Drying-Wetting Cycles: Effect on Deep Soil Carbon

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
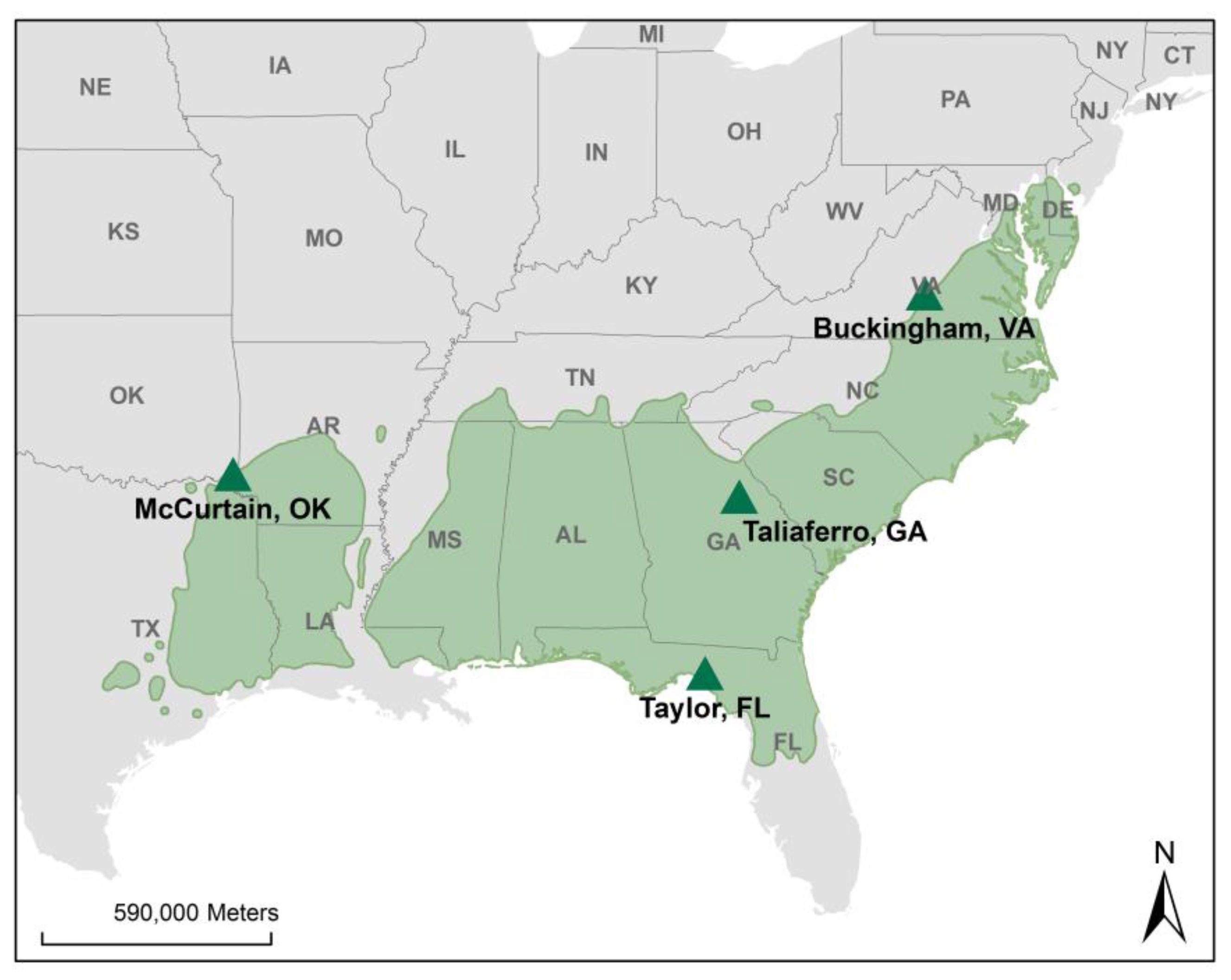
2.1. Site Description
- Oklahoma: the Ruston series is a fine-loamy, siliceous, semiactive, thermic Typic Paleudult. Two profiles (four depth, eight samples) were collected to 100 cm, due to shallow soil solum.
- Virginia: Littlejoe is a fine, mixed, subactive, mesic Typic Hapludult. Due to shallow soil solum, two soil profiles (five depth, 10 samples) were collected to a depth of 150 cm.
- Georgia: the Lloyd series is a fine, kaolinitic, thermic Rhodic Kanhapludult, while the Cecil series is a fine, kaolinitic, thermic Typic Kanhapludult. The Rhodic designation indicates an influence of mafic parental material on soil color. Two soil profiles (eight depth, 16 samples) were collected to 300 cm.
- Florida: Melvina is a sandy, siliceous, thermic Oxyaquic Alorthod; Moriah is loamy, siliceous, superactive, thermic Aquic Arenic Hapludalf; while Lutterloh is loamy, siliceous, subactive, thermic Grossarenic Paleudalf. Due to high water table at the Florida site, two soil profiles (six depth, 12 samples) were collected to 170 cm.
