Lablab Purpureus Influences Soil Fertility and Microbial Diversity in a Tropical Maize-Based No-Tillage System

 ,
,  , ,
, , 
Abstract
1. Introduction
2. Material and Methods
2.1. Site Description
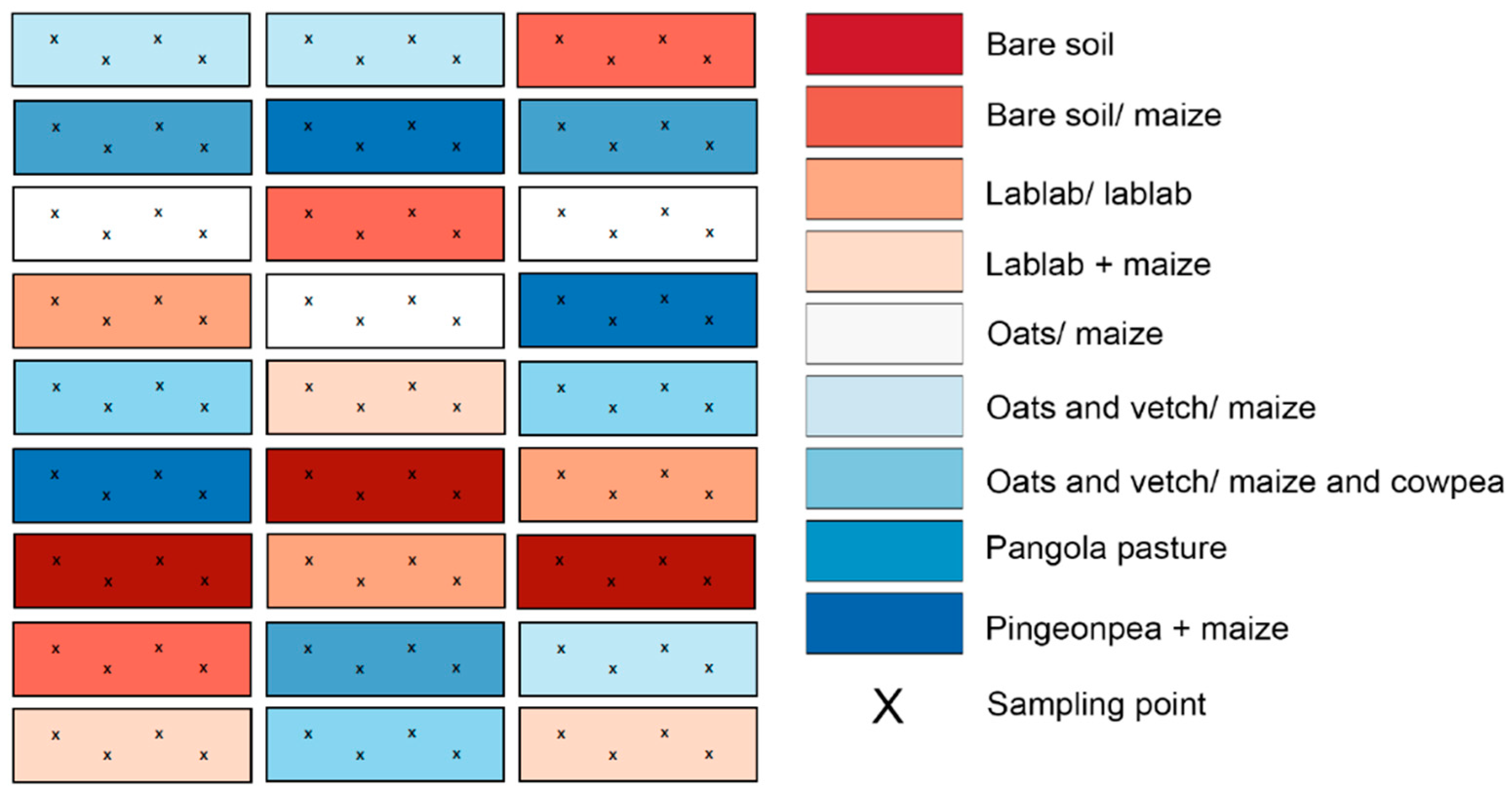
2.2. Experimental Design, Treatments, and Soil Sampling
2.3. Soil Chemical Analysis
2.4. DNA Extraction and Sequencing of the 16S rRNA Gene
2.5. Statistical Analysis
2.6. Multivariate Analysis of Microbial Community Composition
3. Results
3.1. Impacts of Cropping Systems on Soil Chemical Variables
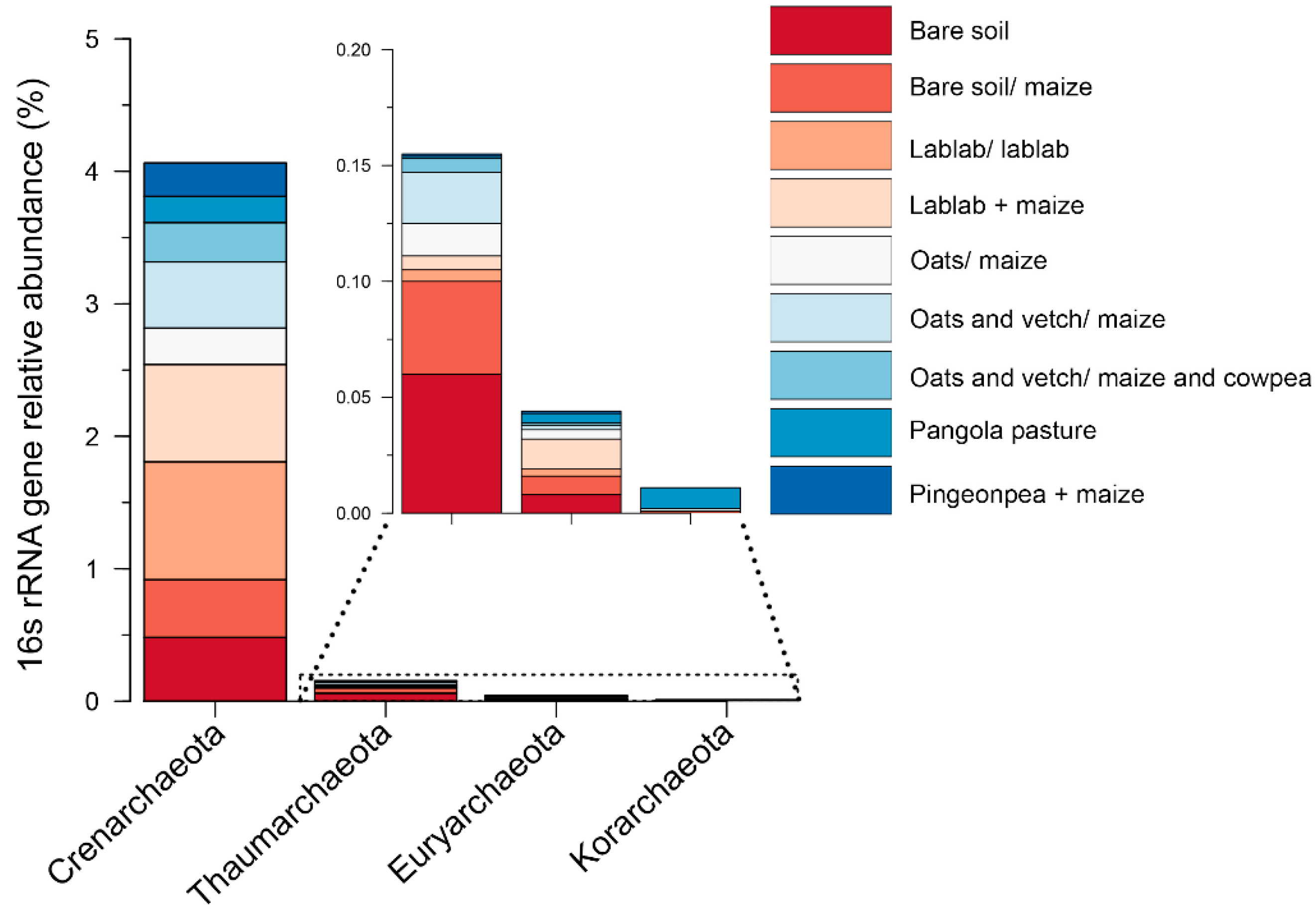
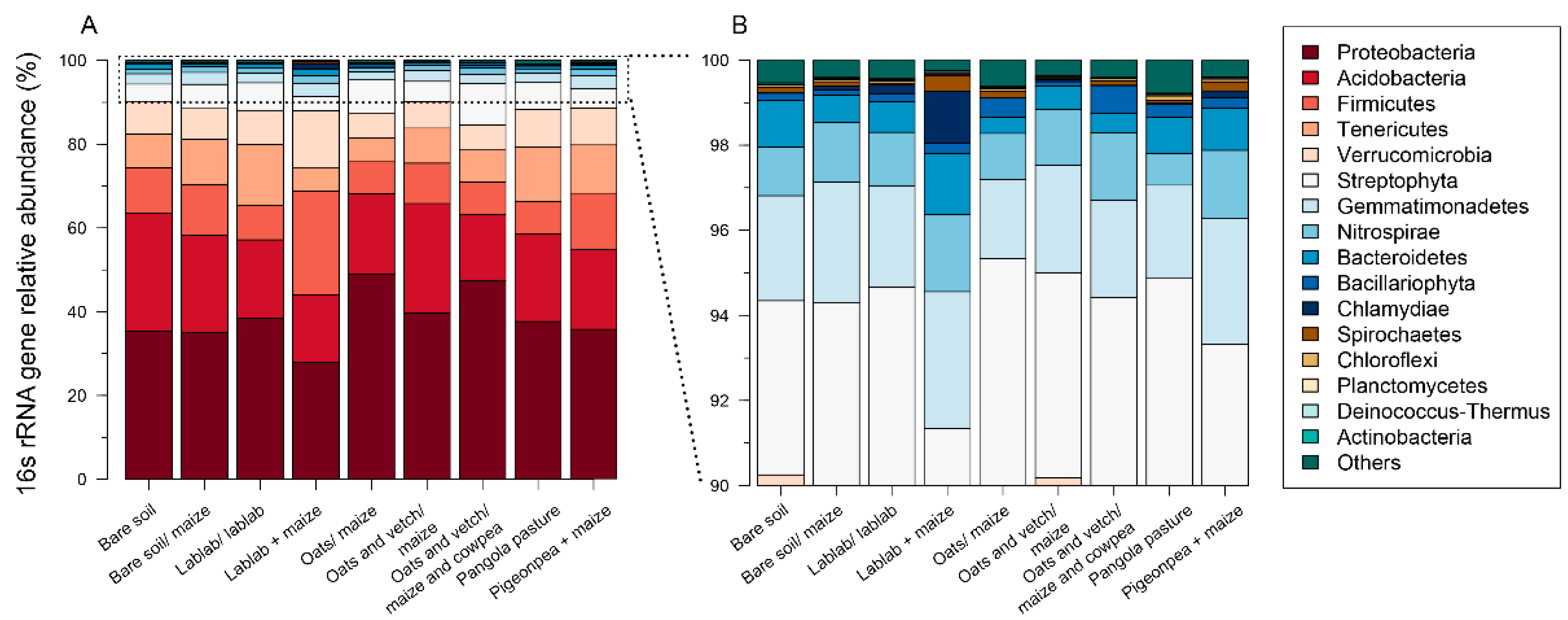
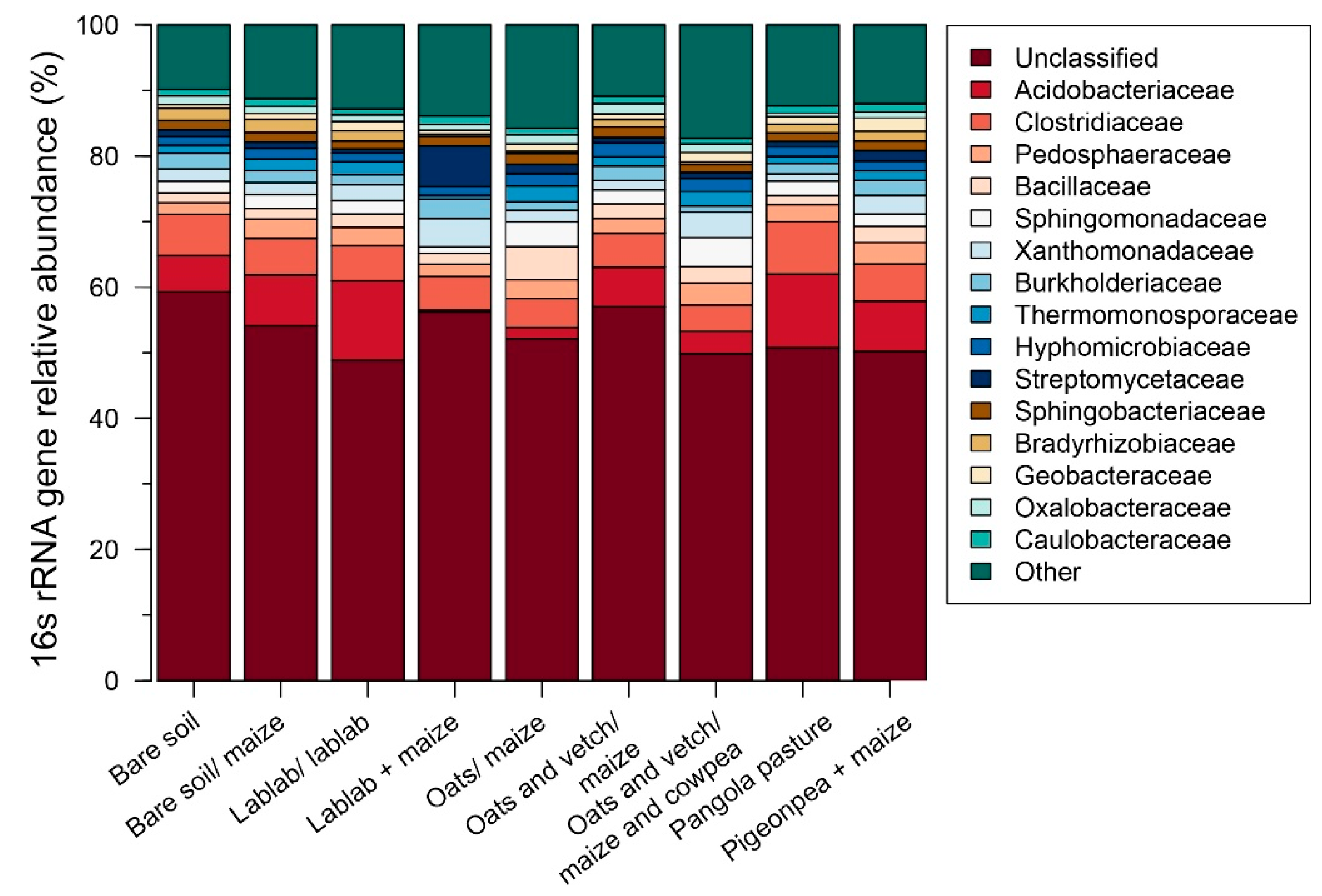
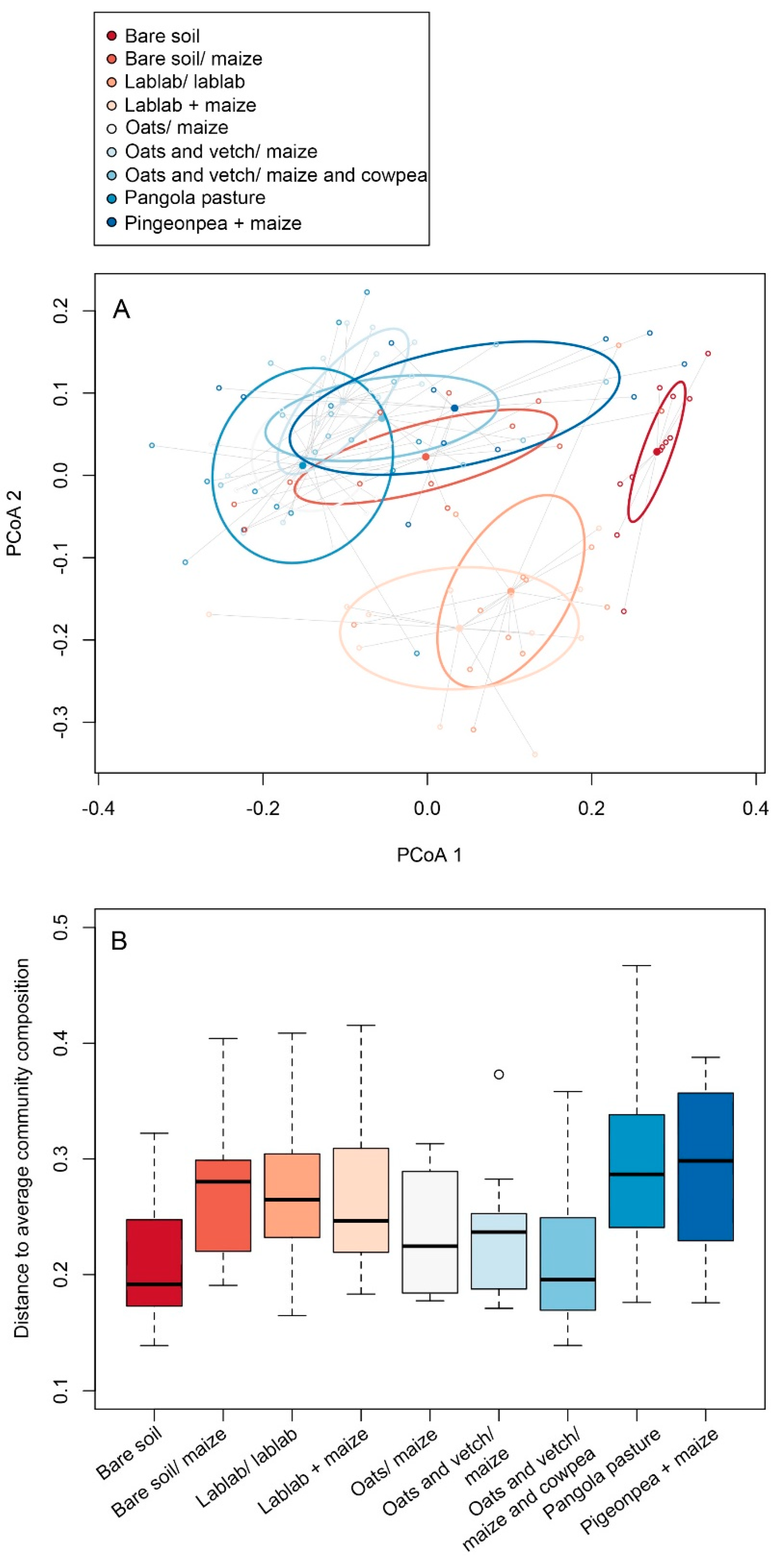
3.2. Impacts of Cropping Systems on Soil Microbial Diversity
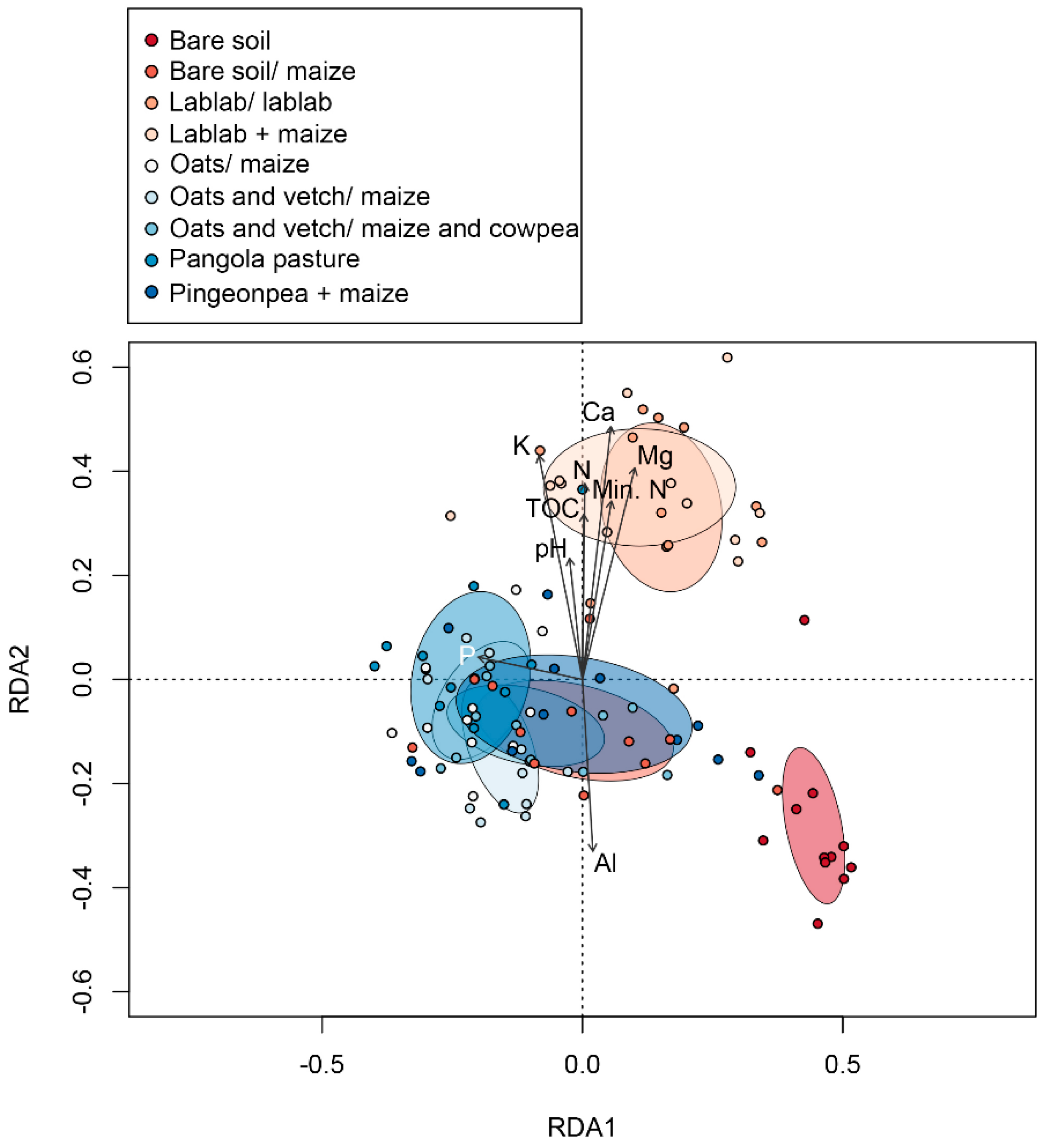
3.3. Linkages between Changes in Soil Chemical Variables and Microbial Diversity
4. Discussion
4.1. Impacts of Legume Cover Crop-Based Cropping Systems on Soil Chemical Variables
4.2. Impacts of Legume Cover Crop-Based Cropping Systems on Soil Microbial Diversity
4.3. Linkages between Changes in Soil Chemical Features and Microbial Diversity
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Martin, A.R.; Cadotte, M.W.; Isaac, M.E.; Milla, R.; Vile, D.; Violle, C. Regional and global shifts in crop diversity through the Anthropocene. PLoS ONE 2019, 14, e0209788. [Google Scholar] [CrossRef]
- Bayer, C.; Mielniczuk, J.; Martin-Neto, L.; Ernani, P.R. Stocks and humification degree of organic matter fractions as affected by no-tillage on subtropical soil. Plant Soil 2002, 238, 133–140. [Google Scholar] [CrossRef]
- Loss, A.; Pereira, M.G.; Perin, A.; Beutler, S.J.; Cunha dos Anjos, L.H.C. Carbon, nitrogen and natural abundance of δ13C e δ15N of light-fraction organic matter under no-tillage and crop-livestock integration systems. Acta Sci. Agron. 2012, 34, 465–472. [Google Scholar] [CrossRef]
- Tilman, D.; Cassman, K.; Matson, P.; Naylor, R. Agricultural sustainability and intensive production practices. Nature 2002, 418, 671–677. [Google Scholar] [CrossRef]
- McDaniel, M.D.; Grandy, A.S.; Tiemann, L.K.; Weintraub, M.N. Crop rotation complexity regulates the decomposition of high and low quality residues. Soil Biol. Biochem. 2014, 78, 243–254. [Google Scholar] [CrossRef]
- Drinkwater, L.E.; Wagoner, P.; Sarrantonio, M. Legume-based cropping systems have reduced carbon and nitrogen losses. Nature 1998, 396, 262–265. [Google Scholar] [CrossRef]
- Drinkwater, L.E.; Snapp, S.S. Nutrients in agroecosystems: Rethinking the management paradigm. Adv. Agron. 2007, 92, 163–186. [Google Scholar]
- Ren, Y.Y.; Liu, J.J.; Wang, Z.L.; Zhang, S.Q. Planting density and sowing proportions of maize/soybean intercrops affected competitive interactions and water-use efficiencies on the Loess Plateau. Eur. J. Agron. 2016, 72, 70–79. [Google Scholar] [CrossRef]
- Aller, D.; Mazur, R.; Moore, K.; Hintz, R.; Laird, D.; Horton, R. Biochar age and crop rotation impacts on soil quality. Soil Sci. Soc. Am. J. 2017, 81, 1157–1167. [Google Scholar] [CrossRef]
- Sharma, N.K.; Singh, R.J.; Mandal, D.; Kumar, A.; Alam, N.M.; Keesstra, S. Increasing farmer’s income and reducing soil erosion using intercropping in rainfed maize-wheat rotation of Himalaya, India. Agric. Ecosyst. Environ. 2017, 247, 43–53. [Google Scholar] [CrossRef]
