Notch-1 Immunopositivity in Brain Lesions Associated with Pharmacoresistant Epilepsy

 , , , ,
, , , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Immunohistochemistry
2.3. Image Analysis
2.4. Classification of Different Malformations
3. Results
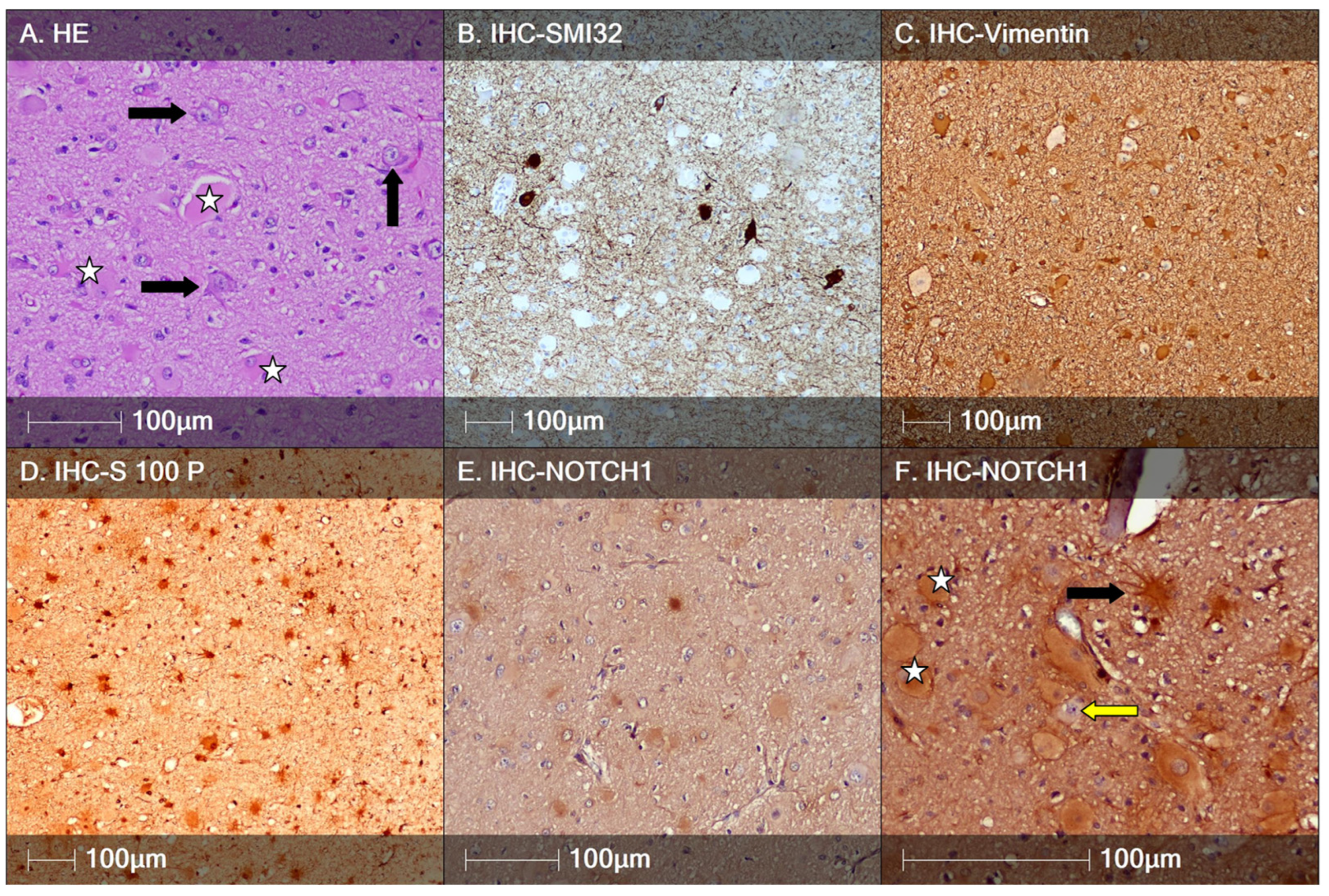
3.1. Focal Cortical Dysplasia Type II
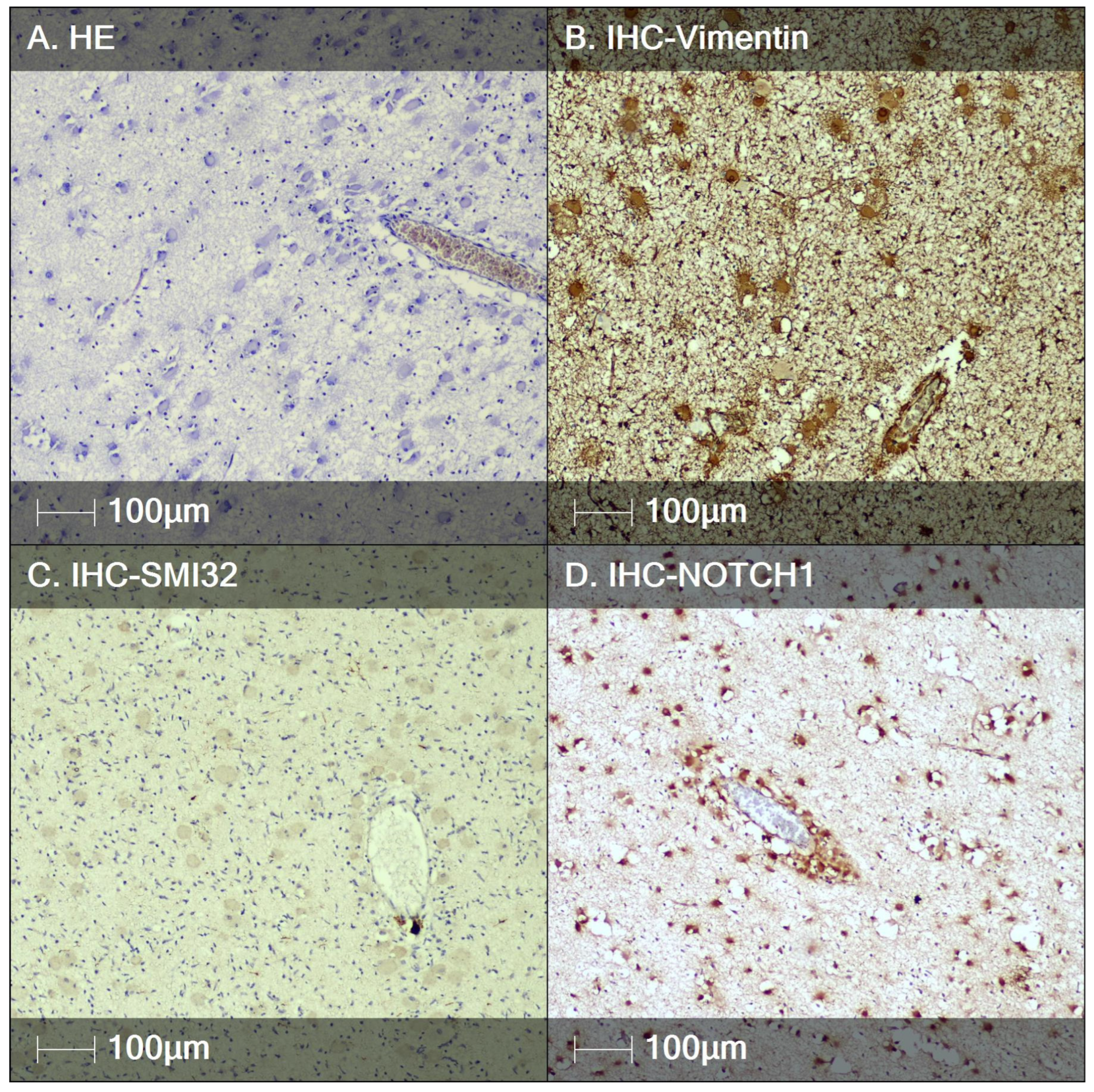
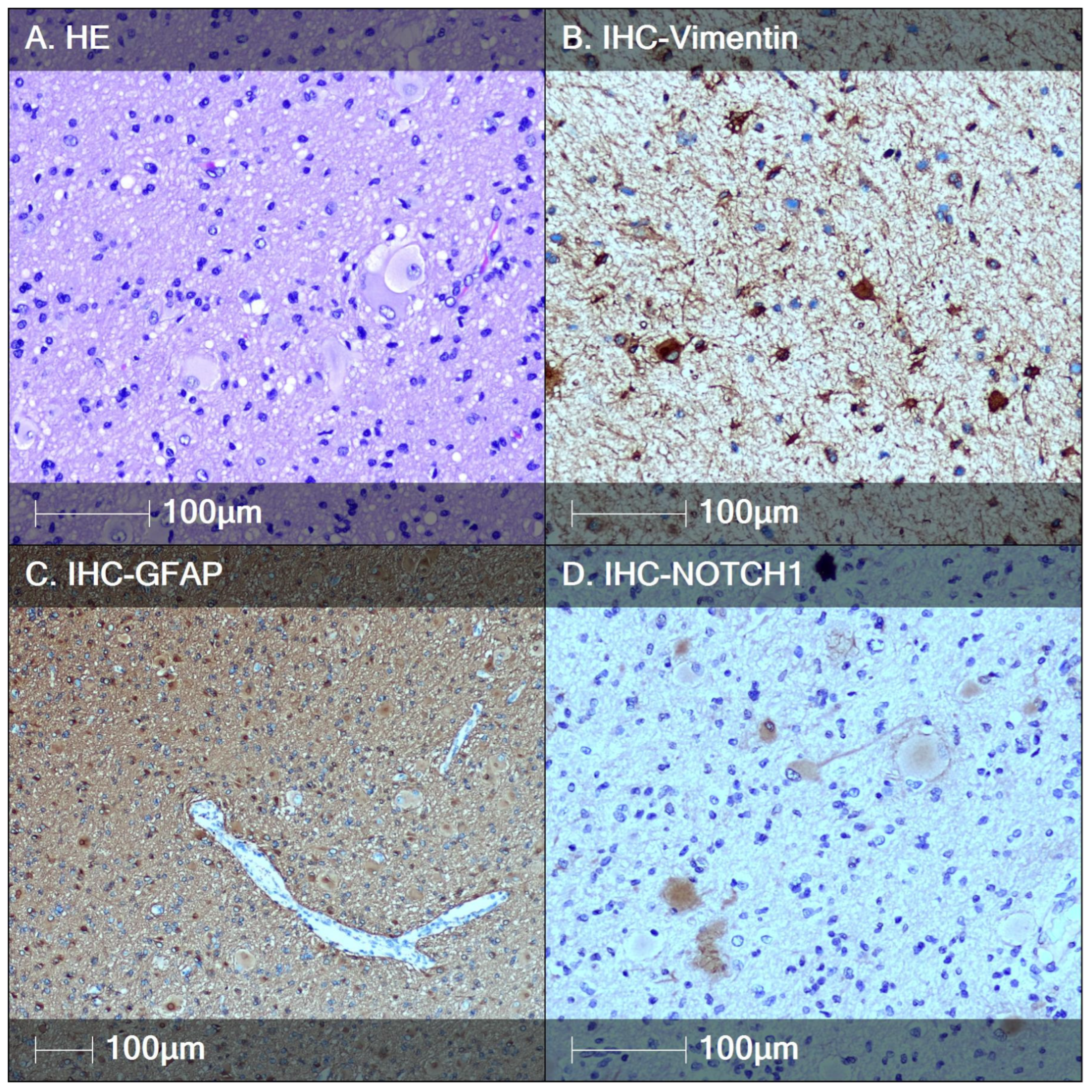
3.2. Tuberous Sclerosis (TS)
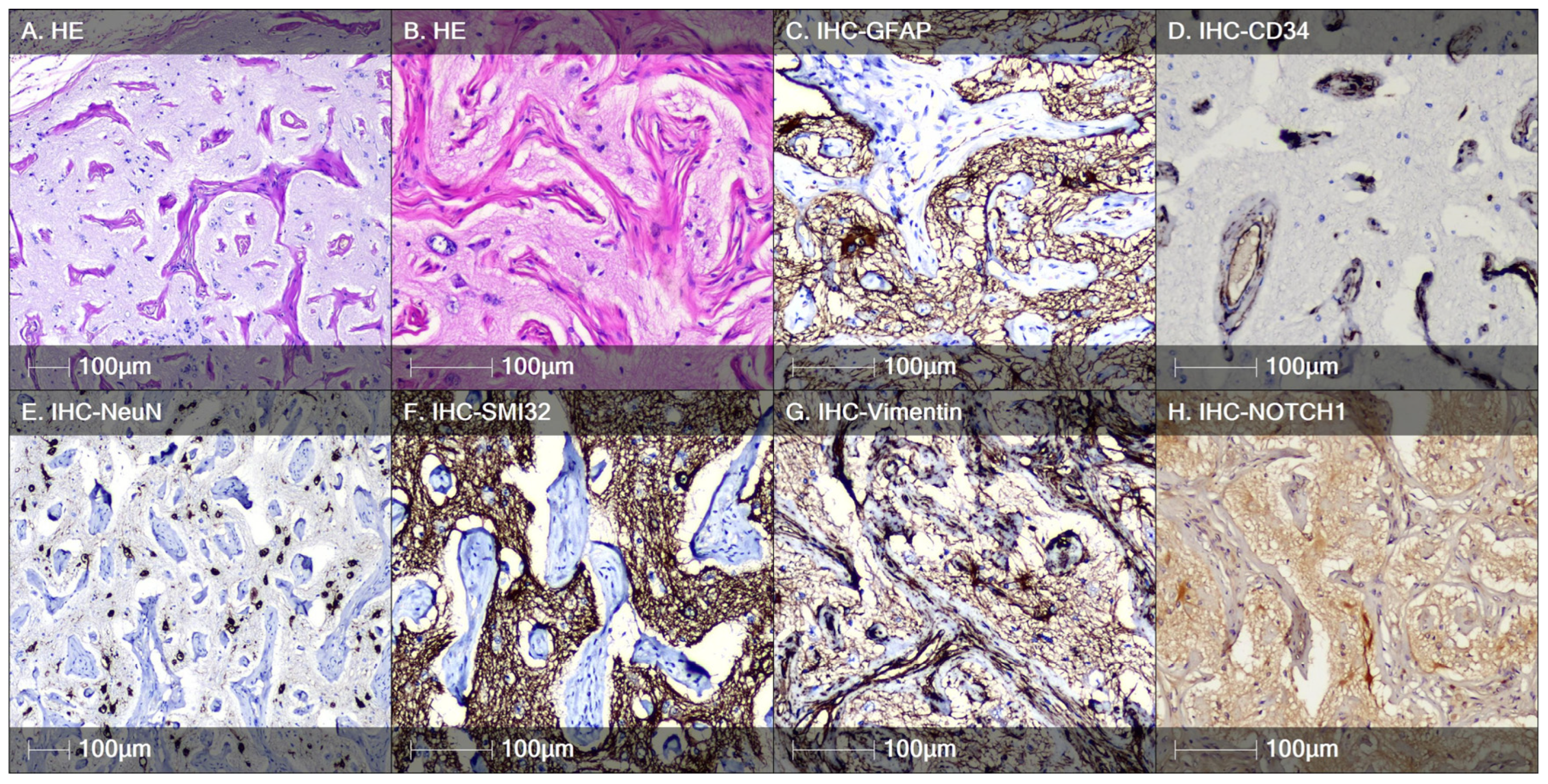
3.3. Case of Meningioangiomatosis
3.4. Quantification Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sultana, B.; Panzini, M.A.; Carpentier, A.V.; Comtois, J.; Rioux, B.; Gore, G.; Bauer, P.R.; Kwon, C.