2.2. Soil Physical and Chemical Analysis
2.3. Soil C Fractionation
2.4. Soil Incubations
2.5. Statistical Analysis
3. Results
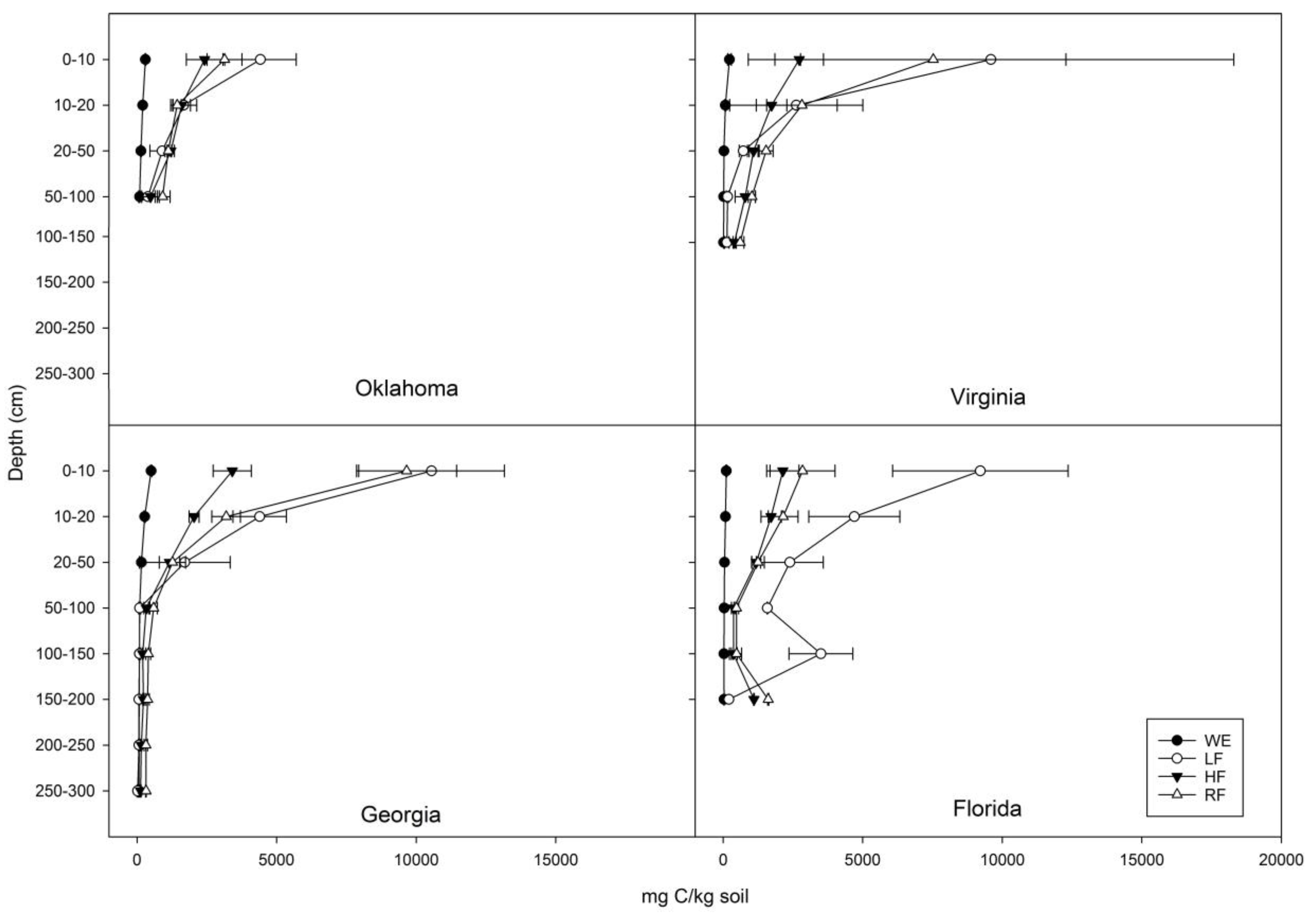
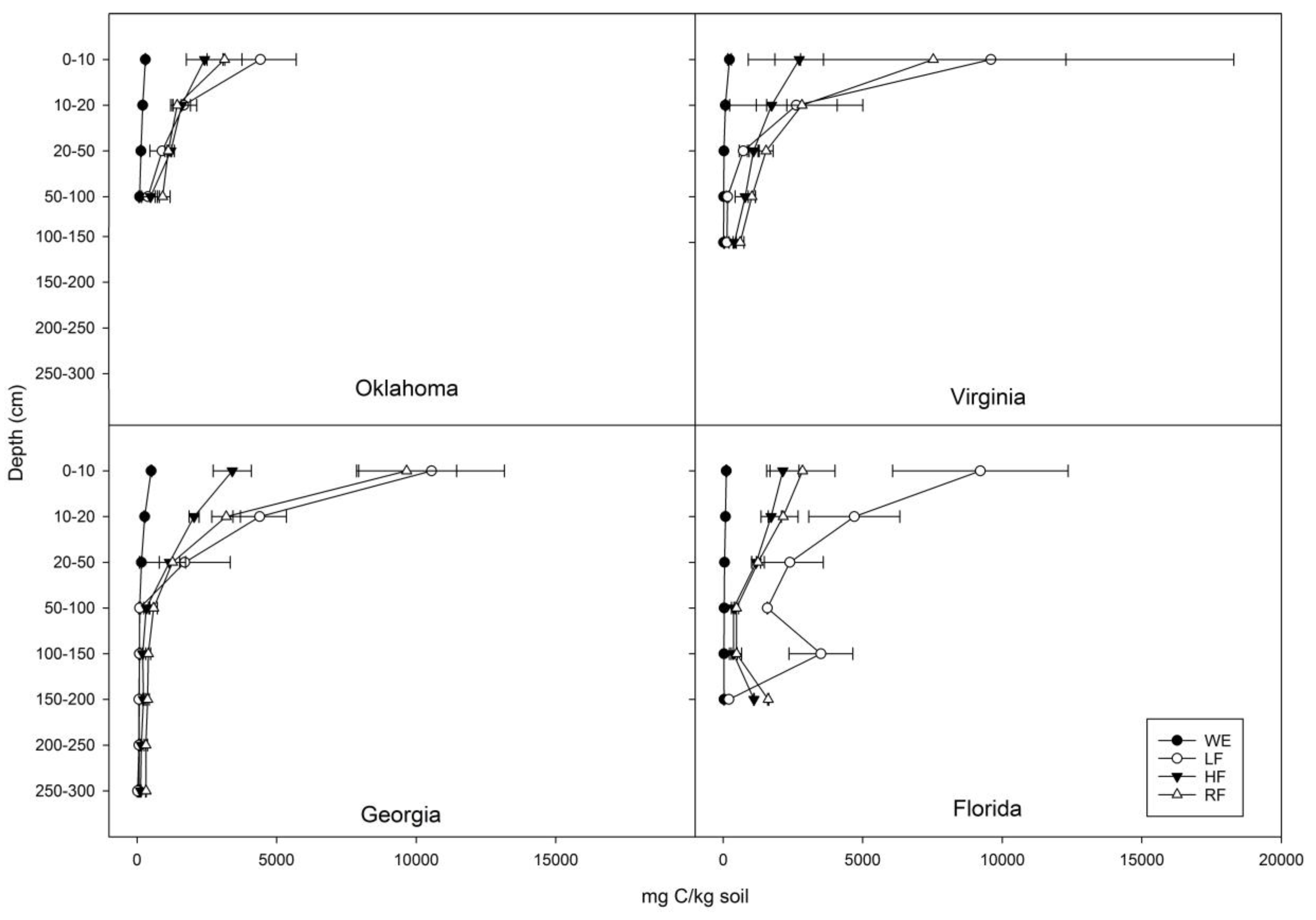
3.1. Distribution of C
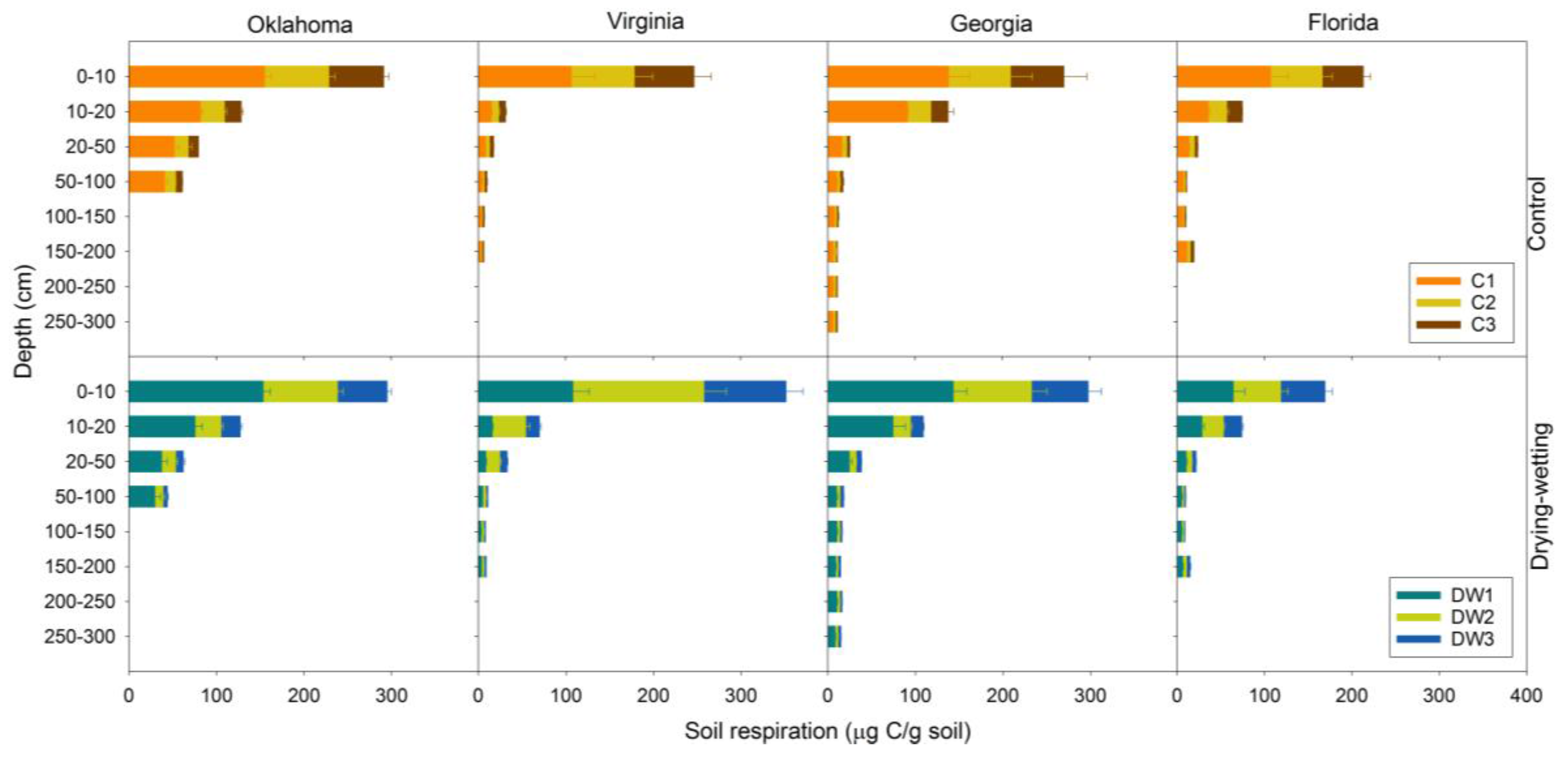
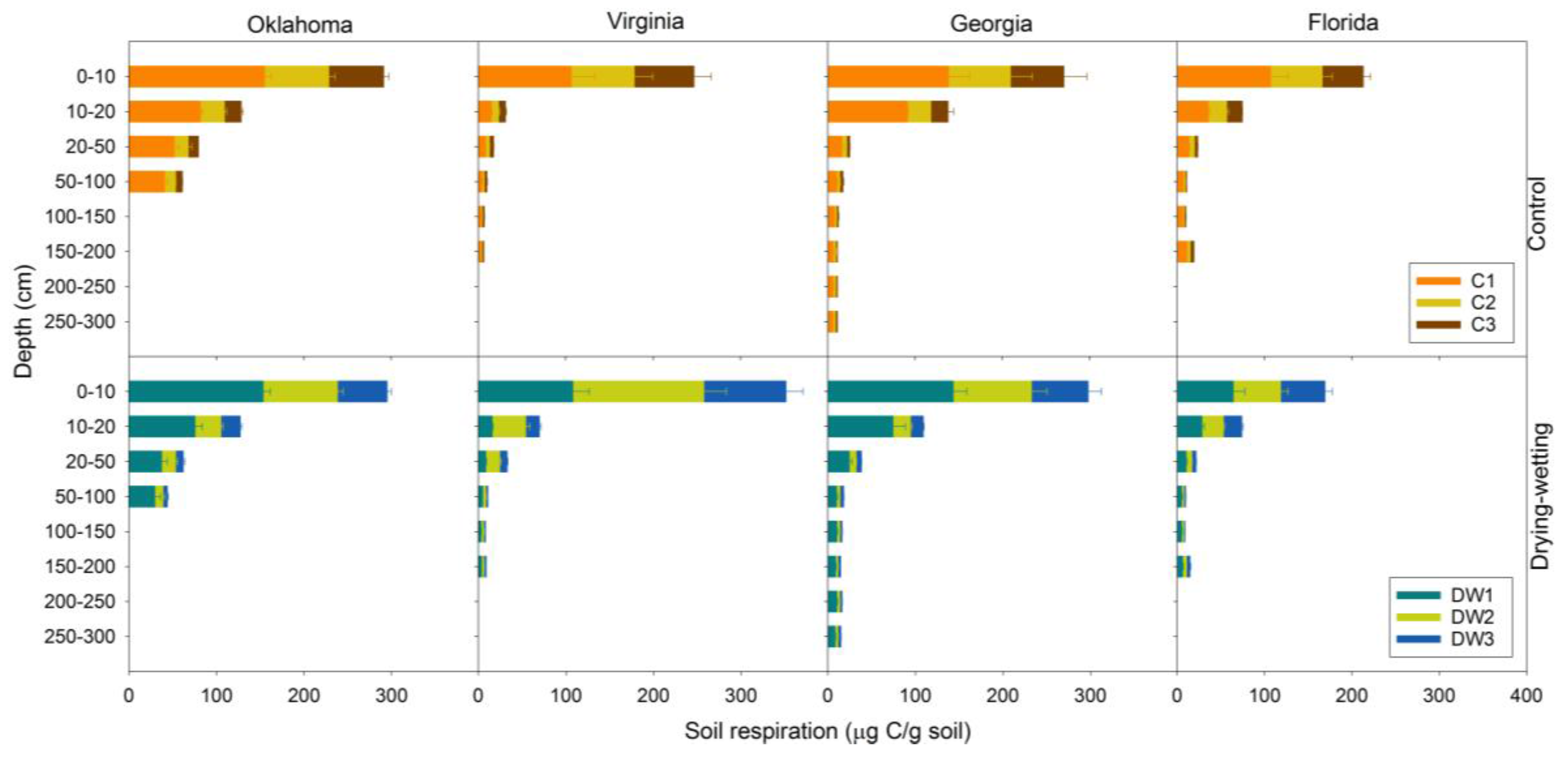
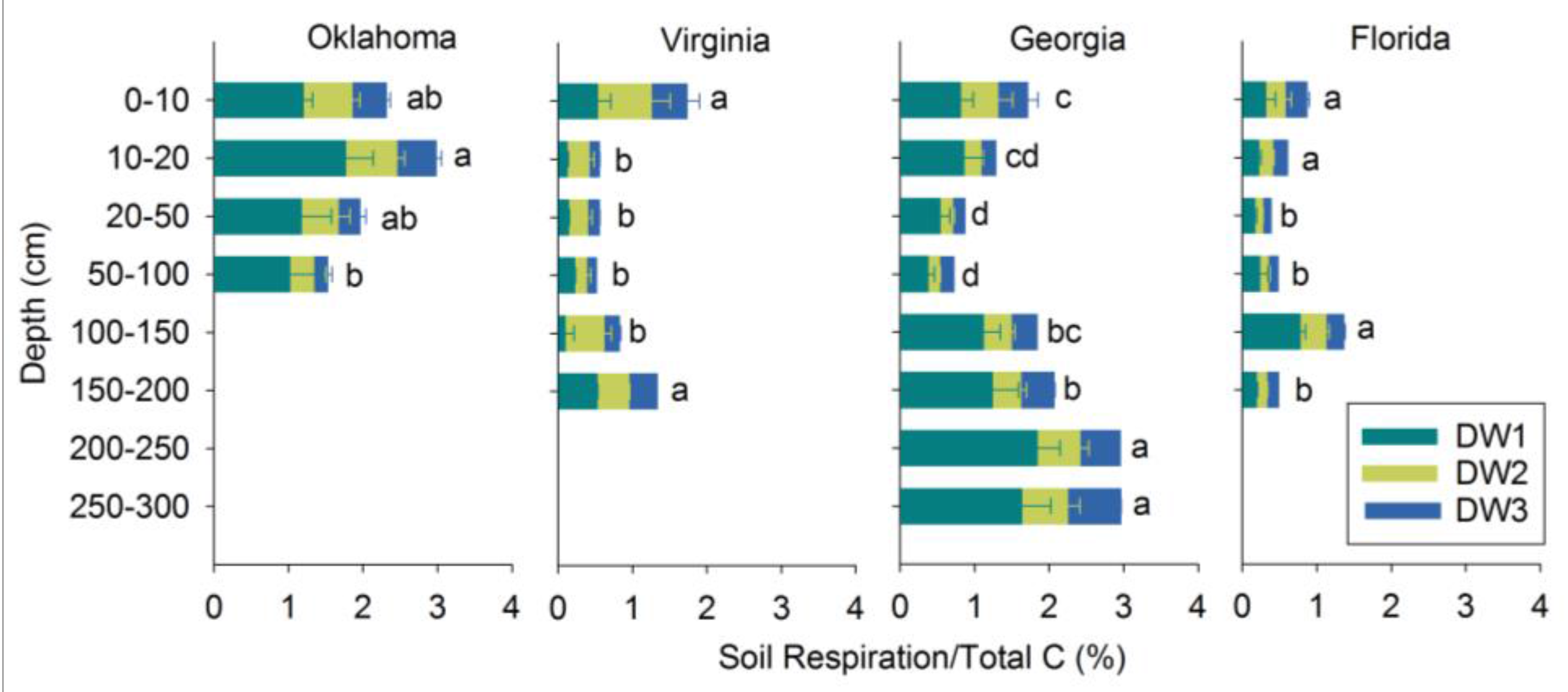
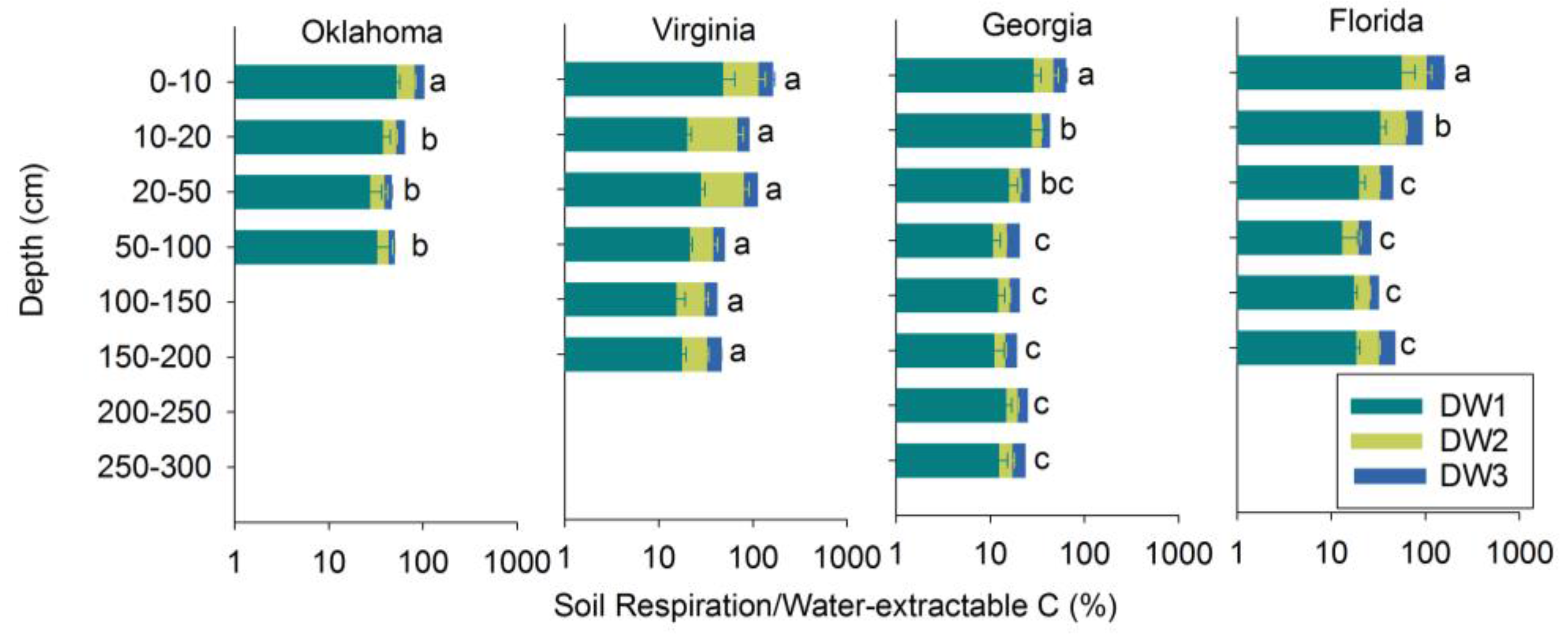
3.2. Soil Respiration
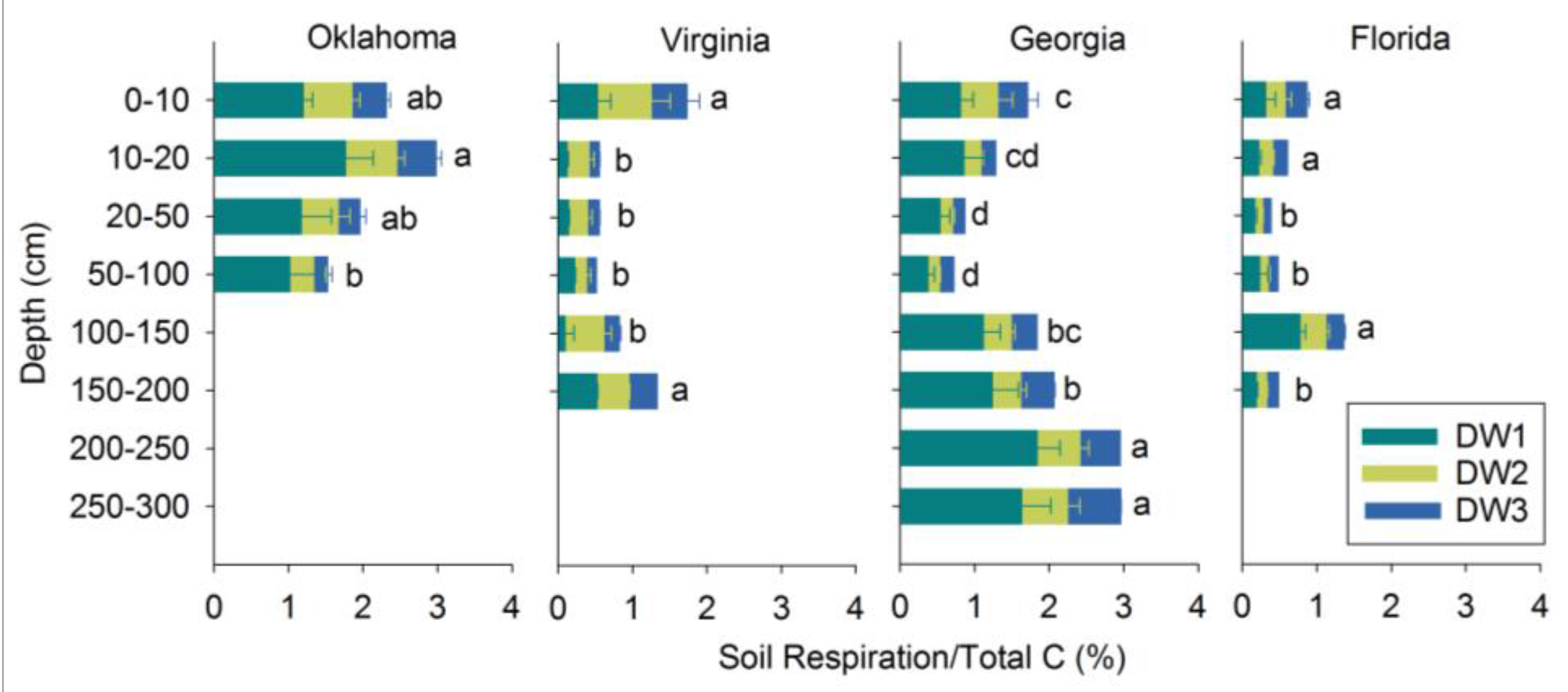
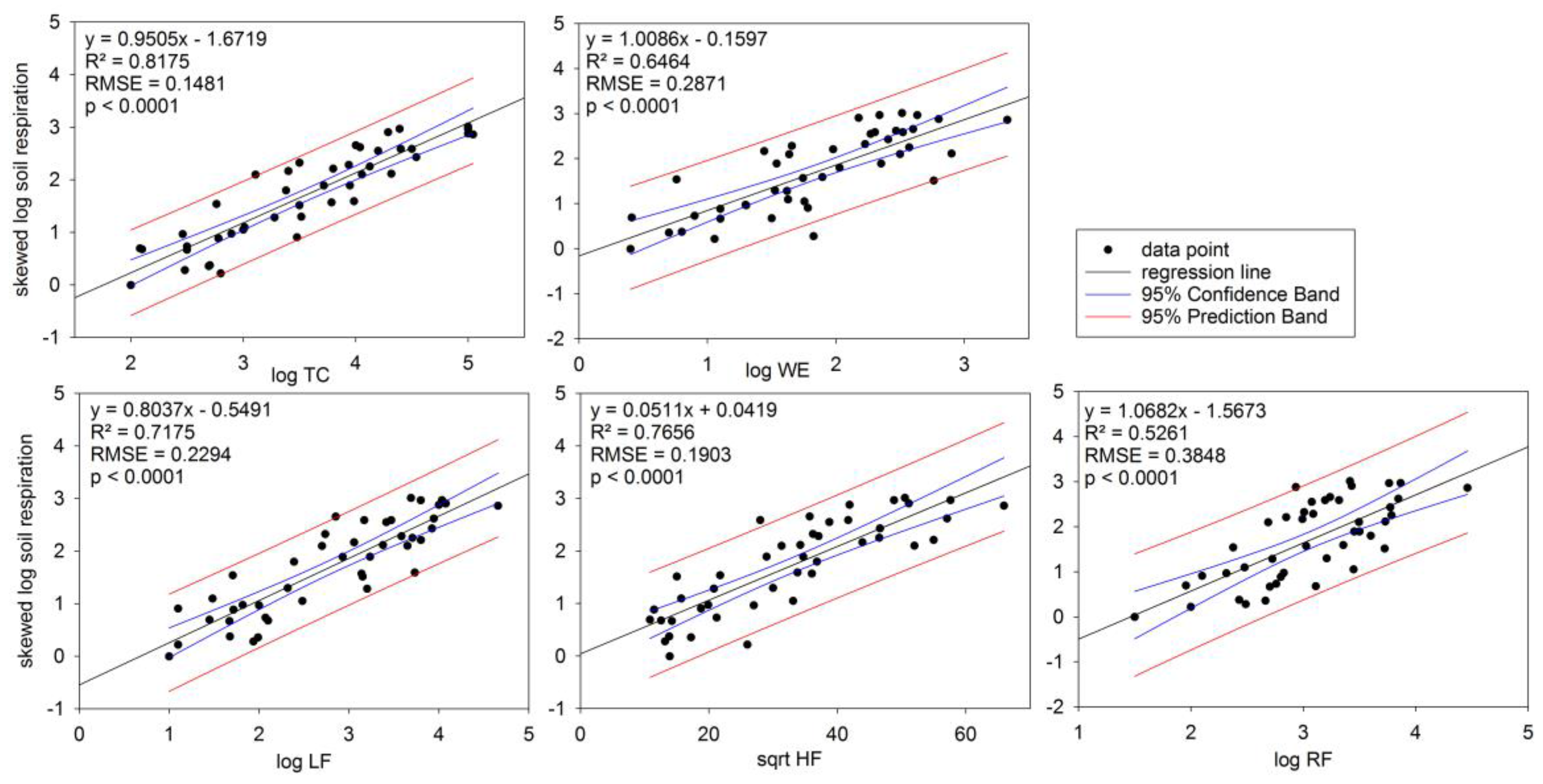
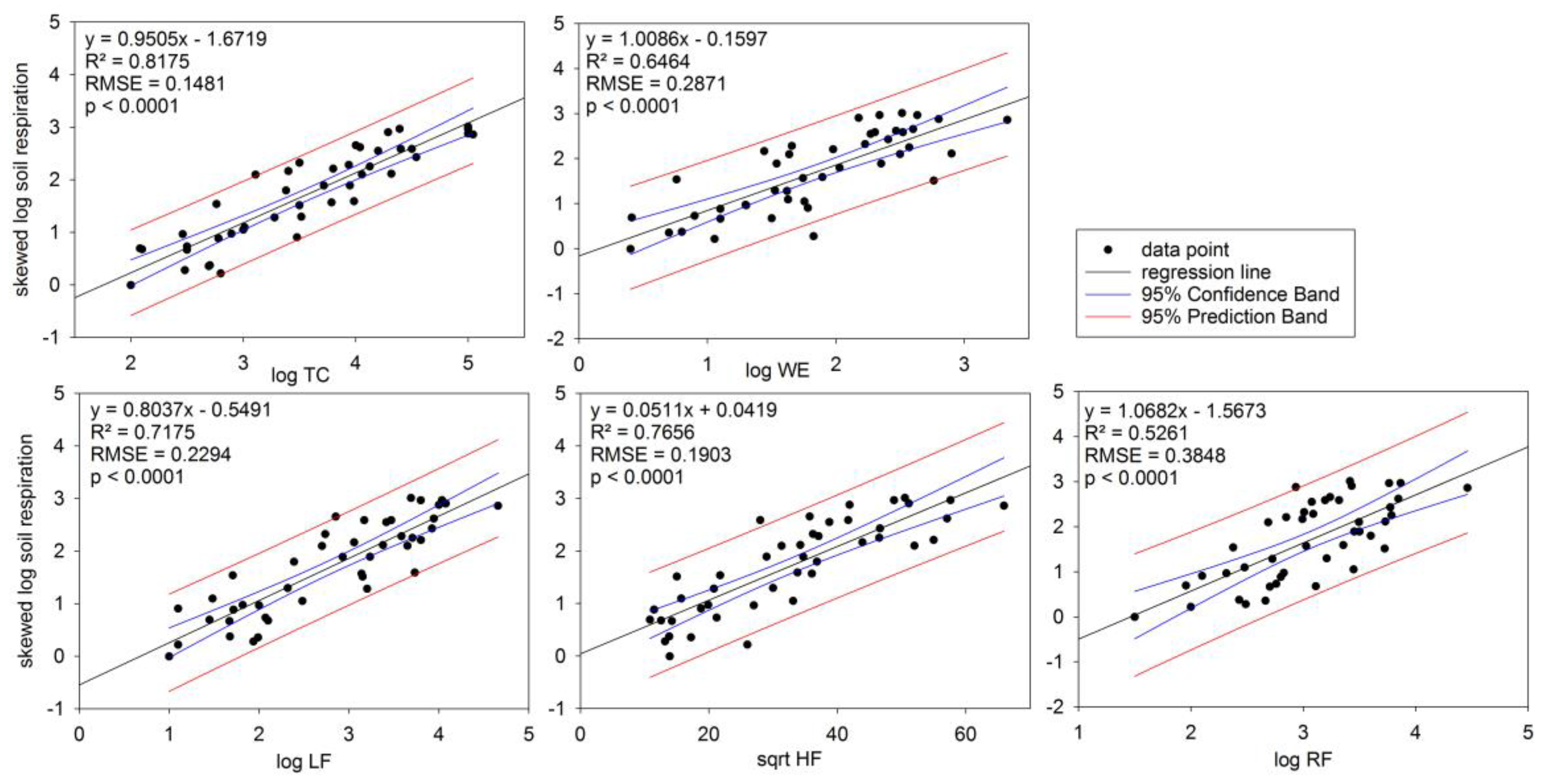
3.3. Soil Respiration in Relation to TC
3.4. Soil Respiration in Relation to Soil C Fractions
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Meehl, G.A.; Tebaldi, C.; Teng, H.; Peterson, T.C. Current and future US weather extremes and El Nino. Geophys. Res. Lett. 2007, 34, L20704. [Google Scholar] [CrossRef]
- Solomon, S.; Qin, D.; Manning, M.; Marquis, M.; Averyt, K.; Tignor, M.M.B.; Miller, H.L., Jr.; Chen, Z. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernamental Panel on Climate Change. In Climate Change 2007: The Physical Science Basis; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Seager, R.; Tzanova, A.; Nakamura, J. Drought in the Southeastern United States: Causes, variability over the last millennium, and the potential for future hydroclimate change. J. Clim. 2009, 22, 5021–5045. [Google Scholar] [CrossRef]
- Borken, W.; Matzner, E. Reappraisal of drying and wetting effects on C and N mineralization and fluxes in soils. Glob. Chang. Biol. 2009, 15, 808–824. [Google Scholar] [CrossRef]
- Birch, H.F. Pattern of humus decomposition in East African soils. Nature 1958, 181, 788. [Google Scholar] [CrossRef]
- Orchard, V.A.; Cook, F.J. Relationship between soil respiration and soil-moisture. Soil Biol. Biochem. 1983, 15, 447–453. [Google Scholar] [CrossRef]
- Jager, G.; Bruins, E.H. Effect of repeated drying at different temperatures on soil organic matter decomposition and characteristics, and on soil microflora. Soil Biol. Biochem. 1975, 7, 153–159. [Google Scholar] [CrossRef]
