Coupled Biological and Abiotic Mechanisms Driving Carbonyl Sulfide Production in Soils

 , , , ,
, , , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Soil Collection and Processing
2.2. Trace Gas Flux Measurements
2.2.1. Trace Gas Flux Procedures and Calculations (Surveys 1 and 2)
2.2.2. Temperature Response Experiment (Survey 1)
2.2.3. Light Response Experiment (Survey 2)
2.3. Soil Characterization
2.3.1. Soil Physical, Chemical, and Microbial Community Characterization (Surveys 1 and 2)
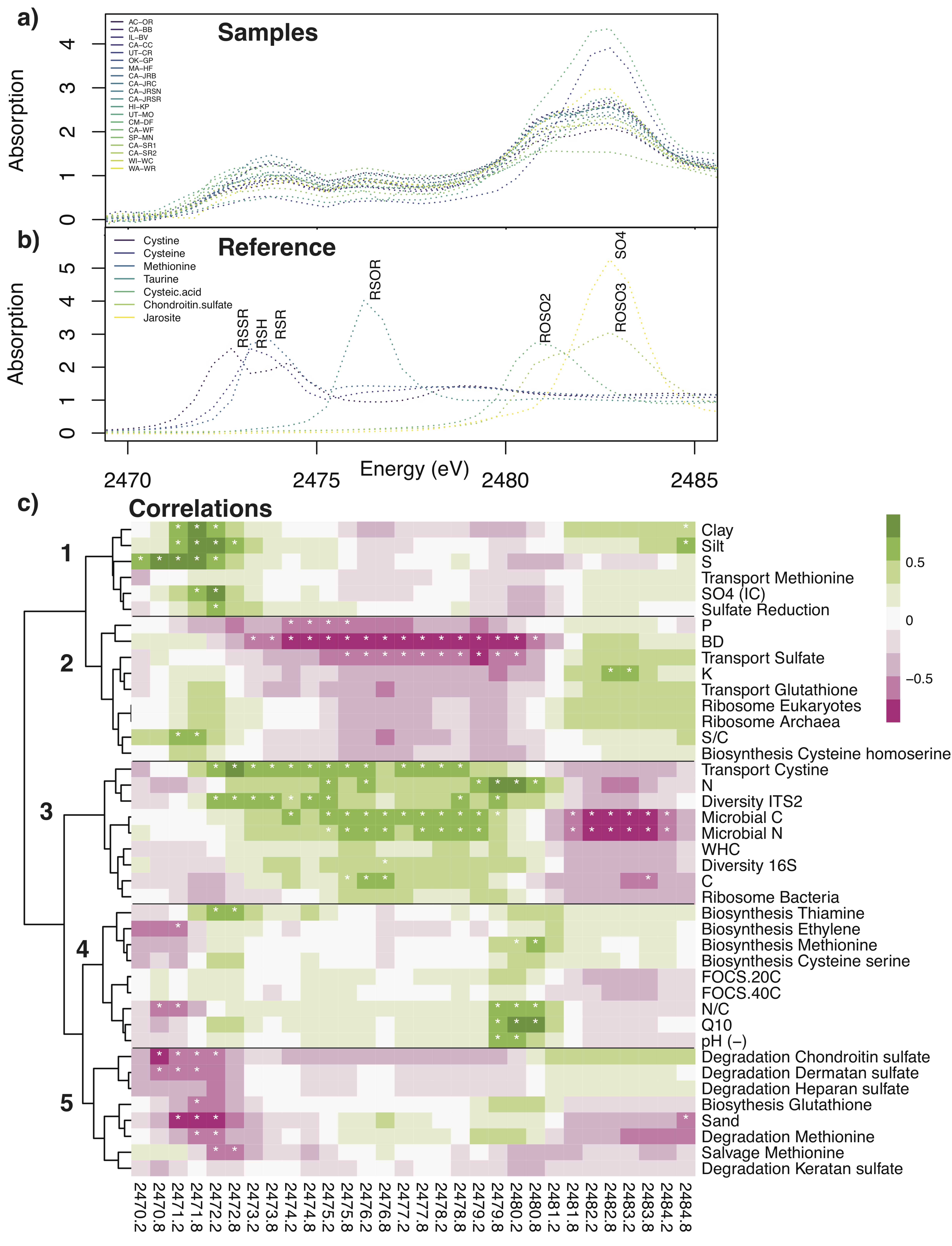
2.3.2. Soil S Speciation (Survey 1)
2.3.3. Soil Microbial Characterization (Survey 1)
2.4. Data Analysis
2.4.1. Predicting Microbial Pathways from Composition and Gene Expression Data
2.4.2. Statistical Tests and Multivariate Data Analysis
3. Results
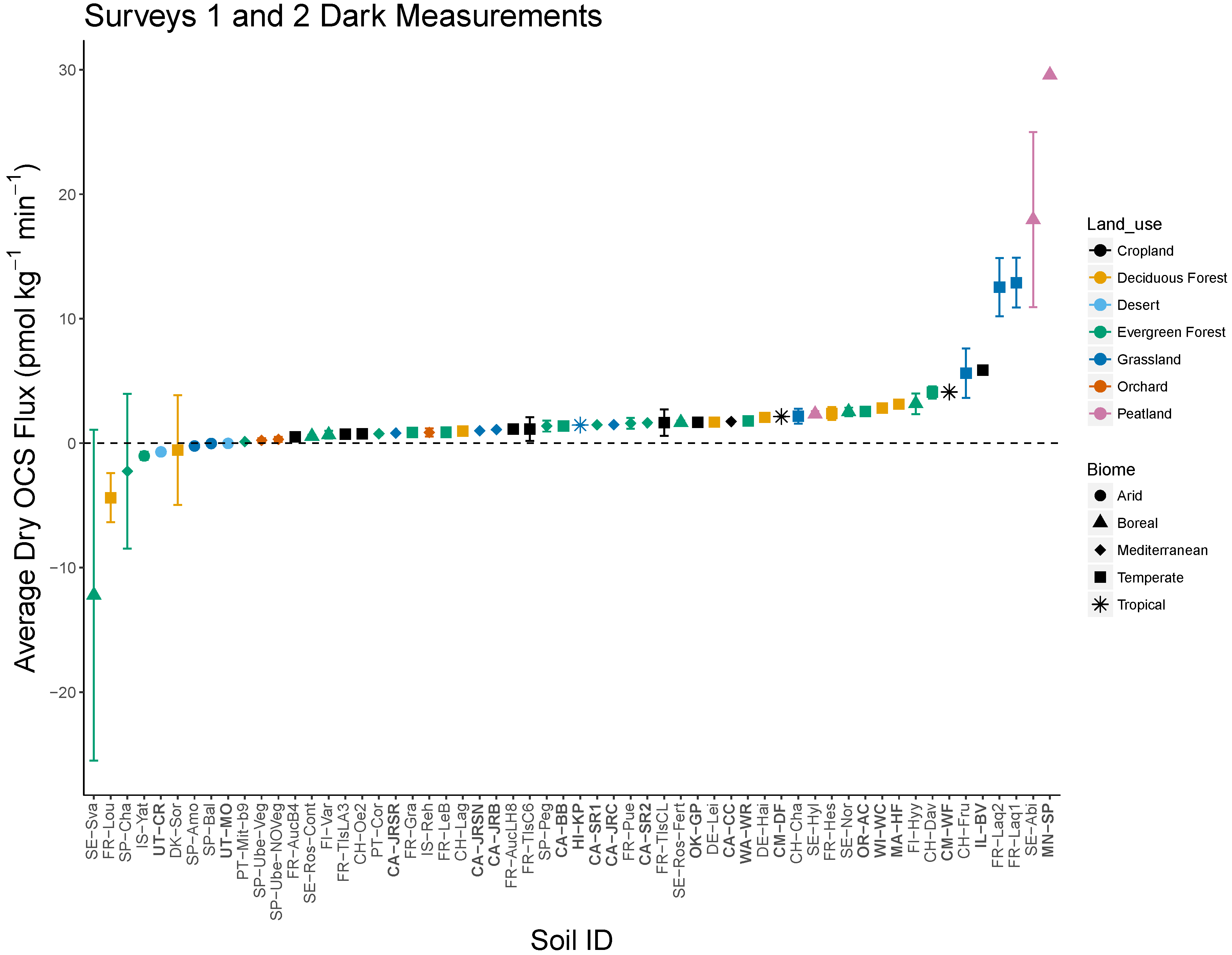
3.1. Patterns in OCS Production with Biome and Land Use
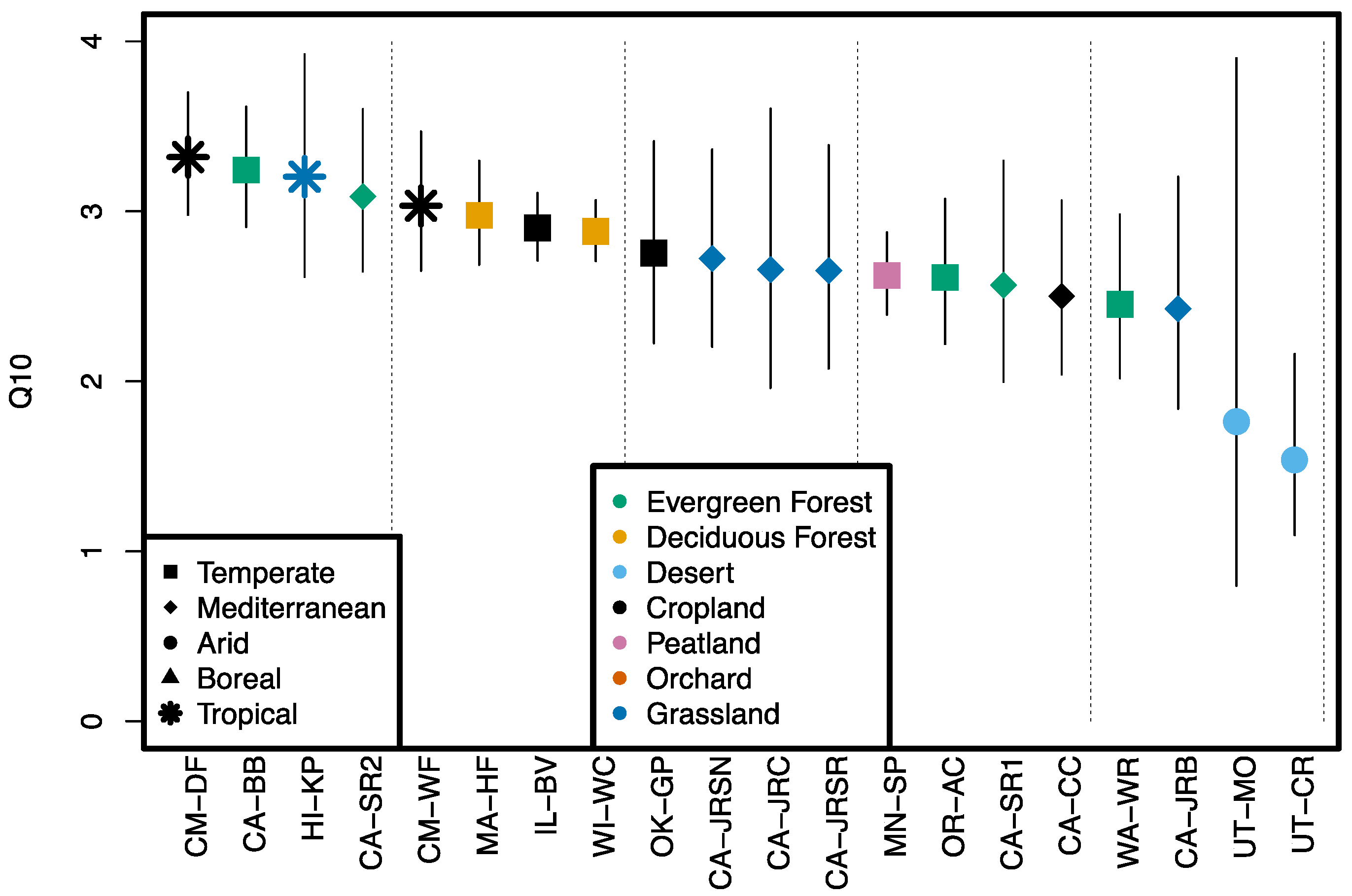
3.2. Temperature Response of OCS Production
3.3. Light Response of OCS Production
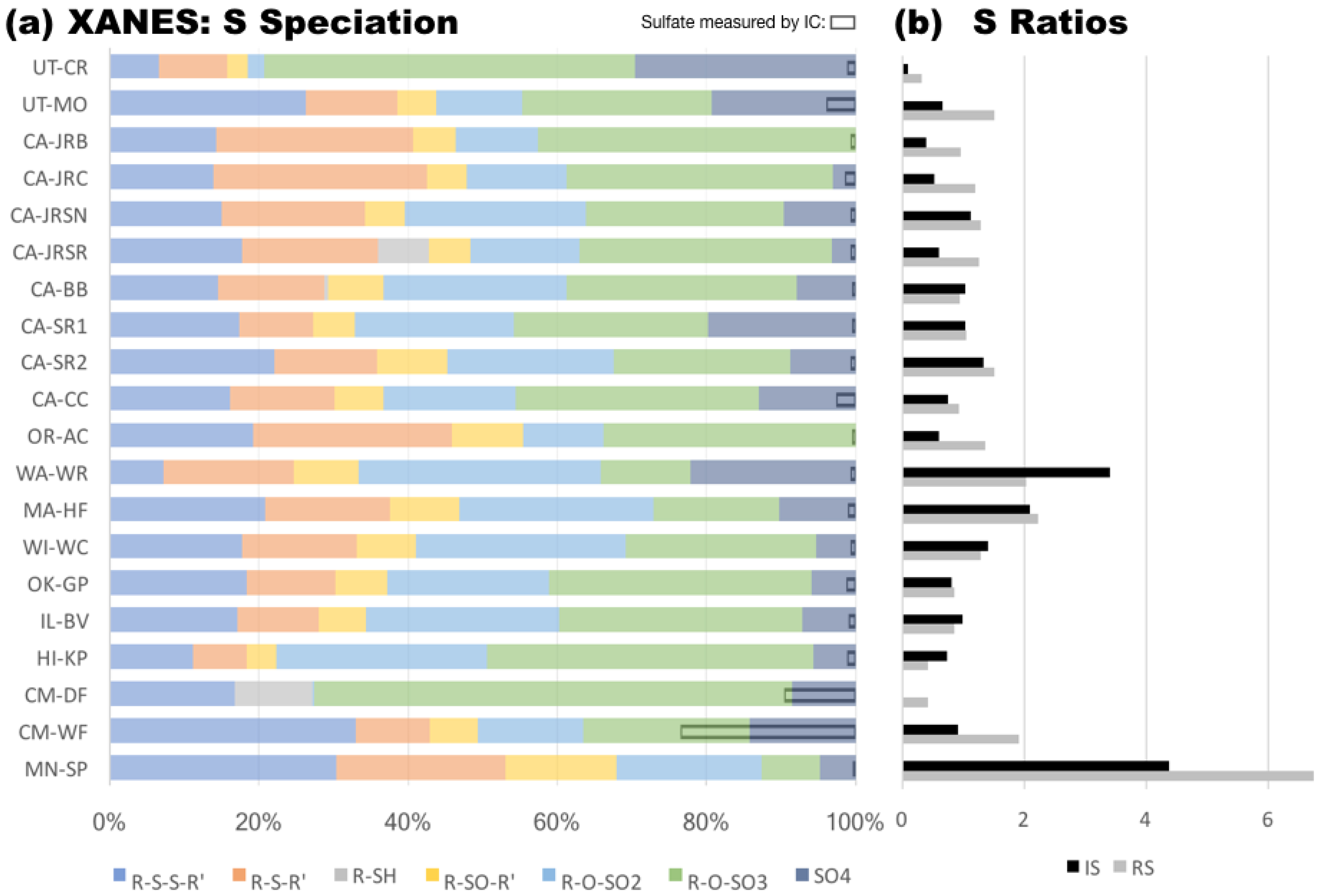
3.4. Soil Sulfur Speciation
3.5. Sulfur Cycling in Soil Microbial Communities
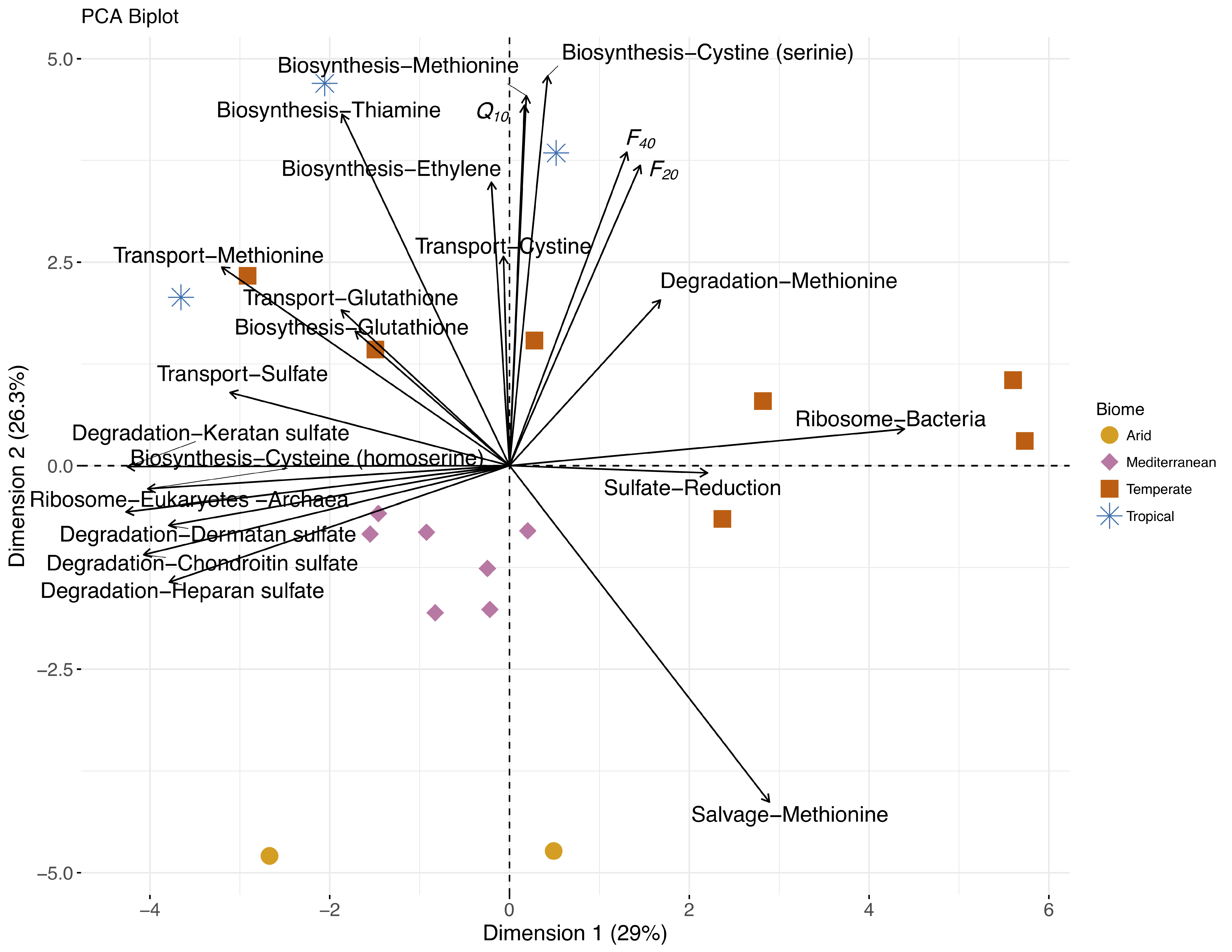
3.6. Multivariate Analysis of Soil Factors Contributing to OCS Production
3.6.1. PLSR Models of Factors Driving F and Q10
3.6.2. Integrated Analysis of Microbial and Chemical Factors Driving OCS Production
4. Discussion
4.1. Ubiquitous OCS Production in Soils
4.2. Mechanisms of OCS Production in Soils
4.2.1. OCS Production from Thermal and Photo Degradation
4.2.2. Direct Microbial OCS Production
4.2.3. Precursors for OCS Production in Soils
4.3. Proposed OCS Production Mechanism of Coupled Biotic-Abiotic OCS Production from S-Containing Amino Acids
5. Conclusions
5.1. Future Research Directions
5.2. Outlook
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site Name | JGI Project ID | Genome Size (bp) | Assembled scnC | Assembled scnC Million Genes−1 | ||
|---|---|---|---|---|---|---|
| BLAST | Pfam | BLAST | Pfam | |||
| CA-CC | 1106757 | 681,529,179 | 44 | 64 | 40 | 28 |
| WI-WC | 1106758 | 671,257,491 | 4 | 82 | 52 | 3 |
| OK-GP | 1106759 | 587,647,789 | 21 | 104 | 73 | 15 |
| CA-SR2 | 1106756 | 521,748,598 | 91 | 82 | 64 | 71 |
| CA-BB | 1106760 | 303,323,377 | 33 | 38 | 51 | 44 |
| CA-JRSN | 1106761 | 245,908,234 | 1 | 40 | 65 | 2 1 |
| HI-KP | 1106762 | 244,931,251 | 10 | 49 | 81 | 17 |
| CM-DF | 1106763 | 146,941,622 | 11 | 12 | 33 | 30 |
| CA-SR1 | 1106764 | 88,712,635 | 5 | 16 | 72 1 | 22 1 |
| IL-BV | 1106755 | 65,589,854 | 43 | 1 | 6 1 | 273 1 |
References
- Kesselmeier, J.; Teusch, N.; Kuhn, U. Controlling variables for the uptake of atmospheric carbonyl sulfide by soil. Biogeochemistry 1999, 104, 11577–11584. [Google Scholar] [CrossRef]
- Whelan, M.E.; Lennartz, S.T.; Gimeno, T.E.; Wehr, R.; Wohlfahrt, G.; Wang, Y.; Kooijmans, L.M.J.; Hilton, T.W.; Belviso, S.; Peylin, P.; et al. Reviews and Syntheses: Carbonyl Sulfide as a Multi-scale Tracer for Carbon and Water Cycles. Biogeosci. Discuss. 2017, 1–97. [Google Scholar] [CrossRef]
- Commane, R.; Meredith, L.K.; Baker, I.T.; Berry, J.A.; Munger, J.W.; Montzka, S.A.; Templer, P.H.; Juice, S.M.; Zahniser, M.S.; Wofsy, S.C. Seasonal fluxes of carbonyl sulfide in a midlatitude forest. Proc. Natl. Acad. Sci. USA 2015, 112, 14162–14167. [Google Scholar] [CrossRef] [PubMed]
- Billesbach, D.P.; Berry, J.A.; Seibt, U.; Maseyk, K.; Torn, M.S.; Fischer, M.L.; Abu-Naser, M.; Campbell, J.E. Growing season eddy covariance measurements of carbonyl sulfide and CO2 fluxes: COS and CO2 relationships in Southern Great Plains winter wheat. Agric. For. Meteorol. 2014, 184, 48–55. [Google Scholar] [CrossRef]
- Kitz, F.; Gerdel, K.; Hammerle, A.; Laterza, T.; Spielmann, F.M.; Wohlfahrt, G. In situ soil COS exchange of a temperate mountain grassland under simulated drought. Oecologia 2017, 183, 851–860. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Maseyk, K.; Lett, C.; Seibt, U. Litter dominates surface fluxes of carbonyl sulfide in a Californian oak woodland. J. Geophys. Res. Biogeosci. 2016, 121, 438–450. [Google Scholar] [CrossRef]
- Maseyk, K.; Berry, J.A.; Billesbach, D.; Campbell, J.E.; Torn, M.S.; Zahniser, M.; Seibt, U. Sources and sinks of carbonyl sulfide in an agricultural field in the Southern Great Plains. Proc. Natl. Acad. Sci. USA 2014, 111, 9064–9069. [Google Scholar] [CrossRef] [PubMed]
