Abstract
The increasing presence of successful invasive plant species can have an impact on wetlands capacity to store and release C. We have investigated the relationships between stocks of different soil organic carbon (SOC) along the soil vertical profile and invasive plant success in a China wetland. In stands dominated by the exotic invasive species Spartina alterniflora and the native invasive Phragmites australis soil organic-carbon concentrations (SOC) were higher (12% and 9%, respectively) than in plots of a native species, Cyperus malaccensis, whereas SOC content (g m−2) was 18% and 17% lower under P. australis than under S. alterniffolia and C. malaccensis, respectively. Soils under both invasive species had the concentrations and contents of light-fraction organic carbon (LFOC), light-fraction organic nitrogen (LFON) at 30–60 cm of soil depth and labile organic carbon (LOC) concentrations at 0–10 cm higher than soils under native species. The invasive species had higher total aboveground, total biomasses and lower shoot:root ratios than the native species. The success of both invasive species was associated with higher growth rates and accumulation of nutrients in biomass than in the native species and also accumulation of C in plant soil system. The stands currently dominated by the invasive species were recently occupied by monospecific stands of the native C. malaccensis, strongly suggesting that all or most of the current soil differences were due to the invasions. Higher sand fraction in C. malaccensis community and higher clay fraction in P. australis community relative to the native species, were correlated with higher soil N and P concentrations in invaded stands. The results suggest that different vegetation cover with distinct shoot/root ratio can change soil structure by favoring sedimentation of different particle size classes. Thus, despite both invasive species have some common traits, the results also showed that different invasive species with partially distinct impacts on soil and nutrient uses can succeed under the same conditions. The traits conferring invasive success are thus not necessarily species-specific. A clear change in the general accumulation of C, N and P in the plant-soil system was related to the invasive plant success in this wetland areas.
1. Introduction
The world contains an estimated 2011 Pg of soil [1]. The importance of soil as a carbon sink is crucial for the ability of the earth to buffer the increasing levels of atmospheric CO2 as a consequence of human activities [2,3,4]. Estuarine wetland soils, due to their location at the river-ocean interface, have a large potential to act as sediment sinks for accumulating C [5,6,7]. Coastal wetlands in China have a pivotal economic and environmental importance. These wetlands cover 5.80 × 104 km2 [8] and provide many services and products [9,10] but several impacts in recent decades have threatened this capacity. Increases in tropical storms and ocean levels have altered flooding intensities, which together with increased pollution and nutrient loads have had impacts on sedimentary processes and the capacity of wetlands to store and release C and nutrients [11,12,13,14,15,16].
Changes in land use and the success of invasive plant species have also had an impact on the capacity of wetlands to store and release C [17,18]. Invasive plant species can promote different patterns in plant-soil nutrient cycling and store than native species and thus change the capacity to accumulate N and P in biomass and soil [10]. The impacts of invasive plant success on the concentrations and stoichiometry of soil nutrients have been widely studied [16,19] and some general trends have emerged [16,20,21]. The metadata analyses in terrestrial ecosystem have shown (data mostly from not flooded ecosystems) that the impacts can differ and even be opposite depending on the natural availability levels of soil nutrients and generally on the capacity of a site to sustain low or high plant production in accordance with the soil and climatic conditions [16,20,21]. A recent metadata study has shown that invasive plants were associated with higher N and P concentration and lower C:N ratio in photosynthetic tissues and with higher soil C and N stocks and N, P and K availabilities in nutrient-poor but not in nutrient-rich environments. The results of this study also suggest higher nutrient resorption efficiencies in invasive than in native species in nutrient-poor environments but higher nutrient uptake capacity in nutrient-rich sites related to higher growth rates of invasive species [22]. The impacts of plant invasion and its relationships with C, N, P concentrations, contents and stoichiometry in flooded ecosystems (permanent or temporally) as wetlands are not well known
The total content of soil organic carbon (SOC) changes over a long time scale. Changes in a soil chronosequence from 50 to 2000 years have been clearly observed [23] but changes in short time scales of few years or few decades are not easily discerned [23]. So, distinguishing between the active-C fraction from the total SOC pool is important for assessing the effect of plant invasion on soil C dynamics. Active C refers to the fraction of soil C that is strongly influenced by plants and microorganisms and is highly susceptible to oxidation and decomposition [24,25]. Different separation and extraction methods can separate active SOC into fractions of dissolved organic carbon (DOC), labile organic carbon (LOC), light-fraction organic carbon (LFOC) and microbial biomass organic carbon (MBC) [26]. These different forms of active SOC have different sensitivities to environmental change [27] but few studies have examined the relationships between the different forms of active SOC and changes in environmental parameters [28,29], especially soil properties. Active SOC is a major source of CO2 and CH4 produced by microbes, so properly managing this pool of active C is important for mitigating global climate change [30,31].
C:nutrient stoichiometry is a good indicator of changes in soil C dynamics in wetlands due to changes in the environment. Stoichiometric data can provide decision makers with the necessary information for developing effective methods to enhance the potential capacity of these ecosystems to fix C and reduce the impact of the emissions of C-source greenhouse gases [32,33]. This information could also improve our understanding of the impacts on the potential uses and regeneration capacity of flooded wetlands by determining the C cycles and balances and the fertility of the soil.
Most available data most suggest that low costs of foliar construction and high phenotypic plasticity in taking up available nutrients frequently contribute to alien success in nutrient-rich environments [16,20,34], whereas most studies suggest that the success of invasive plants in nutrient-poor soils depends on more conservative strategies such as a higher nutrient-use efficiency [19,20,21], especially on short timescales [19]. Too many exceptions, however, prevent this question from being totally resolved [16]. The impacts of plant invasion on total soil C accumulation in the various soil fractions and the further relationships of these impacts on C with other soil traits such as C:N:P ratios are also unresolved questions [16]. Some studies have associated invasive plant success with changes in the C:nutrient ratios and soil C, N and P contents but these relationships have not been universally observed [16]. The relationships, particularly across soil depths, are thus not clear and merit further study. Some studies have discussed soil C dynamics and nutrient stoichiometry [35,36] but few have distinguished them at a fine scale in soil profiles. The relationships of species invasive success with soil C and nutrient status has been mainly focused in the first soil cm [16]. However, different soil textures and soil organic matter decomposition can produce accumulation of some organic carbon fractions along soil deep layers [37,38]. We hypothesized that changes in soil C store along soil profile could be related with different plant species. The knowledge of the variation of C accumulation along the soil vertical profile and its causes is crucial for understanding the capacity of wetlands to act as carbon sinks [39].
We aimed to determine whether the success of plant invasion at a site was based on a similar strategy of C, N and P use irrespective of the species or whether different species could become established in the same area and occupy a similar niche but have different impacts on soil C, N and P contents. We investigated the relationships of different dominant tall-herb wetland communities comprised of one native and two invasive species with the C contents of various soil fraction, soil N and P contents and C:nutrient ratios along soil profiles.