- Clein, J.S.; Schimel, J.P. Reduction in microbial activity in birch litter due to drying and rewetting events. Soil Biol. Biochem. 1994, 26, 403–406. [Google Scholar] [CrossRef]
- Franzluebbers, A.J.; Haney, R.L.; Honeycutt, C.W.; Schomberg, H.H.; Hons, F.M. Flush of carbon dioxide following rewetting of dried soil relates to active organic pools. Soil Sci. Soc. Am. J. 2000, 64, 613–623. [Google Scholar] [CrossRef]
- Fierer, N.; Schimel, J.P. Effects of drying-rewetting frequency on soil carbon and nitrogen transformations. Soil Biol. Biochem. 2002, 34, 777–787. [Google Scholar] [CrossRef]
- Mikha, M.M.; Rice, C.W.; Milliken, G.A. Carbon and nitrogen mineralization as affected by drying and wetting cycles. Soil Biol. Biochem. 2005, 37, 339–347. [Google Scholar] [CrossRef]
- Beare, M.H.; Gregorich, E.G.; St-Georges, P. Compaction effects on CO2 and N2O production during drying and rewetting of soil. Soil Biol. Biochem. 2009, 41, 611–621. [Google Scholar] [CrossRef]
- Butterly, C.R.; Marschner, P.; McNeill, A.M.; Baldock, J.A. Rewetting CO2 pulses in Australian agricultural soils and the influence of soil properties. Biol. Fertil. Soils 2010, 46, 739–753. [Google Scholar] [CrossRef]
- Butterly, C.R.; McNeill, A.M.; Baldock, J.A.; Marschner, P. Changes in water content of two agricultural soils does not alter labile P and C pools. Plant Soil 2011, 348, 185–201. [Google Scholar] [CrossRef]
- Wu, J.; Brookes, P.C. The proportional mineralisation of microbial biomass and organic matter caused by air-drying and rewetting of a grassland soil. Soil Biol. Biochem. 2005, 37, 507–515. [Google Scholar] [CrossRef]
- Fontaine, S.; Barot, S.; Barre, P.; Bdioui, N.; Mary, B.; Rumpel, C. Stability of organic carbon in deep soil layers controlled by fresh carbon supply. Nature 2007, 450, 277–280. [Google Scholar] [CrossRef] [PubMed]
