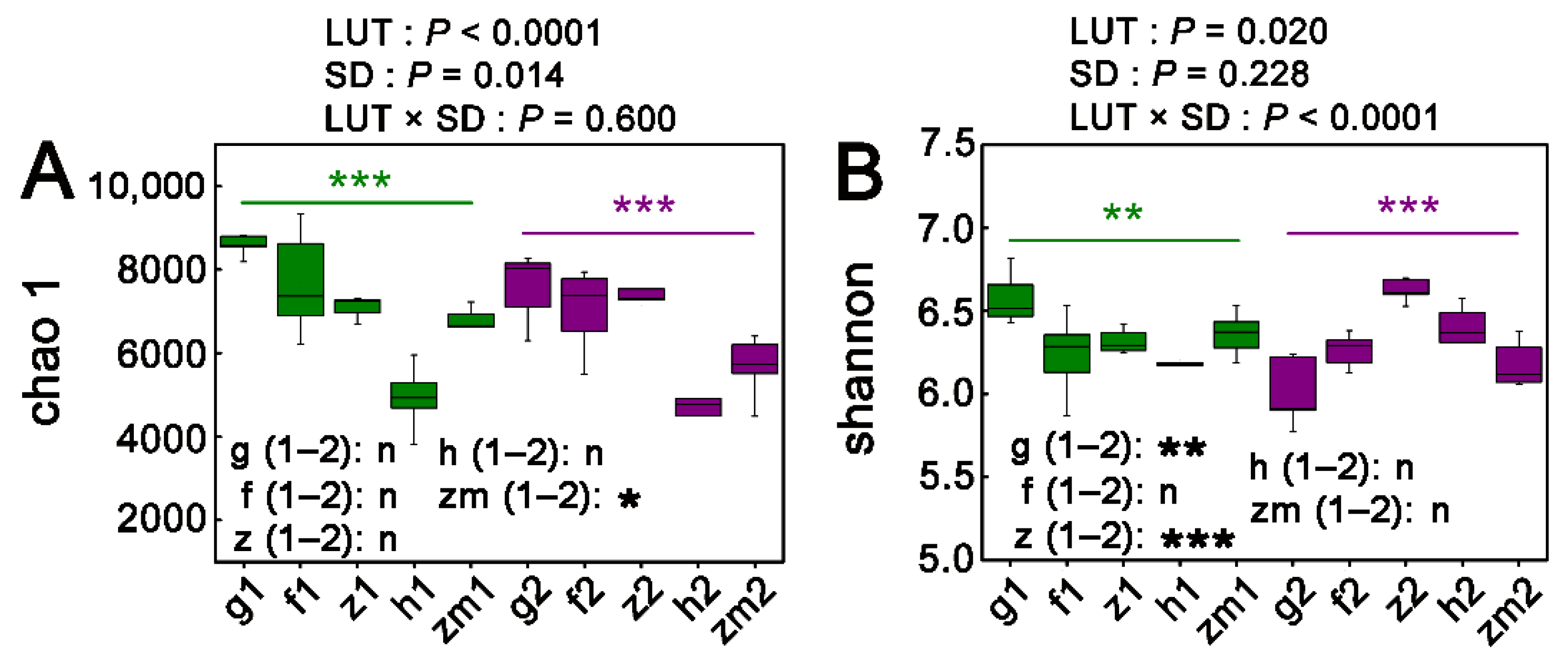
3.2. Alpha Diversity Patterns
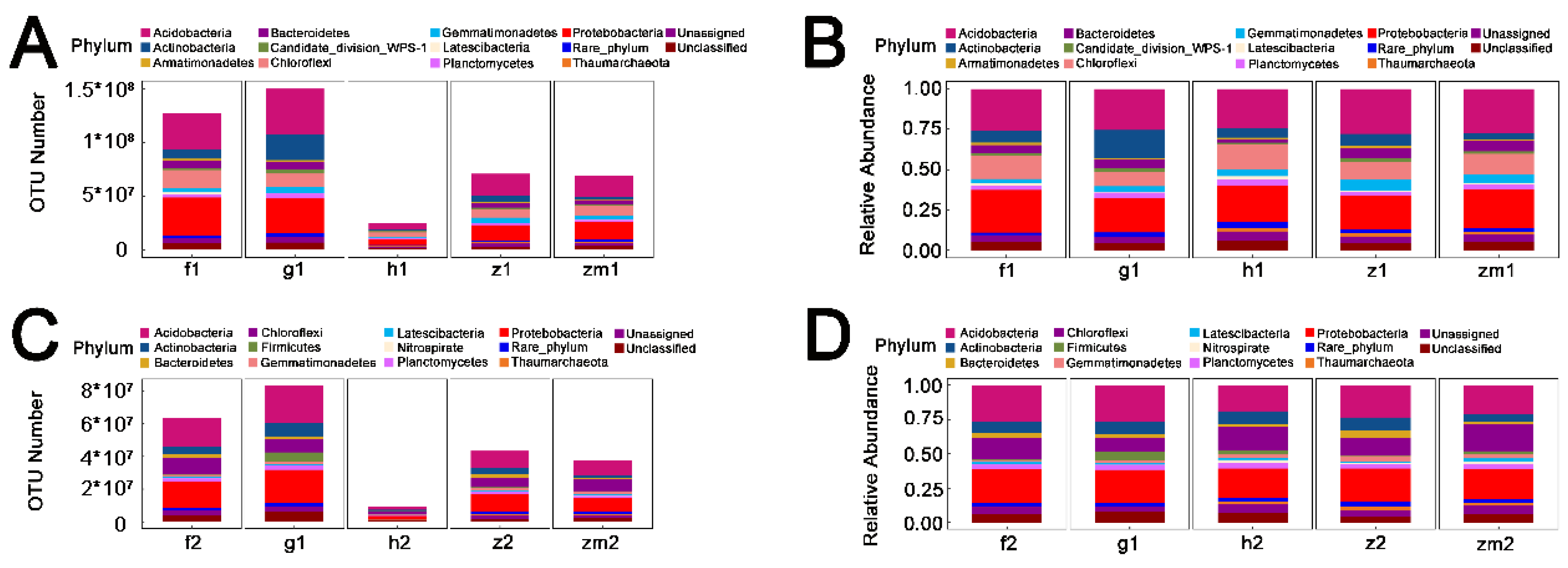
3.3. Absolute Quantification of Soil Bacterial Community
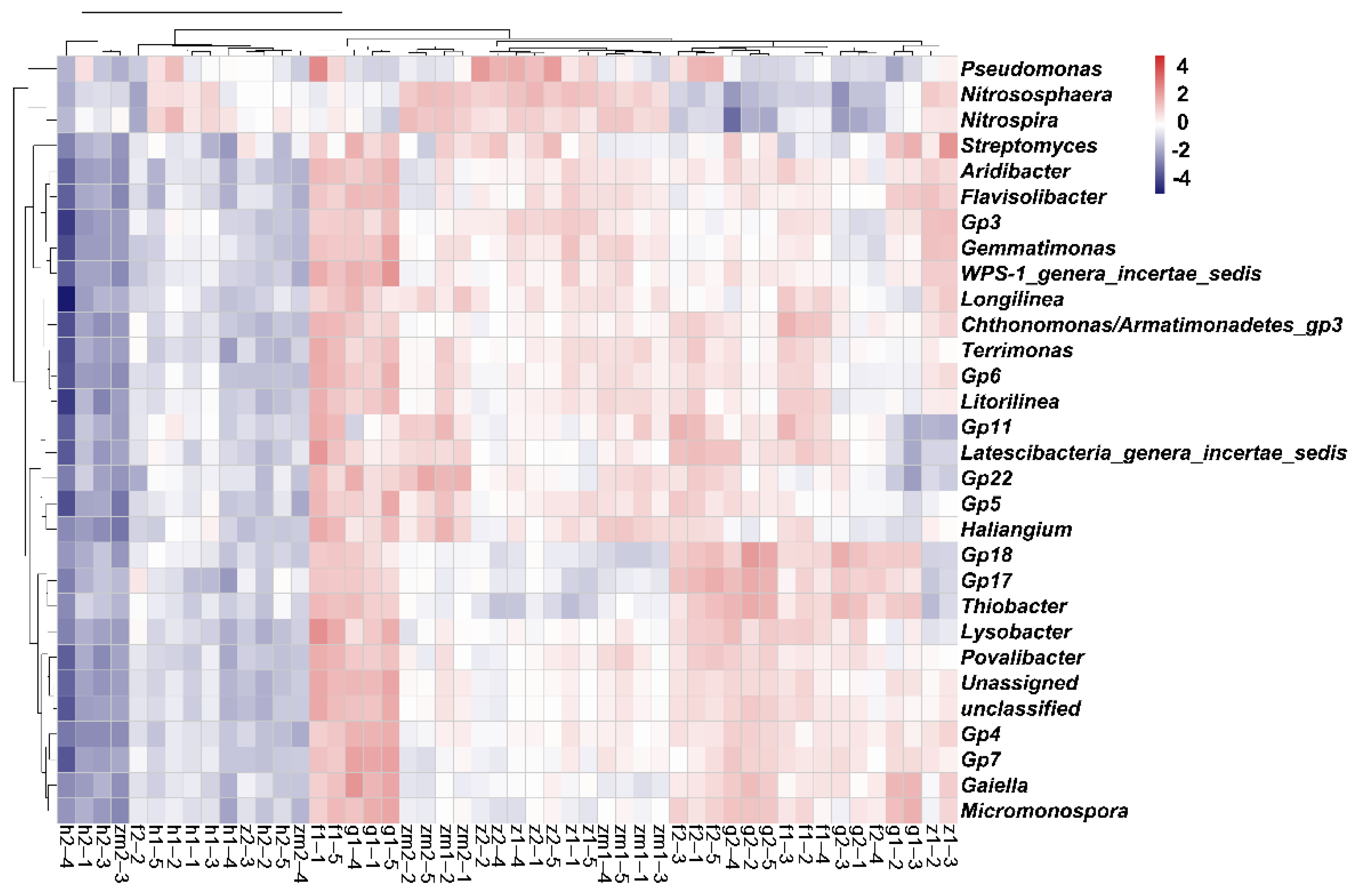
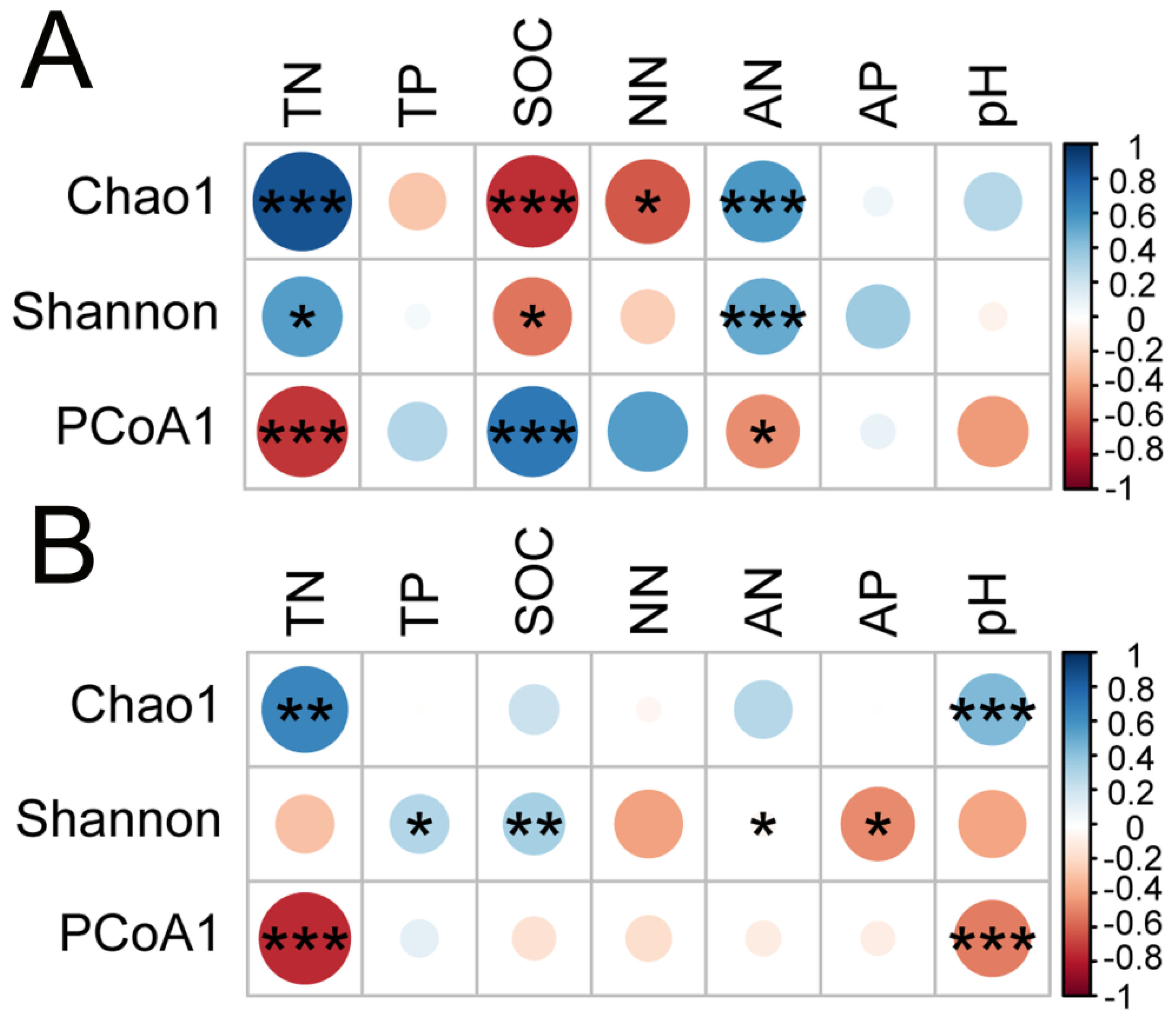
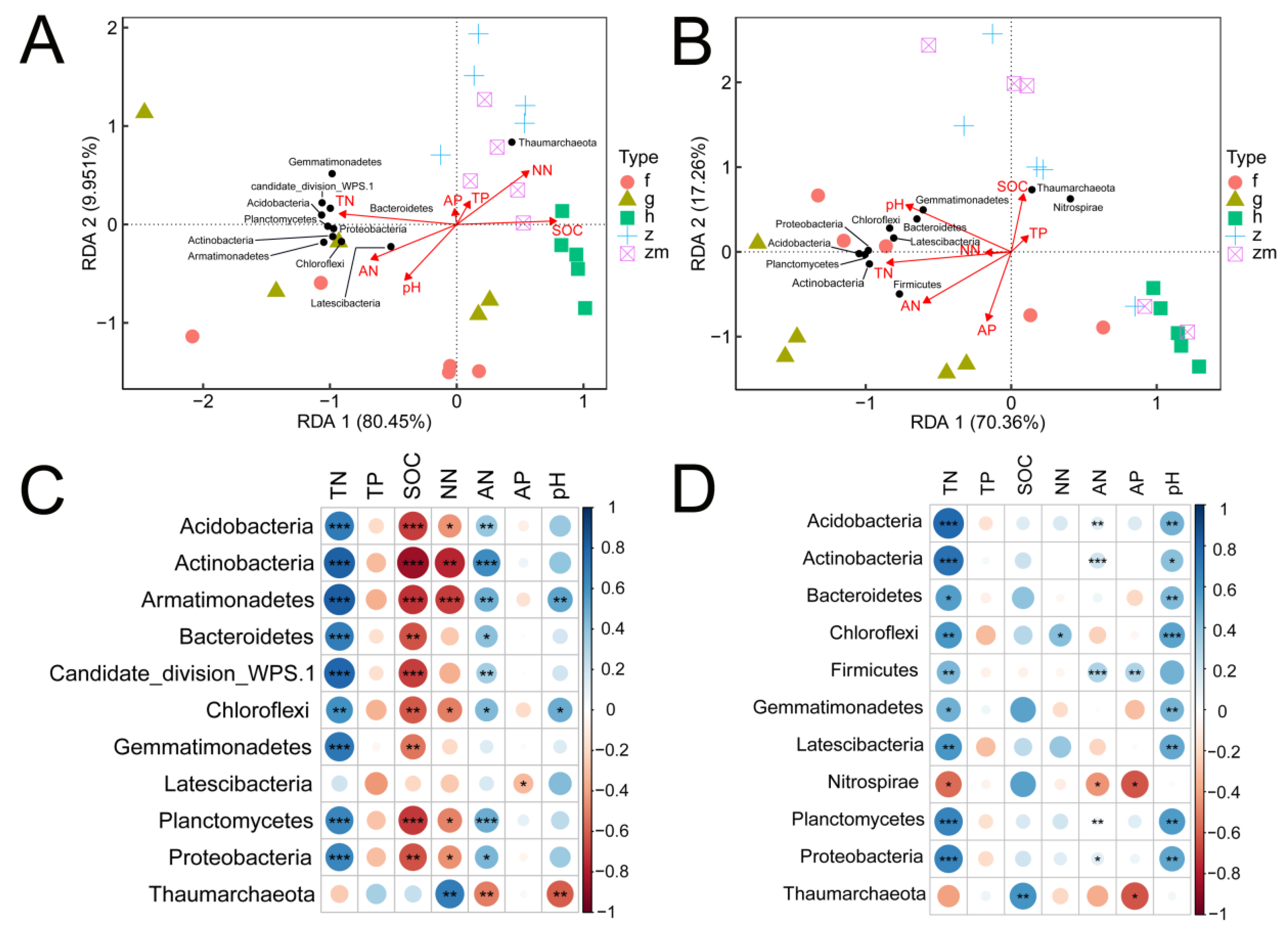
3.4. Relevance of Soil Bacterial Communities and Diversity to Soil Nutrients
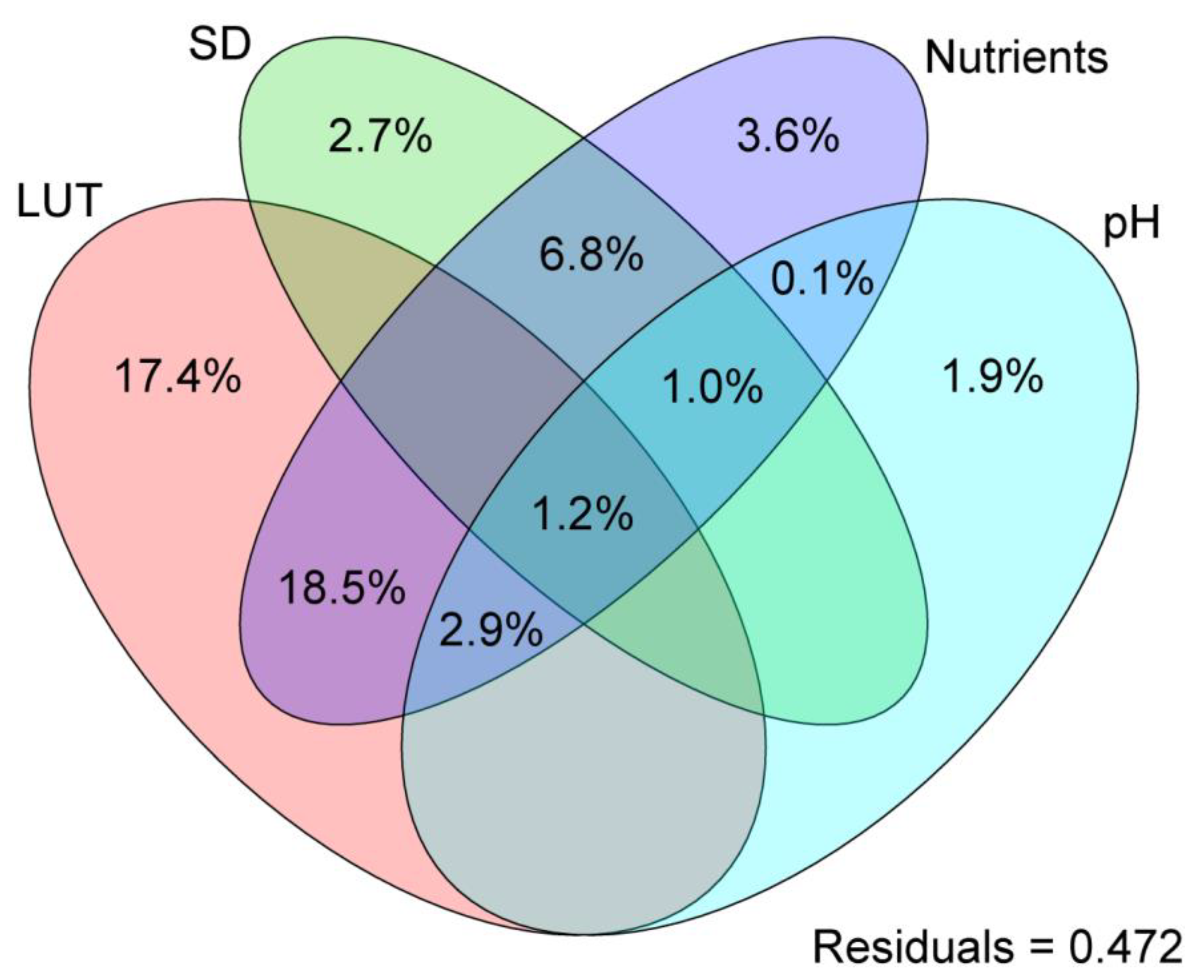
3.5. Structural Equation Model of Soil Nutrients, pH, Land Use Types, and Soil Depth for Bacterial Diversity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sweeting, M.M. Karst in China: Its Geomorphology and Environment; Barsch, D., Ed.; Science & Business Media Springer: Berlin/Heidelberg, Germany, 2012; pp. 242–248. [Google Scholar]
- Huang, Y.; Zhao, P.; Zhang, Z.; Li, X.; He, C.; Zhang, R. Transpiration of Cyclobalanopsis glauca (syn. Quercus glauca) stand measured by sap-flow method in a karst rocky terrain during dry season. Ecol. Res. 2009, 24, 791–801. [Google Scholar] [CrossRef]
- Qi, X.; Wang, K.; Zhang, C. Effectiveness of ecological restoration projects in a karst region of southwest China assessed using vegetation succession mapping. Ecol. Eng. 2013, 54, 245–253. [Google Scholar] [CrossRef]
- Yang, Q.; Jiang, Z.; Ma, Z.; Luo, W.; Xie, Y.; Cao, J. Relationship between karst rocky desertification and its distance to roadways in a typical karst area of Southwest China. Environ. Earth Sci. 2013, 70, 295–302. [Google Scholar] [CrossRef]
- Bárány-Kevei, I. Genetic types, human impact and protection of Hungarian karsts. Acta Climatol. Chorol. Univ. Szeged. 2005, 38–39. [Google Scholar]
- Yuan, D.X. Environmental change and human impact on karst in southern China. Catena. Suppl. 1993, 25, 99–107. [Google Scholar]
- Chen, H.; Zhang, W.; Wang, K.; Hou, Y. Soil organic carbon and total nitrogen as affected by land use types in karst and non-karst areas of northwest Guangxi, China. J. Sci. Food Agric. 2011, 92, 1086–1093. [Google Scholar] [CrossRef] [PubMed]
- Knáb, M.; Szili-Kovács, T.; Márialigeti, K.; Móga, J.; Borsodi, A.K. Bacterial diversity in soils of different Hungarian karst areas. Acta Microbiol. Immunol. Hung. 2018, 65, 439–458. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Hu, Q.; Zhang, C.; Müller, W.E.G.; Schröder, H.C.; Li, Z.; Zhang, Y.; Liu, C.; Jin, Z. The effect of toxicity of heavy metals contained in tailing sands on the organic carbon metabolic activity of soil microorganisms from different land use types in the karst region. Environ. Earth Sci. 2015, 74, 6747–6756. [Google Scholar] [CrossRef]
- Wang, P.; Mo, B.; Chen, Y.; Zeng, Q.; Wang, L. Effect of karst rocky desertification on soil fungal communities in Southwest China. Genet. Mol. Res. 2016, 15. [Google Scholar] [CrossRef] [PubMed]
- Blagodatskaya, E.; Kuzyakov, Y. Active microorganisms in soil: Critical review of estimation criteria and approaches. Soil Biol. Biochem. 2013, 67, 192–211. [Google Scholar]
- Chow, M.L.; Radomski, C.C.; McDermott, J.M.; Davies, J.; Axelrood, P.E. Molecular characterization of bacterial diversity in Lodgepole pine (Pinus contorta) rhizosphere soils from British Columbia forest soils differing in disturbance and geographic source. FEMS Microbiol. Ecol. 2002, 42, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Fan, Z.; Lu, S.; Liu, S.; Guo, H.; Wang, T.; Zhou, J.; Peng, X. Changes in plant rhizosphere microbial communities under different vegetation restoration patterns in karst and Non-karst Ecosystems. Sci. Rep. 2019, 9, 1–12. [Google Scholar]
- McNeill, A.; Unkovich, M. Nutrient Cycling in Terrestrial Ecosystems; Marschner, P., Rengel, Z., Eds.; Springer: Berlin/Heidelberg, Germany, 2007; pp. 37–64. [Google Scholar]