-S.; Jetté, N.; Josephson, C.B.; et al. Incidence and Prevalence of Drug-Resistant Epilepsy. Neurology 2021, 96, 805–817. [Google Scholar] [CrossRef]
- Barkovich, A.J.; Dobyns, W.B.; Guerrini, R. Malformations of cortical development and epilepsy. Cold Spring Harb. Perspect. Med. 2015, 5, a022392. [Google Scholar] [CrossRef] [PubMed]
- Wirrell, E.C.; Wong-Kisiel, L.C.L.; Mandrekar, J.; Nickels, K.C. What predicts enduring intractability in children who appear medically intractable in the first 2 years after diagnosis? Epilepsia 2013, 54, 1056–1064. [Google Scholar] [CrossRef] [PubMed]
- Pasquier, B.; Péoc’H, M.; Fabre-Bocquentin, B.; Bensaadi, L.; Pasquier, D.; Hoffmann, D.; Kahane, P.; Tassi, L.; Le Bas, J.-F.; Benabid, A.L. Surgical pathology of drug-resistant partial epilepsy. A 10-year-experience with a series of 327 consecutive resections. Epileptic Disord. 2002, 4, 99–119. [Google Scholar] [CrossRef]
- Severino, M.; Geraldo, A.F.; Utz, N.; Tortora, D.; Pogledic, I.; Klonowski, W.; Triulzi, F.; Arrigoni, F.; Mankad, K.; Leventer, R.J.; et al. Definitions and classification of malformations of cortical development: Practical guidelines. Brain 2020, 143, 2874–2894. [Google Scholar] [CrossRef]