- Whelan, M.E.; Hilton, T.W.; Berry, J.A.; Berkelhammer, M.; Desai, A.R.; Campbell, J.E. Carbonyl sulfide exchange in soils for better estimates of ecosystem carbon uptake. Atmos. Chem. Phys. 2016, 16, 3711–3726. [Google Scholar] [CrossRef]
- Bunk, R.; Behrendt, T.; Yi, Z.; Andreae, M.O.; Kesselmeier, J. Exchange of carbonyl sulfide (OCS) between soils and atmosphere under various CO2 concentrations. J. Geophys. Res. Biogeosci. 2017, 122, 1343–1358. [Google Scholar] [CrossRef]
- Kaisermann, A.; Ogée, J.; Sauze, J.; Wohl, S.; Jones, S.P.; Gutierrez, A.; Wingate, L. Disentangling the rates of carbonyl sulphide (COS) production and consumption and their dependency with soil properties across biomes and land use types. Atmos. Chem. Phys. Discuss. 2018, 1–27. [Google Scholar] [CrossRef]
- Whelan, M.E.; Rhew, R.C. Carbonyl sulfide produced by abiotic thermal and photodegradation of soil organic matter from wheat field substrate. J. Geophys. Res. Biogeosci. 2015, 120, 54–62. [Google Scholar] [CrossRef]
- Fried, A.; Klinger, L.F.; Erickson, D.J. Atmospheric carbonyl sulfide exchange in bog microcosms. Geophys. Res. Lett. 1993, 20, 129–132. [Google Scholar] [CrossRef]
- Gimeno, T.E.; Ogée, J.; Royles, J.; Gibon, Y.; West, J.B.; Burlett, R.; Jones, S.P.; Sauze, J.; Wohl, S.; Benard, C.; et al. Bryophyte gas-exchange dynamics along varying hydration status reveal a significant carbonyl sulphide (COS) sink in the dark and COS source in the light. New Phytol. 2017, 215, 965–976. [Google Scholar] [CrossRef] [PubMed]
- Berry, J.; Wolf, A.; Campbell, J.E.; Baker, I.; Blake, N.; Blake, D.; Denning, A.S.; Kawa, S.R.; Montzka, S.A.; Seibt, U.; et al. A coupled model of the global cycles of carbonyl sulfide and CO2: A possible new window on the carbon cycle. J. Geophys. Res. Biogeosci. 2013, 118, 842–852. [Google Scholar] [CrossRef]
- Sauze, J.; Ogée, J.; Maron, P.A.; Crouzet, O.; Nowak, V.; Wohl, S.; Kaisermann, A.; Jones, S.P.; Wingate, L. The interaction of soil phototrophs and fungi with pH and their impact on soil CO2, CO18O and OCS exchange. Soil Biol. Biochem. 2017, 115, 371–382. [Google Scholar] [CrossRef] [PubMed]
- Kato, H.; Saito, M.; Nagahata, Y.; Katayama, Y. Degradation of ambient carbonyl sulfide by Mycobacterium spp. in soil. Microbiology 2008, 154, 249–255. [Google Scholar] [CrossRef] [PubMed]
- Masaki, Y.; Ozawa, R.; Kageyama, K.; Katayama, Y. Degradation and emission of carbonyl sulfide, an atmospheric trace gas, by fungi isolated from forest soil. FEMS Microbiol. Lett. 2016, 363. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, T.; Kato, H.; Higashide, M.; Nishimiya, M.; Katayama, Y. Degradation of carbonyl sulfide by Actinomycetes and detection of clade D of β-class carbonic anhydrase. FEMS Microbiol. Lett. 2016, 363, fnw223. [Google Scholar] [CrossRef] [PubMed]
- Smith, N.A.; Kelly, D.P. Oxidation of Carbon Disulphide as the Sole Source of Energy for the Autotrophic Growth of Thiobacillus thioparus Strain TK-m. Microbiology 1988, 134, 3041–3048. [Google Scholar] [CrossRef]
- Sorokin, D.Y.; Abbas, B.; van Zessen, E.; Muyzer, G. Isolation and characterization of an obligately chemolithoautotrophic Halothiobacillus strain capable of growth on thiocyanate as an energy source. FEMS Microbiol. Lett. 2014, 354, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, T.; Noguchi, K.; Saito, M.; Nagahata, Y.; Kato, H.; Ohtaki, A.; Nakayama, H.; Dohmae, N.; Matsushita, Y.; Odaka, M.; et al. Carbonyl Sulfide Hydrolase from Thiobacillus thioparus Strain THI115 Is One of the β-Carbonic Anhydrase Family Enzymes. J. Am. Chem. Soc. 2013, 135, 3818–3825. [Google Scholar] [CrossRef] [PubMed]
- Bezsudnova, E.Y.; Sorokin, D.Y.; Tikhonova, T.V.; Popov, V.O. Thiocyanate hydrolase, the primary enzyme initiating thiocyanate degradation in the novel obligately chemolithoautotrophic halophilic sulfur-oxidizing bacterium Thiohalophilus thiocyanoxidans. Biochim. Biophys. Acta Proteins Proteom. 2007, 1774, 1563–1570. [Google Scholar] [CrossRef] [PubMed]
- Hussain, A.; Ogawa, T.; Saito, M.; Sekine, T.; Nameki, M.; Matsushita, Y.; Hayashi, T.; Katayama, Y. Cloning and expression of a gene encoding a novel thermostable thiocyanate-degrading enzyme from a mesophilic alphaproteobacteria strain THI201. Microbiology 2013, 159, 2294–2302. [Google Scholar] [CrossRef] [PubMed]