- Mishra, A.; Nautiyal, C.S. Functional diversity of the microbial community in the rhizosphere of chickpea grown in diesel fuel-spiked soil amended with Trichoderma ressei using sole-carbon-source utilization profiles. World J. Microbiol. Biotechnol. 2009, 25, 1175–1180. [Google Scholar] [CrossRef]
- Zhang, H.; Li, G.; Song, X.; Yang, D.; Li, Y.; Qiao, J.; Zhang, J.; Zhao, S. Changes in soil microbial functional diversity under different vegetation restoration patterns for Hulunbeier Sandy Land. Acta Ecol. Sin. 2013, 33, 38–44. [Google Scholar] [CrossRef]
- Xu, Z.; Yu, G.; Zhang, X.; Ge, J.; He, N.; Wang, Q.; Wang, D. The variations in soil microbial communities, enzyme activities and their relationships with soil organic matter decomposition along the northern slope of Changbai Mountain. Appl. Soil Ecol. 2015, 86, 19–29. [Google Scholar] [CrossRef]
- Frey, S.D.; Knorr, M.; Parrent, J.L.; Simpson, R.T. Chronic nitrogen enrichment affects the structure and function of the soil microbial community in temperate hardwood and pine forests. For. Ecol. Manag. 2004, 196, 159–171. [Google Scholar] [CrossRef]
- Fierer, N.; Jackson, R.B. The diversity and biogeography of soil bacterial communities. Proc. Natl. Acad. Sci. USA 2006, 103, 626–631. [Google Scholar] [CrossRef] [Green Version]
- Aziz, I.; Mahmood, T.; Islam, K.R. Effect of long term no-till and conventional tillage practices on soil quality. Soil Tillage Res. 2013, 131, 28–35. [Google Scholar] [CrossRef]
- Navarro-Noya, Y.E.; Gómez-Acata, S.; Montoya-Ciriaco, N.; Rojas-Valdez, A.; Suárez-Arriaga, M.C.; Valenzuela-Encinas, C.; Jiménez-Bueno, N.; Verhulst, N.; Govaerts, B.; Dendooven, L. Relative impacts of tillage, residue management and crop-rotation on soil bacterial communities in a semi-arid agroecosystem. Soil Biol. Biochem. 2013, 65, 86–95. [Google Scholar] [CrossRef]
- Sala, O.E.; Chapin, F.S., III; Armesto, J.J.; Berlow, E.; Bloomfield, J.; Dirzo, R.; Huber-Sanwald, E.; Huenneke, L.F.; Jackson, R.B.; Kinzig, A.; et al. Global Biodiversity Scenarios for the Year 2100. Science 2000, 287, 1770–1774. [Google Scholar] [CrossRef] [PubMed]
- van der Wal, A.; van Veen, J.A.; Smant, W.; Boschker, H.T.; Bloem, J.; Kardol, P.; van der Putten, W.H.; de Boer, W. Fungal biomass development in a chronosequence of land abandonment. Soil Biol. Biochem. 2006, 38, 51–60. [Google Scholar] [CrossRef]
- Song, M.; Zou, D.-S.; Du, H.; Peng, W.-X.; Zeng, F.-P.; Tan, Q.-J.; Fan, F.-J. Characteristics of soil microbial populations in depressions between karst hills under different land use patterns. J. Appl. Ecol. 2013, 24, 2471–2478. [Google Scholar]
- Potthast, K.; Hamer, U.; Makeschin, F. Land-use change in a tropical mountain rainforest region of southern Ecuador affects soil microorganisms and nutrient cycling. Biogeochemistry 2012, 111, 151–167. [Google Scholar] [CrossRef]
- Kuramae, E.E.; Yergeau, E.; Wong, L.C.; Pijl, A.S.; Van Veen, J.A.; Kowalchuk, G.A. Soil characteristics more strongly influence soil bacterial communities than land-use type. FEMS Microbiol. Ecol. 2011, 79, 12–24. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.A.; Kim, J.M.; Kim, Y.; Joa, J.-H.; Kang, S.-S.; Ahn, J.-H.; Kim, M.; Song, J.; Weon, H.-Y. Different types of agricultural land use drive distinct soil bacterial communities. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef]
- Agnelli, A.; Ascher, J.; Corti, G.; Ceccherini, M.T.; Nannipieri, P.; Pietramellara, G. Distribution of microbial com-munities in a forest soil profile investigated by microbial biomass, soil respiration and DGGE of total and extracellular DNA. Soil Biol. Biochem. 2004, 36, 859–868. [Google Scholar] [CrossRef]
- Fierer, N.; Schimel, J.P.; Holden, P.A. Variations in microbial community composition through two soil depth profiles. Soil Biol. Biochem. 2003, 35, 167–176. [Google Scholar] [CrossRef]