- Zuber, S.M.; Behnke, G.D.; Nafziger, E.D.; Villamil, M.B. Crop rotation and tillage effects on soil physical and chemical properties in Illinois. Agron. J. 2015, 107, 971–978. [Google Scholar] [CrossRef]
- Nouwakpo, K.S.; Song, J.; Gonzalez, J.M. Soil structural stability assessment with the fluidized bed, aggregate stability, and rainfall simulation on long-term tillage and crop rotation systems. Soil Tillage Res. 2018, 178, 65–71. [Google Scholar] [CrossRef]
- Bayer, C.; Mielniczuck, J.; Amado, T.J.C.; Martin-Neto, L.; Fernandes, S.V. Organic matter storage in a sandy clay loam Acrisol affected by tillage and cropping systems in southern Brazil. Soil Tillage Res. 2000, 54, 101–109. [Google Scholar] [CrossRef]
- Diekow, J.; Mielniczuk, J.; Knicker, H.; Bayer, C.; Dick, D.P.; Kögel-Knabner, I. Soil C and N stocks as affected by cropping systems and nitrogen fertilisation in a southern Brazil Acrisol managed under no-tillage for 17 years. Soil Tillage Res. 2004, 81, 87–95. [Google Scholar] [CrossRef]
- Bayer, C.; Dieckow, J.; Amado, T.J.C.; Eltz, F.L.F.; Vieira, F.C.B. Cover Crop Effects Increasing Carbon Storage in a Subtropical No-Till Sandy Acrisol. Commun. Soil Sci. Plant Anal. 2009, 40, 9–10. [Google Scholar] [CrossRef]
- Schipanski, M.E.; Barbercheck, M.; Douglas, M.R.; Finney, D.M.; Haider, K.; Kaye, J.P.; Kemanian, A.R.; Mortensen, D.A.; Ryan, M.R.; Tooker, J. A framework for evaluating ecosystem services provided by cover crops in agroecosystems. Agric. Syst. 2014, 125, 12–22. [Google Scholar] [CrossRef]
- Lehman, R.M.; Osborne, S.L.; Duke, S.E. Diversified no-till crop rotation reduces nitrous oxide emissions, increases soybean yields, and promotes soil carbon. Accrual. Soil Sci. Soc. Am. J. 2017, 81, 76–83. [Google Scholar] [CrossRef]
- Brennan, E.B.; Acosta-Martinez, V. Cover cropping frequency is the main driver of soil microbial changes during six years of organic vegetable production. Soil Biol. Biochem. 2017, 109, 188–204. [Google Scholar] [CrossRef]
- Comin, J.J.; Ferreira, L.B.; dos Santos, L.H.; Koucher, L.P.; Machado, L.N.; dos Santos, E.; Mafra, A.L.; Kurtz, C.; Souza, M.; Brunetto, G. Carbon and nitrogen contents and aggregation index of soil cultivated with onion for seven years using crop successions and rotations. Soil Tillage Res. 2018, 184, 195–202. [Google Scholar] [CrossRef]
- Manns, H.R.; Martin, R.C. Cropping system yield stability inresponse to plant diversity and soil organic carbon in temperate ecosystems. Agroecol. Sustain. Food Syst. 2018, 42, 724–750. [Google Scholar] [CrossRef]
- Hawkins, B.A.; Mills, N.J.; Jervis, M.A.; Price, P.W. Is the biological control of insects a natural phenomenon? Oikos 1999, 86, 493–506. [Google Scholar] [CrossRef]
- Power, A.G. Ecosystem services and agriculture: Tradeoffs and synergies. Philos. Trans. R. Soc. B 2010, 365, 2959–2971. [Google Scholar] [CrossRef]
- Mendes, R.; Garbeva, P.; Raaijmakers, J.M. The rhizosphere microbiome: Significance of plant beneficial, plant pathogenic, and human pathogenic microorganisms. FEMS Microbiol. Rev. 2013, 37, 634–663. [Google Scholar] [CrossRef]
- Al-Dalain, S.A. Efect of intercropping of zea maize with potato (Solanum tuberosum, L) on potato growth and on the productivity and land equivalent ratio of potato and zea maize. Agric. J. 2009, 4, 164–170. [Google Scholar]