- Katayama, Y.; Narahara, Y.; Inoue, Y.; Amano, F.; Kanagawa, T.; Kuraishi, H. A thiocyanate hydrolase of Thiobacillus thioparus. A novel enzyme catalyzing the formation of carbonyl sulfide from thiocyanate. J. Biol. Chem. 1992, 267, 9170–9175. [Google Scholar] [PubMed]
- Yamasaki, M.; Matsushita, Y.; Nyunoya, H.; Katayama, Y.; Namura, M. Genetic and Immunochemical Characterization of Thiocyanate-Degrading Bacteria in Lake Water Genetic and Immunochemical Characterization of Thiocyanate-Degrading Bacteria in Lake Water. Appl. Environ. Microbiol. 2002, 68, 942–946. [Google Scholar] [CrossRef] [PubMed]
- Welte, C.U.; Rosengarten, J.F.; de Graaf, R.M.; Jetten, M.S.M. SaxA-Mediated Isothiocyanate Metabolism in Phytopathogenic Pectobacteria. Appl. Environ. Microbiol. 2016, 82, 2372–2379. [Google Scholar] [CrossRef] [PubMed]
- Churka Blum, S.; Lehmann, J.; Solomon, D.; Caires, E.F.; Alleoni, L.R.F. Sulfur forms in organic substrates affecting S mineralization in soil. Geoderma 2013, 200–201, 156–164. [Google Scholar] [CrossRef]
- Kertesz, M.A.; Mirleau, P. The role of soil microbes in plant sulphur nutrition. J. Exp. Bot. 2004, 55, 1939–1945. [Google Scholar] [CrossRef] [PubMed]
- Minami, K.; Fukushi, S. Volatilization of carbonyl sulfide from paddy soils treated with sulfur-containing substances. Soil Sci. Plant Nutr. 1981, 27, 339–345. [Google Scholar] [CrossRef]
- Lehmann, S.; Conrad, R. Characteristics of Turnover of Carbonyl Sulfide in 4 Different Soils. J. Atmos. Chem. 1996, 23, 193–207. [Google Scholar] [CrossRef]
- Hanschen, F.S.; Lamy, E.; Schreiner, M.; Rohn, S. Reactivity and Stability of Glucosinolates and Their Breakdown Products in Foods. Angew. Chem. Int. Ed. 2014, 53, 11430–11450. [Google Scholar] [CrossRef] [PubMed]
- Steiger, A.K.; Zhao, Y.; Pluth, M.D. Emerging Roles of Carbonyl Sulfide in Chemical Biology: Sulfide Transporter or Gasotransmitter? Antioxid. Redox Signal. 2017. [Google Scholar] [CrossRef] [PubMed]
- Ferek, R.J.; Andreae, M.O. Photochemical production of carbonyl sulphide in marine surface waters. Nature 1984, 307, 148–150. [Google Scholar] [CrossRef]
- Flöck, O.R.; Andreae, M.O.; Dräger, M. Environmentally relevant precursors of carbonyl sulfide in aquatic systems. Mar. Chem. 1997, 59, 71–85. [Google Scholar] [CrossRef]
- Kelly, D.P.; Wood, A.P.; Jordan, S.L.; Padden, A.N.; Gorlenko, V.M.; Dubinina, G.A. Biological production and consumption of gaseous organic sulphur compounds. Biochem. Soc. Trans. 1994, 22, 1011–1015. [Google Scholar] [CrossRef] [PubMed]
- Zepp, R.G.; Andreae, M.O. Factors affecting the photochemical production of carbonyl sulfide in seawater. Geophys. Res. Lett. 1994, 21, 2813–2816. [Google Scholar] [CrossRef]
- Du, Q.; Mu, Y.; Zhang, C.; Liu, J.; Zhang, Y.; Liu, C. Photochemical production of carbonyl sulfide, carbon disulfide and dimethyl sulfide in a lake water. J. Environ. Sci. 2017, 51, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Geng, C.; Mu, Y. Carbonyl sulfide and dimethyl sulfide exchange between trees and the atmosphere. Atmos. Environ. 2006, 40, 1373–1383. [Google Scholar] [CrossRef]
- Zhang, L.; Walsh, R.S.; Cutter, G.A. Estuarine cycling of carbonyl sulfide: Production and sea–air flux. Mar. Chem. 1998, 61, 127–142. [Google Scholar] [CrossRef]
- Pos, W.H.; Riemer, D.D.; Zika, R.G. Carbonyl sulfide (OCS) and carbon monoxide (CO) in natural waters: Evidence of a coupled production pathway. Mar. Chem. 1998, 62, 89–101. [Google Scholar] [CrossRef]
- Meredith, L.K.; Ogée, J.; Boye, K.; Singer, E.; Wingate, L.; von Sperber, C.; Sengupta, A.; Whelan, M.; Pang, E.; Keiluweit, M.; et al. Soil exchange rates of COS and CO18O shift with the diversity of microbial communities and their carbonic anhydrase enzymes. ISME J. 2018, in press. [Google Scholar]
- Tecon, R.; Or, D. Biophysical processes supporting the diversity of microbial life in soil. FEMS Microbiol. Rev. 2017, 41, 599–623. [Google Scholar] [CrossRef] [PubMed]
- Hobbie, S. Chloroform Fumigation Direct Extraction (CFDE) Protocol for Microbial Biomass Carbon and Nitrogen. Allison Lab Protoc. 2008, 4. [Google Scholar]