- Gittel, A.; Bárta, J.; Kohoutová, I.; Schnecker, J.; Wild, B.; Capek, P.; Kaiser, C.; Torsvik, V.L.; Richter, A.; Schleper, C.; et al. Site- and horizon-specific patterns of microbial community structure and enzyme activities in permafrost-affected soils of Greenland. Front Microbiol. 2014, 5, 541. [Google Scholar] [CrossRef] [PubMed]
- Will, C.; Thürmer, A.; Wollherr, A.; Nacke, H.; Herold, N.; Schrumpf, M.; Gutknecht, J.; Wubet, T.; Buscot, F.; Daniel, R. Horizon-Specific Bacterial Community Composition of German Grassland Soils, as Revealed by Pyrosequencing-Based Analysis of 16S rRNA Genes. Appl. Environ. Microbiol. 2010, 76, 6751–6759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fierer, N.; Allen, A.S.; Schimel, J.P.; Holden, P.A. Controls on microbial CO2 production: A comparison of surface and subsurface soil horizons. Glob. Chang. Biol. 2003, 9, 1322–1332. [Google Scholar] [CrossRef] [Green Version]
- Hartmann, M.; Lee, S.; Hallam, S.J.; Mohn, W.W. Bacterial, archaeal and eukaryal community structures throughout soil horizons of harvested and naturally disturbed forest stands. Environ. Microbiol. 2009, 11, 3045–3062. [Google Scholar] [CrossRef] [PubMed]
- Eilers, K.G.; Debenport, S.; Anderson, S.; Fierer, N. Digging deeper to find unique microbial communities: The strong effect of depth on the structure of bacterial and archaeal communities in soil. Soil Biol. Biochem. 2012, 50, 58–65. [Google Scholar] [CrossRef]
- Fierer, N.; Grandy, A.S.; Six, J.; Paul, E.A. Searching for unifying principles in soil ecology. Soil Biol. Biochem. 2009, 41, 2249–2256. [Google Scholar] [CrossRef]
- Chen, X.; Su, Y.; He, X.; Wei, Y.; Wei, W.; Wu, J. Soil bacterial community composition and diversity respond to cultivation in Karst ecosystems. World J. Microbiol. Biotechnol. 2011, 28, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Schimel, J.P.; Weintraub, M.N. The implications of exoenzyme activity on microbial carbon and nitrogen limitation in soil: A theoretical model. Soil Biol. Biochem. 2003, 35, 549–563. [Google Scholar] [CrossRef]
- Cleveland, C.C.; Townsend, A.R.; Schmidt, S.K. Phosphorus limitation of microbial processes in moist tropical forests: Evidence from short-term laboratory incubations and field studies. Ecosystems 2002, 5, 0680–0691. [Google Scholar] [CrossRef]
- Liu, L.; Zhang, T.; Gilliam, F.S.; Gundersen, P.; Zhang, W.; Chen, H.; Mo, J. Interactive effects of nitrogen and phosphorus on soil microbial communities in a tropical forest. PLoS ONE 2013, 8, e61188. [Google Scholar]
- Fierer, N.; Chadwick, O.A.; Trumbore, S.E. Production of CO2 in Soil Profiles of a California Annual Grassland. Ecosystems 2005, 8, 412–429. [Google Scholar] [CrossRef] [Green Version]
- Rumpel, C.; Kögel-Knabner, I. Deep soil organic matter—A key but poorly understood component of terrestrial C cycle. Plant Soil 2011, 338, 143–158. [Google Scholar] [CrossRef]
- Madsen, E.L. Impacts of Agricultural Practices on Subsurface Microbial Ecology. Adv. Agron. 1995, 54, 1–67. [Google Scholar] [CrossRef]
- Li, C.; Yan, K.; Tang, L.; Jia, Z.; Li, Y. Change in deep soil microbial communities due to long-term fertilization. Soil Biol. Biochem. 2014, 75, 264–272. [Google Scholar] [CrossRef]
- Faust, K.; Raes, J. Microbial interactions: From networks to models. Nat. Rev. Genet. 2012, 10, 538–550. [Google Scholar] [CrossRef]
- Naveed, M.; Herath, L.; Moldrup, P.; Arthur, E.; Nicolaisen, M.; Norgaard, T.; Ferré, T.P.; de Jonge, L.W. Spatial variability of microbial richness and diversity and relationships with soil organic carbon, texture and structure across an agricultural field. Appl. Soil Ecol. 2016, 103, 44–55. [Google Scholar] [CrossRef]
- Seaton, F.M.; George, P.B.L.; Lebron, I.; Jones, D.L.; Creer, S.; Robinson, D.A. Soil textural heterogeneity impacts bacterial but not fungal diversity. Soil Biol. Biochem. 2020, 144, 107766. [Google Scholar] [CrossRef]