2. Materials and Methods
2.1. Study Area
This study was conducted in the Shanyutan wetland (26°01′46″ N, 119°37′31″ E; Figure 1), the largest tidal wetland (approximately 3120 ha) in the estuary of the Minjiang River (Fujian). The climate in this region is relatively warm and wet with a mean annual temperature of 19.6 °C and a mean annual precipitation of 1346 mm [40]. The region has a drop in precipitation only in the November-December period with a total precipitation of 51.2 mm in this period. The lowest temperature is in December. It belongs to the warm temperate climate according to Köppen Cfa climate classification. The river has higher flow in summer and lower in winter but the water flow is currently controlled by human management of the Shuikou reservoir. The soil surface is submerged across the study site beneath 10–120 cm of water for 3–3.5 h during each tidal inundation. The soil surfaces of the wetland are exposed at low tide but the soil remains flooded in some areas. The average annual percentage of water by weight in the soil and the soil redox potential are 116% and 12.6 mV, respectively. The average salinity of the tidal water between May and December 2007 was 4.2 ± 2.5‰. Thus, we chose a very homogeneous area to make very likely that the soil differences were due to the different species cover. Moreover, the bedrock is the same (granite) in all the studies area.
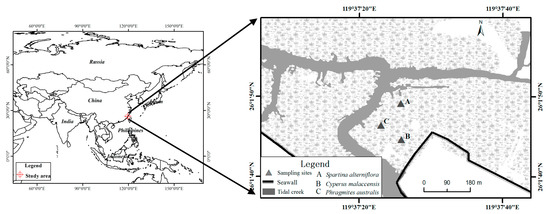
Figure 1.
The location of the study area and sampling sites (▲) in Fujian Province, southeastern China.
Cyperus malaccensis Lam., Phragmites australis (Cav.) Trin. Ex Steud. and Spartina alterniflora Loisel are the three dominant species of plants in the wetland, typically found in the upper (mid to high) portions of mudflats. C. malaccensis is native to the wetland, P. australis is native to China but not of the wetland area and is considered a native invasive species by its expansion in new areas against other native species, P. australis invaded this wetland more than 30 years ago S. alterniflora is an exotic plant that invaded this wetland more than 10 years ago. The decomposition rates of their litter are in the order S. alterniflora < P. australis < C. malaccensis [41]. Wetland soils in areas dominated by S. alterniflora and P. australis generally have a lower pH and bulk density than do soils in areas dominated by C. malaccensis [42]. All areas currently occupied by both invasive species were previously (10–30 years ago) C. malaccensis grasslands. The success of two invasive species in areas previously dominated by a native species supports the premise that differences in soil conditions are associated with the invading species.
2.2. Sample Collection and Measurements
Soil samples were collected in December 2009 in the Shanyutan wetland (Figure 1) from locations dominated by C. malaccensis (native plant), S. alterniflora (invaded more than 10 years ago) and P. australis (invaded more than 30 years ago). Three plots were randomly established in each community and soil profiles (width, 1 m; length, 1 m; depth, 0.6 m) were excavated. The samples were collected with a small sampler (length, 0.3 m; diameter, 0.1 m) from each of six soil layers (0–10, 10–20, 20–30, 30–40, 40–50 and 50–60 cm) at the center and both ends of the soil pits. These three samples from each layer were bulked to form one sample per layer. A total of 54 soil samples (three communities × three plots × six soil layers) were thus collected. The core samples were divided into two parts, with one part unprocessed for the measurement of soil MBC and DOC and the other part air-dried and finely powdered in a ball bill after the removal of all roots and visible plant remains for the determination of total SOC, LFOC and LOC. Total SOC was determined by K2Cr2O7-H2SO4 digestion [43,44], DOC by extracting the soils with deionized water (1:5 ratio) and measuring the liquid supernatant C concentration using a TOC-V CPH total carbon analyzer (Shimadzu Scientific Instruments, Kyoto, Japan), MBC by fumigation–extraction by 0.5 mol L−1 K2SO4 [45] and measuring the liquid supernatant C concentration using a TOC-V CPH total carbon analyzer (Shimadzu Scientific Instruments, Kyoto, Japan), LOC by digestion with 333 mM KMnO4, shake 1 h at 200 rpm, centrifuge 5 min at 2000× g and liquid supernatant measurement using a UV-2450 spectrophotometer (Shimadzu Scientific Instruments, Kyoto, Japan). LFOC was determined by 1.70 g mL−1 NaI separation, shaking 1 h at 200 rpm, centrifuging 20 min at 1000× g, getting the float light LFOC, drying it 72 h at 60 °C [45] and measuring with a Vario EL III Elemental Analyzer (Elementar Scientific Instruments, Hanau, Germany).
Bulk density was measured for the three bulked cores (5 cm diameter, 3 cm depth) collected from each soil layer. Soil salinity, pH and particle-size distribution were measured by a DDS-307 salinity meter (Boqu Scientific Instruments, Shanghai, China), an 868 pH meter (Orion Scientific Instruments, Thermo Fisher Instruments, Waltham, MA, USA) and a Master Sizer 2000 Laser Particle Size Analyzer (Malvern Scientific Instruments, Suffolk, UK), respectively. Soil water content was measured by drying the soil at 105 °C to a constant weight [45] and total soil N concentration was determined by total Kjeldahl digestion (Buchi Scientific Instruments, Flawil, Switzerland) and total soil P concentration was determined by perchloric-acid digestion followed by ammonium-molybdate colorimetry and measurement using a UV-2450 spectrophotometer (Shimadzu Scientific Instruments, Kyoto, Japan).
2.3. Statistical Analyses
We performed general linear models (GLM) with community composition as an independent categorical variable, soil traits (pH and clay concentration) as independent continuous variables and soil C fractions, N and P concentrations, contents and stoichiometric ratios as dependent continuous variables. We used Bonferroni post hoc tests to detect differences among different pair-wise comparisons between species at different soil depth layers. We used Statistica 6.0 (StatSoft, Inc., Tulsa, OK, USA) for the data analyses.
We also performed other multivariate statistical analyses. We determine the overall differences in the changes of the soil C fractions, N and P concentrations, stoichiometric ratios, salinity, pH and soil temperature in the species-specific plant communities using general discriminant analysis (GDA), including the component of the variance due to the different soil layers as an independent categorical variable. General discriminant analyses (GDA) consist of a supervised statistical algorithm that derives an optimal separation between groups established a priori by maximizing between-group variance while minimizing within-group variance [46]. GDA is thus an adequate tool for identifying the variables most responsible for the differences among groups while controlling the component of the variance due to other categorical variables. The GDAs were performed using Statistica 6.0 (StatSoft, Inc., Tulsa, OK, USA).
We also performed structural equation modelling (SEM) to analyze the factors explaining the maximum variability of the soil C fractions, N and P concentrations and stoichiometric ratios as functions of the soil traits (pH and clay concentration) and taking into account the possible effect of invasive plant success, using species factor as a dummy variable. This analysis separately compared the soil of the native community with those of each of the invading communities and provided information for the direct, indirect and total effects of the variables. We fitted the models using the R sem package [47] and identified the minimum adequate model using the Akaike information criterion. Standard errors and the significance level (P) of the direct, indirect and total effects were calculated by bootstrapping (1200 repetitions) [48,49].
3. Results
3.1. Univariate Analyses
The concentrations of total SOC, LOC, MBC, LFOC, LFON, N and P tended to significantly decrease with depth in the soil profile (Figure 2 and Figure 3, Table 1). The LFOC:LFON concentration ratio also tended to significantly decrease with depth (Figure 4, Table 1) but the SOC:P concentration ratio was lowest between depths of 10 and 30 cm. Soil SOC:N and N:P concentration ratios had no clear pattern of variation with soil depth. Average total contents (mg m−2) of DOC, MBC, LFOC, LFON, N and P tended to significantly decrease with depth (Figure 5 and Figure 6, Table 1), whereas total SOC and LOC contents were lowest between 20 and 40 cm.