- Brookes, P.C.; Landman, A.; Pruden, G.; Jenkinson, D.S. Chloroform fumigation and the release of soil nitrogen: A rapid direct extraction method to measure microbial biomass nitrogen in soil. Soil Biol. Biochem. 1985, 17, 837–842. [Google Scholar] [CrossRef]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. Microbial biomass measurements in forest soils: The use of the chloroform fumigation-incubation method in strongly acid soils. Soil Biol. Biochem. 1987, 19, 697–702. [Google Scholar] [CrossRef]
- Kooijmans, L.M.J.; Uitslag, N.A.M.; Zahniser, M.S.; Nelson, D.D.; Montzka, S.A.; Chen, H. Continuous and high precision atmospheric concentration measurements of COS, CO2, CO and H2O using a quantum cascade laser spectrometer (QCLS). Atmos. Meas. Tech. Discuss. 2016, 9, 5293–5314. [Google Scholar] [CrossRef]
- Ogée, J.; Sauze, J.; Kesselmeier, J.; Genty, B.; Van Diest, H.; Launois, T.; Wingate, L. A new mechanistic framework to predict OCS fluxes from soils. Biogeosciences 2016, 13, 2221–2240. [Google Scholar] [CrossRef]
- Fierer, N.; Schimel, J.P. Effects of drying-rewetting frequency on soil carbon and nitrogen transformations. Soil Biol. Biochem. 2002, 34, 777–787. [Google Scholar] [CrossRef]
- Di Stefano, C.; Ferro, V.; Mirabile, S. Comparison between grain-size analyses using laser diffraction and sedimentation methods. Biosyst. Eng. 2010, 106, 205–215. [Google Scholar] [CrossRef]
- Harris, I.; Jones, P.D.; Osborn, T.J.; Lister, D.H. Updated high-resolution grids of monthly climatic observations—The CRU TS3.10 Dataset. Int. J. Climatol. 2014, 34, 623–642. [Google Scholar] [CrossRef]
- Jalilehvand, F. Sulfur: Not a “silent” element any more. Chem. Soc. Rev. 2006, 35, 1256–1268. [Google Scholar] [CrossRef] [PubMed]
- Ravel, B.; Newville, M. ATHENA, ARTEMIS, HEPHAESTUS: Data analysis for X-ray absorption spectroscopy using IFEFFIT. J. Synchrotron Radiat. 2005, 12, 537–541. [Google Scholar] [CrossRef] [PubMed]
- Beck, T.; Joergensen, R.G.; Kandeler, E.; Makeshin, E.; Nuss, E.; Oberholzer, H.R.; Scheu, S. An inter-laboratory comparison of ten different ways of measuring soil microbial biomass C. Soil Biol. Biochem. 1997, 29, 1023–1032. [Google Scholar] [CrossRef]
- Ward, T. BugBase predicts organism-level microbiome phenotypes. bioRxiv 2017, 1–36. [Google Scholar] [CrossRef]
- Kanehisa, M.; Furumichi, M.; Tanabe, M.; Sato, Y.; Morishima, K. KEGG: New perspectives on genomes, pathways, diseases and drugs. Nucleic Acids Res. 2017, 45, D353–D361. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Sato, Y.; Kawashima, M.; Furumichi, M.; Tanabe, M. KEGG as a reference resource for gene and protein annotation. Nucleic Acids Res. 2016, 44, D457–D462. [Google Scholar] [CrossRef] [PubMed]
- Ogata, H.; Goto, S.; Sato, K.; Fujibuchi, W.; Bono, H.; Kanehisa, M. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 1999, 27, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Chen, I.M.A.; Markowitz, V.M.; Chu, K.; Palaniappan, K.; Szeto, E.; Pillay, M.; Ratner, A.; Huang, J.; Andersen, E.; Huntemann, M.; et al. IMG/M: Integrated genome and metagenome comparative data analysis system. Nucleic Acids Res. 2017, 45, D507–D516. [Google Scholar] [CrossRef] [PubMed]
- Wattam, A.R.; Abraham, D.; Dalay, O.; Disz, T.L.; Driscoll, T.; Gabbard, J.L.; Gillespie, J.J.; Gough, R.; Hix, D.; Kenyon, R.; et al. PATRIC, the bacterial bioinformatics database and analysis resource. Nucleic Acids Res. 2014, 42, D581–D591. [Google Scholar] [CrossRef] [PubMed]
- Langille, M.G.I.; Zaneveld, J.; Caporaso, J.G.; McDonald, D.; Knights, D.; Reyes, J.A.; Clemente, J.C.; Burkepile, D.E.; Vega Thurber, R.L.; Knight, R.; et al. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat. Biotechnol. 2013, 31, 814–821. [Google Scholar] [CrossRef] [PubMed]
- Katayama, Y.; Matsushita, Y.; Kaneko, M.; Kondo, M.; Mizuno, T.; Nyunoya, H. Cloning of genes coding for the three subunits of thiocyanate hydrolase of Thiobacillus thioparus THI 115 and their evolutionary relationships to nitrile hydratase. J. Bacteriol. 1998, 180, 2583–2589. [Google Scholar] [PubMed]
- Abdi, H. Partial least squares regression and projection on latent structure regression (PLS Regression). Wiley Interdiscip. Rev. Comput. Stat. 2010, 2, 97–106. [Google Scholar] [CrossRef]
- Carrascal, L.M.; Galván, I.; Gordo, O. Partial least squares regression as an alternative to current regression methods used in ecology. Oikos 2009, 118, 681–690. [Google Scholar] [CrossRef]
