A Look into the Past: Tracing Ancient Sustainable Manuring Practices by Thorough P Speciation of Northern European Anthrosols


Abstract
1. Introduction
2. Materials and Methods
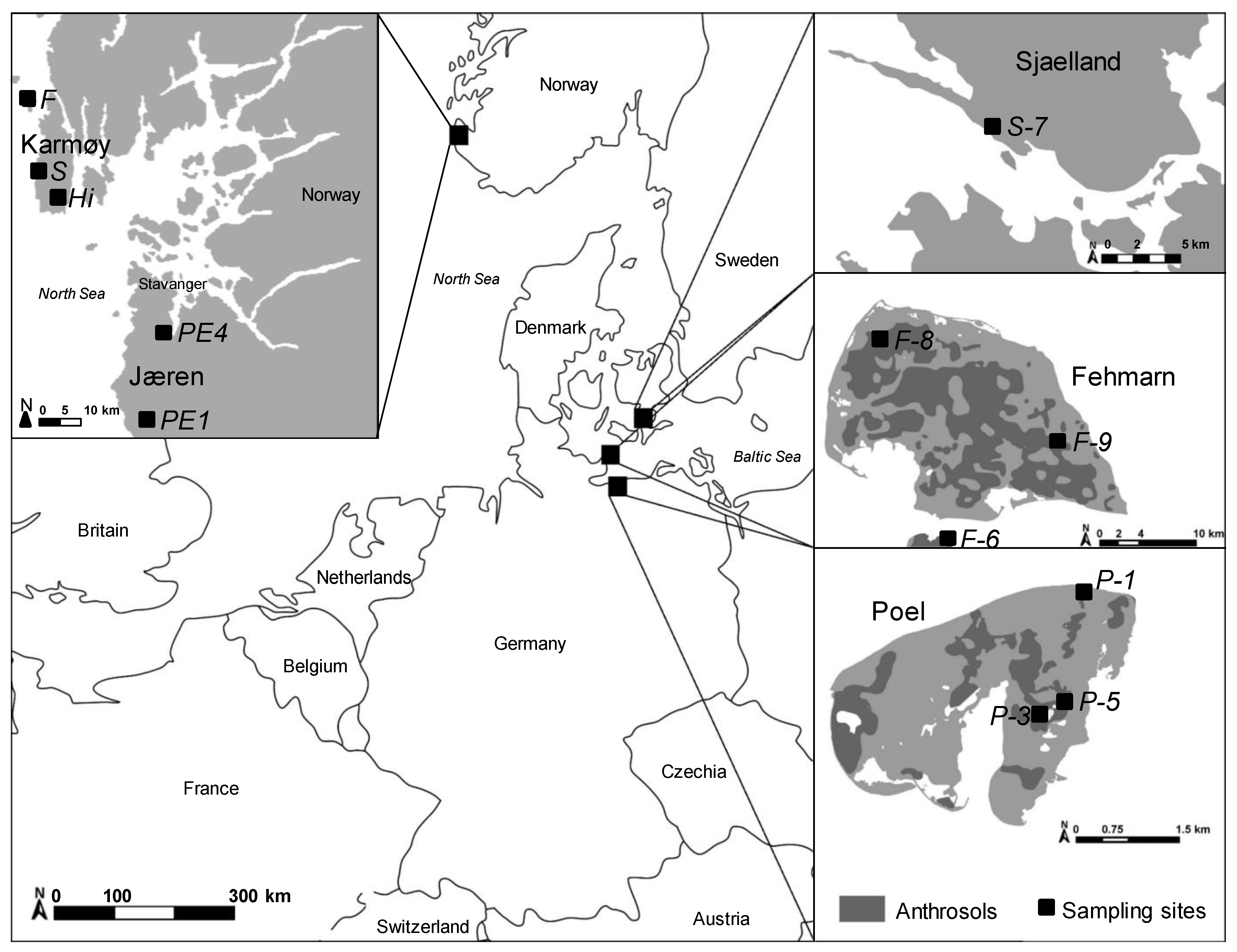
2.1. Study Areas and Soil Sampling
2.2. Sample Pretreatment and Chemical Analyses
2.3. 31P Nuclear Magnetic Resonance Spectroscopy
2.4. P K-Edge X-Ray Absorption Near Edge Structure Spectroscopy
2.5. Data Calculations and Statistical Treatment
3. Results
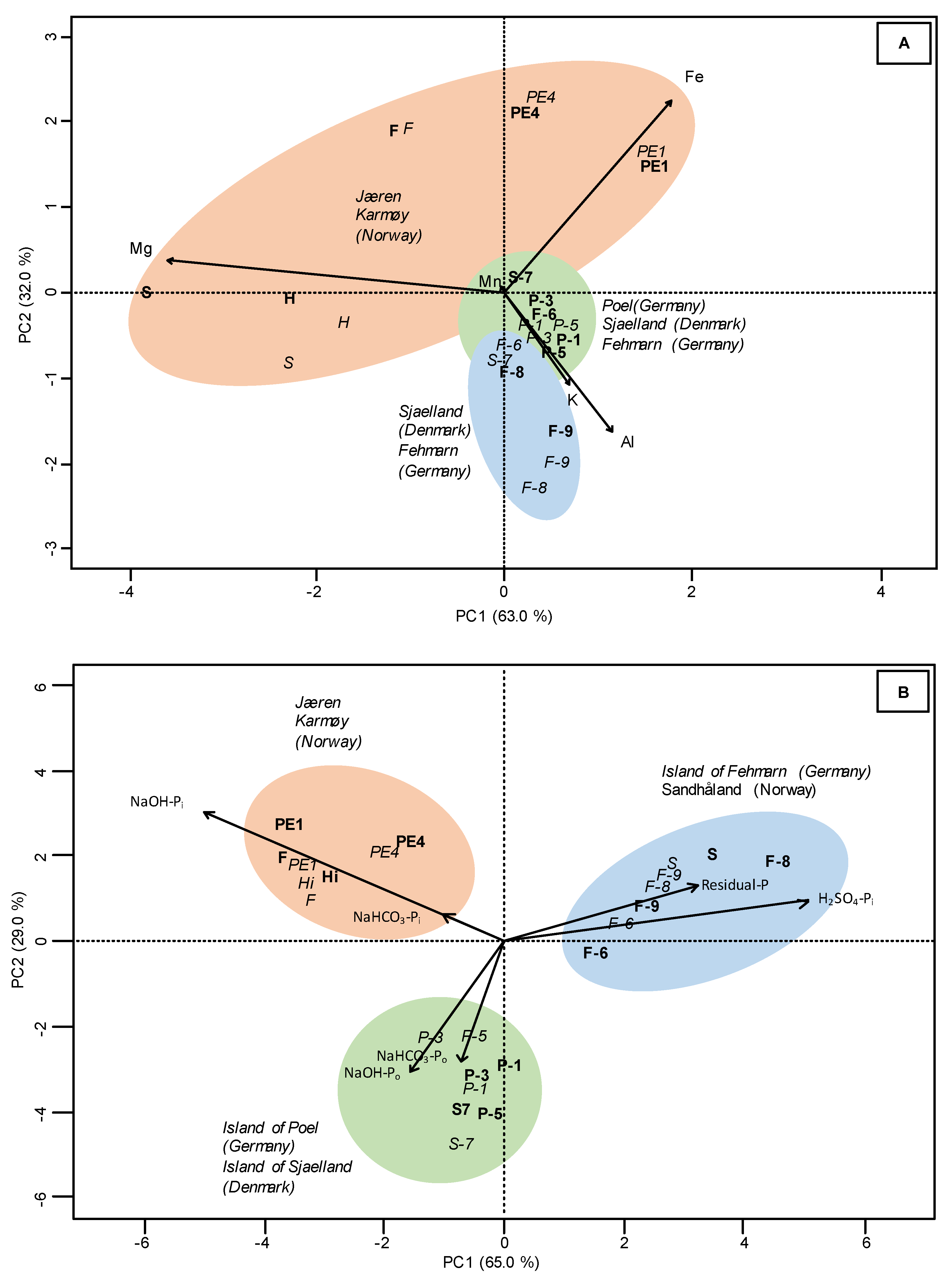
3.1. Total Element Concentrations
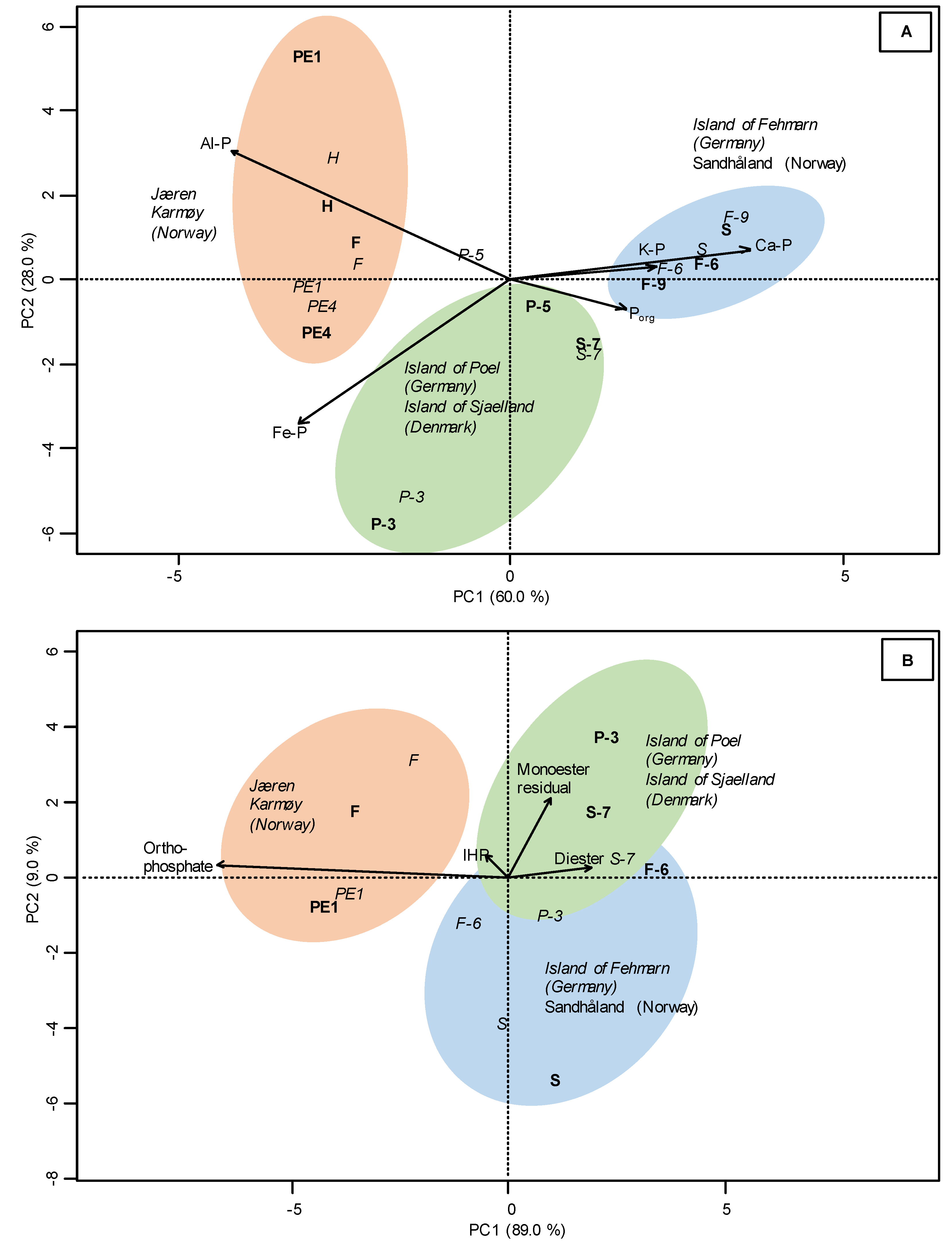
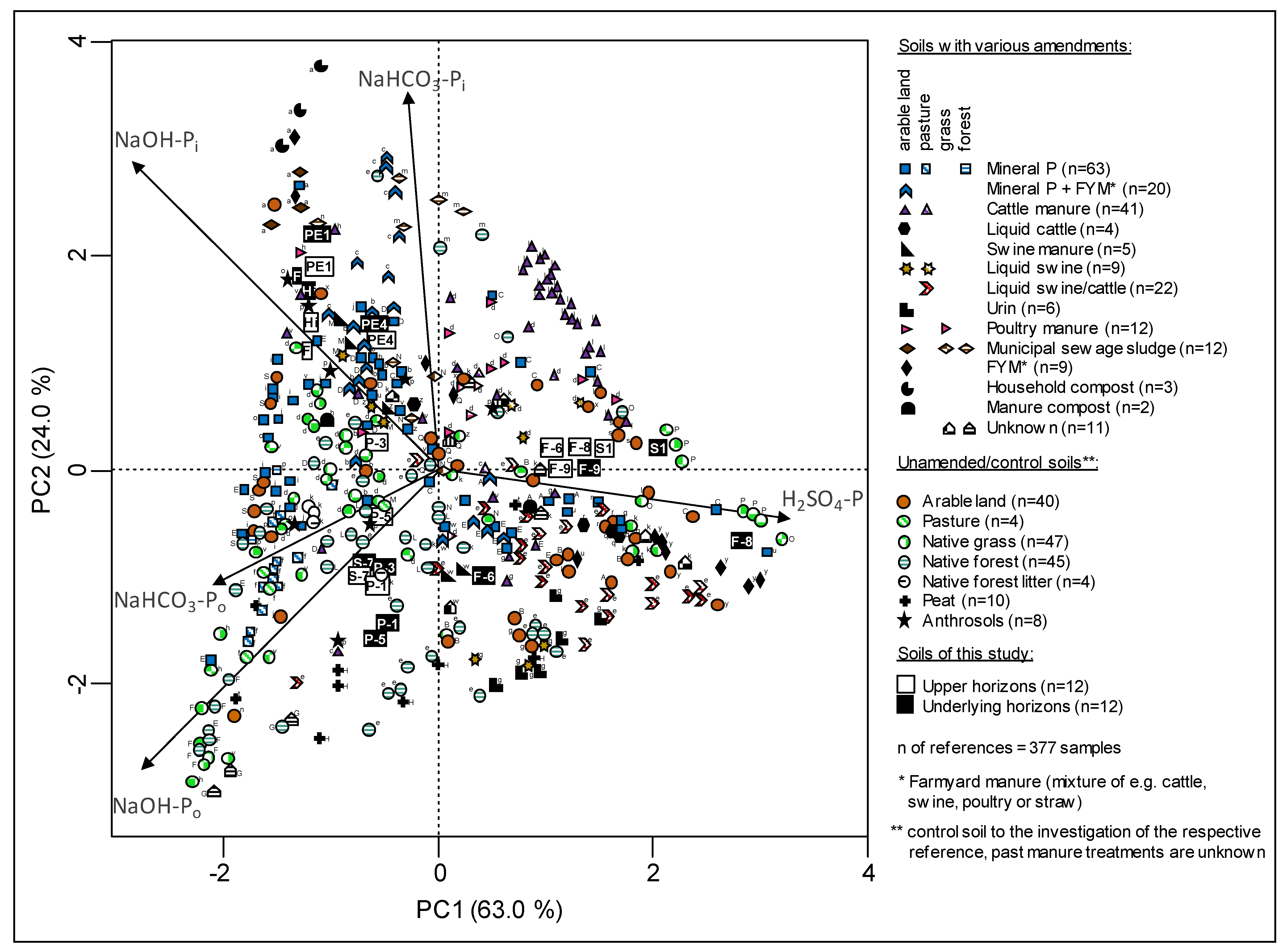
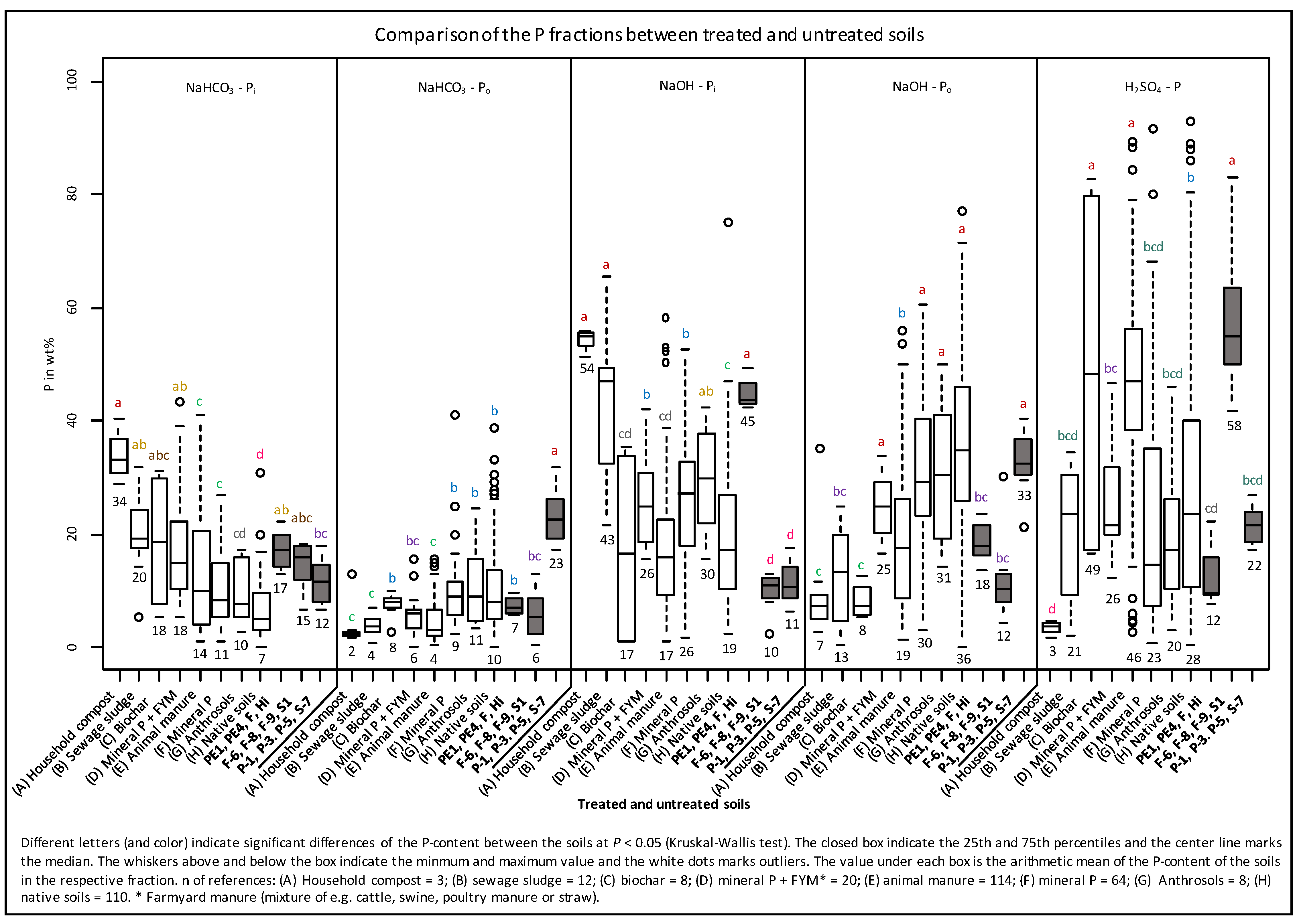
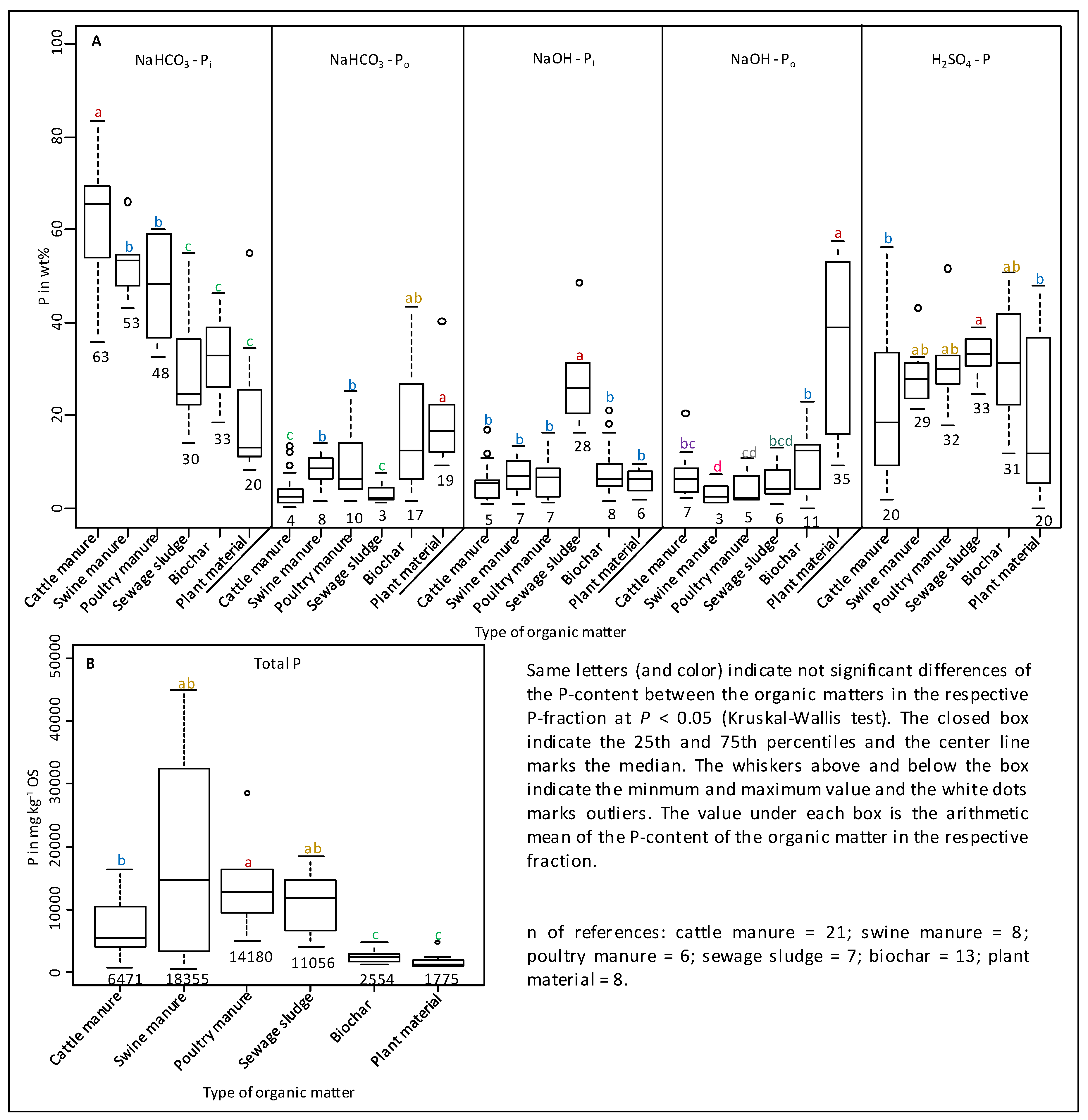
3.2. Sequential P Fractionation
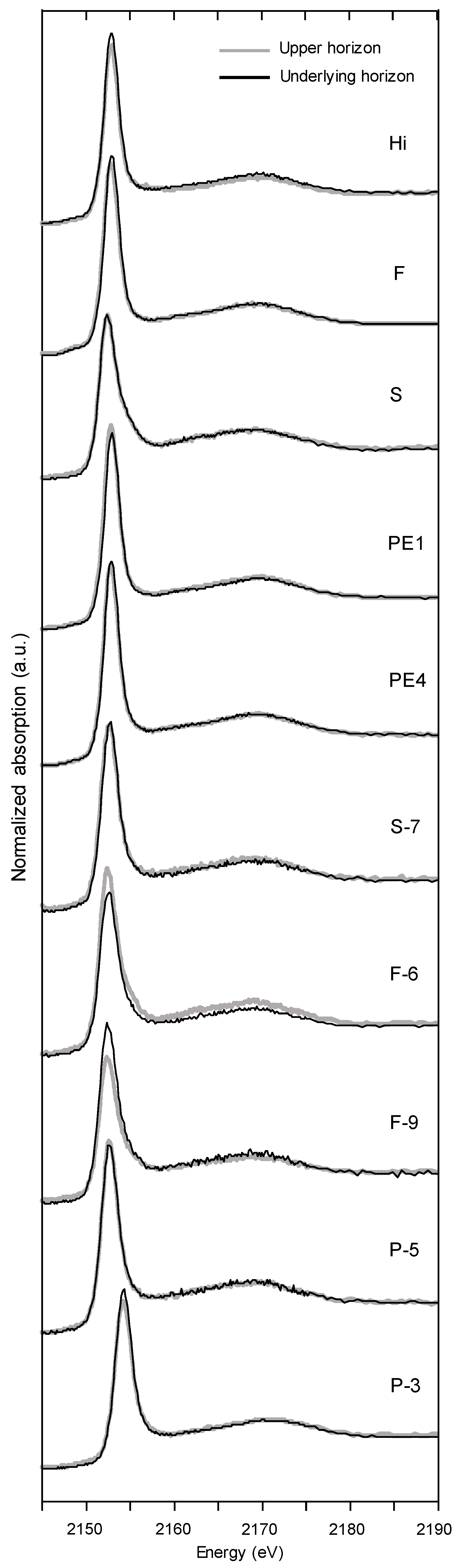
3.3. P K-Edge XANES Spectroscopy
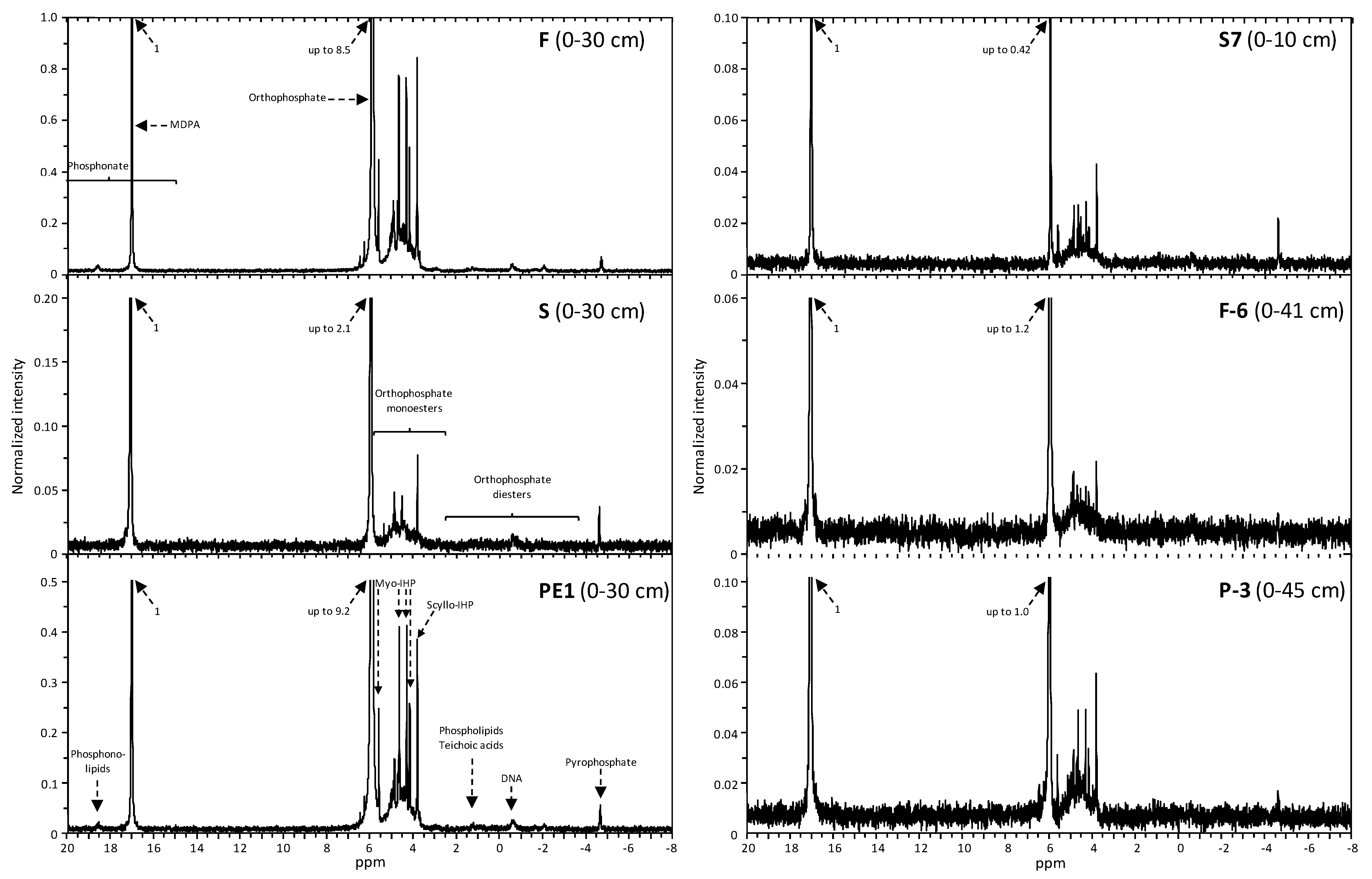
3.4. P Species in the NaOH-EDTA Extract Identified by 31P-NMR Spectroscopy
4. Discussion
4.1. Total P and Other Element Concentrations
4.2. Sequential Phosphorus Fractions
4.3. P Speciation by P K-Edge XANES
4.4. P Speciation by 31P-NMR
4.5. Complementary Archeological Evidence for the Northern European Agricultural History
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Acksel, A.; Amelung, W.; Kühn, P.; Gehrt, E.; Regier, T.; Leinweber, P. Soil organic matter characteristics as indicator of Chernozem genesis in the Baltic Sea region. Geoderma Reg. 2016, 7, 187–200. [Google Scholar] [CrossRef]
- Acksel, A.; Kappenberg, A.; Kühn, P.; Leinweber, P. Human activity formed deep, dark topsoils around the Baltic Sea. Geoderma Reg. 2017, 10, 93–101. [Google Scholar] [CrossRef]
- Acksel, A.; Giani, L.; Stasch, C.; Kühn, P.; Eiter, S.; Potthoff, K.; Regier, T.; Leinweber, P. Humus-rich topsoils in SW Norway—Molecular and isotopic signatures of soil organic matter as indicators for anthropo-pedogenesis. Catena 2019, 172, 831–845. [Google Scholar] [CrossRef]
- Blume, H.P.; Leinweber, P. Plaggen soils: Landscape history, properties, and classification. J. Plant Nutr. Soil Sci. 2004, 167, 319–327. [Google Scholar] [CrossRef]
- Gerlach, R.; Baumewerd-Schmidt, H.; van den Borg, K.; Eckmeier, E.; Schmidt, M.W.I. Prehistoric alteration of soil in the Lower Rhine Basin, Northwest Germany-archaeological, 14C and geochemical evidence. Geoderma 2006, 136, 38–50. [Google Scholar] [CrossRef]
- Gerlach, R.; Fischer, P.; Eckmeier, E.; Hilgers, A. Buried dark soil horizons and archaeological features in the Neolithic settlement region of the Lower Rhine area, NW Germany: Formation, geochemistry and chronostratigraphy. Quat. Int. 2012, 265, 191–204. [Google Scholar] [CrossRef]
- Giani, L.; Makowsky, L.; Mueller, K. Plaggic Anthrosol: Soil of the Year 2013 in Germany: An overview on its formation, distribution, classification, soil function and threats. J. Plant Nutr. Soil Sci. 2014, 177, 320–329. [Google Scholar] [CrossRef]
- Klamm, M. Aufbau und Entstehung eisenzeitlicher Ackerfluren “celtic fields”. Neue Untersuchungen im Gehege Ausselbek, Kr. Schleswig-Flensburg. Archäol. Inf. 1993, 16, 122–124. [Google Scholar]
- Lauer, F.; Pätzold, S.; Gerlach, R.; Protze, J.; Willbold, S.; Amelung, W. Phosphorus status in archaeological arable topsoil relicts-Is it possible to reconstruct conditions for prehistoric agriculture in Germany? Geoderma 2013, 207, 111–120. [Google Scholar] [CrossRef]
- Wiedner, K.; Schneeweiß, J.; Dippold, M.A.; Glaser, B. Anthropogenic dark earth in Northern Germany—The Nordic Analogue to terra preta de Índio in Amazonia. Catena 2015, 132, 114–125. [Google Scholar] [CrossRef]
- Davidson, D.; Dercon, G.; Simpson, I.; Dalsgaard, K.; Spek, T.; Plant, D.A. The identification and significance of inputs to Anthrosols in North-West Europe. Atti Della Soc. Toscana Sci. Nat.-Mem. Ser. A 2007, 112, 79–83. [Google Scholar]
- Dercon, G.; Davidson, D.A.; Dalsgaard, K.; Simpson, I.A.; Spek, T.; Thomas, J. Formation of sandy anthropogenic soils in NW Europe: Identification of inputs based on particle size distribution. Catena 2005, 59, 341–356. [Google Scholar] [CrossRef]
- Domhof, J. Strooiselwinning voor potstallen in verband met de profielopbouw van heide-en oude bouwlandgronden. Boor En Spade 1953, 6, 192–203. [Google Scholar]
- Pape, J.C. Plaggen soils in the Netherlands. Geoderma 1970, 4, 229–255. [Google Scholar] [CrossRef]
- Van Smeerdijk, D.G.; Spek, T.; Kooistra, M.J. Anthropogenic soil formation and agricultural history of the open fields of Valthe (Drenthe, the Netherlands) in mediaeval and early modern times. In Neogene and Quaternary Geology of North-West Europe; Contributions on the Occasion of Waldo, H. Zagwijn’s Retirement; Mededelingen/Rijks Geologische Dienst: Haarlem, The Netherlands, 1995; Volume 52, pp. 451–479. [Google Scholar]
- Spek, M. The Age of Plaggen Soils. In An Evaluation of Dating Methods for Plaggen Soils in The Netherlands and Northern Germany; Food and Agriculture Organization: Rome, Italy, 1992. [Google Scholar]
- Bastiaens, J. Bodemsporen van beddenbouw in het zuidelijk deel van het plaggenlandbouwareaal. Hist. Geogr. Tijdschr. 1994, 12, 8. [Google Scholar]
- Langohr, R. Anthropic impact on the soilscape of the agricultural land in Belgium since early Neolithic: Contribution from archeopedology. In Etude et Gestion des Sols; Food and Agriculture Organization: Rome, Italy, 2001. [Google Scholar]
- Dalsgaard, K.; Karlsen, A.D.; Larsen, L. The farmer and the landscape. In Between Sea and Heather—Landscape and Settlements of the Ulfborg District Until 1700; Dalsgaard, K., Eriksen, P., Jensen, J.V., Rbmer, J.R., Eds.; Aarhus University Press: Aarhus, Denmark, 2000; pp. 87–103. (In Danish) [Google Scholar]
- Gormsen, G. Traditional heathland farming in Western Denmark. Reconstruction of an agricultural system from a peasant diary. Ethnol. Scand. 1991, 21, 105–125. [Google Scholar]
- Nielsen, N.H.; Kristiansen, S.M. Identifing ancient manuring: Traditional phosphate vs. multi-element analysis of archaeological soil. J. Archaeol. Sci. 2014, 42, 390–398. [Google Scholar] [CrossRef]
- Stoklund, B. Turf manuring on the Danish island of Laeso. Geogr. Tidsskr. 1999, 1, 209–214. [Google Scholar]
- Wilson, C.A.; Davidson, D.A.; Cresser, M.S. Multi-element soil analysis: An assessment of its potential as an aid to archaeological interpretation. J. Archaeol. Sci. 2008, 35, 412–424. [Google Scholar] [CrossRef]
- Conry, M.J. Plaggen soils. A review of man-made raised soils. Soils Fertil. 1974, 37, 319–326. [Google Scholar]
- Barber, J.W. Excavations on Iona, 1979. In Proceedings of the Society of Antiquaries of Scotland; National Museum of Antiquities of Scotland: Edinburgh, UK, 1981; Volume 111, pp. 282–380. [Google Scholar]
- Carter, S. A reassessment of the origins of the St Andrews «garden soil». Tayside Fife Archaeol. J. 2001, 7, 87–92. [Google Scholar]