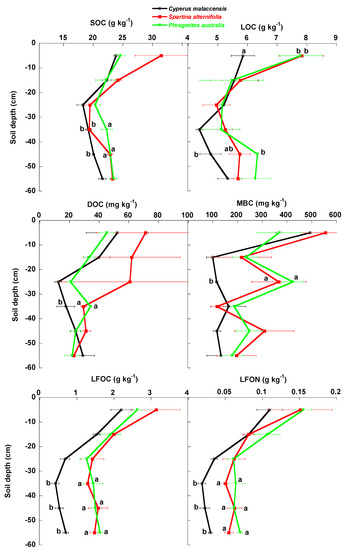
Figure 2.
Mean ± SE (g kg−1 soil dry weight) SOC, LOC, DOC, MBC, LFOC and LFON concentrations in the soils of the Cyperus malaccensis, Spartina alterniflora and Phragmites australis communities at different depths. Different letters indicate significant differences in the Bonferroni post-hoc tests (p < 0.05) among the species communities.
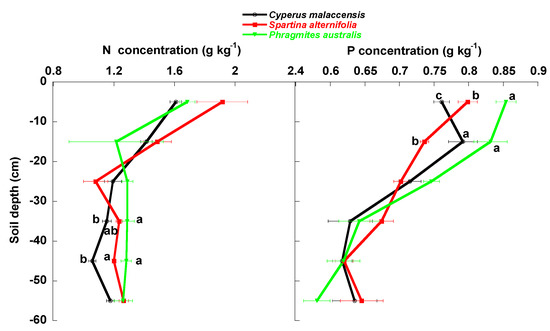
Figure 3.
Mean ± SE (g kg−1 soil dry weight) N and P concentrations in the soils of the Cyperus malaccensis, Spartina alterniflora and Phragmites australis communities at different depths. Different letters indicate significant differences in the Bonferroni post-hoc tests (p < 0.05) among the species communities.

Table 1.
Levels (mean ± SE) of the variables in the general linear models, with species community and soil layer as independent variables and the soil stoichiometric variables as dependent factors. We used Bonferroni post hoc test to detect differences among different pair-wise comparisons between species and soil depth layers.
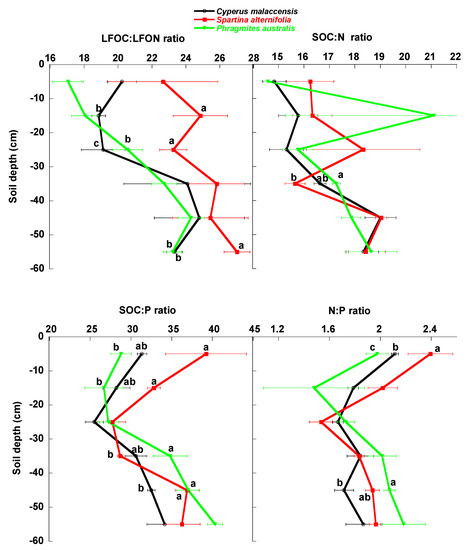
Figure 4.
Mean ± SE LFOC:LFON, SOC:N, SOC:P and N:P ratios in the soils of the Cyperus malaccensis, Spartina alterniflora and Phragmites australis communities at different depths. Different letters indicate significant differences in the Bonferroni post-hoc tests (p < 0.05) among the species communities.
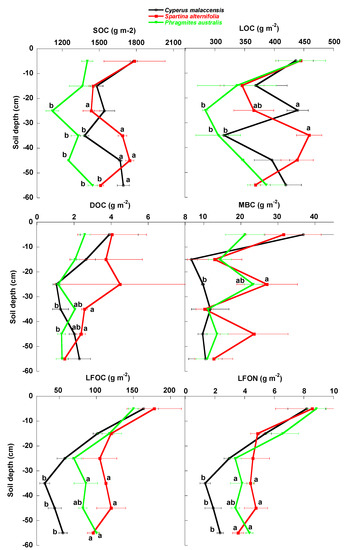
Figure 5.
Mean ± SE (g m−2 soil dry weight) total SOC, LOC, DOC, MBC, LFOC and LFON contents in the soils of the Cyperus malaccensis, Spartina alterniflora and Phragmites australis communities at different depths. Different letters indicate significant differences in the Bonferroni post-hoc tests (p < 0.05) among the species communities.
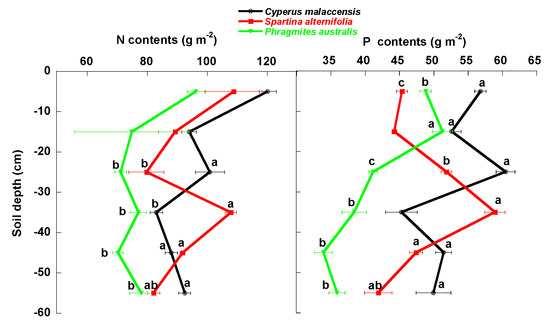
Figure 6.
Mean ± SE (g m−2 soil dry weight) N and P total contents in the soils of the Cyperus malaccensis, Spartina alterniflora and Phragmites australis communities at different depths. Different letters indicate significant differences in the Bonferroni post-hoc tests (p < 0.05) among the species communities.
The native C. malaccensis community had more total soil (0–60 cm) SOC and N contents than the P australis but not the S. alterniflora communities (Figure 7, Table 1) and more total P content than both invaded communities. The soil of the P. australis community had the lowest SOC, N and P contents, mainly due to the lower contents below 30 cm (Figure 2, Figure 3, Figure 4, Figure 5 and Figure 6, Table 1), whereas the LFOC, MBC and DOC were clearly higher in the topsoil than in the subsoil. The distribution of LFOC and LFON along the soil profile was more homogeneous under the two invasive species than under the native C. malaccensis (Table 2). Notably, the ratio for SOC is not different from 1 in the soils under any of the three studied species (Table 2), showing a constant distribution of total C along the first 60 cm of soil.
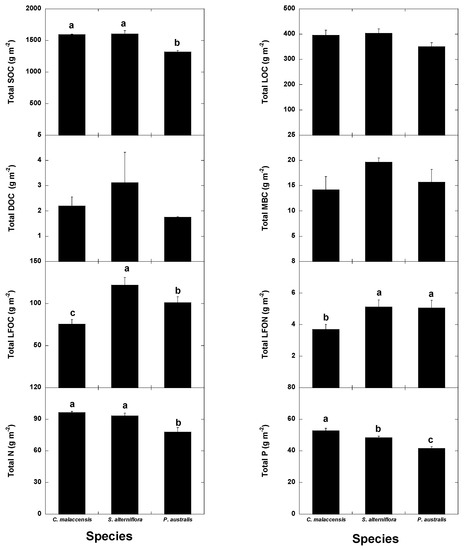
Figure 7.
Mean ± SE (g m−2 soil dry weight) SOC, LOC, DOC, MBC, LFOC, LFON, N and P contents in the soils of the Cyperus malaccensis, Spartina alterniflora and Phragmites australis communities at different depths. Different letters indicate significant differences in the Bonferroni post-hoc tests (p < 0.05) among the species communities.
Table 2.