- Liu, J.; Sui, Y.; Yu, Z.; Shi, Y.; Chu, H.; Jin, J.; Liu, X.; Wang, G. High throughput sequencing analysis of biogeographical distribution of bacterial communities in the black soils of northeast China. Soil Biol. Biochem. 2014, 70, 113–122. [Google Scholar] [CrossRef]
- Rousk, J.; Brookes, P.C.; Bååth, E. Contrasting soil pH effects on fungal and bacterial growth suggest functional redundancy in carbon mineralization. Appl. Environ. Microbiol. 2009, 75, 1589–1596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandén, T.; Zavattaro, L.; Spiegel, H.; Grignani, C.; Sandén, H.; Baumgarten, A.; Tiirola, M.; Mikkonen, A. Out of sight—Profiling soil characteristics, nutrients and microbial communities affected by organic amendments down to one meter in a long-term maize cultivation experiment. Appl. Soil Ecol. 2018, 134, 54–63. [Google Scholar] [CrossRef]
- Gu, Y.; Wang, Y.; Lu, S.; Xiang, Q.; Yu, X.; Zhao, K.; Zou, L.; Chen, Q.; Tu, S.; Zhang, X. Long-term fertilization structures bacterial and archaeal communities along soil depth gradient in a paddy soil. Front. Microbiol. 2017, 8, 1516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frouz, J.; Prach, K.; Pižl, V.; Háněl, L.; Stary, J.; Tajovský, K.; Materna, J.; Balík, V.; Kalčík, J.; Řehounková, K. Interactions between soil development, vegetation and soil fauna during spontaneous succession in post mining sites. Eur. J. Soil Biol. 2008, 44, 109–121. [Google Scholar] [CrossRef]
- Yuan, Y.; Zhao, Z.; Niu, S.; Li, X.; Wang, Y.; Bai, Z. Reclamation promotes the succession of the soil and vegetation in opencast coal mine: A case study from Robinia pseudoacacia reclaimed forests, Pingshuo mine, China. Catena 2018, 165, 72–79. [Google Scholar] [CrossRef]
- Nelson, D.W.; Sommers, L.E. Total Carbon, Organic Carbon, and Organic Matter. In Methods of Soil Analysis: Part 2. Chemical and Microbiological Properties; Page, A.L., Ed.; Soil Science Society of America and American Society of Agronomy: Madison, WI, USA, 1982; pp. 539–579. [Google Scholar] [CrossRef]
- Zhao, J.; Zhang, R.; Xue, C.; Xun, W.; Sun, L.; Xu, Y.; Shen, Q. Pyrosequencing reveals contrasting soil bacterial diversity and community structure of two main winter wheat cropping systems in China. Microb. Ecol. 2014, 67, 443–453. [Google Scholar] [CrossRef] [PubMed]
- Tkacz, A.; Hortala, M.; Poole, P.S. Absolute quantitation of microbiota abundance in environmental samples. Microbiome 2018, 6, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Smets, W.; Leff, J.W.; Bradford, M.A.; McCulley, R.L.; Lebeer, S.; Fierer, N. A method for simultaneous measurement of soil bacterial abundances and community composition via 16S rRNA gene sequencing. Soil Biol. Biochem. 2016, 96, 145–151. [Google Scholar]
- Mou, J.; Li, Q.; Shi, W.; Qi, X.; Song, W.; Yang, J. Chain conformation, physicochemical properties of fucosylated chondroitin sulfate from sea cucumber Stichopus chloronotus and its in vitro fermentation by human gut microbiota. Carbohydr. Polym. 2020, 228, 115359. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.Q.; Yu, Y.N.; Gao, R.W.; Wang, H.; Zhang, J.; Li, R.; Long, X.H.; Shen, Q.R.; Chen, W.; Cai, F. High-throughput absolute quantification sequencing reveals the effect of different fertilizer applications on bacterial com-munity in a tomato cultivated coastal saline soil. Sci. Total Environ. 2019, 687, 601–609. [Google Scholar] [CrossRef] [PubMed]
- Cai, F.; Pang, G.; Li, R.-X.; Li, R.; Gu, X.-L.; Shen, Q.-R.; Chen, W. Bioorganic fertilizer maintains a more stable soil microbiome than chemical fertilizer for monocropping. Biol. Fertil. Soils 2017, 53, 861–872. [Google Scholar] [CrossRef]
- Huang, W.; Xian, Z.; Kang, X.; Tang, N.; Li, Z. Genome-wide identification, phylogeny and expression analysis of GRAS gene family in tomato. BMC Plant Biol 2015, 15, 209. [Google Scholar] [CrossRef] [Green Version]