- Aitchison, J. The Statistical Analysis of Compositional Data, Monographs on Statistics and Applied Probability; Chapman and Hall: London, UK, 1986. [Google Scholar]
- Wehr, R.; Saleska, S.R. The long-solved problem of the best-fit straight line: Application to isotopic mixing lines. Biogeosciences 2017, 14, 17–29. [Google Scholar] [CrossRef]
- Manceau, A.; Nagy, K.L. Quantitative analysis of sulfur functional groups in natural organic matter by XANES spectroscopy. Geochim. Cosmochim. Acta 2012, 99, 206–223. [Google Scholar] [CrossRef]
- Ma, J.; Wang, Z.-Y.; Stevenson, B.A.; Zheng, X.-J.; Li, Y. An inorganic CO2 diffusion and dissolution process explains negative CO2 fluxes in saline/alkaline soils. Sci. Rep. 2013, 3, 2025. [Google Scholar] [CrossRef] [PubMed]
- Saito, M.; Honna, T.; Kanagawa, T.; Katayama, Y. Microbial Degradation of Carbonyl Sulfide in Soils. Microbes Environ. 2002, 17, 32–38. [Google Scholar] [CrossRef]
- Baker, N.R.; Allison, S.D.; Frey, S.D. Ultraviolet photodegradation facilitates microbial litter decomposition in a Mediterranean climate. Ecology 2015, 96, 1994–2003. [Google Scholar] [CrossRef] [PubMed]
- Thompson, L.R.; Sanders, J.G.; McDonald, D.; Amir, A.; Ladau, J.; Locey, K.J.; Prill, R.J.; Tripathi, A.; Gibbons, S.M.; Ackermann, G.; et al. A communal catalogue reveals Earth’s multiscale microbial diversity. Nature 2017, 551, 457–463. [Google Scholar] [CrossRef] [PubMed]
- Katayama, Y.; Kanagawa, T.; Kuraishi, H. Emission of Carbonyl Sulfide by Thiobacillus-Thioparus Grown with Thiocyanate in Pure and Mixed Cultures. FEMS Microbiol. Lett. 1993, 114, 223–228. [Google Scholar] [CrossRef]
- Smeulders, M.J.; Pol, A.; Venselaar, H.; Barends, T.R.M.; Hermans, J.; Jetten, M.S.M.; Op den Camp, H.J.M. Bacterial CS2 hydrolases from Acidithiobacillus thiooxidans strains are homologous to the archaeal catenane CS2 hydrolase. J. Bacteriol. 2013, 195, 4046–4056. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Lerdau, M.; He, Y. Widespread production of nonmicrobial greenhouse gases in soils. Glob. Chang. Biol. 2017, 23, 4472–4482. [Google Scholar] [CrossRef] [PubMed]
- Minami, K.; Fukushi, S. Detection of carbonyl sulfide among gases produced by the decomposition of cystine in paddy soils. Soil Sci. Plant Nutr. 1981, 27, 105–109. [Google Scholar] [CrossRef]
- Morra, M.J.; Dick, W.A. Production of Thiocysteine (Sulfide) in Cysteine Amended Soils. Soil Sci. Soc. Am. J. 1985, 49, 882–886. [Google Scholar] [CrossRef]
- Sekowska, A.; Dénervaud, V.; Ashida, H.; Michoud, K.; Haas, D.; Yokota, A.; Danchin, A. Bacterial variations on the methionine salvage pathway. BMC Microbiol. 2004, 4, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Freney, J.R. Aerobic transformation of cysteine to sulphate in soil. Nature 1958, 182, 1318–1319. [Google Scholar] [CrossRef]
- Devai, I.; DeLaune, R.D. Formation of volatile sulfur compounds in salt marsh sediment as influenced by soil redox condition. Org. Geochem. 1995, 23, 283–287. [Google Scholar] [CrossRef]
- Delgado-Baquerizo, M.; Oliverio, A.M.; Brewer, T.E.; Benavent-González, A.; Eldridge, D.J.; Bardgett, R.D.; Maestre, F.T.; Singh, B.K.; Fierer, N. A global atlas of the dominant bacteria found in soil. Science 2018, 359, 320–325. [Google Scholar] [CrossRef] [PubMed]





| Compound | Classification | Non-Photochemical Production | Photo-Chemical Production (UV) | References |
|---|---|---|---|---|
| Cysteine (CYS) | Thiol, R-SH | N | Y | [33,34,36,37,40] |
| Methionine (MET) | Thioether (organic sulfide) R-S-R’ | N | uncertain | [33,34,37] |
| Glutathione (GSH) | Thiol, R-SH | Y | Y | [33,34] |
| 3-Mercaptopropionic acid (3-MPA) | Thiol, R-SH | N | Y | [34] |
| Methyl mercaptan (MeSH) | Thiol, R-SH | N | Y | [34] |
| Sulfide | Sulfide, HS− | - | Y | [34] |
| Dimeric disulfide (Na-GSSG) | Organic disulfide, R-S-S-R’ | N | N | [34] |
| Methanesulfonic acid (MSA) | Sulfonate, R-O-SO2 | N | N | [34] |
| Soil Collection | Survey 1 | Survey 2 |
| Sites | 20 | 38 |
| Region | United States, Cambodia | Europe |
| Biomes | Arid, Mediterranean, Boreal, Temperate, Tropical | Arid, Mediterranean, Boreal, Temperate |
| Land use | Cropland, Desert, Grassland, Deciduous Forest, Evergreen Forest, Peatland | Cropland, Orchard, Grassland, Deciduous Forest, Evergreen Forest, Peatland |