- Zhang, W.P.; Liu, G.C.; Sun, J.H.; Fornara, D.; Zhang, L.Z.; Zhang, F.F.; Li, L. Temporal dynamics of nutrient uptake by neighboring plant species: Evidence from intercropping. Funct. Ecol. 2017, 31, 469–479. [Google Scholar] [CrossRef]
- Dong, N.; Tang, M.M.; Zhang, W.P.; Bao, X.G.; Wang, Y.; Christie, P.; Li, L. Temporal Differentiation of Crop Growth as One of the Drivers of Intercropping Yield Advantage. Nat. Sci. Rep. 2018, 8, 3110. [Google Scholar] [CrossRef]
- Bevivino, A.; Dalmastri, C. Impact of Agricultural Land Management on Soil Bacterial Community: A Case Study in the Mediterranean Area. In Soil Biological Communities and Ecosystem Resilience; Chapter: Sustainability in Plant and Crop Protection Series; Lukac, M., Grenni, P., Gamboni, M., Eds.; Springer Nature: Cham, Switzerland, 2017; Volume 1, pp. 77–95. [Google Scholar]
- Peralta, A.L.; Sun, Y.; Marshall, D.; McDaniel, J.; Lennon, T. Crop rotational diversity increases disease suppressive capacity of soil microbiomes. Ecosphere 2018, 9, e02235. [Google Scholar] [CrossRef]
- McDaniel, M.D.; Grandy, A.S. Soil microbial biomass and function are altered by 12 years of crop rotation. Soil 2016, 2, 583–599. [Google Scholar] [CrossRef]
- Hooper, D.U.; Bignell, D.E.; Brown, V.K.; Brussaard, L.; Mark, J.D.; Wall, D.H.; Wardle, D.A.; Coleman, D.C.; Giller, K.E.; Lavelle, P. Interactions between aboveground and belowground biodiversity in terrestrial ecosystems: Patterns, mechanisms, and feedbacks. Bioscience 2000, 50, 1049–1061. [Google Scholar] [CrossRef]
- Kent, D.; Triplett, E.W. Microbial communities and their interactions in soil and rhizosphere ecosystems. Annu. Rev. Microbiol. 2002, 5, 211–236. [Google Scholar] [CrossRef]
- Zak, D.R.; Holmes, H.E.; White, D.C.; Peacock, A.D.; Tilman, D. Plant diversity, soil microbial communities, and ecosystem function: Are there any links? Ecology 2002, 84, 2042–2050. [Google Scholar] [CrossRef]
- Hooper, D.U.; Chapin, F.; Ewel, J.; Hector, A.; Inchausti, P.; Lavorel, S.; Lawton, J.; Lodge, D.; Loreau, M.; Naeem, S. Effects of biodiversity on ecosystem functioning: A consensus of current knowledge. Ecol. Monogr. 2005, 75, 3–35. [Google Scholar] [CrossRef]
- Berg, G.; Smalla, K. Plant species and soil type cooperatively shape the structure and function of microbial communities in the rhizosphere. FEMS Microbiol. Ecol. 2009, 68, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Finney, D.M.; Murrell, E.G.; White, C.M.; Baraibar, B.; Barbercheck, M.E.; Bradley, B.A.; Cornelisse, S.; Hunter, M.C.; Kaye, J.P.; Mortensen, D.A. Ecosystem Services and Disservices Are Bundled in Simple and Diverse Cover Cropping Systems. Agric. Environ. Lett. 2017, 3, 170033. [Google Scholar] [CrossRef]
- Wood, S.A.; Karp, D.S.; DeClerck, F.; Kremen, C.; Naeem, S.; Palm, C.A. Functional traits in agriculture: Agrobiodiversity and ecosystem services. Trends Ecol. Evol. 2015, 30, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Wood, S.A.; Bradford, M.A. Leveraging a New Understanding of How Belowground Food Webs Stabilize Soil Organic Matter to Promote Ecological Intensification of Agriculture. In Soil Carbon Storage—Modulators, Mechanisms and Modeling, 1st ed.; Brajesh Singh, Ed.; Elsevier: London, UK, 2018; Volume 4, Chapter 4; pp. 117–136. [Google Scholar]
- Yu, P.; Hochholdinger, F. The Role of Host Genetic Signatures on Root–Microbe Interactions in the Rhizosphere and Endosphere. Front. Plant Sci. 2018, 9, 1896. [Google Scholar] [CrossRef]