Ratios (Mean ± SE) of the total contents between the contents of the studied variables between topsoil and subsoil (0–30 cm/30–60 cm). Different letters in a single row indicate significant differences in the post-hoc Bonferroni test (p < 0.05) among species.
SOC content was positively correlated with bulk density and N and P contents (Table 3). SOC and LOC concentrations were positively correlated with N concentration and N:P ratio (Table 3) but were not correlated with P concentration (Table 3). Of the C fractions, only LFOC concentrations and contents were positively correlated with both N and P concentrations (Table 3). As expected, soil water content was positively correlated with SOC concentrations, an effect associated with anoxic conditions and slow rates of mineralization of soil matter (Table 3).
Table 3.
Pearson correlations among the soil and plant nutrients and nutrient ratios. Bonferroni correction of α inflation was applied.
Soil water content did not differ among the soils of the three communities (Figure 8). pH and bulk density were lower in the soils for both invasive species than the native species and bulk density was lower in the P. australis soil than the S. alterniflora soil. Soil salinity was highest in the P. australis soil, followed by the C. malaccensis and S. alterniflora soils.
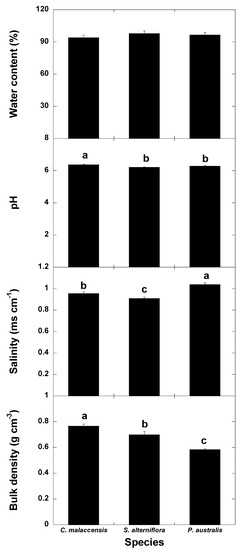
Figure 8.
Mean ± SE soil water content (%), pH, salinity (ms cm−1) and bulk density (g cm−3) in the Cyperus malaccensis, Spartina alterniflora and Phragmites australis communities at different depths. Different letters indicate significant differences in the Bonferroni post-hoc tests (p < 0.05) among the species communities.
3.2. Multivariate Analyses
The GDA of all variables significantly separated the soils of the three plant communities (Figure 9, Table 4). The soil of the native C. malaccensis community was mainly separated from those of the two invasive species by its higher bulk density and SOC, N and P concentrations and lower salinity. The GDA identified SOC concentration, bulk density and salinity as the most significant variables explaining the differences between the soils (Table 5). This analysis showed that the soils under P. australis had higher soil clay content and soil salinity and lower DOC, LOC and SOC than the soils under the other two species, whereas the native species C. malaccensis had higher soil N, P and SOC concentrations, soil bulk density and pH than the other two species (Figure 9).
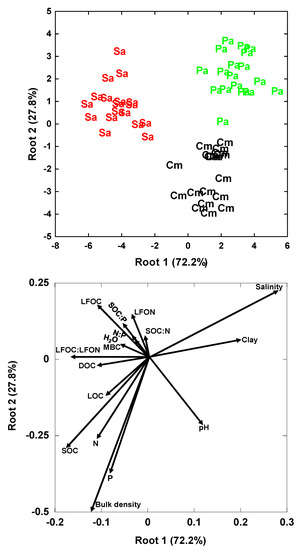
Figure 9.
Biplots of the standardized canonical discriminate function coefficients for the first two roots representing the soil variables as independent variables and the various grouping dependent factors corresponding to the plant communities. Cm = Cyperus malaccensis community, Sa = Spartina alterniflora community and Pa = Phragmites australis community.
Table 4.
Test statistics for squared Mahalanobis distances (M) among the plant communities, with soil salinity; clay %; pH; bulk density; N, P, SOC, LOC, DOC, MBC, LFOC and LFON concentrations and N:P, SOC:N and SOC:P concentration ratios as variables. Bold type indicates a significant effect of the variable in the model (p < 0.05).
Table 5.
Main effects of the variables in the GDA analysis. Statistics (Wilks’ λ and P) of the discriminant functional analysis among the plant communities, with soil salinity, clay %, pH, bulk density, N, P, SOC, LOC, DOC, MBC, LFOC and LFON concentrations and N:P, SOC:N and SOC:P concentration ratios as variables. The soil layers were included as a controlling categorical independent variable. Bold type indicates a significant effect of the variable in the model (p < 0.05).
The best SEM models comparing the native species with each of the invasive species indicated that soil bulk density, total N content and salinity best associated the invasions with the changes in SOC contents (Figure 9). A comparison of the soil data for C. malaccensis and P. australis communities suggested that invasive success was associated with a decrease in SOC content by its negative relationships with bulk density and N content of the C. malaccensis soil and not by a direct effect (Figure 10A). S. alterniflora success had no significant effect on total SOC content but the SEM showed that this invasive species had a negative effect on SOC content by its negative relationships with bulk density and salinity (Figure 10B). This indirect negative effect was counteracted by a direct positive effect to produce no total effect.
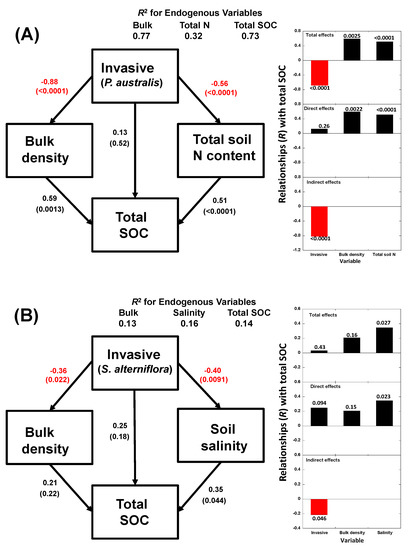
Figure 10.
Diagrams of the structural equation models that best explained the maximum variance of the SOC content in comparing Cyperus malaccensis (native) versus Phragmites australis (invasive) (A) and Cyperus malaccensis (native) versus Spartina alterniflora (invasive) (B) plant communities. Black and red arrows indicate positive and negative relationships, respectively.
4. Discussion
4.1. Changes of Carbon Fractions and N and P along Soil Profile
SOC, LOC, DOC and MBC concentrations and contents showed a slight decrease along the first 30 cm of soil profile and more or less constant concentrations and contents from 30 cm to 60 cm of soil depth. This pattern of decrease along the first 30 cm of soil depth was clearer in LFOC fraction. The results showed a slight reduction of soil organic concentrations and contents along the first 60 cm of soil profile. Only LFOC directly coming from leaf litter showed a clear decrease of contents in the 30–60 cm of soil depth with respect the first 30 cm of soil depth. The increase in the studied C:N ratios (LFOC:LFON and SOC:N) showed a great capacity of these soils to accumulate carbon in deep soil layers. Contrary to this result, most current studies show a decrease in C:N ratio with soil depth [50,51]. Thus, this interesting and particular result of these studied wetland soils is very probably related to the continuous sediment loadings that these soils received and the anoxic conditions that prevent the fast and complete decomposition of the litter, which is continuously covered by new sediments and progressively buried to deeper soil layers. A trend to uniform organic carbon contents along the vertical soil profile in wetlands has been observed in soils receiving continuous loads of sediments [37,52], which is related with the physical protection of soil organic carbon linked to sedimentation mainly in seasonally-flooded wetlands [53].