- Tian, Q.; Taniguchi, T.; Shi, W.-Y.; Li, G.; Yamanaka, N.; Du, S. Land-use types and soil chemical properties influence soil microbial communities in the semiarid Loess Plateau region in China. Sci. Rep. 2017, 7, srep45289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, M.; Lee, K.; Scow, K. DNA fingerprinting reveals links among agricultural crops, soil properties, and the composition of soil microbial communities. Geoderma 2003, 114, 279–303. [Google Scholar] [CrossRef]
- Seuradge, B.J.; Maren, O.; Neufeld, J.D. Depth-dependent influence of different land use systems on bacterial biogeography. Microbiol. Ecol. 2017, 93, fiw239. [Google Scholar]
- Fu, X.; Shao, M.; Wei, X.; Horton, R. Soil organic carbon and total nitrogen as affected by vegetation types in Northern Loess Plateau of China. Geoderma 2010, 155, 31–35. [Google Scholar] [CrossRef]
- Yimer, F.; Ledin, S.; Abdelkadir, A. Soil organic carbon and total nitrogen stocks as affected by topographic aspect and vegetation in the Bale Mountains, Ethiopia. Geoderma 2006, 135, 335–344. [Google Scholar] [CrossRef]
- Karlen, D.L.; Mausbach, M.J.; Doran, J.W.; Cline, R.G.; Harris, R.F.; Schuman, G.E. Soil Quality: A Concept, Definition, and Framework for Evaluation (A Guest Editorial). Soil Sci. Soc. Am. J. 1997, 61, 4–10. [Google Scholar] [CrossRef] [Green Version]
- Potthast, K.; Hamer, U.; Makeschin, F. Response of soil microbial activity and community structure to land use changes in a mountain rainforest region of Southern Ecuador. EGU Gen. Assem. Conf. Abstr. 2010, 2010, 11495. [Google Scholar]
- Ge, J.; Wang, S.; Fan, J.; Gongadze, K.; Wu, L. Soil nutrients of different land-use types and topographic positions in the water-wind erosion crisscross region of China’s Loess Plateau. CATENA 2019, 184, 104243. [Google Scholar] [CrossRef]
- Dadi, F.; Endalkachew, K.; Zerihun, K. Rethinking eucalyptus globulus labill. based land use systems in smallholder farmers livelihoods: A case of kolobo watershed, west shewa, Ethiopia. Ekológia 2018, 37, 57–68. [Google Scholar]
- Duguma, L.; Hager, H.; Sieghardt, M. Effects of land use types on soil chemical properties in smallholder farmers of central highland Ethiopia. Ekológia 2010, 29, 1–14. [Google Scholar] [CrossRef]
- Rhoades, C.C.; Coleman, E.D.C. Soil Carbon Differences among Forest, Agriculture, and Secondary Vegetation in Lower Montane Ecuador. Ecol. Appl. 2000, 10, 497–505. [Google Scholar] [CrossRef]
- Gong, J.; Chen, L.; Fu, B.; Huang, Y.; Huang, Z.; Peng, H. Effect of land use on soil nutrients in the loess hilly area of the Loess Plateau, China. Land Degrad. Dev. 2005, 17, 453–465. [Google Scholar] [CrossRef]
- Aerts, R.; Chapin, F.S., III. The mineral nutrition of wild plants revisited: A re-evaluation of processes and patterns. Adv. Ecol. Res. 2000, 30, 1–67. [Google Scholar]
- Chen, X.; Li, B.L. Change in soil carbon and nutrient storage after human disturbance of a primary Korean pine forest in Northeast China. For. Ecol. Manag. 2003, 186, 197–206. [Google Scholar] [CrossRef]
- Lajtha, K.; Schlesinger, W.H. The biogeochemistry of phosphorus cycling and phosphorus availability along a sesert soil chronosequence. Ecology 1998, 69, 24–39. [Google Scholar] [CrossRef]
- Fetene, E.M.; Amera, M.Y. The effects of land use types and soil depth on soil properties of Agedit watershed, Northwest Ethiopia. Ethiop. J. Sci. Technol. 2018, 11, 39. [Google Scholar] [CrossRef] [Green Version]
- Solomon, D.; Lehmann, J.; Mamo, T.; Fritzsche, F.; Zech, W. Phosphorus forms and dynamics as influenced by land use changes in the sub-humid Ethiopian highlands. Geoderma 2002, 105, 21–48. [Google Scholar] [CrossRef]