- Smalla, K.; Wieland, G.; Buchner, A.; Zock, A.; Parzy, J.; Kaiser, S.; Roskot, N.; Heuer, H.; Berg, G. Bulk and rhizosphere soil bacterial communities studied by denaturing gradient gel electrophoresis: Plant-dependent enrichment and seasonal shifts revealed. Appl. Environ. Microbiol. 2001, 67, 4742–4751. [Google Scholar] [CrossRef]
- Li, X.Z.; Rui, J.P.; Xiong, J.B.; Li, J.B.; He, Z.L.; Zhou, J.Z.; Yannarell, A.C.; Mackie, R.I. Functional Potential of Soil Microbial Communities in the Maize Rhizosphere. PLoS ONE 2014, 9, e112609. [Google Scholar] [CrossRef]
- Wen, X.; Wang, M.; Ti, J.; Wu, Y.; Chen, F. Bacterial community composition in the rhizosphere of maize cultivars widely grown in different decades. Biol. Fertil. Soils 2017, 53, 221–229. [Google Scholar] [CrossRef]
- Pfeifer, J.A.; Ley, R.E. Exploring the maize rhizosphere microbiome in the field: A glimpse into a highly complex system. Commun. Integr. Biol. 2013, 6, e25177. [Google Scholar] [CrossRef]
- Walters, A.W.; Jin, Z.; Youngblut, N.; Wallace, J.G.; Sutter, J.; Zhang, W.; González-Peña, A.; Peiffer, J.; Koren, O.; Shi, Q. Large-scale replicated field study of maize rhizosphere identifies heritable microbes. Proc. Natl. Acad. Sci. USA 2018, 115, 7368–7373. [Google Scholar] [CrossRef] [PubMed]
- Soman, S.; Li, M.W.; Kent, M. Long-term fertilizer and crop-rotation treatments differentially affect soil bacterial community structure. Plant Soil 2017, 413, 145–159. [Google Scholar] [CrossRef]
- Benitez, M.S.; Osborne, S.; Lehman, R.M. Previous crop and rotation history effects on maize seedling health and associated rhizosphere microbiome. Nat. Sci. Rep. 2017, 7, 15709. [Google Scholar] [CrossRef] [PubMed]
- Tedesco, M.J.; Gianello, C.; Bissani, C.A.; Bohnen, H.; Volkweiss, S.J. Análises de Solo, Plantas e Outros Materiais; UFRGS: Porto Alegre, Brazil, 1995. (In Portuguese) [Google Scholar]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2010, 108, 4516–4522. [Google Scholar] [CrossRef] [PubMed]
- Fagen, J.R.; Giongo, A.; Brown, C.T.; Davis-Richardson, A.; Gano, K.A.; Triplett, E.W. Characterization of the relative abundance of the citrus pathogen Ca. Liberibacter asiaticus in the microbiome of its insect 525vecor, Diaphorina citri, using high throughput 16S rRNA sequencing. Open Microbiol. J. 2012, 6, 29–33. [Google Scholar] [CrossRef] [PubMed]
- Dias, R.; Xavier, M.G.; Rossi, F.D.; Neves, M.V.; Lange, T.A.P.; Giongo, A.; De Rose, C.A.F.; Triplett, E.W. MPI-blastn and NCBI-TaxCollector: Improving metagenomic analysis with high performance classification and wide taxonomic attachment. J. Bioinform. Comput. Biol. 2014, 12, 14500131–145001317. [Google Scholar] [CrossRef] [PubMed]
- Davis-Richardson, A.G.; Ardissone, A.N.; Dias, R.; Simell, V.; Leonard, M.T.; Kemppainen, K.M.; Drew, J.C.; Schatz, D.; Atkinson, M.A.; Kolaczkowski, B.; et al. Bacteroides dorei dominates gut microbiome prior to autoimmunity in Finnish children at high risk for type 1 diabetes. Front. Microbiol. 2014, 5, 678. [Google Scholar] [CrossRef] [PubMed]
- Cole, J.R.; Wang, Q.; Fish, J.A.; Chai, B.; McGarrell, D.M.; Sun, Y.; Brown, C.T.; Porras-Alfaro, A.; Kuske, C.R.; Tiedje, J.M. Ribosomal Database Project: Data and tools for high throughput rRNA analysis. Nucl. Acids Res. 2014, 42, D63342. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P. Vegan: Community Ecology Package. R Package Version 2.5-4. Available online: https://CRAN.R-project.org/package=vegan (accessed on 10 January 2019).