- Davidson, D.A.; Carter, S.P. Micromorphological evidence of past agricultural practices in cultivated soils: The impact of a traditional agricultural system on soils in Papa Stour, Shetland. J. Archaeol. Sci. 1998, 25, 827–838. [Google Scholar] [CrossRef]
- Davidson, D.A.; Simpson, I.A. The formation of deep topsoils in Orkney. Earth Surf. Process. Landf. 1984, 9, 75–81. [Google Scholar] [CrossRef]
- Entwistle, J.A.; Abrahams, P.W. Multi-element analysis of soils and sediments from Scottish historical sites. The potential of inductively coupled plasma-mass spectrometry for rapid site investigation. J. Archaeol. Sci. 1997, 24, 407–416. [Google Scholar] [CrossRef]
- Simpson, I.A. The chronology of anthropogenic soil formation in Orkney. Scott. Geogr. Mag. 1993, 109, 4–11. [Google Scholar] [CrossRef]
- Simpson, I.A. Relict properties of anthropogenic deep top soils as indicators of infield management in Marwick, West Mainland, Orkney. J. Archaeol. Sci. 1997, 24, 365–380. [Google Scholar] [CrossRef]
- Simpson, I.A.; Dockrill, S.J.; Bull, I.D.; Evershed, R.P. Early anthropogenic soil formation at Tofts Ness, Sanday, Orkney. J. Archaeol. Sci. 1998, 25, 729–746. [Google Scholar] [CrossRef]
- Simpson, I.A.; van Bergen, P.F.; Perret, V.; Elhmmali, M.M.; Roberts, D.J.; Evershed, R.P. Lipid biomarkers of manuring practice in relict anthropogenic soils. Holocene 1999, 9, 223–229. [Google Scholar] [CrossRef]
- Giani, L.; Chertov, O.; Gebhardt, C.; Kalinina, O.; Nadporozhskaya, M.; Tolkdorf-Lienemann, E. Plagganthrepts in northwest Russia? Genesis, properties and classification. Geoderma 2004, 121, 113–122. [Google Scholar] [CrossRef]
- Hubbe, A.; Chertov, O.; Kalinina, O.; Nadporozhskaya, M.; Tolksdorf-Lienemann, E.; Giani, L. Evidence of plaggen soils in European North Russia (Arkhangelsk region). J. Plant Nutr. Soil Sci. 2007, 170, 329–334. [Google Scholar] [CrossRef]
- Kalinina, O.; Chertov, O.; Nadporozhskaya, M.; Giani, L. Properties of soil organic matter of Plaggic Anthrosols from Northwest Germany, Northwest and North Russia. Arch. Agron. Soil Sci. 2009, 55, 477–492. [Google Scholar] [CrossRef]
- Glaser, B.; Haumaier, L.; Guggenberger, G.; Zech, W. The ‘Terra Preta’ phenomenon: A model for sustainable agriculture in the humid tropics. Naturwissenschaften 2001, 88, 37–41. [Google Scholar] [CrossRef] [PubMed]
- Liang, B.; Lehmann, J.; Solomon, D.; Kinyangi, J.; Grossman, J.; O’Neill, B.; Skjemstad, J.O.; Thies, J.; Luizão, F.J.; Petersen, J.; et al. Black Carbon increases cation exchange capacity in Soils. Soil Sci. Soc. Am. J. 2006, 70, 1719. [Google Scholar] [CrossRef]
- Solomon, D.; Lehmann, J.; Thies, J.; Schäfer, T.; Liang, B.; Kinyangi, J.; Neves, E.; Petersen, J.; Luizao, F.; Skjemstad, J. Molecular signature and sources of biochemical recalcitrance of organic C in Amazonian Dark Earths. Geochim. Cosmochim. Acta 2007, 71, 2285–2298. [Google Scholar] [CrossRef]
- Holliday, V.T.; Gartner, W.G. Methods of soil P analysis in archaeology. J. Archaeol. Sci. 2007, 34, 301–333. [Google Scholar] [CrossRef]
- Bethell, P.; Máté, I. The Use of Soil Phosphate Analysis in Archaeology: A Critique. I: Scientific Analysis in Archaeology and its Interpretation; Henderson, J., Ed.; Monograph No. 19; Oxford University Committee for Archaeology: Oxford, UK, 1989. [Google Scholar]
- Eidt, R.C. Advances in Abandoned Settlement Analysis: Application to Prehistoric Anthrosols in Colombia, South America; Center for Latin America, University of Wisconsin-Milwaukee Milwaukee: Milwaukee, WI, USA, 1984; ISBN 0-930450-53-1. [Google Scholar]
- Proudfoot, B. The Analysis and Interpretation of Soil Phosphorus in Archaeological Contexts. In Geoarchaeology; Davidson, D.A., Shakley, M.L., Eds.; Duckworth: London, UK, 1976; pp. 93–113. [Google Scholar]
- Provan, D.M.J. Soil phosphate analysis as a tool in archaeology. Nor. Archaeol. Rev. 1971, 4, 37–50. [Google Scholar] [CrossRef]
- Condron, L.M.; Newman, S. Revisiting the fundamentals of phosphorus fractionation of sediments and soils. J. Soils Sediments 2011, 11, 830–840. [Google Scholar] [CrossRef]
- Negassa, W.; Leinweber, P. How does the hedley sequential phosphorus fractionation reflect impacts of land use and management on soil phosphorus: A review. J. Plant Nutr. Soil Sci. 2009, 172, 305–325. [Google Scholar] [CrossRef]
- Hedley, M.J.; Stewart, J.W.B.; Chauhan, B.S. Changes in inorganic and organic soil phosphorus fractions induced by cultivation practices and by laboratory incubations 1. Soil Sci. Soc. Am. J. 1982, 46, 970. [Google Scholar] [CrossRef]
- Tiessen, H.; Moir, J.O. Characterization of available P by sequential extraction. Soil Sampl. Methods Anal. 1993, 7, 5–229. [Google Scholar]
- Cross, A.F.; Schlesinger, W.H. A literature review and evaluation of the. Hedley fractionation: Applications to the biogeochemical cycle of soil phosphorus in natural ecosystems. Geoderma 1995, 64, 197–214. [Google Scholar] [CrossRef]
- Leinweber, P.; Haumaier, L.; Zech, W. Sequential extractions and 31P-NMR spectroscopy of phosphorus forms in animal manures, whole soils and particle-size separates from a densely populated livestock area in northwest Germany. Biol. Fertil. Soils 1997, 25, 89–94. [Google Scholar] [CrossRef]
- Scherer, H.; Sharma, S. Phosphorus fractions and phosphorus delivery potential of a luvisol derived from loess amended with organic materials. Biol. Fertil. Soils 2002, 35, 414–419. [Google Scholar]
- Su, J.; Wang, H.; Kimberley, M.O.; Beecroft, K.; Magesan, G.N.; Hu, C. Fractionation and mobility of phosphorus in a sandy forest soil amended with biosolids. Environ. Sci. Pollut. Res.-Int. 2007, 14, 529–535. [Google Scholar] [CrossRef]
- Qian, P.; Schoenau, J.J.; Wu, T.; Mooleki, P. Phosphorus amounts and distribution in a Saskatchewan soil after five years of swine and cattle manure application. Can. J. Soil Sci. 2004, 84, 275–281. [Google Scholar] [CrossRef]
- Kruse, J.; Abraham, M.; Amelung, W.; Baum, C.; Bol, R.; Kühn, O.; Lewandowski, H.; Niederberger, J.; Oelmann, Y.; Rüger, C. Innovative methods in soil phosphorus research: A review. J. Plant Nutr. Soil Sci. 2015, 178, 43–88. [Google Scholar] [CrossRef]
- Abdala, D.B.; da Silva, I.R.; Vergütz, L.; Sparks, D.L. Long-term manure application effects on phosphorus speciation, kinetics and distribution in highly weathered agricultural soils. Chemosphere 2015, 119, 504–514. [Google Scholar] [CrossRef]
- Ajiboye, B.; Akinremi, O.O.; Racz, G.J. Laboratory characterization of phosphorus in fresh and oven-dried organic amendments. J. Environ. Qual. 2004, 33, 1062–1069. [Google Scholar] [CrossRef]
- Ajiboye, B.; Akinremi, O.O.; Hu, Y.; Jürgensen, A. XANES speciation of phosphorus in organically amended and fertilized Vertisol and Mollisol. Soil Sci. Soc. Am. J. 2008, 72, 1256. [Google Scholar] [CrossRef]