- Chemeda, M.; Kibret, K.; Fite, T. Influence of different land use types and soil depths on selected soil properties related to soil fertility in Warandhab area, Horo Guduru Wallaga Zone, Oromiya, Ethiopia. Int. J. Environ. Res. 2017, 4, 555634. [Google Scholar]
- Kiflu, A.; Beyene, S. Effects of different land use systems on selected soil properties in South Ethiopia. J. Soil Sci. Environ. Manag. 2013, 4, 100–207. [Google Scholar] [CrossRef]
- Van der Eijk, D.; Janssen, B.H., Oenema. Initial and residual effects of fertilizer phosphorus on soil phosphorus and maize yields on phosphorus fixing soils. A case study in south—West Kenya. Agric. Ecosyst. Environ. 2006, 116, 104–120. [Google Scholar] [CrossRef]
- Zhang, P.; Li, L.; Pan, G.; Ren, J. Soil quality changes in land degradation as indicated by soil chemical, biochemical and microbiological properties in a karst area of southwest Guizhou, China. Environ. Earth Sci. 2006, 51, 609–619. [Google Scholar] [CrossRef]
- Williams, P.W. Environmental change and human impact on karst terrains. Catena Suppl. 1993, 25, 1–19. [Google Scholar]
- Chen, G.; Gan, L.; Wang, S.; Wu, Y.; Wan, G. A comparative study on the microbiological characteristics of soils un-der different land use conditions from karst areas of southwest China. Chin. J. Geochem. 2001, 20, 52–58. [Google Scholar] [CrossRef]
- Iovieno, P.; Morra, L.; Leone, A.; Pagano, L.; Alfani, A. Effect of organic and mineral fertilizers on soil respiration and enzyme activities of two Mediterranean horticultural soils. Biol. Fertil. Soils 2009, 45, 555–561. [Google Scholar] [CrossRef]
- Nayak, D.R.; Babu, Y.J.; Adhya, T. Long-term application of compost influences microbial biomass and enzyme activities in a tropical Aeric Endoaquept planted to rice under flooded condition. Soil Biol. Biochem. 2007, 39, 1897–1906. [Google Scholar] [CrossRef]
- da C Jesus, E.; Marsh, T.L.; Tiedje, J.M.; de S Moreira, F.M. Changes in land use alter the structure of bacterial communities in western amazon soils. Isme J. 2009, 3, 1004–1011. [Google Scholar] [CrossRef]
- Mueller, R.C.; Belnap, J.; Kuske, C.R. Soil bacterial and fungal community responses to nitrogen addition across soil depth and microhabitat in an arid shrubland. Front. Microbiol. 2015, 6, 891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.-S.; Wu, L.-K.; Chen, J.; Khan, M.A.; Luo, X.-M.; Lin, W.-X. Biochemical and microbial properties of rhizospheres under maize/peanut intercropping. J. Integr. Agric. 2016, 15, 101–110. [Google Scholar] [CrossRef] [Green Version]
- Xiao, D.; Xiao, S.; Ye, Y.; Zhang, W.; He, X.; Wang, K. Microbial biomass, metabolic functional diversity, and activity are affected differently by tillage disturbance and maize planting in a typical karst calcareous soil. J. Soils Sediments 2019, 19, 809–821. [Google Scholar] [CrossRef]
- Xiao, S.; Zhang, W.; Ye, Y.; Zhao, J.; Wang, K. Soil aggregate mediates the impacts of land uses on organic carbon, total nitrogen, and microbial activity in a Karst ecosystem. Sci. Rep. 2017, 7, srep41402. [Google Scholar] [CrossRef] [Green Version]
- Lynn, T.M.; Liu, Q.; Hu, Y.; Yuan, H.; Wu, X.; Khai, A.A.; Wu, J.; Ge, T. Influence of land use on bacterial and archaeal diversity and community structures in three natural ecosystems and one agricultural soil. Arch. Microbiol. 2017, 199, 711–721. [Google Scholar]
- Miethling, R.; Wieland, G.; Backhaus, H.; Tebbe, C. Variation of Microbial Rhizosphere Communities in Response to Crop Species, Soil Origin, and Inoculation with Sinorhizobium meliloti L33. Microb. Ecol. 2000, 40, 43–56. [Google Scholar] [CrossRef]