- Neto, R.C.A.; Miranda, N.O.; Duda, G.P.; Góes, G.B.; Lima, A.S. Crescimento e produtividade do sorgo forrageiro BR 601 sob adubação verde. Rev. Bras. Eng. Agrícola Ambient. 2010, 14, 124–130. [Google Scholar] [CrossRef]
- Von Uexküll, H.R.; Mutert, E. Global Extent, Development and Economic Impact of Acid Soils. Plant Soil 1995, 171, 1–15. [Google Scholar] [CrossRef]
- Venter, Z.S.; Jacobs, K.; Hawkins, H.J. The impact of crop rotation on soil microbial diversity: A meta-analysis. Pedobiologia 2016, 59, 215–223. [Google Scholar] [CrossRef]
- Smith, R.G.; Mortensen, D.A.; Ryan, M.R. A new hypothesis for the functional role of diversity in mediating resource pools and weed–crop competition in agroecosystems. Weed Res. 2009, 50, 37–48. [Google Scholar] [CrossRef]
- Costa, R.; Götz, M.; Mrotzek, N.; Lottmann, J.; Berg, G.; Smalla, K. Effects of site and plants species on rhizosphere community structure as revealed by molecular analysis of microbial guilds. FEMS Microbiol. Ecol. 2006, 56, 236–249. [Google Scholar] [CrossRef] [PubMed]
- Badri, D.V.; Vivanco, J.M. Regulation and function of root exudates. Plant Cell Environ. 2009, 32, 666–681. [Google Scholar] [CrossRef]
- Geisseler, D.; Scow, K.M. Does long-term use of mineral fertilizers affect the soil microbial biomass? Better Crop. Plant Food 2014, 98, 13–15. [Google Scholar]
- Coyne, M.S.; Mikkelsen, R. Soil microorganisms contribute to plant nutrition and root health. Better Crop. Plant Food 2015, 99, 18–20. [Google Scholar]
- Zhalnina, K.; Louie, K.B.; Hao, Z.; Mansoori, N.; Rocha, U.N.; Shi, S.; Cho, H.; Karaoz, U.; Loqué, D.; Bowen, B.P. Dynamic root exudate chemistry and microbial substrate preferences drive patterns in rhizosphere microbial community assembly. Nat. Microbiol. 2018, 3, 470–480. [Google Scholar] [CrossRef]
- Lynch, J.M.; Whipps, J.M. Substrate flow in the rhizosphere. Plant Soil 1990, 129, 1–10. [Google Scholar] [CrossRef]
- Bulgarelli, D.; Schlaeppi, K.; Spaepen, S.; van Themaat, E.V.L.; Schulze-Lefert, P. Structure and functions of the bacterial microbiota of plants. Annu. Rev. Plant Biol. 2013, 64, 807–838. [Google Scholar] [CrossRef]
- Corral-Lugo, A.; Daddaoua, A.; Ortega, A.; Espinosa-Urgel, M.; Krell, T. Rosmarinic acid is a homoserine lactone mimic produced by plants that activates a bacterial quorum-sensing regulator. Sci. Signal. 2016, 9. [Google Scholar] [CrossRef]
- Jurgens, G.; Lindström, K.; Saano, A. Novel group within the kingdom Crenarchaeota from boreal forest soil. Appl. Environ. Microbiol. 1997, 63, 803–805. [Google Scholar] [PubMed]
- Ochsenreiter, T.; Selezi, D.; Quaiser, A.; Bonch-Osmolovskaya, L.; Schleper, C. Diversity and abundance of Crenarchaeota in terrestrial habitats studied by 16S RNA surveys and real time PCR. Environ. Microbiol. 2003, 5, 787–797. [Google Scholar] [CrossRef] [PubMed]
- Lehtovirta, L.E.; Prosser, J.E.; Nicol, G.W. Soil pH regulates the abundance and diversity of Group 1.1c Crenarchaeota. FEMS Microbiol. Ecol. 2009, 70, 367–376. [Google Scholar] [CrossRef] [PubMed]
- Kemnitz, D.; Kolb, S.; Conrad, R. High abundance of Crenarchaeota in a temperate acidic forest soil. FEMS Microbiol. Ecol. 2007, 60, 442–448. [Google Scholar] [CrossRef] [PubMed]