- Fierer, N.; Allen, A.S.; Schimel, J.P.; Holden, P.A. Controls on microbial CO2 production: A comparison of surface and subsurface soil horizons. Glob. Chang. Biol. 2003, 9, 1322–1332. [Google Scholar] [CrossRef]
- Batjes, N.H. Total carbon and nitrogen in the soils of the world. Eur. J. Soil Sci. 1996, 47, 151–163. [Google Scholar] [CrossRef]
- Harrison, R.B.; Footen, P.W.; Strahm, B.D. Deep Soil Horizons: Contribution and Importance to Soil Carbon Pools and in Assessing Whole-Ecosystem Response to Management and Global Change. For. Sci. 2011, 57, 67–76. [Google Scholar]
- Gaudinski, J.B.; Trumbore, S.E.; Davidson, E.A.; Zheng, S. Soil Carbon Cycling in a Temperate Forest: Radiocarbon-Based Estimates of Residence Times, Sequestration Rates and Partitioning of Fluxes. Biogeochemistry 2000, 51, 33–69. [Google Scholar] [CrossRef]
- Wood, B.D.; Keller, C.K.; Johnstone, D.L. In situ measurement of microbial activity and controls on microbial CO2 production in the unsaturated zone. Water Resour. Res. 1993, 29, 647–659. [Google Scholar] [CrossRef]
- Trumbore, S.E.; Vogel, J.S.; Southon, J.R. AMS 14C measurements of fractionated soil organic matter: An approach to deciphering the soil carbon cycle. Radiocarbon 1989, 31, 644–654. [Google Scholar] [CrossRef]
- Trumbore, S.E.; Davidson, E.A.; Decamargo, P.B.; Nepstad, D.C.; Martinelli, L.A. Belowground cycling of carbon in forests and pastures of Eastern Amazonia. Glob. Biogeochem. Cycles 1995, 9, 515–528. [Google Scholar] [CrossRef]
- Trumbore, S. Age of soil organic matter and soil respiration: Radiocarbon constraints on belowground C dynamics. Ecol. Appl. 2000, 10, 399–411. [Google Scholar] [CrossRef]
- Trumbore, S.E.; Zheng, S.H. Comparison of fractionation methods for soil organic matter 14C analysis. Radiocarbon 1996, 38, 219–229. [Google Scholar] [CrossRef]