- Beauchemin, S.; Hesterberg, D.; Chou, J.; Beauchemin, M.; Simard, R.R.; Sayers, D.E. Speciation of phosphorus in phosphorus-enriched agricultural soils using X-ray absorption near-edge structure spectroscopy and chemical fractionation. J. Environ. Qual. 2003, 32, 1809. [Google Scholar] [CrossRef] [PubMed]
- Kar, G.; Hundal, L.S.; Schoenau, J.J.; Peak, D. Direct chemical speciation of P in sequential chemical extraction residues using P K-edge X-ray absorption near-edge structure spectroscopy. Soil Sci. 2011, 176, 589–595. [Google Scholar] [CrossRef]
- Khatiwada, R.; Hettiarachchi, G.M.; Mengel, D.B.; Fei, M. Speciation of phosphorus in a fertilized, reduced-till soil system: In-field treatment incubation study. Soil Sci. Soc. Am. J. 2012, 76, 2006. [Google Scholar] [CrossRef]
- Koch, M.; Kruse, J.; Eichler-löbermann, B.; Zimmer, D.; Willbold, S.; Leinweber, P.; Siebers, N. Geoderma Phosphorus stocks and speciation in soil profiles of a long-term fertilizer experiment: Evidence from sequential fractionation, P K-edge XANES, and P NMR spectroscopy. Geoderma 2018, 316, 115–126. [Google Scholar] [CrossRef]
- Liu, J.; Yang, J.; Cade-Menun, B.J.; Liang, X.; Hu, Y.; Liu, C.W.; Zhao, Y.; Li, L.; Shi, J. Complementary phosphorus speciation in agricultural soils by sequential fractionation, solution 31P nuclear magnetic resonance, and phosphorus K-edge X-ray absorption near-edge structure spectroscopy. J. Environ. Qual. 2013, 42, 1763–1770. [Google Scholar] [CrossRef]
- Liu, J.; Hu, Y.; Yang, J.; Abdi, D.; Cade-Menun, B.J. Investigation of soil legacy phosphorus transformation in long-term agricultural fields using sequential fractionation, P K-edge XANES and solution P NMR spectroscopy. Environ. Sci. Technol. 2014, 49, 168–176. [Google Scholar] [CrossRef]
- Liu, J.; Cade-Menun, B.J.; Yang, J.; Hu, Y.; Liu, C.W.; Tremblay, J.; LaForge, K.; Schellenberg, M.; Hamel, C.; Bainard, L.D. Long-term land use affects phosphorus speciation and the composition of phosphorus cycling genes in agricultural soils. Front. Microbiol. 2018, 9, 1–14. [Google Scholar] [CrossRef]
- Lombi, E.; Scheckel, K.G.; Armstrong, R.D.; Forrester, S.; Cutler, J.N.; Paterson, D. Speciation and distribution of phosphorus in a fertilized soil. Soil Sci. Soc. Am. J. 2006, 70, 2038. [Google Scholar] [CrossRef]
- Luo, L.; Ma, Y.; Sanders, R.L.; Xu, C.; Li, J.; Myneni, S.C.B. Phosphorus speciation and transformation in long-term fertilized soil: Evidence from chemical fractionation and P K-edge XANES spectroscopy. Nutr. Cycl. Agroecosyst. 2017, 107, 215–226. [Google Scholar] [CrossRef]
- Peak, D.; Kar, G.; Hundal, L.; Schoenau, J. Kinetics and mechanisms of phosphorus release in a soil amended with biosolids or inorganic fertilizer. Soil Sci. 2012, 177, 183–187. [Google Scholar] [CrossRef]
- Prietzel, J.; Thieme, J.; Paterson, D. Phosphorus speciation of forest-soil organic surface layers using P K-edge XANES spectroscopy. J. Plant Nutr. Soil Sci. 2010, 173, 805–807. [Google Scholar] [CrossRef]
- Sato, S.; Solomon, D.; Hyland, C.; Ketterings, Q.M.; Lehmann, J. Phosphorus speciation in manure and manure-amended soils using XANES spectroscopy. Environ. Sci. Technol. 2005, 39, 7485–7491. [Google Scholar] [CrossRef] [PubMed]
- Toor, G.S.; Peak, J.D.; Sims, J.T. Phosphorus speciation in broiler litter and turkey manure produced from modified diets. J. Environ. Qual. 2005, 34, 687–697. [Google Scholar] [CrossRef] [PubMed]
- Annaheim, K.E.; Doolette, A.L.; Smernik, R.J.; Mayer, J.; Oberson, A.; Frossard, E.; Bünemann, E.K. Long-term addition of organic fertilizers has little effect on soil organic phosphorus as characterized by 31P NMR spectroscopy and enzyme additions. Geoderma 2015, 257, 67–77. [Google Scholar] [CrossRef]
- Doolette, A.L.; Smernik, R.J.; Dougherty, W.J. Spiking Improved Solution Phosphorus-31 Nuclear Magnetic Resonance Identification of Soil Phosphorus Compounds. Soil Sci. Soc. Am. J. 2009, 73, 919. [Google Scholar] [CrossRef]
- Dou, Z.; Ramberg, C.F.; Toth, J.D.; Wang, Y.; Sharpley, A.N.; Boyd, S.E.; Chen, C.R.; Williams, D.; Xu, Z.H. Phosphorus speciation and sorption-desorption characteristics in heavily manured soils. Soil Sci. Soc. Am. J. 2009, 73, 93. [Google Scholar] [CrossRef]
- Hansen, J.C.; Cade-Menun, B.J.; Strawn, D.G. Phosphorus speciation in manure-amended alkaline soils. J. Environ. Qual. 2004, 33, 1521–1527. [Google Scholar] [CrossRef]
- Hawkes, G.E.; Powlson, D.S.; Randall, E.W.; Tate, K.R. A 31P nuclear magnetic resonance study of the phosphorus species in alkali extracts of soils from long-term field experiments. J. Soil Sci. 1984, 35, 35–45. [Google Scholar] [CrossRef]
- Koopmans, G.F.; Chardon, W.J.; Dolfing, J.; Oenema, O.; Van der Meer, P.; Van Riemsdijk, W.H. Wet chemical and phosphorus-31 nuclear magnetic resonance analysis of phosphorus speciation in a sandy soil receiving long-term fertilizer or animal manure applications. J. Environ. Qual. 2003, 32, 287–295. [Google Scholar] [CrossRef]
- Koopmans, G.F.; Chardon, W.J.; McDowell, R.W. Phosphorus movement and speciation in a sandy soil profile after long-term animal manure applications. J. Environ. Qual. 2007, 36, 305. [Google Scholar] [CrossRef]
- Li, M.; Zhang, J.; Wang, G.; Yang, H.; Whelan, M.J.; White, S.M. Organic phosphorus fractionation in wetland soil profiles by chemical extraction and phosphorus-31 nuclear magnetic resonance spectroscopy. Appl. Geochem. 2013, 33, 213–221. [Google Scholar] [CrossRef]
- McDowell, R.W.; Stewart, I. Phosphorus in fresh and dry dung of grazing dairy cattle, deer, and sheep. J. Environ. Qual. 2005, 34, 598–607. [Google Scholar] [CrossRef] [PubMed]
- McDowell, R.W.; Condron, L.M.; Stewart, I.; Cave, V. Chemical nature and diversity of phosphorus in New Zealand pasture soils using 31P nuclear magnetic resonance spectroscopy and sequential fractionation. Nutr. Cycl. Agroecosyst. 2005, 72, 241–254. [Google Scholar] [CrossRef]
- Shafqat, M.N.; Pierzynski, G.M.; Xia, K. Phosphorus source effects on soil organic phosphorus: A 31P NMR study. Commun. Soil Sci. Plant Anal. 2009, 40, 1722–1746. [Google Scholar] [CrossRef]
- Guggenberger, G.; Christensen, B.T.; Rubæk, G.; Zech, W. Land-use and fertilization effects on P forms in two European soils: Resin extraction and 31P-NMR analysis. Eur. J. Soil Sci. 1996, 47, 605–614. [Google Scholar] [CrossRef]
- Solomon, D.; Lehmann, J. Loss of phosphorus from soil in semi-arid northern Tanzania as a result of cropping: Evidence from sequential extraction and 31P-NMR spectroscopy. Eur. J. Soil Sci. 2000, 51, 699–708. [Google Scholar] [CrossRef]
- Turrion, M.B.; Lafuente, F.; Aroca, M.J.; López, O.; Mulas, R.; Ruipérez, C. Characterization of soil phosphorus in a fire-affected forest Cambisol by chemical extractions and 31P-NMR spectroscopy analysis. Sci. Total Environ. 2010, 408, 3342–3348. [Google Scholar] [CrossRef]