- De La Torre, J.R.; Walker, C.B.; Imgalls, A.E.; Könneke, M.; Stahl, D.A. Cultivation of a thermophilic ammonia oxidizing archaeon synthesizing crenarchaeol. Environ. Microbiol. 2007, 10, 810–818. [Google Scholar] [CrossRef] [PubMed]
- Lehtovirta-Morley, L.E.; Stoecker, K.; Vilcinskas, A.; Prosser, J.I.; Nicol, G.W. Cultivation of an obligate acidophilic ammonia oxidizer from a nitrifying acid soil. Proc. Natl. Acad. Sci. USA 2011, 108, 15892–15897. [Google Scholar] [CrossRef] [PubMed]
- Tourna, M.; Srieglmeier, M.; Spang, A.; Könneke, M.; Schintlmeister, A.; Urich, T.; Engel, M.; Schloter, M.; Wagner, M.; Richter, A.; et al. Nitrososphaera viennensis, an ammonia oxidizing archaeon from soil. Proc. Natl. Acad. Sci. USA 2011, 20, 8420–8425. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.K.; Jung, M.Y.; Yu, D.S.; Park, S.J.; Oh, T.K.; Rhee, S.K.; Kim, J.F. Genome sequence of an ammonia-oxidizing soil archaeon, “Candidatus Nitrosoarchaeum koreensis” MY1. J. Bacteriol. 2011, 193, 5539–5540. [Google Scholar] [CrossRef] [PubMed]
- Blainey, P.C.; Mosier, A.C.; Potanina, A.; Francis, C.A.; Quake, S.R. Genome of a low-salinity ammonia-oxidizing archaeon determined by single-cell and metagenomic analysis. PLoS ONE 2011, 6, 1–12. [Google Scholar] [CrossRef]
- Zhalnina, K.; Dias, R.; Leonard, M.T.; Dörr de Quadros, P.; Camargo, F.A.O.; Drew, J.C.; Farmerie, W.G.; Daroub, S.H.; Triplett, E.W. Genome sequence of Candidatus Nitrososphaera evergladensis from group I.1b isolated from Everglades soil reveals novel genomic features of the ammonia-oxidizing archaea. PLoS ONE 2014, 9, e101648. [Google Scholar] [CrossRef] [PubMed]
- Pedrós-Alió, C. The rare bacterial biosphere. Ann. Rev. Mar. Sci. 2012, 4, 449–466. [Google Scholar] [CrossRef] [PubMed]
- Teira, E.; Lekunberri, I.; Gasol, J.M.; Nieto-Cid, M.; Alvarez-Salgado, X.A.; Figueiras, P.G. Dynamics of the hydrocarbon-degrading Cycloclasticus bacteria during mesocosm-simulated oil spills. Environ. Microbiol. 2007, 9, 2551–2562. [Google Scholar] [CrossRef] [PubMed]
- Santoyo, G.; Moreno-Hagelsieb, G.; Orozco-Mosqueda, M.D.C.; Glick, B.R. Plant growth-promoting bacterial endophytes. Microbiol. Res. 2016, 183, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Da, K.; Nowak, J.; Flinn, B. Potato cytosine methylation and gene expression changes induced by a beneficial bacterial endophyte, Burkholderia phytofirmans PsJN. Plant Physiol. Biochem. 2012, 50, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Talbi, C.; Delgado, M.J.; Girard, L.; Ramírez-Trujillo, A.; Caballero-Mellado, J.; Bedmar, E.J. Burkholderia phymatum strains capable of nodulating Phaseolus vulgaris are present in Moroccan soils. Appl. Environ. Microbiol. 2010, 76, 4587–4591. [Google Scholar] [CrossRef] [PubMed]
- Bournaud, C.; de Faria, S.M.; dos Santos, J.M.F.; Tisseyre, P.; Silva, M.; Chaintreuil, C.; Farmerie, W.G.; Daroub, S.H.; Triplett, E.W. Burkholderia species are the most common and preferred nodulating symbionts of the Piptadenia group (Tribe mimoseae). PLoS ONE 2013, 8, e6347. [Google Scholar] [CrossRef] [PubMed]
- Garbeva, P.; van Veen, J.A.; van Elsas, J.D. Predominant Bacillus spp. In agricultural soil under different management regimes detected via PCR-DGGE. Microb. Ecol. 2003, 45, 302–316. [Google Scholar] [CrossRef] [PubMed]