The effect of invasive species on vertical profile was often significant for soil variables. The clearest effect was for the higher concentrations and contents of LFOC and LFON at 30–60 cm of soil depth in both invasive than in the native species. This could be related with the higher LOC concentrations observed at 0–10 cm of soil depth also in both invasive species than in native ones, suggesting that both invasive species produce more LOC, probably by litter fall, which is then accumulated in deeper soil layers. The SOC content along soil profile was, however, similar between the native species and the invasive S. alternifolia but lower in the invasive P. australis than in the other both species. The data thus showed that the invasive effect on total soil C storing capacity would depend of the invasive species. In this case, P. australis seems to be the species less adequate to confer to the wetlands system a high capacity to act as C sink.
4.2. Relationships of Species Invasion with Soil C Concentrations and Contents
The invasive species had lower soil C concentrations, especially of the more labile organic-C fractions and mainly below 30 cm. The total amounts of soil C per unit of soil surface, however, were higher, with a trend of significantly higher total C accumulation in the soils of the native than the invasive species. This difference between C concentration with C content per unit of soil surface was related to the lower bulk density in the invasive than the native communities. Bulk density has been positively correlated with organic C, N and P concentrations in similar wetland areas in the soils of Carex sp. and P. australis communities [54]. Differences and changes in bulk density have also been frequently associated with the success of invasive plant species [55,56,57].
These changes in soil texture can be associated with the changes in plant structure between native and invasive species. A previous study reported higher total aboveground and total biomass in invasive than native species. Mean shoot biomass was 643, 599 and 1845 g m−2 for the native P. australis, C. malaccensis and the invasive S. alterniflora respectively [58]. Mean root biomass was 2322, 3168 and 2085 g m−2 for the native P. australis, C. malaccensis and the invasive S. alterniflora respectively [58]. Mean shoot: root ratios were thus 0.26, 0.19 and 0.88 [58]. These differences in the amount and distribution of biomass, with higher aboveground biomass in the invasive plants, may underlie the changes in soil texture and thereby total C, N and P contents, as demonstrated by the SEM analysis of total SOC content.
The C, N and P contents of biomass were not analyzed in this study but the much higher total biomass of both invasive species strongly suggest that some of these decreases in soil C, N and P contents were associated with increases in plant N and P contents from higher plant uptake and with a higher accumulation of C in the biomass. All three species were tall-grass plants of similar age, so the higher biomasses in the invasive species at the end of the growth period was likely due to higher growth rates in both invasive species than in the native species. Higher growth rates together with a decrease in soil N and P contents and the higher shoot:root biomass ratios of both invasive species suggest that invasive success was associated with higher nutrient uptake, proportional allocation of resources to aboveground biomass and growth rate. The data also strongly suggested that invasive plant success was associated with a large shift of N and P from soil to plants and with a change in soil C fractions from less to more labile forms in the invaded than the native stands.
This study did not determine whether the success of plant invasion in these wetlands involved higher phenotypic plasticity in taking up the available nutrients, which is expected in nutrient-rich sites [16,20,34], or involved more conservative strategies such as a higher nutrient-use efficiency, which is generally expected in nutrient-poor sites [19,20,21]. The invasive species in our study had a mix of properties between properties related to nutrient-rich sites with properties related to nutrient-poor sites. They had higher growth rates and shoot:root ratios, suggesting that the invasive species likely took up more nutrients, consistent with nutrient-rich sites where higher nutrient uptake increases growth and production [22]. The possible increase in the accumulation of nutrients in biomass and the decrease in soil contents for the invasive species, however, also suggests a higher nutrient-use and resorption efficiencies and also higher capacity for nutrient conservation in invasive species, consistent with nutrient-poor sites. These trends have been also observed in other wetland areas of the same estuarine region where C. malaccensis native communities have been invaded by Spartina alterniflora [59].
4.3. Similarities and Differences in the Impacts of Invasive Success between S. alterniflora and P. australis
The lower soil bulk densities in both invaded communities, relative to the native community, were mainly related to the different impacts on soil texture. The percentage of clay in the soils was in the order P. australis > C. malaccensis > S. alterniflora (37.6 ± 1.2, 34.2 ± 1.7 and 29.2 ± 1.1%, respectively), the percentage of sand was in the order S. alterniflora > C. malaccensis > P. australis (12.7 ± 0.9, 8.94 ± 1.03 and 6.18 ± 0.50%, respectively) but silt content did not differ significantly among the three monospecific stands. The highest biomass and densest root system coupled with the higher aboveground biomass of S. alterniflora was thus associated with the highest soil sand and lowest clay contents. In contrast, the lowest biomass and least dense root system of P. australis was associated with the highest clay and lowest sand contents. The success of both invasive species was thus associated with decreases in bulk density and total soil C, N and P contents per unit of surface area but these changes were associated with different changes in the soil textures for both invasive species.
These results suggest that the mechanisms underlying invasive plant success under the same conditions can differ with species. S. alterniflora had much higher total and aboveground biomasses than P. australis but also a higher root biomass and lower shoot:root ratio than P. australis. We were unable to clearly discern if these soil textures differed prior to plant invasion but the differences in biomass and in its distribution between aerial and root structures could be responsible for these textural differences, at least in part. As in other studies reporting differences in soil bulk density and elemental composition between invaded and native communities, the causes of these differences in our study were unclear. The differences may have existed prior to invasion, which may have favored alien success, or may be consequences of the invasion [60]. Anyway, previous studies on C. malaccensis communities conducted in other areas of the same Estuary showed soils with similar texture and nutrient contents those observed in C. malaccensis communities in the current study [59] suggesting that plant invasion can be related with the observed shifts in soil conditions.
The stands currently dominated by both invasive species were recently occupied by monospecific stands of the native C. malaccensis, strongly suggesting that all or most of the current soil differences were due to the invading species. Moreover, the soil properties were fairly homogeneous within each community but different between communities and the three communities were close to each other, also consistent with the hypothesis that the observed soil differences were completely or partially due to species invasion. This hypothesis is also reinforced by the fact that all the studied area has the same bedrock type (granite) and the studied sites are very close. Changes in soil traits such as SOC concentration [61], texture [62,63] and other soil variables [64] due to plant invasion have been correlated with differences in plant biomass and/or shoot:root ratio between native and invasive plants. In fact, soil properties such as texture can change in wetlands within 10 years, as observed in other studies [40,65]. Some studies have observed changes in soil bulk density [66], C contents [67,68,69,70] and several other traits [70,71] as a consequence of invasive plant success. The previously discussed similar SOC values along the soil vertical profile suggesting that these wetlands are sinks of sediments reinforce the idea that sites with different vegetation cover with different structure and shoot/root ratio can change soil structure by favoring sedimentation of different particle size classes.
Independent of any soil differences prior to or a consequence of species invasion, both invasive species differed in their requirements of and/or impacts on soil, indicating that different alien species can succeed under the same or very similar ecological conditions. Our study is also consistent with previous studies observing differences in soil texture related to invasive plant success [39,65,66,72]. The evidence from many studies generally suggest that the differences in soils in invaded relative to native communities are associated with the distinct structural and functional traits of the invasive and native species, such as nutrient uptake, N2-fixing capacity, litter production, foliar resorption or growth rate [73,74,75].
5. Conclusions
The invasive success of both species was associated with a decrease in soil bulk density and C stocks in one of the invasive species (P. australis) but not the other (S. alterniflora). Both invasive species had the concentrations and contents of LFOC and LFON at 30–60 cm of soil depth higher than native species. This could be related with the higher LOC concentrations observed at 0–10 cm of soil depth also in both invasive species than in native ones.