| Sampling depth | 0 to 10 cm (after removing litter) | 0 to 10 cm (after removing litter) |
| Samples | 3 within a 1-m sampling radius | 3–5 within 5-m radius |
| Soil Processing | Survey 1 | Survey 2 |
| Sampling depth | Replicates kept separate | Replicates homogenized |
| Sieve mesh size | 2 mm (Humboldt Mfg., Elgin, IL, USA) | 5 mm |
| Pre-treatment | Air-dried 3 days, wet to 30% WHC, incubated 7 days at 22.5 °C in the dark | Soils at field moisture untreated |
| Final treatment | Air-dried for a median of 45 days | Air-dried for 7–14 days |
| Flux Measurements | Survey 1 | Survey 2 |
| Soil amount | 80 g dry soil | 350–400 g dry soil |
| Measurement chamber | 1-L PFA chambers (100-1000-01, Savillex, Eden Prairie, MN, USA). Surface area of 0.0078 m2 | 0.825 L glass jars with customized glass lid with stainless steel and PTFE ports. Surface area of 0.0062 m2 |
| Acclimation | 24 h dark at 22.5 °C | 2–3 days under 12 h dark/12 h light photoperiod at 17 °C |
| Treatments | Temperature ramp (11.5 °C to 37.5 °C; dark only) | Light conditioning and temperature artifact (dark at 17 °C; light at 23 °C) |
| Inlet air source | Room air passed through buffer volume (2 L PFA chamber) | Scrubbed ambient air with CO2 and OCS added to approximately 420 ppm CO2 and 500 ppt OCS. |
| Flow rate | 0.3 L min−1 | 0.25 L min−1 |
| Temperature control | Water bath [8] | Customized climate-control chamber (MD1400, Snijders, Tillburg, The Netherlands) |
| Dynamic flow sequence | Inlet (10 min), N2 tank background (10 min), and outlet (40 min) | Each component 2 min |
| Time averaged | Last 4 min inlet, 4 min for N2, and 8 min for outlet measurements. | Last 20 s for all measurements. |
| Soil Characterization | Survey 1 | Survey 2 |
| Measured properties (pre-treatment) | WHC, microbial biomass, microbial community composition (DNA), metatranscriptomes (RNA) | WHC, soil moisture |
| Measured properties (final treatment) | OCS and CO2 fluxes, BD, C, N, pH, texture, soil moisture, S, other elements, SO4, XANES | OCS and CO2 fluxes, BD, C, N, pH, texture, redox potential |
| pH method | 1:2.5 soil-water ratio | 1:5 soil-water ratio |
| Texture method | Multi-wavelength laser diffraction particle analyzer (LS 13 320 MW, Beckman Coulter, Brea, CA, USA) | Sedimentation method (INRA method SOL-0302). |
| C and N method | Elemental analyzer (NA-1500, Carlo-Erba, Milan, Italy) | Gas chromatography and catharometer (corrected for CaCO3; INRA methods NF-ISO-13878, NF-ISO-10694) |
| Microbial biomass method | Chloroform fumigation-potassium sulfate extraction method [43] | Chloroform fumigation-potassium sulfate extraction method [44,45]. |
| Survey 1 | |||||||||||
| Soil ID | Latitude | Longitude | Biome | Land Use | pH | C/N | Clay (%) | Silt (%) | Sand (%) | MAT (°C) | MAP (mm) |
| UT-CR | 38.68 | −109.42 | Arid | Desert | 8.8 | 88 | 14 | 34 | 52 | 9.7 | 295 |
| UT-MO | 38.87 | −109.81 | Arid | Desert | 9.4 | 172 | 36 | 58 | 6 | 10.9 | 225 |
| CA-JRB | 37.4 | −122.23 | Mediterranean | Grassland | 7.5 | 12 | 11 | 37 | 52 | 14.6 | 618 |
| CA-JRC | 37.41 | −122.23 | Mediterranean | Grassland | 7.3 | 11 | 11 | 39 | 50 | 14.6 | 618 |
| CA-JRSN | 37.41 | −122.23 | Mediterranean | Grassland | 6.5 | 11 | 17 | 48 | 35 | 14.6 | 618 |
| CA-JRSR | 37.41 | −122.23 | Mediterranean | Grassland | 7.4 | 12 | 10 | 43 | 47 | 14.6 | 618 |
| CA-SR1 | 34.09 | −118.66 | Mediterranean | Evergreen Forest | 7.3 | 21 | 13 | 36 | 51 | 14.8 | 450 |
| CA-SR2 | 34.09 | −118.66 | Mediterranean | Evergreen Forest | 7.6 | 17 | 8 | 33 | 59 | 14.8 | 450 |
| CA-CC | 37.43 | −122.18 | Mediterranean | Cropland | 8.2 | 12 | 13 | 30 | 57 | 14.6 | 618 |
| CA-BB | 37.19 | −122.22 | Temperate | Evergreen Forest | 6.4 | 25 | 10 | 36 | 54 | 14.6 | 618 |
| OR-AC | 42.18 | −122.8 | Temperate | Evergreen Forest | 6.5 | 32 | 6 | 25 | 69 | 10.5 | 676 |
| WA-WR | 45.82 | −121.95 | Temperate | Evergreen Forest | 5.3 | 33 | 1 | 4 | 95 | 8.4 | 1850 |
| MA-HF | 42.54 | −72.17 | Temperate | Deciduous Forest | 4.3 | 21 | 4 | 26 | 70 | 8.8 | 1167 |
| WI-WC | 45.81 | −90.08 | Temperate | Deciduous Forest | 5.8 | 15 | 3 | 15 | 82 | 4.7 | 812 |
| IL-BV | 40.01 | −88.29 | Temperate | Cropland | 5.8 | 11 | 7 | 37 | 56 | 11.4 | 1013 |
| OK-GP | 36.61 | −97.49 | Temperate | Cropland | 4.8 | 10 | 13 | 62 | 25 | 16.0 | 972 |