- Köberl, M.; Müller, H.; Ramadan, E.M.; Berg, G. Desert farming benefits from microbial potential in arid soils and promotes diversity and plant health. PLoS ONE 2011, 6, e24452. [Google Scholar] [CrossRef]
- De Quadros, P.D.; Zhalnina, K.; Davis-Richardson, A.; Fagen, J.R.; Drew, J.; Bayer, C.; Camargo, F.A.; Triplett, E.W. The effect of tillage system and crop rotation on soil microbial diversity and composition in a subtropical Acrisol. Diversity 2012, 4, 375–395. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Oats + Vetch/Maize | Oats +Vetch/Maize + Cowpea | Oat/Maize | Fallow/Fallow | Lablab/Lablab | Maize/Pigeon pea Bean | Maize/Lablab Bean | Pangola/Pangola | Fallow/Maize |
|---|---|---|---|---|---|---|---|---|---|
| pH | 4.7 ± 0.4 c | 4.7 ± 0.2 de | 4.9 ± 0.3 cd | 5.0 ± 0.2bc | 5.1 ± 0.2 bc | 4.6 ± 0.2 e | 5.3 ± 0.1 ab | 5.5 ± 0.2 a | 5.0 ± 0.4 bc |
| P (mg dm−3) | 22.9 ± 2.6 b | 21.4 ± 2.7 b | 34.7 ± 8.2 a | 8.4 ± 1.0 cd | 14.2 ± 1.6 c | 32.7 ± 3.4 a | 29.1 ± 5.2 a | 7.3 ± 1.1 d | 31.9 ± 7.8 a |
| K (mg dm−3) | 232 ± 19 bc | 224 ± 24.4 bc | 220 ± 24.3 bc | 130 ± 5.6 e | 253 ± 41.6 b | 219 ± 16.9 c | 317 ± 37.6 a | 174 ± 14.1 c | 221 ± 31.1 bc |
| Al (cmolc dm−3) | 0.75 ± 0.7 a | 0.97 ± 0.4 a | 0.77 ± 0.5 a | 0.70 ± 0.3 a | 0.15 ± 0.1 b | 0.75 ± 0.3 a | 0.07 ± 0.05 b | 0.10 ± 0.01b | 0.60 ± 0.5 ab |
| Ca (cmolc dm−3) | 2.37 ± 1.1 bc | 2.05 ± 0.4 c | 2.02 ± 0.5 c | 1.93 ± 0.4 c | 4.37 ± 0.3 a | 3.09 ± 0.3 b | 3.95 ± 0.3 a | 2.95 ± 0.3 b | 1.95 ± 0.49 c |
| Mg (cmolc dm−3) | 1.35 ± 0.7 cd | 1.02 ± 0.3 d | 1.02 ± 0.3 d | 1.35 ± 0.3 cd | 2.00 ± 0.2 ab | 1.55 ± 0.3 c | 2.08 ± 0.2 a | 1.65 ± 0.2 bc | 1.27 ± 0.32 cd |
| TOC (%) | 1.56 ± 0.06 c | 1.77 ± 0.09 d | 1.42 ± 0.06 f | 1.20 ± 0.08 g | 2.24 ± 0.12 b | 2.59 ± 0.2 a | 2.03 ± 0.2 c | 1.86 ± 0.1 de | 1.34 ± 0.1 fg |
| Total N (%) | 0.13 ± 0.01 e | 0.15 ± 0.01 d | 0.12 ± 0.01 f | 0.09 ± 0.01 g | 0.18 ± 0.01 b | 0.20 ± 0.01 a | 0.17 ± 0.01 c | 0.15 ± 0.01 e | 0.11 ± 0.01 f |
| Mineral N (mg kg−1) | 8.16 ± 2.1 c | 6.31 ± 0.8 cd | 4.27 ± 1.2d | 1.83 ± 0.9 e | 12.12 ± 1.5 b | 16.73 ± 3.4 a | 15.26 ± 4.2 a | 4.01 ± 1.4 de | 4.29 ± 0.7 de |
| Cropping Systems | Averaged Reads per Treatment | Nº of Replicates per Treatment | Sum of Reads / All Replicates |
|---|---|---|---|
| Bare Soil | 53,531 | 12 | 642,375 |
| Fallow/ Maize | 56,992 | 12 | 683,900 |
| Oats/ Maize | 47,800 | 12 | 525,804 |
| Oats + Vetch / Maize | 54,351 | 12 | 652,212 |
| Oats + Vetch / Maize + Cowpea | 65,135 | 12 | 781,620 |
| Pigeon pea + Maize | 49,137 | 12 | 589,645 |
| Lablab + Maize | 65,176 | 12 | 782,115 |
| Pangola pasture | 79,635 | 12 | 875,985 |
| Lablab/ Lablab | 68,380 | 12 | 820,562 |
| TOTAL (SUM) | 540,138 | N = 108 | 5,378,124 |
| Cropping Systems | Fisher Alpha Diversity | Richness | ||||||
|---|---|---|---|---|---|---|---|---|
| Phylum | Family | Genus | Species | Phylum | Family | Genus | Species | |
| Bare Soil | 2.5 ± 0.78 a | 15.4 ± 0.14 a | 43.0 ± 0.15 b | 152.7 ± 0.15 b | 24 ± 1.8 | 103 ± 12 b | 305 ± 52 b | 898 ± 166 b |
| Fallow/Maize | 2.4 ± 0.80 ab | 14.5 ± 0.14 ab | 42.5 ± 0.22 c | 144.3 ± 0.22 c | 23 ± 1.9 | 105 ± 18 b | 312 ± 53 b | 878 ± 170 b |
| Oats/Maize | 2.3 ± 0.91 b | 13.2 ± 0.22 c | 39.7 ± 0.30 c | 125.0 ± 0.30 d | 22± 1.5 | 106 ± 15 b | 280 ± 48 c | 736 ± 175 bc |