The invasive success of both species was associated with an increase in soil N and P concentrations in some soil layers but also with a decrease in total N and P contents due to the lower bulk densities in the soils of both invasive species relative to that in the soil of the native species.
The lower soil bulk densities for both invasive species were produced by different impacts on soil texture, an effect likely due to the different total biomasses and shoot:root ratios in the two invasive species. This result is very interesting, because it suggests that different invasive species with distinct mechanisms in the use of nutrients and with different consequences on nutrient plant-soil cycles can succeed under the same conditions. The traits conferring alien success are thus not necessarily specific by a determined site and also not species-specific.
Thus, the higher accumulation of C in soil and C and nutrients in plant suggest that the success of both invasive species was associated with higher growth rates and shoot:root ratios and likely with higher capacities to take up nutrients, all of which are expected in nutrient-rich sites where a higher nutrient uptake is used to increase growth and production. The possible increase in the accumulation of nutrients in biomass in invasive species, however, also suggests a higher capacity for nutrient conservation by retaining nutrients in plant-soil system.
Acknowledgments
This work was supported by grants from the National Science Foundation of China (41571287), Natural Science Foundation Key Programs of Fujian Province (2018R1101006-1), Fujian Provincial Outstanding Young Scientists Program (2017), the European Research Council Synergy grant ERC-2013-SyG-610028 IMBALANCE-P, the Spanish Government grant CGL2013-48074-P and the Catalan Government grant SGR 2014-274.
Author Contributions
W.W. and J.S. conceived and designed the experiments; W.W. and C.W. performed the experiments; W.W. and J.S. analyzed the data; W.W., J.S., J.P. and C.W. contributed reagents/materials/analysis tools; all the authors contribute to write the paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- International Panel on Climate Change. Global Carbon Cycle Overview. Summary for Police Makers. In Land Use, Land-Use Change and Forestry; IPCC special Reports; Watson, R.T., Noble, I.R., Bolin, B., Ravindranath, N.H., Verarde, D.J., Dokken, D.J., Eds.; IPCC: Geneva, Switzerland, 2000; p. 375. [Google Scholar]
- Joiner, D.W.; Lafleur, P.M.; McCaughey, J.H.; Bartlett, P.A. Interannual variability in carbon dioxide exchanges at a boreal wetland in the BOREAS northern study area. J. Geophys. Res. 1999, 104, 27663–27672. [Google Scholar] [CrossRef]
- Smith, P. Soils as carbon sinks: The global context. Soil Use Manag. 2004, 20, 212–218. [Google Scholar] [CrossRef]
- Bonan, G.B.; van Cleve, K. Soil temperature, nitrogen mineralization and carbon source-sink relationships in boreal forests. Can. J. For. Res. 2011, 22, 629–639. [Google Scholar] [CrossRef]
- Bianchi, T.S.; Allison, M.A.; Zhao, J.; Li, X.X.; Comeaux, R.S.; Feagin, R.A.; Kulawardhana, R.W. Historical reconstruction of mangrove expansion in the Gulf of Mexico: Linking climate change with carbon sequestration in coastal wetlands. Estuar. Coast. Shelf Sci. 2013, 119, 7–16. [Google Scholar] [CrossRef]
- Burden, A.; Garbutt, R.A.; Evans, C.D.; Jones, D.L.; Cooper, D.M. Carbon sequestration and biogeochemical cycling in a saltmarsh subject to coastal managed realignment. Estuar. Coast. Shelf Sci. 2013, 120, 12–20. [Google Scholar] [CrossRef]
- Mitsch, W.J.; Bernal, B.; Nahlik, A.M.; Mander, U.; Zhang, L.; Anderson, C.J.; Jorgensen, S.E.; Brix, H. Wetlands, carbon, and climate change. Landsc. Ecol. 2013, 28, 583–597. [Google Scholar] [CrossRef]
- Wetland China. The Report on the Second National Wetland Resources Survey (2009–2013). 2014. Available online: http://www.shidi.org/zt/2014xwfbh/ (accessed on 12 June 2017).
- Liu, J.Q.; Zeng, C.S.; Chen, N. Research of Minjiang River Estuary Wetland; Science Press: Beijing, China, 2006. [Google Scholar]
- Wang, W.; Sardans, J.; Zeng, C-S.; Tong, C.; Peñuelas, J. Impact of plant invasion and increasing floods on total soil phosphorus and its fractions in the Minjiang river estuarine wetlands, China. Wetlands 2016, 36, 21–36. [Google Scholar] [CrossRef]
- Schewe, J.; Levermann, A.; Meinshausen, M. Climate change under a scenario near 1.5 degrees C of global warming: Monsoon intensification, ocean warming and stric sea level rise. Earth Syst. Dyn. 2011, 2, 25–35. [Google Scholar] [CrossRef]
- Mendelsohn, R.; Emanuel, K.; Chonabayashi, S.; Bakkensen, L. The impact of climate change on tropical cyclone damage. Nat. Clim. Chang. 2012, 2, 205–209. [Google Scholar] [CrossRef]
- Sardans, J.; Rivas-Ubach, A.; Peñuelas, J. The C:N:P stoichiometry of organisms and ecosystems in a changing world: A review and perspectives. Perspect. Plant Ecol. Evol. Syst. 2012, 14, 33–47. [Google Scholar] [CrossRef]
- Ramsar. The Ramsar Manual, 6th ed.; Ramsar Convention Secretariat: Gland, Switzerland, 2013; p. 110. [Google Scholar]
- Piecuch, C.G.; Ponte, R.M. Mechanisms of global-mean steric sea level change. J. Clim. 2014, 27, 824–834. [Google Scholar] [CrossRef]
- Sardans, J.; Peñuelas, J. The role of plants in the effects of Global Change on nutrient availability and stoichiometry in the plant-soil system. Plant Physiol. 2012, 160, 1741–1761. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Sardans, J.; Zeng, C.; Zhong, C.; Li, Y.; Peñuelas, J. Responses of soil nutrient concentrations and stoichiometry to different human land uses in a subtropical tidal wetland. Geoderma 2014, 232–234, 459–470. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wang, C.; Sardans, J.; Tong, C.; Jia, R.; Zeng, C.; Peñuelas, J. Flood regime affects soil stoichiometry and the distribution of the invasive plants in subtropical estuarine wetlands in China. Catena 2015, 128, 144–154. [Google Scholar] [CrossRef]
- Funk, J.L.; Vitousek, P.M. Resource-use efficiency and plant invasion in low-resource systems. Nature 2007, 446, 1079–1081. [Google Scholar] [CrossRef] [PubMed]