- Lauber, C.L.; Hamady, M.; Knight, R.; Fierer, N. Pyrosequencing-Based Assessment of Soil pH as a Predictor of Soil Bacterial Community Structure at the Continental Scale. Appl. Environ. Microbiol. 2009, 75, 5111–5120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rousk, J.; Bååth, E.; Brookes, P.C.; Lauber, C.L.; Lozupone, C.; Caporaso, J.G.; Knight, R.; Fierer, N. Soil bacterial and fungal communities across a pH gradient in an arable soil. ISME J. 2010, 4, 1340–1351. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.; Cao, J.; Lan, G.; Liang, Y.; Wang, H.; Li, Q. The Influence of Land Use Patterns on Soil Bacterial Community Structure in the Karst Graben Basin of Yunnan Province, China. Forest 2019, 11, 51. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Li, J.; Wang, J.; Liu, G.; Wang, G.; Guo, L.; Peng, S. Decreased temporary turnover of bacterial communities along soil depth gradient during a 35-year grazing exclusion period in a semiarid grassland. Geoderma 2019, 351, 49–58. [Google Scholar] [CrossRef]
- Cheng, J.; Jing, G.; Wei, L.; Jing, Z. Long-term grazing exclusion effects on vegetation characteristics, soil properties and bacterial communities in the semi-arid grasslands of China. Ecol. Eng. 2016, 97, 170–178. [Google Scholar] [CrossRef]
- He, S.; Guo, L.; Niu, M.; Miao, F.; Jiao, S.; Hu, T.; Long, M. Ecological diversity and co-occurrence patterns of bacterial community through soil profile in response to long-term switchgrass cultivation. Sci. Rep. 2017, 7, 3608. [Google Scholar] [CrossRef]
- Jiménez-Bueno, N.; Valenzuela-Encinas, C.; Marsch, R.; Ortiz-Gutiérrez, D.; Verhulst, N.; Govaerts, B.; Dendooven, L.; Na-varro-Noya, Y. Bacterial indicator taxa in soils under different long-term agricultural management. J. Appl. Microbiol. 2016, 120, 921–933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansel, C.M.; Fendorf, S.; Jardine, P.M.; Francis, C.A. Changes in Bacterial and Archaeal Community Structure and Functional Diversity along a Geochemically Variable Soil Profile. Appl. Environ. Microbiol. 2008, 74, 1620–1633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, X.; Liu, C.; Li, J.; Luo, Y.; Yang, Q.; Zhang, W.; Yang, P.; Feng, B. Responses of rhizosphere soil properties, enzyme activities and microbial diversity to intercropping patterns on the Loess Plateau of China. Soil Till. Res. 2019, 195, 104355. [Google Scholar] [CrossRef]
- Liu, C.; Gong, X.; Dang, K.; Li, J.; Yang, P.; Gao, X.; Deng, X.; Feng, B. Linkages between nutrient ratio and the microbial community in rhizosphere soil following fertilizer management. Environ. Res. 2020, 184, 109261. [Google Scholar] [CrossRef] [PubMed]
- Lei, Y.; Xiao, Y.; Li, L.; Jiang, C.; Zu, C.; Li, T.; Cao, H. Impact of tillage practices on soil bacterial diversity and composition under the tobacco-rice rotation in China. J. Microbiol. 2017, 55, 349–356. [Google Scholar] [CrossRef] [PubMed]
- Fierer, N.; Bradford, M.A.; Jackson, R.B. Toward an ecological classification of soil bacteria. Ecology 2007, 88, 1354–1364. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Xiao, X.; Yin, X.; Wang, F. Bacterial community along a historic lake sediment core of Ardley Island, west Antarctica. Extremophiles 2006, 10, 461–467. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Cong, J.; Lu, H.; Li, G.; Qu, Y.; Su, X.; Zhou, J.; Li, D. Community structure and elevational diversity patterns of soil Acidobacteria. J. Enviro. Sci. 2014, 26, 1717–1724. [Google Scholar] [CrossRef]
- Chu, H.; Fierer, N.; Lauber, C.L.; Caporaso, J.G.; Knight, R.; Grogan, P. Soil bacterial diversity in the Arctic is not fundamentally different from that found in other biomes. Environ. Microbiol. 2010, 12, 2998–3006. [Google Scholar] [CrossRef]
- Chen, H.; Zhang, W.; Wang, K.; Fu, W. Soil moisture dynamics under different land uses on karst hillslope in northwest Guangxi, China. Environ. Earth Sci. 2010, 61, 1105–1111. [Google Scholar] [CrossRef]