| Oats + Vetch/ Maize | 2.3 ± 0.82 bc | 13.5 ± 0.19 c | 37.4 ± 0.22 d | 118.3 ± 0.22 e | 22 ± 2 | 98 ± 11 bc | 270 ± 37 c | 716 ± 107 bc |
| Oats + Vetch/ Maize + Cowpea | 2.2 ± 0.91 c | 13.3 ± 0.20 c | 41.0 ± 0.17 c | 136.4 ± 0.17 c | 21 ± 2.4 | 97 ± 20 bc | 301 ± 73 b | 833 ± 238 b |
| Pigeon pea + Maize | 1.9 ± 0.86 d | 11.4 ± 0.58 d | 32.2 ± 0.24 d | 99.6 ± 0.29 e | 19 ± 2.2 | 82 ± 10 c | 234 ± 49 d | 611 ± 141 c |
| Lablab + Maize | 2.3 ± 0.97 bc | 14.9 ± 0.12 a | 51.4 ± 0.33 a | 182.0 ± 0.24 a | 22 ± 2.2 | 129 ± 11 a | 342 ± 46 a | 967 ± 256 a |
| Pangola pasture | 2.2 ± 0.78 c | 14.4 ± 0.25 b | 43.4 ± 0.33 b | 137.0 ± 0.33 bc | 22 ± 1.7 | 119 ± 16 ab | 303 ± 60 b | 838 ± 227 b |
| Lablab/Lablab | 2.2 ± 0.88 c | 14.4 ± 0.10 b | 46.0 ± 0.19 b | 159.3 ± 0.19 b | 22 ± 1.7 | 122 ± 10 a | 366 ± 37 a | 1066 ± 185 a |
| Cropping Systems | Clostridium | Bacillus | Sphingomonas | Candidatus Koribacter | Burkholderia | Rhodoplanes | Bradyrhizobium | Geobacter |
|---|---|---|---|---|---|---|---|---|
| Bare Soil | 0.103 d | 0.769 a | 1.898 a | 0.795 a | 0.625 b | 0.476 cd | 0.542 a | 0.113 b |
| Fallow/Maize | 3.823 bc | 1.479 a | 1.322 b | 1.086 a | 0.820 b | 0.559 bcd | 0.584 a | 0.636 a |
| Oats/Maize | 6.200 a | 1.229 a | 1.093 bcd | 0.921 a | 0.644 b | 0.494 cd | 0.459 a | 0.739 a |
| Oats + Vetch/Maize | 3.862 b | 1.074 a | 1.023 cd | 1.219 a | 0.544 b | 0.578 bcd | 0.551 a | 0.929 a |
| Oats + Vetch/Maize + Cowpea | 3.820 bc | 1.349 a | 0.846 cd | 1.238 a | 0.506 b | 0.645 abc | 0.625 a | 0.892 a |
| Pigeon pea + Maize | 2.996 bcd | 0.068 a | 0.613 d | 1.069 a | 0.886 b | 0.816 a | 0.652 a | 0.489 ab |
| Lablab + Maize | 1.700 cd | 1.597 a | 1.713 ab | 0.698 a | 0.878 b | 0.701 ab | 0.464 a | 0.152 b |
| Pangola pasture | 5.642 a | 1.256 a | 0.506 d | 0.631 a | 0.423 b | 0.454 d | 0.515 a | 0.513 ab |
| Lablab/Lablab | 0.860 cd | 1.374 a | 0.754 cd | 0.911 a | 1.920 a | 0.657 abc | 0.668 a | 0.128 b |
| ANOVA p-value | <0.001 | 0.333 | <0.001 | 0.061 | <0.001 | <0.001 | 0.182 | <0.001 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dörr de Quadros, P.; R. Martin, A.; Zhalnina, K.; Dias, R.; Giongo, A.; Fulthorpe, R.; Bayer, C.; W. Triplett, E.; A. de O. Camargo, F. Lablab Purpureus Influences Soil Fertility and Microbial Diversity in a Tropical Maize-Based No-Tillage System. Soil Syst. 2019, 3, 50. https://doi.org/10.3390/soilsystems3030050
Dörr de Quadros P, R. Martin A, Zhalnina K, Dias R, Giongo A, Fulthorpe R, Bayer C, W. Triplett E, A. de O. Camargo F. Lablab Purpureus Influences Soil Fertility and Microbial Diversity in a Tropical Maize-Based No-Tillage System. Soil Systems. 2019; 3(3):50. https://doi.org/10.3390/soilsystems3030050
Chicago/Turabian StyleDörr de Quadros, Patricia, Adam R. Martin, Kateryna Zhalnina, Raquel Dias, Adriana Giongo, Roberta Fulthorpe, Cimelio Bayer, Eric W. Triplett, and Flávio A. de O. Camargo. 2019. "Lablab Purpureus Influences Soil Fertility and Microbial Diversity in a Tropical Maize-Based No-Tillage System" Soil Systems 3, no. 3: 50. https://doi.org/10.3390/soilsystems3030050
APA StyleDörr de Quadros, P., R. Martin, A., Zhalnina, K., Dias, R., Giongo, A., Fulthorpe, R., Bayer, C., W. Triplett, E., & A. de O. Camargo, F. (2019). Lablab Purpureus Influences Soil Fertility and Microbial Diversity in a Tropical Maize-Based No-Tillage System. Soil Systems, 3(3), 50. https://doi.org/10.3390/soilsystems3030050