- Will, R.E.; Fox, T.; Akers, M.; Domec, J.C.; Gonzalez-Benecke, C.; Jokela, E.J.; Kane, M.; Laviner, M.A.; Lokuta, G.; Markewitz, D.; et al. A range-wide experiment to investigate nutrient and soil moisture interactions in loblolly pine plantations. Forests 2015, 6, 2014–2028. [Google Scholar] [CrossRef]
- NOAA National Weather Service Cooperative Observation Program. Available online: http://www.ncdc.noaa.gov (accessed on 15 February 2016).
- USDA Soil Classification. Available online: https://soilseries.sc.egov.usda.gov (accessed on 12 February 2016).
- Gee, G.W.; Or, D. 2.4 Particle-size analysis. In Methods of Soil Analysis: Part 4 Physical Methods SSSA Book Series; Soil Science Society of America (SSSA): Fitchburg, WI, USA, 2002; pp. 255–293. [Google Scholar]
- Thomas, G.W. Soil pH and soil acidity. In Methods of Soil Analysis: Part 3—Chemical Methods SSSA Book Series; Soil Science Society of America (SSSA): Fitchburg, WI, USA, 1996; pp. 475–490. [Google Scholar]
- Bremner, J.M. Nitrogen-total. In Methods of Soil Analysis: Part 3—Chemical Methods SSSA Book Series; Soil Science Society of America (SSSA): Fitchburg, WI, USA, 1996; pp. 1085–1121. [Google Scholar]
- Jones, D.L.; Willett, V.B. Experimental evaluation of methods to quantify dissolved organic nitrogen (DON) and dissolved organic carbon (DOC) in soil. Soil Biol. Biochem. 2006, 38, 991–999. [Google Scholar] [CrossRef]
- Golchin, A.; Oades, J.M.; Skjemstad, J.O.; Clarke, P. Study of free and occluded particulate organic-matter in soils by solid-state 13C CP/MAS NMR-spectroscopy and scanning electron-microscopy. Aust. J. Soil Res. 1994, 32, 285–309. [Google Scholar] [CrossRef]
- Zibilske, L.M. Carbon mineralization 1. In Methods of Soil Analysis: Part 2—Microbiological and Biochemical Properties SSSA Book Series; Soil Science Society of America (SSSA): Fitchburg, WI, USA, 1994; pp. 835–863. [Google Scholar]
- Magid, J.; Kjaergaard, C.; Gorissen, A.; Kuikman, P.J. Drying and rewetting of a loamy sand soil did not increase the turnover of native organic matter, but retarded the decomposition of added C-14-labelled plant material. Soil Biol. Biochem. 1999, 31, 595–602. [Google Scholar] [CrossRef]