- Murphy, J.; Riley, J.P. A modified single solution method for the determination of phosphate in natural waters. Anal. Chim. Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Acksel, A.; Baumann, K.; Hu, Y.; Leinweber, P. A critical review and evaluation of some P-research methods. Submitt. Commun. Soil. Sci. Plant Anal. 2019, 9. [Google Scholar] [CrossRef]
- Baumann, K.; Glaser, K.; Mutz, J.-E.; Karsten, U.; MacLennan, A.; Hu, Y.; Michalik, D.; Kruse, J.; Eckhardt, K.-U.; Schall, P. Biological soil crusts of temperate forests: Their role in P cycling. Soil Biol. Biochem. 2017, 109, 156–166. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. 2013. Available online: https://cran.biodisk.org/web/packages/dplR/vignettes/intro-dplR.pdf (accessed on 29 October 2019).
- Schnepel, C.; Potthoff, K.; Eiter, S.; Giani, L. Evidence of plaggen soils in SW Norway. J. Plant Nutr. Soil Sci. 2014, 177, 638–645. [Google Scholar] [CrossRef]
- Kidd, P.S.; Domínguez-Rodríguez, M.J.; Díez, J.; Monterroso, C. Bioavailability and plant accumulation of heavy metals and phosphorus in agricultural soils amended by long-term application of sewage sludge. Chemosphere 2007, 66, 1458–1467. [Google Scholar] [CrossRef] [PubMed]
- Sui, Y.; Thompson, M.L.; Shang, C. Fractionation of phosphorus in a Mollisol amended with biosolids. Soil Sci. Soc. Am. J. 1999, 63, 1174–1180. [Google Scholar] [CrossRef]
- Xu, G.; Shao, H.; Zhang, Y.; Junna, S. Nonadditive effects of biochar amendments on soil phosphorus fractions in two contrasting soils. Land Degrad. Dev. 2018, 29, 2720–2727. [Google Scholar] [CrossRef]
- Agbenin, J.O.; Goladi, J.T. Dynamics of phosphorus fractions in a savanna Alfisol under continuous cultivation. Soil Use Manag. 1998, 14, 59–64. [Google Scholar] [CrossRef]
- Keller, M.; Oberson, A.; Annaheim, K.E.; Tamburini, F.; Mäder, P.; Mayer, J.; Frossard, E.; Bünemann, E.K. Phosphorus forms and enzymatic hydrolyzability of organic phosphorus in soils after 30 years of organic and conventional farming. J. Plant Nutr. Soil Sci. 2012, 175, 385–393. [Google Scholar] [CrossRef]
- Lehmann, J.; Lan, Z.; Hyland, C.; Sato, S.; Solomon, D.; Ketterings, Q.M. Long-term dynamics of phosphorus forms and retention in manure-amended soils. Environ. Sci. Technol. 2005, 39, 6672–6680. [Google Scholar] [CrossRef]
- Mao, X.; Xu, X.; Lu, K.; Gielen, G.; Luo, J.; He, L.; Donnison, A.; Xu, Z.; Xu, J.; Yang, W. Effect of 17 years of organic and inorganic fertilizer applications on soil phosphorus dynamics in a rice–wheat rotation cropping system in eastern China. J. Soils Sedim. 2015, 15, 1889–1899. [Google Scholar] [CrossRef]
- Reddy, D.D.; Rao, A.S.; Takkar, P.N. Effects of repeated manure and fertilizer phosphorus additions on soil phosphorus dynamics under a soybean-wheat rotation. Biol. Fertil. Soils 1999, 28, 150–155. [Google Scholar] [CrossRef]
- Tran, T.S.; N’dayegamiye, A. Long-term effects of fertilizers and manure application on the forms and availability of soil phosphorus. Can. J. Soil Sci. 1995, 75, 281–285. [Google Scholar] [CrossRef]
- Campbell, C.A.; Biederbeck, V.O.; Selles, F.; Schnitzer, M.; Stewart, J.W.B. Effect of manure and P fertilizer on properties of a Black Chernozem in southern Saskatchewan. Can. J. Soil Sci. 1986, 66, 601–614. [Google Scholar] [CrossRef]
- Couto, R.d.R.; Ferreira, P.A.A.; Ceretta, C.A.; Lourenzi, C.R.; Facco, D.B.; Tassinari, A.; Piccin, R.; Conti, L.D.; Gatiboni, L.C.; Schapanski, D. Phosphorus fractions in soil with a long history of organic waste and mineral fertilizer addition. Bragantia 2017, 76, 155–166. [Google Scholar] [CrossRef]
- Guardini, R.; Comin, J.J.; Schmitt, D.E.; Tiecher, T.; Bender, M.A.; dos Santos, D.R.; Mezzari, C.P.; Oliveira, B.S.; Gatiboni, L.C.; Brunetto, G. Accumulation of phosphorus fractions in typic Hapludalf soil after long-term application of pig slurry and deep pig litter in a no-tillage system. Nutr. Cycl. Agroecosyst. 2012, 93, 215–225. [Google Scholar] [CrossRef]
- Hao, X.; Godlinski, F.; Chang, C. Distribution of phosphorus forms in soil following long-term continuous and discontinuous cattle manure applications. Soil Sci. Soc. Am. J. 2008, 72, 90–97. [Google Scholar] [CrossRef]
- López-Contreras, A.Y.; Hernández-Valencia, I.; López-Hernández, D. Fractionation of soil phosphorus in organic amended farms located on savanna sandy soils of Venezuelan Amazonian. Biol. Fertil. Soils 2007, 43, 771–777. [Google Scholar] [CrossRef]
- Motavalli, P.P.; Miles, R.J. Soil phosphorus fractions after 111 years of animal manure and fertilizer applications. Biol. Fertil. Soils 2002, 36, 35–42. [Google Scholar]
- Sharpley, A.N.; McDowell, R.W.; Kleinman, P.J.A. Amounts, forms, and solubility of phosphorus in soils receiving manure. Soil Sci. Soc. Am. J. 2004, 68, 2048–2057. [Google Scholar] [CrossRef]
- Zheng, Z.; Simard, R.R.; Lafond, J.; Parent, L.E. Pathways of soil phosphorus transformations after 8 years of cultivation under contrasting cropping practices. Soil Sci. Soc. Am. J. 2002, 66, 999–1007. [Google Scholar] [CrossRef]
- Zheng, Z.; MacLeod, J.A.; Sanderson, J.B.; Lafond, J. Soil phosphorus dynamics after ten annual applications of mineral fertilizers and liquid dairy manure: Fractionation and path analyses. Soil Sci. 2004, 169, 449–456. [Google Scholar] [CrossRef]
- Condron, L.M.; Goh, K.M. Effects of long-term phosphatic fertilizer applications on amounts and forms of phosphorus in soils under irrigated pasture in New Zealand. J. Soil Sci. 1989, 40, 383–395. [Google Scholar] [CrossRef]
- Lehmann, J.; Campos, C.V.; de Macêdo, J.L.V.; German, L. Sequential P fractionation of relict anthropogenic dark earths of Amazonia. Amaz. Dark Earths Explor. Space Time 2004, 26416, 113. [Google Scholar]
- Magid, J. Vegetation effects on phosphorus fractions in set-aside soils. Plant Soil 1993, 149, 111–119. [Google Scholar] [CrossRef]
- Richards, J.E.; Bates, T.E.; Sheppard, S.C. Changes in the forms and distribution of soil phosphorus due to long-term corn production. Can. J. Soil Sci. 1995, 75, 311–318. [Google Scholar] [CrossRef]
- Verma, S.; Subehia, S.K.; Sharma, S.P. Phosphorus fractions in an acid soil continuously fertilized with mineral and organic fertilizers. Biol. Fertil. Soils 2005, 41, 295–300. [Google Scholar] [CrossRef]
- Agbenin, J.O.; Tiessen, H. Phosphorus transformations in a toposequence of lithosols and cambisols from semi-arid northeastern Brazil. Geoderma 1994, 62, 345–362. [Google Scholar] [CrossRef]
- Alt, F.; Oelmann, Y.; Herold, N.; Schrumpf, M.; Wilcke, W. Phosphorus partitioning in grassland and forest soils of Germany as related to land-use type, management intensity, and land use–related pH. J. Plant Nutr. Soil Sci. 2011, 174, 195–209. [Google Scholar] [CrossRef]
- Araújo, M.S.B.; Schaefer, C.E.R.; Sampaio, E.V.S.B. Soil phosphorus fractions from toposequences of semi-arid Latosols and Luvisols in northeastern Brazil. Geoderma 2004, 119, 309–321. [Google Scholar] [CrossRef]