- González, A.L.; Kominoski, J.S.; Danger, M.; Ishida, S.; Iwai, N.; Rubach, A. Can ecological stoichiometry help explain patterns of biological invasion? Oikos 2010, 119, 779–790. [Google Scholar] [CrossRef]
- Matzek, V. Superior performance and nutrient-use efficiency of invasive plants over non-invasive congeners in a resource-limited environment. Biol. Invasions 2011, 13, 3005–3014. [Google Scholar] [CrossRef]
- Sardans, J.; Bartrons, M.; Margalef, O.; Gargallo-Garriga, A.; Janssens, I.A.; Ciais, P.; Obersteiner, M.; Sigurdsson, B.D.; Chen, H.Y.; Peñuelas, J. Plant invasion is associated with higher plant-soil nutrient concentrations in nutrient-poor environments. Glob. Chang. Biol. 2017, 23, 1282–1291. [Google Scholar] [CrossRef] [PubMed]
- Wissing, L.; Kölbl, A.; Vogelsang, V.; Fu, J.; Cao, Z.; Kögel-Knabner, I. Organic carbon accumulation in a 2000-year chronosequence of paddy soil evolution. Catena 2011, 87, 376–385. [Google Scholar] [CrossRef]
- Kimura, M.; Murase, J.; Lu, Y. Carbon cycling in rice field ecosystems in the context of input, decomposition and translocation of organic materials and the fates of their end products (CO2 and CH4). Soil Biol. Biochem. 2004, 36, 1399–1416. [Google Scholar] [CrossRef]
- Chen, H.L.; Zhou, J.M.; Xiao, B.H. Characterization of dissolved organic matter derived from rice straw at different stages of decay. J. Soil Sediment. 2010, 10, 915–922. [Google Scholar] [CrossRef]
- Wang, W.; Lai, D.Y.F.; Wang, C.; Pan, T.; Zeng, C. Effects of rice straw incorporation on active soil organic carbon pools in a subtropical paddy field. Soil Tillage Res. 2015, 152, 8–16. [Google Scholar] [CrossRef]
- Gu, L.; Post, W.M.; King, A.W. Fast labile carbon turnover obscures sensitivity of heterotrophic respiration from soil to temperature: A model analysis. Glob. Biogeochem. Cycles 2004, 18. [Google Scholar] [CrossRef]
- Xu, M.; Lou, Y.; Sun, X.; Wang, W.; Baniyamuddin, M.; Zhao, K. Soil organic carbon active fractions as early indicators for total carbon change under straw incorporation. Biol. Fertil. Soils 2011, 47, 745–752. [Google Scholar] [CrossRef]
- Zhao, F.; Yang, G.; Han, X.; Feng, Y.; Ren, G. Stratification of carbon fractions and carbon management index in deep soil affected by the Grain-to-Green Program in China. PLoS ONE 2014, 9, e99657. [Google Scholar] [CrossRef] [PubMed]
- Knoblauch, C.; Maarifat, A.A.; Pfeiffer, E.M.; Faefele, S.M. Degradability of black carbon and its impact on trace gas fluxes and carbon turnover in paddy soils. Soil Biol. Biochem. 2011, 43, 1768–1778. [Google Scholar] [CrossRef]
- Hanke, A.; Cerli, C.; Muhr, J.; Borken, W.; Kalbitz, K. Redox control on carbon mineralization and dissolved organic matter along a chronosequence of paddy soils. Eur. J. Soil Sci. 2013, 64, 476–487. [Google Scholar] [CrossRef]
- Peñuelas, J.; Sardans, J.; Rivas-Ubach, A.; Janssens, I.A. The human-induced imbalance between C, N and P in Earth’s life system. Glob. Chang. Biol. 2012, 189, 5–8. [Google Scholar] [CrossRef]
- Peñuelas, J.; Poulter, B.; Sardans, J.; Ciais, P.; van der Velde, M.; Bopp, L.; Boucher, O.; Godderis, Y.; Llusià, J.; Nardin, E.; et al. Human-induced nitrogen-Phosphorus imbalances alter natural and managed ecosystems across the globe. Nat. Commun. 2013, 4, 2934. [Google Scholar] [CrossRef] [PubMed]
- Daehler, C.C. Performance comparisons of co-occuring native and alien invasive plants: Implications for conservation and restoration. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 183–211. [Google Scholar] [CrossRef]
- Tian, H.; Chen, G.; Zhang, C.; Melillo, J.; Hall, C. Pattern and variation of C: N: P ratios in China’s soils: A synthesis of observational data. Biogeochemistry 2010, 98, 139–151. [Google Scholar] [CrossRef]
- Schipper, L.A.; Sparling, G.P. Accumulation of soil organic C and change in C:N ratio after establishment of pastures on reverted scrubland in New Zealand. Biogeochemistry 2011, 104, 49–58. [Google Scholar] [CrossRef]
- Senga, Y.; Hiroki, M.; Nakamura, Y.; Watarai, Y.; Watanable, Y.; Nohara, S. Vertical profiles of DIN, DOC, and microbial activities in the wetland soil of Kushiro mire, northeastern Japan. Limnology 2011, 12, 17–23. [Google Scholar] [CrossRef]
- Xiang, J.; Liu, D.Y.; Ding, W.X.; Yuan, J.J.; Lin, Y.X. Invasion chronosequence of Spartina alterniflora on methane emission and organic carbon sequestration in a coastal salt marsh. Atmos. Environ. 2015, 112, 72–80. [Google Scholar] [CrossRef]
- Craft, C.; Reader, J.; Sacco, J.N.; Broome, S.W. Twenty-five years of ecosystem development of constructed Spartina alterniflora marshes. Ecol. Appl. 1999, 9, 1405–1419. [Google Scholar] [CrossRef]
- Zheng, C.H.; Zeng, C.S.; Chen, Z.Q.; Lin, M.C. A study on the changes of landscape pattern of estuary wetlands of the Minjiang River. Wetl. Sci. 2006, 4, 29–34. [Google Scholar]
- Tong, C.; Liu, B.G. Litter decomposition and nutrient dynamics in different tidal water submergence environments of estuarine tidal wetland. Geogr. Res. 2009, 28, 118–128. [Google Scholar]
- Jia, R.X.; Tong, C.; Wang, W.Q.; Zeng, C.S. Organic carbon concentrations and storages in the salt marsh sediments in the Min River estuary. Wetl. Sci. 2008, 6, 492–499. [Google Scholar]
- Sorrell, B.; Brix, H.; Schierup, H.H.; Lorenzen, B. Die-back of Phragmites australis: Influence on the distribution and rate of sediment methanogenesis. Biogeochemistry 1997, 36, 173–188. [Google Scholar] [CrossRef]
- Bai, J.H.; Yang, H.O.; Deng, W.; Zhu, Y.M.; Zhang, X.L.; Wang, Q.G. Spatial distribution characteristics of organic matter and total nitrogen of marsh soils in river marginal wetlands. Geoderma 2005, 124, 181–192. [Google Scholar] [CrossRef]
- Lu, R.K. Analysis Methods of Soil Science and Agricultural Chemistry; Agriculture Science and Technology Press: Beijing, China, 1999. [Google Scholar]
- Raamsdonk, L.M.; Teusink, B.; Broadhurst, D.; Zhang, N.S.; Hayes, A.; Walsh, M.C.; Berden, J.A.; Brudle, K.M.; Kell, D.K.; Rowland, J.J.; et al. A functional genomics strategy that uses metabolome data to reveal the phenotype of silent mutations. Nat. Biotechnol. 2001, 19, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Fox, J.; Nie, Z.; Byrnes, J. SEM: Structural Equation Models. 2013. Available online: http://cran.r-project.org/package=sem (accessed on 24 July 2017).