- Franzluebbers, K.; Weaver, R.W.; Juo, A.S.R.; Franzluebbers, A.J. Carbon and nitrogen mineralization from cowpea plants part decomposing in moist and in repeatedly dried and wetted soil. Soil Biol. Biochem. 1994, 26, 1379–1387. [Google Scholar] [CrossRef]
- Curtin, D.; Selles, F.; Wang, H.; Campbell, C.A.; Biederbeck, V.O. Carbon dioxide emissions and transformation of soil carbon and nitrogen during wheat straw decomposition. Soil Sci. Soc. Am. J. 1998, 62, 1035–1041. [Google Scholar] [CrossRef]
- Kruse, J.S.; Kissel, D.E.; Cabrera, M.L. Effects of drying and rewetting on carbon and nitrogen mineralization in soils and incorporated residues. Nutr. Cycl. Agroecosyst. 2004, 69, 247–256. [Google Scholar] [CrossRef]
- Nelson, P.N.; Baldock, J.A.; Oades, J.M. Changes in dispersible clay content, organic carbon content, and electrolyte composition following incubation of sodic soil. Soil Res. 1998, 36, 883–898. [Google Scholar] [CrossRef]
- Xiang, S.-R.; Doyle, A.; Holden, P.A.; Schimel, J.P. Drying and rewetting effects on C and N mineralization and microbial activity in surface and subsurface California grassland soils. Soil Biol. Biochem. 2008, 40, 2281–2289. [Google Scholar] [CrossRef]
- Rumpel, C.; Kögel-Knabner, I. Deep soil organic matter-a key but poorly understood component of terrestrial C cycle. Plant Soil 2011, 338, 143–158. [Google Scholar] [CrossRef]
- Adu, J.K.; Oades, J.M. Physical factors influencing decomposition of organic materials in soil aggregates. Soil Biol. Biochem. 1978, 10, 109–115. [Google Scholar] [CrossRef]
- Eusterhues, K.; Rumpel, C.; Kleber, M.; Kögel-Knabner, I. Stabilisation of soil organic matter by interactions with minerals as revealed by mineral dissolution and oxidative degradation. Org. Geochem. 2003, 34, 1591–1600. [Google Scholar] [CrossRef]