- Chen, C.R.; Condron, L.M.; Davis, M.R.; Sherlock, R.R. Effects of afforestation on phosphorus dynamics and biological properties in a New Zealand grassland soil. Plant Soil 2000, 220, 151–163. [Google Scholar] [CrossRef]
- Cross, A.F.; Schlesinger, W.H. Biological and geochemical controls on phosphorus fractions in semiarid soils. Biogeochemistry 2001, 52, 155–172. [Google Scholar] [CrossRef]
- Derry, D.D.; Voroney, R.P.; Briceño, J.A. Long-term effects of short-fallow frijol tapado on soil phosphorus pools in Costa Rica. Agric. Ecosyst. Environ. 2005, 110, 91–103. [Google Scholar] [CrossRef]
- Schlichting, A.; Leinweber, P.; Meissner, R.; Altermann, M. Sequentially extracted phosphorus fractions in peat-derived soils. J. Plant Nutr. Soil Sci. 2002, 165, 290–298. [Google Scholar] [CrossRef]
- Schoenau, J.J.; Stewart, J.W.B.; Bettany, J.R. Forms and cycling of phosphorus in prairie and boreal forest soils. Biogeochemistry 1989, 8, 223–237. [Google Scholar] [CrossRef]
- Tchienkoua, M.; Zech, W. Chemical and spectral characterization of soil phosphorus under three land uses from an Andic Palehumult in West Cameroon. Agric. Ecosyst. Environ. 2003, 100, 193–200. [Google Scholar] [CrossRef]
- Tiessen, H.; Salcedo, I.H.; Sampaio, E.V.S.B. Nutrient and soil organic matter dynamics under shifting cultivation in semi-arid northeastern Brazil. Agric. Ecosyst. Environ. 1992, 38, 139–151. [Google Scholar] [CrossRef]
- Tiessen, H.; Stewart, J.W.B.; Moir, J.O. Changes in organic and inorganic phosphorus composition of two grassland soils and their particle size fractions during 60–90 years of cultivation. J. Soil Sci. 1983, 34, 815–823. [Google Scholar] [CrossRef]
- Malik, M.A.; Khan, K.S.; Marschner, P.; Ali, S. Organic amendments differ in their effect on microbial biomass and activity and on P pools in alkaline soils. Biol. Fertil. Soils 2013, 49, 415–425. [Google Scholar] [CrossRef]
- Edmeades, D.C. The long-term effects of manures and fertilisers on soil productivity and quality: A review. Nutr. Cycl. Agroecosyst. 2003, 66, 165–180. [Google Scholar] [CrossRef]
- Myhre, B. Arable fields and farm structure. Archaeol. Environ. 1985, 4, 69–82. [Google Scholar]
- Rose, C.; Parker, A.; Jefferson, B.; Cartmell, E. The characterization of feces and urine: A review of the literature to inform advanced treatment technology. Crit. Rev. Environ. Sci. Technol. 2015, 45, 1827–1879. [Google Scholar] [CrossRef]
- Faridullah, F.; Khalid, Z.; Irshad, M.; Alam, A.; Ahmed, T.; Bhatti, Z. Fractionation of phosphorus in human and animal wastes. Minerva Biotecnol. 2015, 27, 63–70. [Google Scholar]
- Feachem, R.G. Health Aspects of Excreta and Wastewater Management; review and analysis, part 1; World Bank: Washington, DC, USA, 1978. [Google Scholar]
- Strauss, M. Health Aspects of Nightsoil and Sludge Use in Agriculture and Aquaculture. Part II: Pathogen Survival; International Reference Centre for Waste Disposal: Duebendorf, Switzerland, 1985. [Google Scholar]
- Li, G.; Li, H.; Leffelaar, P.A.; Shen, J.; Zhang, F. Characterization of phosphorus in animal manures collected from three (dairy, swine, and broiler) farms in China. PLoS ONE 2014, 9, e102698. [Google Scholar] [CrossRef] [PubMed]
- Gagnon, B.; Demers, I.; Ziadi, N.; Chantigny, M.H.; Parent, L.-E.; Forge, T.A.; Larney, F.J.; Buckley, K.E. Forms of phosphorus in composts and in compost-amended soils following incubation. Can. J. Soil Sci. 2012, 92, 711–721. [Google Scholar] [CrossRef]
- Gagnon, B.; Simard, R.R. Soil P fractions as affected by on-farm composts in a controlled incubation study. Can. J. Soil Sci. 2003, 83, 223–226. [Google Scholar] [CrossRef]
- Larrea-Marín, M.T.; Pomares-Alfonso, M.S.; Gómez-Juaristi, M.; Sánchez-Muniz, F.J.; de la Rocha, S.R. Validation of an ICP-OES method for macro and trace element determination in Laminaria and Porphyra seaweeds from four different countries. J. Food Compos. Anal. 2010, 23, 814–820. [Google Scholar] [CrossRef]
- Mæhre, H.K.; Malde, M.K.; Eilertsen, K.E.; Elvevoll, E.O. Characterization of protein, lipid and mineral contents in common Norwegian seaweeds and evaluation of their potential as food and feed. J. Sci. Food Agric. 2014, 94, 3281–3290. [Google Scholar] [CrossRef] [PubMed]
- Biancarosa, I.; Belghit, I.; Bruckner, C.G.; Liland, N.S.; Waagbø, R.; Amlund, H.; Heesch, S.; Lock, E.J. Chemical characterization of 21 species of marine macroalgae common in Norwegian waters: Benefits of and limitations to their potential use in food and feed. J. Sci. Food Agric. 2018, 98, 2035–2042. [Google Scholar] [CrossRef]
- Huth, V.; Günther, A.; Augustin, J.; Borraz, E.; Ellerbrock, R.; Giebels, M.; Hierold, W.; Hoffmann, M.; Jurasinski, G.; Rosskopf, N.; et al. Treibhausgas-Emissionen aus wiedervernässten Niedermooren Nordostdeutschlands. In Proceedings of the DBG Mitteilungen Band 116-2013-Exkursionsführer Rostock, Rostock, Germany, 7–12 September 2013; Volume 116, p. 180, ISBN 0343-1071. [Google Scholar]
- Lehmann, J.; Pereira, J.; Steiner, C.; Nehls, T.; Zech, W.; Glaser, B. Nutrient availability and leaching in an archaeological Anthrosol and a Ferralsol of the Central Amazon basin: Fertilizer, manure and charcoal amendments. Plant Soil 2003, 249, 343–357. [Google Scholar] [CrossRef]
- Schmidt, M.J.; Py-daniel, A.R.; Paula, C.D.; Valle, R.B.M.; Caromano, C.F.; Texeira, W.G.; Barbosa, C.A.; Fonseca, J.A.; Magalhães, M.P.; Silva, D.; et al. Dark earths and the human built landscape in Amazonia: A widespread pattern of anthrosol formation. J. Archaeol. Sci. 2014, 42, 152–165. [Google Scholar] [CrossRef]
- Reimann, C.; Birke, M.; Demetriades, A.; Filzmoser, P.; O’Connor, P. Chemistry of Europe’s Agricultural Soils, Part A; Schweitzerbart: Stuttgart, Germany, 2014. [Google Scholar]
- Barej, J.A.M.; Pätzold, S.; Perkons, U.; Amelung, W. Phosphorus fractions in bulk subsoil and its biopore systems. Eur. J. Soil Sci. 2014, 65, 553–561. [Google Scholar] [CrossRef]
- Chang, A.C.; Page, A.L.; Sutherland, F.H.; Grgurevic, E. Fractionation of Phosphorus in Sludge-Affected Soils 1. J. Environ. Qual. 1983, 12, 286–290. [Google Scholar] [CrossRef]
- Qian, P.; Schoenau, J.J. Fractionation of P in soil as influenced by a single addition of liquid swine manure. Can. J. Soil Sci. 2000, 80, 561–566. [Google Scholar] [CrossRef]
- van Zwieten, L.; Kimber, S.; Morris, S.; Chan, K.Y.; Downie, A.; Rust, J.; Joseph, S.; Cowie, A. Effects of biochar from slow pyrolysis of papermill waste on agronomic performance and soil fertility. Plant Soil 2010, 327, 235–246. [Google Scholar] [CrossRef]
- Peng, X.; Ye, L.L.; Wang, C.H.; Zhou, H.; Sun, B. Temperature- and duration-dependent rice straw-derived biochar: Characteristics and its effects on soil properties of an Ultisol in southern China. Soil Tillage Res. 2011, 112, 159–166. [Google Scholar] [CrossRef]