- Sun, B.; Roberts, D.M.; Dennis, P.G.; Caul, S.; Daniell, T.; Hallett, P.; Hopkins, D.W. Microbial properties and nitrogen contents of arable soils under different tillage regimes. Soil Use Manag. 2013, 30, 152–159. [Google Scholar] [CrossRef]
- Jones, R.T.; Robeson, M.S.; Lauber, C.L.; Hamady, M.; Knight, R.; Fierer, N. A comprehensive survey of soil acido-bacterial diversity using pyrosequencing and clone library analyses. ISME J. 2009, 3, 442–453. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.; Xiong, J.; Zhang, H.; Feng, Y.; Lin, X.; Li, X.; Liang, W.; Chu, H. Soil pH drives the spatial distribution of bacterial communities along elevation on Changbai Mountain. Soil Biol. Biochem. 2013, 57, 204–211. [Google Scholar] [CrossRef]
- Griffiths, B.S.; Philippot, L. Insights into the resistance and resilience of the soil microbial community. FEMS Microbiol. Rev. 2013, 37, 112–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kielak, A.; Rodrigues, J.L.M.; Kuramae, E.E.; Chain, P.S.G.; Van Veen, J.A.; Kowalchuk, G.A. Phylogenetic and metagenomic analysis of Verrucomicrobia in former agricultural grassland soil. FEMS Microbiol. Ecol. 2010, 71, 23–33. [Google Scholar] [CrossRef]
- Wang, P.; Chen, B.; Zhang, H. High throughput sequencing analysis of bacterial communities in soils of a typical Poyang Lake wetland. Acta Ecol. Sin. 2017, 37, 1650–1658. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | df | SOC | TN | TP | NN | AN | AP | pH | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| F | p | F | p | F | p | F | p | F | p | F | p | F | p | ||
| LUT | 4 | 58.7 | <0.0001 | 18.5 | <0.0001 | 3.2 | 0.023 | 13.2 | <0.0001 | 174.5 | <0.0001 | 22.6 | <0.0001 | 90.0 | <0.0001 |
| SD | 1 | 46.3 | <0.0001 | 14.9 | 0.0004 | 5.2 | 0.028 | 14.7 | 0.0004 | 127.4 | <0.0001 | 40.2 | <0.0001 | 76.6 | <0.0001 |
| LUT × SD | 4 | 59.1 | <0.0001 | 1.2 | 0.309 | 1.0 | 0.399 | 18.3 | <0.0001 | 64.4 | <0.0001 | 14.5 | <0.0001 | 75.3 | <0.0001 |
| Treatment | df | Acidobacteria | Actinobacteria | Bacteroidetes | Chloroflexi | Gemmatimonadetes | Planctomycetes | Proteobacteria | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| F | p | F | p | F | p | F | p | F | p | F | p | F | p | ||
| LUT | 4 | 11.1 | <0.0001 | 27.0 | <0.0001 | 5.4 | 0.001 | 16.3 | <0.0001 | 4.3 | 0.006 | 9.8 | <0.0001 | 8.0 | <0.0001 |
| SD | 1 | 16.5 | 0.0002 | 19.3 | <0.0001 | 22.4 | <0.0001 | 17.7 | 0.0001 | 24.8 | <0.0001 | 7.4 | 0.004 | 9.1 | 0.004 |
| LUT × SD | 4 | 0.8 | 0.545 | 8.1 | <0.0001 | 1.8 | 0.144 | 1.6 | 0.200 | 1.3 | 0.294 | 0.8 | 0.539 | 0.9 | 0.461 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Gong, J.; Liu, J.; Hou, W.; Moroenyane, I.; Liu, Y.; Jin, J.; Liu, J.; Xiong, H.; Cheng, C.; et al. Effects of Different Land Use Types and Soil Depth on Soil Nutrients and Soil Bacterial Communities in a Karst Area, Southwest China. Soil Syst. 2022, 6, 20. https://doi.org/10.3390/soilsystems6010020
Li Y, Gong J, Liu J, Hou W, Moroenyane I, Liu Y, Jin J, Liu J, Xiong H, Cheng C, et al. Effects of Different Land Use Types and Soil Depth on Soil Nutrients and Soil Bacterial Communities in a Karst Area, Southwest China. Soil Systems. 2022; 6(1):20. https://doi.org/10.3390/soilsystems6010020
Chicago/Turabian StyleLi, Yuke, Jiyi Gong, Jie Liu, Wenpeng Hou, Itumeleng Moroenyane, Yinglong Liu, Jie Jin, Jie Liu, Han Xiong, Chen Cheng, and et al. 2022. "Effects of Different Land Use Types and Soil Depth on Soil Nutrients and Soil Bacterial Communities in a Karst Area, Southwest China" Soil Systems 6, no. 1: 20. https://doi.org/10.3390/soilsystems6010020
APA StyleLi, Y., Gong, J., Liu, J., Hou, W., Moroenyane, I., Liu, Y., Jin, J., Liu, J., Xiong, H., Cheng, C., Malik, K., Wang, J., & Yi, Y. (2022). Effects of Different Land Use Types and Soil Depth on Soil Nutrients and Soil Bacterial Communities in a Karst Area, Southwest China. Soil Systems, 6(1), 20. https://doi.org/10.3390/soilsystems6010020