- Kieft, T.L.; Soroker, E.; Firestone, M.K. Microbial biomass response to a rapid increase in water potential when dry soil is wetted. Soil Biol. Biochem. 1987, 19, 119–126. [Google Scholar] [CrossRef]
- Rovira, P.; Vallejo, V.R. Labile and recalcitrant pools of carbon and nitrogen in organic matter decomposing at different depths in soil: An acid hydrolysis approach. Geoderma 2002, 107, 109–141. [Google Scholar] [CrossRef]
- Silveira, M.L.; Comerford, N.B.; Reddy, K.R.; Cooper, W.T.; El-Rifai, H. Characterization of soil organic carbon pools by acid hydrolysis. Geoderma 2008, 144, 405–414. [Google Scholar] [CrossRef]
- Harper, R.J.; Tibbett, M. The hidden organic carbon in deep mineral soils. Plant Soil 2013, 368, 641–648. [Google Scholar] [CrossRef]
- Koarashi, J.; Hockaday, W.C.; Masiello, C.A.; Trumbore, S.E. Dynamics of decadally cycling carbon in subsurface soils. J. Geophys. Res. Biogeosci. 2012, 117, G03033. [Google Scholar] [CrossRef]
- Scharlemann, J.P.W.; Tanner, E.V.J.; Hiederer, R.; Kapos, V. Global soil carbon: Understanding and managing the largest terrestrial carbon pool. Carbon Manag. 2014, 5, 81–91. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Lat (°N) | Long (°W) | Avg Max Temp (°C) | Avg Min Temp (°C) | Avg Ppt (m) | Soil Series |
|---|---|---|---|---|---|---|
| McCurtain, Oklahoma | 35.15 | 94.97 | 23.9 | 9.4 | 1.3 | Ruston |
| Buckingham, Virginia | 37.57 | 78.53 | 19.4 | 6.7 | 1.1 | Littlejoe |
| Taliaferro, Georgia | 33.57 | 82.88 | 22.7 | 10.1 | 1.1 | Lloyd-Cecil |
| Taylor, Florida | 30.11 | 83.58 | 27.3 | 13.6 | 1.4 | Melvina-Moriah-Lutterloh |
| Location | Depth (cm) | Texture | pH | C (%) | N (%) |
|---|---|---|---|---|---|
| Oklahoma | 0–10 | loam | 4.8 ± 0.5 | 1.29 ± 0.25 | 0.07 ± 0.03 |
| 10–20 | loam | 4.2 ± 0.2 | 0.43 ± 0.12 | 0.04 ± 0.01 | |
| 20–50 | loam | 3.9 ± 0.0 | 0.32 ± 0.14 | 0.03 ± 0.00 | |
| 50–100 | clay loam | 3.6 ± 0.2 | 0.29 ± 0.03 | 0.03 ± 0.01 | |
| Virginia | 0–10 | silt loam | 4.2 ± 0.1 | 3.89 ± 0.54 | 0.14 ± 0.03 |