- Frossard, E.; Tekely, P.; Grimal, J.Y. Characterization of phosphate species in urban sewage sludges by high-resolution solid-state 31P NMR. Eur. J. Soil Sci. 1994, 45, 403–408. [Google Scholar] [CrossRef]
- Khan, K.S.; Joergensen, R.G. Changes in microbial biomass and P fractions in biogenic household waste compost amended with inorganic P fertilizers. Bioresour. Technol. 2009, 100, 303–309. [Google Scholar] [CrossRef] [PubMed]
- Ngo, P.-T.; Rumpel, C.; Ngo, Q.-A.; Alexis, M.; Vargas, G.V.; de la Luz Mora Gil, M.; Dang, D.-K.; Jouquet, P. Biological and chemical reactivity and phosphorus forms of buffalo manure compost, vermicompost and their mixture with biochar. Bioresour. Technol. 2013, 148, 401–407. [Google Scholar] [CrossRef]
- Sharpley, A.; Moyer, B. Phosphorus forms in manure and compost and their release during simulated rainfall. J. Environ. Qual. 2000, 29, 1462–1469. [Google Scholar] [CrossRef]
- Xu, G.; Zhang, Y.; Shao, H.; Sun, J. Pyrolysis temperature affects phosphorus transformation in biochar: Chemical fractionation and 31P NMR analysis. Sci. Total Environ. 2016, 569, 65–72. [Google Scholar] [CrossRef]
- Zvomuya, F.; Helgason, B.L.; Larney, F.J.; Janzen, H.H.; Akinremi, O.O.; Olson, B.M. Predicting phosphorus availability from soil-applied composted and non-composted cattle feedlot manure. J. Environ. Qual. 2006, 35, 928. [Google Scholar] [CrossRef]
- Hinedi, Z.R.; Chang, A.C.; Lee, R.W.K. Mineralization of phosphorus in sludge-amended soils monitored by phosphorus-31-nuclear magnetic resonance spectroscopy. Soil Sci. Soc. Am. J. 1988, 52, 1593–1596. [Google Scholar] [CrossRef]
- Hinedi, Z.R.; Chang, A.C. Solubility and phosphorus-31 magic angle spinning nuclear magnetic resonance of phosphorus in sludge-amended soils. Soil Sci. Soc. Am. J. 1989, 53, 1057–1061. [Google Scholar] [CrossRef]
- Leytem, A.B.; Plumstead, P.W.; Maguire, R.O.; Kwanyuen, P.; Brake, J. What aspect of dietary modification in broilers controls litter water-soluble phosphorus. J. Environ. Qual. 2007, 36, 453–463. [Google Scholar] [CrossRef] [PubMed]
- Giles, C.; Cade-Menun, B.; Hill, J. The inositol phosphates in soils and manures: Abundance, cycling, and measurement. Can. J. Soil Sci. 2011, 91, 397–416. [Google Scholar] [CrossRef]
- L’annunziata, M.; Gonzalez, J. Soil metabolic transformations of carbon-14-myo-inositol, carbon-14-phytic acid and carbon-14-iron (III) phytate. In Soil Organic Matter Studies; International Atomic Energy Agency: Vienna, Austria, 1977. [Google Scholar]
- Smith, D.H.; Clark, F.E. Anion-exchange chromatography of inositol phosphates from soil. Soil Sci. 1951, 72, 353–360. [Google Scholar] [CrossRef]
- Caldwell, A.; Black, C. Inositol hexaphosphate: II. Synthesis by soil microorganisms 1. Soil Sci. Soc. Am. J. 1958, 22, 293–296. [Google Scholar] [CrossRef]
- Cosgrove, D.J. Inositol polyphosphates in activated sludge 1. J. Environ. Qual. 1973, 2, 483–485. [Google Scholar] [CrossRef]
- Martin, J.-B.; Laussmann, T.; Bakker-Grunwald, T.; Vogel, G.; Klein, G. Neo-inositol polyphosphates in the amoeba entamoeba histolytica. J. Biol. Chem. 2000, 275, 10134–10140. [Google Scholar] [CrossRef]
- Simons, P.C.M.; Jongbloed, A.W. Influence of nutritional and physiological factors on phosphorus content of animal manures. In Sponsored by the Commission of the European Communities, Directorate-General for Agriculture, and Directorate-General for Research, Science and Education, Proceedings of the Phosphorus in Sewage Sludge and Animal Waste Slurries: EEC Seminar/Organized Jointly by the CEC and the Institute for Soil Fertility, Haren (Gr.), Groningen, Netherlands, 12–13 June 1980; Hucker, T.W.G., Catroux, G., Eds.; Dordrecht: Holland, The Netherlands; D. Reidel Pub. Co.: Boston, MA, USA; Sold: Hingham, MA, USA, 1981. [Google Scholar]
- Barnett, G.M. Phosphorus forms in animal manure. Bioresour. Technol. 1994, 49, 139–147. [Google Scholar] [CrossRef]
- Turner, B.L.; Mahieu, N.; Condron, L.M. The phosphorus composition of temperate pasture soils determined by NaOH-EDTA extraction and solution 31P NMR spectroscopy. Org. Geochem. 2003, 34, 1199–1210. [Google Scholar] [CrossRef]
- Murphy, P.N.C.; Bell, A.; Turner, B.L. Phosphorus speciation in temperate basaltic grassland soils by solution 31P NMR spectroscopy. Eur. J. Soil Sci. 2009, 60, 638–651. [Google Scholar] [CrossRef]
- Tate, K.R.; Newman, R.H. Phosphorus fractions of a climosequence of soils in New Zealand tussock grassland. Soil Biol. Biochem. 1982, 14, 191–196. [Google Scholar] [CrossRef]
- Lavahun, M.F.E.; Joergensen, R.G.; Meyer, B. Activity and biomass of soil microorganisms at different depths. Biol. Fertil. Soils 1996, 23, 38–42. [Google Scholar] [CrossRef]
- Fraser, D.G.; Doran, J.W.; Sahs, W.W.; Lesoing, G.W. Soil microbial populations and activities under conventional and organic management. J. Environ. Qual. 1988, 17, 585–590. [Google Scholar] [CrossRef]
- Martyniuk, S.; Wagner, G.H. Quantitative and qualitative examination of soil microflora associated with different management systems. Soil Sci. 1978, 125, 343–350. [Google Scholar] [CrossRef]
- Condron, L.M.; Frossard, E.; Tiessen, H.; Newmans, R.H.; Stewart, J.W.B. Chemical nature of organic phosphorus in cultivated and uncultivated soils under different environmental conditions. J. Soil Sci. 1990, 41, 41–50. [Google Scholar] [CrossRef]
- Canellas, L.P.; Espindola, J.A.A.; Guerra, J.G.M.; Teixeira, M.G.; Velloso, A.C.X.; Rumjanek, V.M. Phosphorus analysis in soil under herbaceous perennial leguminous cover by nuclear magnetic spectroscopy. Pesqui. Agropecu. Bras. 2004, 39, 589–596. [Google Scholar] [CrossRef][Green Version]
- Vincent, A.G.; Schleucher, J.; Gröbner, G.; Vestergren, J.; Persson, P.; Jansson, M.; Giesler, R. Changes in organic phosphorus composition in boreal forest humus soils: The role of iron and aluminium. Biogeochemistry 2012, 108, 485–499. [Google Scholar] [CrossRef]
- McDowell, R.W.; Cade-Menun, B.; Stewart, I. Organic phosphorus speciation and pedogenesis: Analysis by solution 31P nuclear magnetic resonance spectroscopy. Eur. J. Soil Sci. 2007, 58, 1348–1357. [Google Scholar] [CrossRef]
- Chapman, V.J. Seaweeds and Their Uses, 2nd ed.; Springer: Methuen, MA, USA; London, UK, 1970; p. 304. [Google Scholar]
- Stutter, M.I. The composition, leaching, and sorption behavior of some alternative sources of phosphorus for soils. Ambio 2015, 44, 207–216. [Google Scholar] [CrossRef]
- Hernæs, P. Karmoys Historie–som det Stiger Frem. Bind 1: Fra Istid til 1050; Karmoy Kommune: Kopervik, Norway, 1997. [Google Scholar]
- Statistics Norway Tabell: 09594: Arealbruk Og Arealressurser, Etter Arealklasser (km2). 2017. Available online: https://www.ssb.no/statistikkbanken (accessed on 30 November 2017).