| HI-KP | 20.15 | −155.83 | Tropical | Grassland | 6.6 | 10 | 7 | 26 | 67 | 20.3 | 2680 |
| CM-DF | 11.51 | 105.01 | Tropical | Cropland | 5.5 | 9 | 28 | 62 | 10 | 28.2 | 1453 |
| CM-WF | 11.51 | 105.01 | Tropical | Cropland | 4.6 | 10 | 25 | 59 | 16 | 28.2 | 1453 |
| MN-SP | 47.51 | −93.45 | Boreal | Peatland | 4 | 33 | n.d. | n.d. | n.d. | 4.1 | 725 |
| Survey 2 | |||||||||||
| Soil ID | Latitude | Longitude | Biome | Land Use | pH | C/N | Clay (%) | Silt (%) | Sand (%) | MAT (°C) | MAP (mm) |
| SP-Amo | 36.83 | −2.25 | Arid | Grassland | 8.6 | 9 | 15 | 35 | 50 | 18.1 | 291 |
| SP-Bal | 39.94 | −2.03 | Arid | Grassland | 8.4 | 19 | 18 | 28 | 54 | 13.8 | 427 |
| IS-Yat | 31.35 | 35.05 | Arid | Evergreen Forest | 8.1 | 30 | 29 | 49 | 22 | 21.0 | 217 |
| IS-Reh | 31.91 | 34.81 | Mediterranean | Orchard | 7.8 | 18 | 15 | 7 | 78 | 20.3 | 485 |
| SP-Ube_NOVeg | 37.92 | −3.23 | Mediterranean | Orchard | 8.4 | 42 | 55 | 40 | 5 | 15.7 | 422 |
| SP-Ube_Veg | 37.91 | −3.23 | Mediterranean | Orchard | 8.6 | 13 | 23 | 35 | 42 | 15.7 | 422 |
| FR-Pue | 43.74 | 3.6 | Mediterranean | Evergreen Forest | 6.9 | 18 | 42 | 32 | 26 | 13.8 | 755 |
| PT-Cor | 39.14 | −8.33 | Mediterranean | Evergreen Forest | 5.7 | 18 | 4 | 16 | 80 | 16.7 | 811 |
| PT-Mit-amb | 38.54 | −8 | Mediterranean | Evergreen Forest | 5.5 | n.d. | n.d. | n.d. | n.d. | 16.7 | 811 |
| PT-Mit-b9 | 38.54 | −8 | Mediterranean | Evergreen Forest | 5.9 | 18 | 4 | 9 | 87 | 16.7 | 811 |
| PT-Mit-mid | 38.54 | −8 | Mediterranean | Evergreen Forest | 6 | n.d. | n.d. | n.d. | n.d. | 16.7 | 811 |
| SP-Cha | 40.65 | 0.21 | Mediterranean | Evergreen Forest | 5.5 | 19 | 13 | 20 | 67 | 15.3 | 490 |
| SP-Peg | 40.38 | 4.19 | Mediterranean | Evergreen Forest | 6.2 | 11 | 3 | 9 | 88 | 16.7 | 603 |
| CH-Cha | 47.21 | 8.41 | Temperate | Grassland | 6.3 | 27 | 28 | 44 | 28 | 9.5 | 1136 |
| CH-Fru | 47.12 | 8.54 | Temperate | Grassland | 4.9 | 10 | 42 | 47 | 11 | 9.2 | 1282 |
| FR-Laq1 | 45.64 | 2.74 | Temperate | Grassland | 4.6 | 11 | 18 | 59 | 23 | 8.2 | 985 |
| FR-Laq2 | 45.64 | 2.74 | Temperate | Grassland | 5.7 | 11 | 21 | 57 | 22 | 8.2 | 985 |
| CH-Dav | 46.81 | 9.86 | Temperate | Evergreen Forest | 4.3 | 25 | 22 | 25 | 53 | 2.6 | 2135 |
| FR-Gra | 44.76 | 0.6 | Temperate | Evergreen Forest | 4.6 | 27 | 4 | 5 | 91 | 13.2 | 794 |
| FR-LeB | 44.72 | 0.77 | Temperate | Evergreen Forest | 4.8 | 25 | 4 | 3 | 93 | 13.2 | 794 |
| CH-Lag | 47.12 | 8.54 | Temperate | Deciduous Forest | 6.3 | 13 | 42 | 43 | 15 | 9.2 | 1282 |
| DE-Hai | 51.08 | 10.45 | Temperate | Deciduous Forest | 6 | 13 | 48 | 49 | 4 | 8.6 | 672 |
| DE-Lei | 51.33 | 10.37 | Temperate | Deciduous Forest | 5.2 | 14 | 19 | 77 | 4 | 8.6 | 672 |
| DK-Sor | 55.49 | 11.64 | Temperate | Deciduous Forest | 4.2 | 19 | 15 | 23 | 62 | 9.0 | 584 |
| FR_Rou | 45.01 | 0.97 | Temperate | Deciduous Forest | 6.5 | n.d. | n.d. | n.d. | n.d. | 12.7 | 830 |
| FR-Hes | 48.67 | 7.07 | Temperate | Deciduous Forest | 5.4 | 14 | 24 | 61 | 15 | 10.3 | 743 |
| FR-Lou | 43.08 | −0.04 | Temperate | Deciduous Forest | 7.9 | 23 | 14 | 38 | 48 | 12.8 | 845 |
| CH-Oe2 | 47.29 | 7.73 | Temperate | Cropland | 7.3 | 10 | 42 | 47 | 11 | 9.1 | 1220 |
| FR_TlsC6 | 43.54 | 1.51 | Temperate | Cropland | 8.6 | 22 | 18 | 35 | 47 | 13.9 | 660 |
| FR_TlsLA3 | 43.53 | 1.5 | Temperate | Cropland | 8.5 | 15 | 28 | 45 | 27 | 13.9 | 660 |
| FR-AucB4 | 43.62 | 0.57 | Temperate | Cropland | 8.4 | 30 | 33 | 49 | 18 | 13.9 | 712 |
| FR-AucLH8 | 43.64 | 0.6 | Temperate | Cropland | 7.8 | 8 | 47 | 35 | 18 | 13.9 | 712 |
| FR-TlsCL | 43.53 | 1.51 | Temperate | Cropland | 5.7 | 8 | 33 | 42 | 25 | 13.9 | 660 |
| SE-Abi | 68.36 | 19.05 | Boreal | Peatland | 4.4 | 39 | n.d. | n.d. | n.d. | −3.1 | 690 |
| SE-Hyl | 56.1 | 13.42 | Boreal | Peatland | 3.8 | 26 | 14 | 35 | 51 | 7.7 | 849 |
| FI-Hyy | 61.85 | 24.3 | Boreal | Evergreen Forest | 4.6 | 36 | 14 | 32 | 54 | 3.9 | 579 |
| FI-Var | 67.76 | 29.62 | Boreal | Evergreen Forest | 5.3 | 31 | 4 | 17 | 79 | −0.6 | 578 |
| SE-Nor | 60.09 | 17.47 | Boreal | Evergreen Forest | 4.4 | 31 | 13 | 26 | 61 | 5.9 | 576 |
| SE-Ros_Cont | 64.17 | 19.75 | Boreal | Evergreen Forest | 5.2 | 41 | 4 | 15 | 81 | 2.0 | 635 |
| SE-Ros_Fert | 64.17 | 19.75 | Boreal | Evergreen Forest | 4.5 | 27 | 4 | 30 | 66 | 2.0 | 635 |