| 10–20 | loam | 4.5 ± 0.3 | 1.00 ± 0.24 | 0.04 ± 0.01 | |
| 20–50 | silty clay | 4.5 ± 0.4 | 0.52 ± 0.16 | 0.03 ± 0.00 | |
| 50–100 | silty clay loam | 4.9 ± 0.1 | 0.23 ± 0.05 | 0.02 ± 0.00 | |
| 100–150 | silty clay loam | 4.9 ± 0.0 | 0.09 ± 0.01 | 0.00 ± 0.00 | |
| Georgia | 0–10 | loam | 5.5 ± 0.5 | 1.92 ± 0.67 | 0.10 ± 0.03 |
| 10–20 | clay loam | 5.4 ± 0.2 | 0.87 ± 0.26 | 0.06 ± 0.01 | |
| 20–50 | clay | 5.4 ± 0.1 | 0.44 ± 0.13 | 0.04 ± 0.00 | |
| 50–100 | clay | 5.2 ± 0.4 | 0.25 ± 0.02 | 0.03 ± 0.00 | |
| 100–150 | clay | 5.8 ± 0.0 | 0.07 ± 0.03 | 0.00 ± 0.00 | |
| 150–200 | clay loam | 5.6 ± 0.1 | 0.06 ± 0.03 | 0.00 ± 0.00 | |
| 200–250 | clay loam | 5.4 ± 0.3 | 0.05 ± 0.02 | 0.00 ± 0.00 | |
| 250–300 | loam | 5.3 ± 0.2 | 0.05 ± 0.01 | 0.01 ± 0.00 | |
| Florida | 0–10 | sand | 5.5 ± 0.2 | 2.06 ± 0.36 | 0.07 ± 0.02 |
| 10–20 | sand | 5.6 ± 0.1 | 1.30 ± 0.10 | 0.04 ± 0.01 | |
| 20–50 | sand | 5.8 ± 0.3 | 0.62 ± 0.09 | 0.02 ± 0.00 | |
| 50–100 | sand | 6.2 ± 0.4 | 0.22 ± 0.03 | 0.01 ± 0.00 | |
| 100–150 | sandy clay loam | 5.2 ± 0.1 | 0.07 ± 0.01 | 0.00 ± 0.00 | |
| 150–170 | sandy clay loam | 4.8 ± 0.3 | 0.33 ± 0.03 | 0.00 ± 0.00 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qi, J.; Markewitz, D.; Foroughi, M.; Jokela, E.; Strahm, B.; Vogel, J. Drying-Wetting Cycles: Effect on Deep Soil Carbon. Soil Syst. 2018, 2, 3. https://doi.org/10.3390/soils2010003
Qi J, Markewitz D, Foroughi M, Jokela E, Strahm B, Vogel J. Drying-Wetting Cycles: Effect on Deep Soil Carbon. Soil Systems. 2018; 2(1):3. https://doi.org/10.3390/soils2010003
Chicago/Turabian StyleQi, Ji, Daniel Markewitz, Maryam Foroughi, Eric Jokela, Brian Strahm, and Jason Vogel. 2018. "Drying-Wetting Cycles: Effect on Deep Soil Carbon" Soil Systems 2, no. 1: 3. https://doi.org/10.3390/soils2010003
APA StyleQi, J., Markewitz, D., Foroughi, M., Jokela, E., Strahm, B., & Vogel, J. (2018). Drying-Wetting Cycles: Effect on Deep Soil Carbon. Soil Systems, 2(1), 3. https://doi.org/10.3390/soils2010003