- Puschmann, O.; Reid, S.J.; Fjellstad, W.J.; Hofsten, J.; Dramstad, W. Tilstandsbeskrivelse av Norske Jordbruksregioner ved Bruk av Statistikk. NIJOS-rapport 17. 2004. Available online: https://nibio.brage.unit.no/nibio-xmlui/handle/11250/2558439 (accessed on 29 October 2019).
- Lundberg, A. Karmøys Flora: Biologisk Mangfald i Eit Kystlandskap; Bergen-Sandviken Fagbokforl: Sandviken, Norway, 1998; ISBN 82-7674-472-9. [Google Scholar]
- Lundberg, A.; Handegaard, T. Changes in the spatial structure and function of coastal cultural landscapes. GeoJournal 1996, 39, 167–178. [Google Scholar] [CrossRef]
- Lundberg, A. Changes in the land and the regional identity of western Norway. The case of Sandhåland, Karmøy. In Nordic Landscapes: Region and Belonging on the Northern Edge of Europe; University of Minnesota Press: Minneapolis, MA, USA, 2008; pp. 344–371. [Google Scholar]
- Kvamme, M. En vegetasjonshistorisk undersøkelse av kulturlandskapets utvikling på Lurekalven, Lindås hd., Hordaland. Unpublished Cand. Real Thesis, University of Bergen, Bergen, Norway, 1982. [Google Scholar]
- Myhre, B. The Early Viking Age in Norway. Acta Archaeol. 2000, 71, 35–47. [Google Scholar] [CrossRef]
- Opedal, A. Homslands large bosetningshistorie. Fra Haug Ok Heidni 1994, 3, 19–21. [Google Scholar]
- Sølvberg, I.Ø. Driftsmåter i Vestnorsk Jordbruk Ca. 600–1350; Universitets-forl.: Bergen, Norway, 1976; Volume 4, ISBN 82-00-01571-8. [Google Scholar]
- Donaldson, J. General View of the Agriculture of the County of Nairn, the Eastern Coast of Inverness-shire, and the Parish of Dyke, and Part of Edenkeillie, in the County of Elgin, and Forres. By James Donaldson… Drawn Up for the Consideration of the Board of Agriculture and Internal Improvement; B. Millan: London, UK, 1794. [Google Scholar]
- Lübke, H.; Lüth, F.; Terberger, T.; Landesmuseum, A.; für Bodendenkmalpflege, L. Neue Forschungen zur Steinzeit im südlichen Ostseegebiet. Beitr. Zur 2004, 46, 221–241. [Google Scholar]
- Preuß, J. Das Neolithikum in Mitteleuropa. Kulturen–Wirtschaft–Umwelt vom 6. Bis 3. Jahrtausend vu Z. In Übersichten zum Stand der Forschung; Beier & Beran: Weissbach, Germany, 1998; p. 700. [Google Scholar]
- Schier, W. Extensiver Brandfeldbau und die Ausbreitung der neolithischen Wirtschaftsweise in Mitteleuropa und Südskandinavien am Ende des 5. Jahrtausends v. Chr. Prähistorische Z. 2009, 84, 15–43. [Google Scholar] [CrossRef]
- Heinrich, D. Die Tierknochen des frühneolithischen Wohnplatzes Wangels LA 505. Ein Vorbericht. Offa 1999, 54, 43–48. [Google Scholar]
- Schmölcke, U. Küstenleben vor 6200 Jahren-Archäozoologische Untersuchungen der Tierknochen von Timmendorf-Nordmole (Insel Poel). Natur- und Landeskunde; Zeitschrift für Schleswig-Holstein: Hamburg, Germany; Mecklenburg, Germany, 2003; Volume 110, pp. 100–105. [Google Scholar]
- Schleinert, D. Die Schwedische Landesaufnahme der Insel Poel 1698; Edition Temmen: Bremen, Germany, 2009. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Depth (cm) | Pt | Al | Ca | Fe | K | Mg | Mn | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| mg kg−1 | mg kg−1 soil (% of mol kg−1 Soil) | |||||||||||||
| Island of Feøy (SW Norway) | ||||||||||||||
| F | 0–30 | 3087 c | 18967 c | (38) | 9163 c | (12) | 30374 ab | (30) | 714 kl | (1) | 7902 b | (18) | 1220 cd | (1) |
| 30–45 | 3682 a | 19383 c | (39) | 7135 c | (10) | 31593 a | (31) | 662 l | (1) | 7763 b | (17) | 1587 ab | (2) | |
| Island of Karmøy (SW Norway) | ||||||||||||||
| Hi | 0–30 | 3196 b | 24735 ab | (44) | 8332 bc | (10) | 28436 c | (24) | 1662 efg | (2) | 9599 a | (19) | 1796 a | (2) |
| 30–45 | 2223 g | 25363 a | (43) | 7848 c | (9) | 29809 bc | (24) | 1565 fgh | (2) | 11422 a | (21) | 1347 bc | (1) | |
| S | 0–30 | 1118 h | 8374 hi | (13) | 68987 ab | (72) | 8955 k | (7) | 1099 i | (1) | 3667 e | (6) | 595 h | (<1) |
| 30–45 | 869 i | 6364 k | (7) | 114252 a | (83) | 7375 n | (4) | 1010 ij | (1) | 4060 d | (5) | 425 m | (<1) | |
| Jæren region (SW Norway) | ||||||||||||||
| PE1 | 0–30 | 2623 d | 10998 gh | (43) | 3915 hi | (10) | 18374 fg | (35) | 1122 i | (3) | 1580 j | (7) | 498 i | (1) |
| 35–60 | 2690 d | 11806 fg | (45) | 3125 ij | (8) | 19374 f | (36) | 1084 i | (3) | 1634 j | (7) | 651 g | (1) | |
| PE4 | 0–30 | 2584 e | 13586 ef | (39) | 6098 d | (12) | 24502 e | (34) | 1718 def | (3) | 3524 f | (11) | 887 f | (1) |
| 35–60 | 2490 f | 14319 e | (39) | 5592 de | (10) | 26179 d | (34) | 2318 bc | (4) | 3946 d | (12) | 892 ef | (1) | |
| Island of Sjaelland (Denmark) | ||||||||||||||
| S-7 | 0–10 | 323 q | 4096 l | (42) | 1641 l | (11) | 5145 o | (26) | 1078 i | (8) | 1012 o | (12) | 173 q | (1) |
| 10–45 | 318 q | 3588 l | (41) | 1438 m | (11) | 5105 o | (28) | 841 jk | (7) | 871 p | (11) | 211 pq | (1) | |
| Island of Fehmarn (Germany) | ||||||||||||||
| F-6 | 0–41 | 527 l | 7544 ij | (43) | 2886 jk | (11) | 9378 j | (26) | 1668 efg | (7) | 1783 i | (11) | 423 m | (1) |
| 41–53 | 264 s | 11316 gh | (45) | 3101 ij | (8) | 14895 i | (29) | 2057 cd | (6) | 2347 hi | (10) | 982 de | (2) | |
| F-8 | 0–26 | 567 k | 17595 cd | (50) | 4572 fg | (9) | 17586 gh | (24) | 3547 a | (7) | 3325 f | (10) | 341 op | (<1) |
| 26–34 | 304 r | 21186 bc | (47) | 4318 gh | (7) | 25425 de | (28) | 3681 a | (6) | 4891 c | (12) | 624 gh | (1) | |
| F-9 | 0–35 | 621 j | 14019 e | (47) | 4769 ef | (11) | 14993 i | (24) | 3325 ab | (8) | 2447 gh | (9) | 459 l | (1) |
| 35–75 | 444 n | 14946 de | (47) | 5442 de | (12) | 16551 h | (25) | 2857 abc | (6) | 2622 g | (9) | 471 kl | (1) | |
| Island of Poel (Germany) | ||||||||||||||
| P-1 | 0–20 | 349 p | 6441 k | (45) | 1564 l | (7) | 8475 lm | (28) | 1521 gh | (7) | 1413 kl | (11) | 416 mn | (1) |
| 50–90 | 305 r | 6387 k | (46) | 1998 k | (10) | 8151 mn | (28) | 1121 i | (6) | 1156 n | (9) | 478 jk | (2) | |
| P-3 | 0–45 | 477 m | 6803 jk | (45) | 1556 l | (7) | 8952 k | (29) | 1722 de | (8) | 1451 k | (11) | 409 n | (1) |
| 45–75 | 404 o | 6307 k | (44) | 1613 l | (8) | 8536 l | (29) | 1392 h | (7) | 1352 lm | (11) | 402 n | (1) | |
| P-5 | 0–47 | 437 n | 6582 k | (45) | 1555 l | (7) | 8664 l | (29) | 1440 h | (7) | 1308 m | (10) | 496 ij | (2) |
| 47–80 | 244 t | 4305 l | (46) | 1047 m | (8) | 5407 o | (28) | 786 kl | (6) | 824 p | (10) | 363 o | (2) | |
| Site | Depth (cm) | Pt | Sequentially Extracted Fractions | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Resin-P | NaHCO3-Pi | NaHCO3-Po | NaOH-Pi | NaOH-Po | H2SO4-P | Residual-P | |||
| mg kg−1 | mg P kg−1 (% of Pt) | ||||||||
| Island of Feøy (SW Norway) | |||||||||
| F | 0–30 | 3370 b | 76 ab (2) | 415 cd (12) | 325 a (10) | 1438 ab (43) | 790 a (23) | 326 cd (10) | 0 j (0) |
| 30–45 | 3916 a | 69 abc (2) | 592 a (15) | 312 a (8) | 1919 a (49) | 729 a (19) | 294 ef (8) | 0 j (0) | |
| Island of Karmøy (SW Norway) | |||||||||
| Hi | 0–30 | 3320 b | 96 a (3) | 494 ab (15) | 235 ab (7) | 1455 ab (44) | 736 a (22) | 305 de (9) | 0 hij (0) |
| 30–45 | 2127 e | 77 ab (3) | 329 de (15) | 142 cd (6) | 945 ef (43) | 431b (19) | 204 ij (9) | 95 bcde (4) | |
| S | 0–30 | 956 ef | 38 ef (3) | 112 gh (10) | 22 kl (2) | 125 fg (11) | 92 gh (8) | 567 a (51) | 162 abcd (15) |
| 30–45 | 666 f | 25 ghi (3) | 95 hi (11) | 3 m (<1) | 52 h (6) | 39 lm (5) | 451 bc (52) | 203 ab (23) | |
| Jæren region (SW Norway) | |||||||||
| PE1 | 0–30 | 2708 c | 98 a (4) | 469 bc (17) | 247 ab (9) | 1189 cd (44) | 424 b (16) | 281 ef (10) | 0 hij (0) |