- Davison, A.C.; Hinkley, D.V.; Schechtman, E. Efficient Bootstrap Simulation. Biometrika 1986, 73, 555–566. [Google Scholar] [CrossRef]
- Mitchell-Olds, T. Jackknife, bootstrap and other resampling methods in regression analysis. Ann. Stat. 1986, 14, 1316–1318. [Google Scholar] [CrossRef]
- Lu, G.; Li, Z.; Zhao, X.; Jia, Q.Y.; Xie, Y.; Zhou, G. Vertical distribution of soil organic carbon and total nitrogen in reed wetland. Chin. J. Appl. Ecol. 2006, 17, 384–389. [Google Scholar]
- Ostrowska, A.; Porebska, G. Assessment of the C/N ratio as indicator of the decomposability of organic matter in forest soils. Ecol. Indic. 2015, 49, 104–109. [Google Scholar] [CrossRef]
- Alongi, D.M.; Wattayakorn, G.; Pfitzner, J.; Tirendi, F.; Zagorskis, I.; Brunskill, G.J.; Davidson, A.; Clough, B.F. Organic carbon accumulation and metabolic pathways in sediments of mangrove forests in southern Thailand. Mar. Geol. 2001, 179, 85–103. [Google Scholar] [CrossRef]
- Maynard, J.J.; Dahlgren, R.A.; O’Geen, A.T. Soil carbon cycling and sequestration in a seasonally satured wetland receiving agricultural runoff. Biogeosciences 2011, 8, 3391–3406. [Google Scholar] [CrossRef]
- Peng, P.; Zhang, W.; Tong, C.; Qiu, S.; Zhang, W. Soil C, N and P contents and their relationships with soil physical properties in wetlands of Dongting lake floodplain. Chin. J. Ecol. 2005, 16, 1872–1878. [Google Scholar]
- Miller, M.E.; Belnap, J.; Beattty, S.W.; Reynolds, R.L. Performance of Bromus tectorum L. in relation to soil properties, water additions, and chemical amendments in calcareous soils of southwestern Utah, USA. Plant Soil 2006, 288, 1–18. [Google Scholar] [CrossRef]
- Lortie, C.J.; Cushman, J.H. Effects of a directional abiotic gradient on plant community dynamics and invasion in a coastal dune system. J. Ecol. 2007, 95, 468–481. [Google Scholar] [CrossRef]
- Pande, A.; Williams, C.L.; Lant, C.L.; Gibson, D.J. Using map algebra to determine the mesoscale distribution of invasive plants: The case of Celastrus orbicularis in Southern Illinois, USA. Biol. Invasions 2007, 9, 419–431. [Google Scholar] [CrossRef]
- Tong, C.; Zhang, L.; Wang, W.; Gauci, V.; Marrs, R.; Liu, B.; Jia, R.; Zeng, C. Contrasting nutrient stocks and litter decomposition in stands of native and invasive species in a sub-tropical estuarine marsh. Environ. Res. 2011, 111, 909–916. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wang, C.; Sardans, J.; Zeng, C.; Tong, C.; Peñuelas, J. Plant invasive success associated with higher N-use efficiency and stoichiometric shifts in the soil–plant system in the Minjiang River tidal estuarine wetlands of China. Wetl. Ecol. Manag. 2015, 23, 865–880. [Google Scholar] [CrossRef]
- Boxwell, J.; Drohan, P.J. Surface soil physical and hydrological characteristics in Bromus tectorum L. (cheatgrass) versus Artemisia tridentate Nutt. (big sagebrush) habitat. Geoderma 2009, 149, 305–311. [Google Scholar] [CrossRef]
- Zhou, L.Y.; Yin, S.L.; An, S.Q.; Yang, W.; Deng, Q.; Xie, D.; Ji, H.T.; Ouyang, Y.; Cheng, X.L. Spartina alterniflora invasion alters carbon exchange and soil organic carbon in eastern salt Marsh of China. Clean-Soil Air Water 2015, 43, 569–576. [Google Scholar] [CrossRef]
- Uselman, S.M.; Synder, K.A.; Leger, E.A.; Duke, S.E. First-Year establishment, biomass and seed production of early vs. late seral natives in two medusahead (Taeniatherum caput-medusae) invaded soils. Invasive Plant Sci. Manag. 2014, 7, 291–302. [Google Scholar] [CrossRef]
- Haubensak, K.A.; D’Antonio, C.M.; Embry, S.; Blank, R. A comparison of Bromus tectorum growth and mycorrhizal colonization in salt deserts vs. sagebrush habitats. Rangel. Ecol. Manag. 2014, 67, 275–284. [Google Scholar] [CrossRef]
- Blank, R.R.; Morgan, T.; Allen, F. Suppression of annual Bromus tectorum by perennial Agropyron cristatus: Roles of soil nitrogen availability and biological soil space. AoB Plants 2015, 7. [Google Scholar] [CrossRef] [PubMed]
- Craft, C.; Broome, S.; Campbell, C. Fifteen years of vegetation and soil development after brackish-water marsh creation. Restor. Ecol. 2002, 10, 248–258. [Google Scholar] [CrossRef]
- Zhang, C.B.; Wang, J.; Qian, B.Y.; Li, W.H. Effects of the invader Solidago Canadensis on soil properties. Appl. Soil Ecol. 2009, 43, 163–169. [Google Scholar] [CrossRef]
- Koutika, L.S.; Vanderhoeven, S.; Chapius-Lardy, L.; Dassonville, N.; Meerts, P. Assessment of changes in soil organic matter after invasion by exotic plant species. Biol. Fertil. Soils 2007, 44, 331–341. [Google Scholar] [CrossRef]
- Koteen, L.E.; Baldocchi, D.D.; Harte, J. Invasion of non-native grasses causes a drop in soil carbon storage in California grasslands. Environ. Res. Lett. 2011, 6, 044001. [Google Scholar] [CrossRef]
- Throop, H.L.; Laitha, K.; Kramer, M. Density fractionation and 13C reveal changes in soil carbon following woody encoarchment in a desert ecosystem. Biogeochemistry 2013, 112, 409–422. [Google Scholar] [CrossRef]
- Yu, F.K.; Huang, X.H.; Duan, C.Q.; He, S.Z.; Zhang, G.S.; Liu, C.E.; Fu, D.G.; Shao, H.B. Impacts of Ageratina adenophora invasion on soil physical-chemical properties of Eucalyptus plantation and implications for constructing agro-forest ecosystem. Ecol. Eng. 2014, 64, 130–135. [Google Scholar] [CrossRef]
- Souza-Alonso, P.; Guisande-Collazo, A.; Gonzalez, L. Gradualism in Acacia dealbata link invasion: Impacts on soil chemistry and microbial community over a chronological sequence. Soil Biol. Biochem. 2015, 80, 315–323. [Google Scholar] [CrossRef]
- Surrette, S.B.; Brewer, J.S. Inferring relationships between native plant diversity and Lonicera japonica in upland forests in north Mississippi, USA. Appl. Veg. Sci. 2008, 11, 205–214. [Google Scholar] [CrossRef]
- Lindsay, E.A.; French, K. Litterfall and nitrogen cycling following invasion by Chrysanthemoides moniifera ssp. rotundata in coastal Australia. J. Appl. Ecol. 2005, 42, 556–566. [Google Scholar] [CrossRef]
- Titus, J.H. Nitrogen-fixers Alnus and Lupinus influence soil characteristics but not colonization by later successional species in primary succession on Mount St. Helens. Plant Ecol. 2009, 203, 289–301. [Google Scholar] [CrossRef]
- Novoa, A.; Rodríguez, R.; Richardson, D.; Gonzàlez, L. Soil quality: A key factor in understanding plant invasión? The case of Capobrotus edulis (L.) N.E.Br. Biol. Invasions 2014, 16, 429–443. [Google Scholar] [CrossRef]
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).