| SE-Sva | 64.17 | 19.78 | Boreal | Evergreen Forest | 4.7 | 47 | 9 | 28 | 63 | 2.0 | 635 |
| Effect | Survey 1 | Survey 2 | ||
|---|---|---|---|---|
| F20 | F40 | Q10 | F20 | |
| ANOVA p-value | ||||
| Biome | <0.001 | <0.001 | <0.001 | <0.001 |
| Land use | 2 | 2 | 2 | <0.001 |
| Lsmeans | (pmol OCS kg−1 min−1) | - | (pmol OCS kg−1 min−1) | |
| Arid | −0.46 B | 0.44 C | 1.65 C | −2.93 B |
| Mediterranean | 1.73 A | 8.74 B | 2.66 B | 0.38 AB |
| Boreal | 3 | 3 | 3 | 0.59 AB |
| Temperate | 3.44 A | 21.48 A | 2.83 AB | 3.72 A |
| Tropical | 3.37 A | 23.90 A | 3.18 A | 1 |
| Deciduous Forest | 2 | 2 | 2 | −2.02 B |
| Cropland | 2 | 2 | 2 | −1.62 B |
| Evergreen Forest | 2 | 2 | 2 | 0.33 B |
| Orchard | 1 | 1 | 1 | 0.84 AB |
| Grassland | 2 | 2 | 2 | 4.64 A |
| Peatland | 3 | 3 | 3 | 1 |
| Desert | 2 | 2 | 2 | 1 |
| Site | S (%) | C/N | C/S | N/S | P/S | SO4 (IC) (mg/kgS) |
|---|---|---|---|---|---|---|
| UT-CR | 0.03 | 88 | 81 | 1 | 2 | 0.9 |
| UT-MO | 0.05 | 172 | 37 | 0 | 2 | 2.7 |
| CA-JRB | 0.02 | 12 | 67 | 6 | 1 | 0.3 |
| CA-JRC | 0.04 | 11 | 61 | 6 | 2 | 0.4 |
| CA-JRSN | 0.02 | 11 | 64 | 6 | 1 | 0.3 |
| CA-JRSR | 0.02 | 12 | 108 | 9 | 1 | 0.3 |
| CA-BB | 0.04 | 25 | 125 | 5 | 4 | 0.2 |
| CA-SR1 | 0.04 | 21 | 41 | 2 | 3 | 0.2 |
| CA-SR2 | 0.06 | 17 | 63 | 4 | 2 | 0.4 |
| CA-CC | 0.04 | 12 | 34 | 3 | 2 | 1.8 |
| OR-AC | 0.06 | 32 | 105 | 3 | 1 | 0.2 |
| WA-WR | 0.03 | 33 | 141 | 4 | 3 | 4.2 |
| MA-HF | 0.12 | 21 | 70 | 3 | 1 | 3.5 |
| WI-WC | 0.09 | 15 | 53 | 4 | 1 | 0.6 |
| IL-BV | 0.03 | 11 | 73 | 7 | 1 | 0.5 |
| OK-GP | 0.03 | 10 | 37 | 4 | 0 | 0.9 |
| HI-KP | 0.10 | 10 | 50 | 5 | 7 | 1.2 |
| CM-DF | 0.05 | 9 | 22 | 3 | 1 | 12.4 |
| CM-WF | 0.10 | 10 | 23 | 2 | 0 | 43.2 |
| MN-SP | 0.66 | 33 | 64 | 2 | 1 | S 0.2 |
| Survey 1 | Survey 2 | |||||||
|---|---|---|---|---|---|---|---|---|
| FOCS,20C | FOCS,40C | Q10 | FOCS,20C | |||||
| C1 R2 | C2 R2 | C1 R2 | C2 R2 | C1 R2 | C2 R2 | C1 R2 | C2 R2 | |
| 0.51 | 0.07 | 0.55 | 0.07 | 0.74 | 0.15 | 0.30 | 0.04 | |
| Predictors | C1 w | C2 w | C1 w | C2 w | C1 w | C2 w | C1 w | C2 w |
| BD | 2 | 0.45 | 2 | 0.51 | −0.24 | 2 | 1 | 1 |
| pH | −0.49 | 2 | −0.51 | 2 | −0.39 | 2 | −0.30 | −0.29 |
| Clay (clr) | 2 | 2 | 2 | 2 | 2 | 0.26 | 2 | −0.68 |
| Silt (clr) | 2 | 2 | 2 | 2 | 2 | 0.22 | 2 | 2 |
| Sand (clr) | 2 | 2 | 2 | 2 | 2 | −0.25 | 2 | 0.45 |
| GWC | 2 | 2 | 2 | 2 | 0.22 | 2 | 1 | 1 |
| Microbial C | 0.22 | −0.29 | 0.22 | −0.29 | 2 | −0.27 | 1 | 1 |
| Microbial N | 2 | −0.29 | 2 | −0.30 | 2 | −0.28 | 1 | 1 |
| C/N | −0.35 | 2 | −0.30 | 2 | −0.43 | 2 | −0.46 | 2 |
| C/S | 2 | 2 | 2 | 2 | 2 | −0.22 | 1 | 1 |
| C (clr) | 2 | −0.26 | 2 | 2 | 2 | −0.42 | −0.44 | 0.23 |
| N (clr) | 0.34 | 2 | 0.30 | −0.23 | 0.40 | 2 | 0.44 | −0.23 |
| P (clr) | −0.28 | 2 | −0.28 | 2 | 2 | 2 | 1 | 1 |
| K (clr) | 2 | 0.35 | 2 | 0.30 | −0.23 | 0.21 | 1 | 1 |
| ISO4 (mg/kgS) | 0.28 | 0.23 | 0.37 | 0.28 | 2 | 2 | 1 | 1 |
| R-S-S-R’ (clr) | 2 | 0.22 | 2 | 2 | 2 | 0.26 | 1 | 1 |
| R-S-R’ (clr) | −0.24 | −0.40 | −0.27 | −0.38 | 2 | −0.30 | 1 | 1 |
| R-SO-R’ (clr) | 2 | 2 | 2 | 2 | 0.33 | 0.21 | 1 | 1 |
| R-O-SO3 (clr) | −0.21 | 2 | 2 | 2 | 2 | 0.22 | 1 | 1 |
| XSO4 (clr) | 2 | 2 | 2 | 0.24 | 2 | 2 | 1 | 1 |
| Redox | 1 | 1 | 1 | 1 | 1 | 1 | 0.38 | 0.28 |
| FCO2 | 1 | 1 | 1 | 1 | 1 | 1 | 2 | 2 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meredith, L.K.; Boye, K.; Youngerman, C.; Whelan, M.; Ogée, J.; Sauze, J.; Wingate, L. Coupled Biological and Abiotic Mechanisms Driving Carbonyl Sulfide Production in Soils. Soil Syst. 2018, 2, 37. https://doi.org/10.3390/soilsystems2030037
Meredith LK, Boye K, Youngerman C, Whelan M, Ogée J, Sauze J, Wingate L. Coupled Biological and Abiotic Mechanisms Driving Carbonyl Sulfide Production in Soils. Soil Systems. 2018; 2(3):37. https://doi.org/10.3390/soilsystems2030037
Chicago/Turabian StyleMeredith, Laura K., Kristin Boye, Connor Youngerman, Mary Whelan, Jérôme Ogée, Joana Sauze, and Lisa Wingate. 2018. "Coupled Biological and Abiotic Mechanisms Driving Carbonyl Sulfide Production in Soils" Soil Systems 2, no. 3: 37. https://doi.org/10.3390/soilsystems2030037
APA StyleMeredith, L. K., Boye, K., Youngerman, C., Whelan, M., Ogée, J., Sauze, J., & Wingate, L. (2018). Coupled Biological and Abiotic Mechanisms Driving Carbonyl Sulfide Production in Soils. Soil Systems, 2(3), 37. https://doi.org/10.3390/soilsystems2030037