| 35–60 | 2760 c | 72 abc (3) | 534 ab (19) | 168 bc (6) | 1356 bc (49) | 372 b (13) | 259 fg (9) | 0 ghi (0) | |
| PE4 | 0–30 | 2376 d | 56 bcd (2) | 248 fg (10) | 130 cde (5) | 1020 de (39) | 393 b (15) | 528 ab (20) | 208 ab (8) |
| 35–0 | 2279 d | 48 cde (2) | 275 ef (11) | 125 de (5) | 963 ef (39) | 379 b (15) | 489 bc (20) | 211 a (8) | |
| Island of Sjaelland (Denmark) | |||||||||
| S-7 | 0–10 | 344 j | 13 kl (4) | 32 mn (9) | 109 ef (32) | 22 lm (6) | 108 d (32) | 59 o (17) | 0 ij (0) |
| 10–45 | 315 k | 14 jkl (4) | 28 n (9) | 79 gh (25) | 29 k (9) | 105 def (33) | 61 no (19) | 3 hij (1) | |
| Island of Fehmarn (Germany) | |||||||||
| F-6 | 0–41 | 451 g | 33 efg (6) | 39 lm (7) | 58 hi (11) | 55 h (10) | 46 jk (9) | 220 hi (42) | 76 defg (14) |
| 41–53 | 204 n | 6 mn (2) | 12 p (5) | 14 klm (5) | 25 kl (10) | 61 hi (23) | 86 l (32) | 59 defg (22) | |
| F-8 | 0–26 | 416 h | 29 fgh (5) | 46 jk (8) | 30 jk (5) | 43 ij (8) | 42 kl (7) | 226 ghi (40) | 151 abcd (27) |
| 26–34 | 222 mn | 3 n (1) | 12 p (4) | 8 lm (3) | 5 n (2) | 10 m (3) | 185 jk (61) | 82 cdef (27) | |
| F-9 | 0–35 | 451 g | 38 def (6) | 31 n (5) | 47 i (8) | 50 hi (8) | 55 ij (9) | 230 gh (37) | 170 abc (27) |
| 35–75 | 307 k | 17 ijk (4) | 40 klm (9) | 8 lm (2) | 30 k (7) | 41 kl (9) | 172 k (39) | 137 abcd (31) | |
| Island of Poel (Germany) | |||||||||
| P-1 | 0–20 | 343 j | 8 lm (2) | 27 n (8) | 69 h (20) | 38 j (11) | 119 c (34) | 81 lm (23) | 6 hij (2) |
| 50–90 | 276 l | 7 mn (2) | 14 op (5) | 47 ij (15) | 28 kl (9) | 107 de (35) | 74 mn (24) | 29 efgh (9) | |
| P-3 | 0–45 | 478 g | 19 ijk (4) | 66 ij (14) | 122 de (26) | 83 g (17) | 101 efg (21) | 85 l (18) | 0 hij (0) |
| 45–75 | 384 i | 7 lm (2) | 19 o (5) | 101 fg (25) | 52 h (13) | 121 c (30) | 85 l (21) | 20 fghi (5) | |
| P-5 | 0–47 | 401 hi | 21 hij (5) | 43 kl (10) | 77 gh (18) | 58 h (13) | 118 c (27) | 84 l (19) | 36 efgh (8) |
| 47–80 | 237 m | 6 mn (2) | 14 o (6) | 46 i (19) | 18 mn (7) | 96 fg (39) | 56 o (23) | 8 hij (3) | |
| Site | Depth (cm) | Porg | Fe | Al | Ca | K |
|---|---|---|---|---|---|---|
| % | ||||||
| Island of Feøy (SW Norway) | ||||||
| F | 0–30 | 5 | 65 | 24 | 0 | 6 |
| 30–45 | 9 | 61 | 26 | 0 | 0 | |
| Island of Karmøy (SW Norway) | ||||||
| Hi | 0–30 | 10 | 55 | 35 | 0 | 0 |
| 30–45 | 8 | 60 | 32 | 0 | 0 | |
| S | 0–30 | 12 | 47 | 0 | 31 | 11 |
| 30–45 | 12 | 44 | 0 | 36 | 9 | |
| Jæren region (SW Norway) | ||||||
| PE1 | 0–30 | 5 | 70 | 25 | 0 | 0 |
| 35–60 | 9 | 45 | 45 | 0 | 0 | |
| PE4 | 0–30 | 7 | 70 | 22 | 0 | 0 |
| 35–60 | 6 | 74 | 21 | 0 | 0 | |
| Island of Sjaelland (Denmark) | ||||||
| S-7 | 0–10 | 30 | 55 | 0 | 7 | 8 |
| 10–45 | 30 | 55 | 0 | 7 | 8 | |
| Island of Fehmarn (Germany) | ||||||
| F-6 | 0–41 | 13 | 48 | 0 | 24 | 15 |
| 41–53 | 25 | 44 | 0 | 16 | 15 | |
| F-9 | 0–35 | 18 | 39 | 0 | 23 | 20 |
| 35–75 | 21 | 48 | 0 | 13 | 19 | |
| Island of Poel (Germany) | ||||||
| P-3 | 0–45 | 17 | 83 | 0 | 0 | 0 |
| 45–75 | 11 | 89 | 0 | 0 | 0 | |
| P-5 | 0–47 | 21 | 56 | 17 | 4 | 3 |
| 480 | 28 | 54 | 7 | 6 | 4 | |
| Site | Depth (cm) | P Groups in NaOH-EDTA Extract | |||||
|---|---|---|---|---|---|---|---|
| Pt in NaOH-EDTA Extract | Phosphonate/PHOSPHONOLIPIDS | Orthophosphate | Orthophosphate Monoester | Orthophosphate Diester | Pyrophosphate | ||
| mg P kg−1 | mg P kg−1 (% of Pt) | ||||||
| Island of Feøy and Karmøy (SW Norway) | |||||||
| F | 0–30 | 3492 | 15 (<1) | 1958 (56) | 1353 (39) | 145 (4) | 22 (<1) |
| 30–45 | 4012 | 9 (<1) | 2653 (66) | 1308 (33) | 33 (1) | 10 (<1) | |
| S | 0–30 | 694 | 0 (0) | 416 (37) | 174 (16) | 97 (9) | 8 (1) |
| 30–45 | 397 | 0 (0) | 217 (25) | 114 (13) | 63 (7) | 3 (<1) | |
| Jæren region (SW Norway) | |||||||
| PE1 | 0–30 | 2634 | 4 (<1) | 1834 (68) | 670 (25) | 113 (4) | 12 (<1) |
| 35–60 | 2759 | 4 (<1) | 2004 (73) | 638 (23) | 108 (4) | 5 (<1) | |
| Island of Sjaelland (Denmark) | |||||||
| S-7 | 0–10 | 244 | 0 (0) | 65 (19) | 109 (32) | 66 (19) | 4 (1) |
| 10-–45 | 233 | 0 (0) | 72 (23) | 110 (35) | 49 (15) | 1 (<1) | |
| Island of Fehmarn (Germany) | |||||||
| F-6 | 0–41 | 402 | 0 (0) | 243 (46) | 115 (22) | 41 (8) | 4 (1) |
| 41–53 | 166 | 0 (0) | 32 (12) | 80 (30) | 52 (20) | 2 (1) | |
| Island of Poel (Germany) | |||||||
| P-3 | 0–45 | 367 | 0 (0) | 159 (33) | 121 (25) | 85 (18) | 1 (<1) |
| 45–75 | 313 | 0 (0) | 87 (22) | 168 (42) | 58 (14) | 1 (<1) | |
| Amendments/Soils | Pt | Ca | Al | Fe | Mg | K | Mn | References |
|---|---|---|---|---|---|---|---|---|
| mg kg−1 (Dry Basis) | ||||||||
| Various Organic Amendments | ||||||||
| Human excrement | 4300–16000 | 2680–4270 | - | 200000 | 930–2860 | 1780–7160 | [127,128,129,130] | |
| Swine manure | 32950 | 33225 | 1375 | 1950 | 12950 | - | 400 | [56,131] |
| Cattle manure | 5125 | 24100 | 8800 | 4525 | 16200 | - | 200 | [56,79,105,131] |
| Sheep manure | 8000 | 13000 | 3500 | 1300 | - | - | - | [79] |
| Sewage sludge manure | 15550 | 51850 | 23900 | 13600 | - | - | - | [56] |
| Deer manure | 7400 | 11700 | 5300 | 2600 | - | - | - | [79] |
| Poultry manure | 9500 | 29000 | 600 | 2850 | 8800 | 11000 | 265 | [124,131] |
| Farm yard manure | 4000 | 77000 | - | 6000 | 9000 | 8400 | 320 | [124] |
| Cattle compost | 4000–18200 | 24000 | 3400 | 2500 | 5000 | 22000 | - | [132,133] |
| Sheep compost | 19100 | [133] | ||||||
| Swine compost | 7000 | 35000 | 800 | 2000 | 7000 | 6000 | - | [132] |
| Poultry (litter) compost | 14000–29700 | 12000 | 2000 | 3100 | 5900 | 17000 | - | [132,133] |
| Sewage sludge compost | 7000 | 25000 | 3100 | 8350 | 5900 | 8650 | 410 | [124,132] |
| Seaweed of Norway, Baltic sea and France | 600–2500 | 5300–17500 | 40 | 70–2000 | 800–21400 | 9400–35700 | −3900 | [134,135,136] |
| Peat and Anthrosols | ||||||||
| Dry peat of NO Germany | 14000 | 100000 | 56000 | 266000 | 9400 | 11300 | 1100 | [137] |
| Anthrosol of Amazonia | 2400 | 13000 | - | 140 | 4000 | 17300 | 210 | [138] |
| Anthrosol of Amazonia | 3600 | 4200 | 30000 | - | 460 | 860 | 240 | [139] |
| Anthrosol of Germany | 1000 | 6600 | - | 6700 | 890 | 710 | 650 | [10] |
| Agriculture Soils of SW Norway | ||||||||
| (Ap: 0–20 cm, n = 2218, 1 site 2500 km²) | 900–1600 | 1700–3200 | 6900–11300 | 10200–17600 | 370–1500 | 240–720 | 80–250 | [140] |
| Agriculture Soils of NO Germany | ||||||||
| (Ap: 0–20 cm, n = 2218, 1 site 2500 km²) | 207–460 | 1701–3200 | 2500–6900 | 10201–17600 | 371–1500 | 720–1300 | 250–460 | [140] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Acksel, A.; Baumann, K.; Hu, Y.; Leinweber, P. A Look into the Past: Tracing Ancient Sustainable Manuring Practices by Thorough P Speciation of Northern European Anthrosols. Soil Syst. 2019, 3, 72. https://doi.org/10.3390/soilsystems3040072
Acksel A, Baumann K, Hu Y, Leinweber P. A Look into the Past: Tracing Ancient Sustainable Manuring Practices by Thorough P Speciation of Northern European Anthrosols. Soil Systems. 2019; 3(4):72. https://doi.org/10.3390/soilsystems3040072
Chicago/Turabian StyleAcksel, Andre, Karen Baumann, Yongfeng Hu, and Peter Leinweber. 2019. "A Look into the Past: Tracing Ancient Sustainable Manuring Practices by Thorough P Speciation of Northern European Anthrosols" Soil Systems 3, no. 4: 72. https://doi.org/10.3390/soilsystems3040072
APA StyleAcksel, A., Baumann, K., Hu, Y., & Leinweber, P. (2019). A Look into the Past: Tracing Ancient Sustainable Manuring Practices by Thorough P Speciation of Northern European Anthrosols. Soil Systems, 3(4), 72. https://doi.org/10.3390/soilsystems3040072