- Northrup, H.; Arronow, M.E.; Bebin, M.; Bissler, J.; Darling, T.N.; de Vries, P.J.; Frost, M.D.; Gosnell, E.S.; Gupta, N.; Jansen, A.C.; et al. Updated International Tuberous Sclerosis Complex Diagnostic Criteria and Surveillance and Management Recommendations. Pediatr. Neurol. 2021, 123, 50–66. [Google Scholar] [CrossRef]
- Najm, I.; Lal, D.; Vanegas, M.A.; Cendes, F.; Lopes-Cendes, I.; Palmini, A.; Paglioli, E.; Sarnat, H.B.; Walsh, C.A.; Wiebe, S.; et al. The ILAE consensus classification of focal cortical dysplasia: An update proposed by an ad hoc task force of the ILAE diagnostic methods commission. Epilepsia 2022, 63, 1899–1919. [Google Scholar] [CrossRef] [PubMed]
- Rossini, L.; Villani, F.; Granata, T.; Tassi, L.; Tringali, G.; Cardinale, F.; Aronica, E.; Spreafico, R.; Garbelli, R. FCD Type II and mTOR pathway: Evidence for different mechanisms involved in the pathogenesis of dysmorphic neurons. Epilepsy Res. 2017, 129, 146–156. [Google Scholar] [CrossRef]
- Arruda, I.L.; Arruda, R.F.; da Silveira, R.M.B.; Duarte, J.T.C.; Guaranha, M.S.B.; Guilhoto, L.M.; Júnior, H.C.; Stavale, J.N.; Centeno, R.S.; Yacubian, E.M.T.; et al. A controversial question: Can morphometry and clinical history be enough to diagnose hippocampal dysplasia? Epileptic Disord. 2024, 26, 382–391. [Google Scholar] [CrossRef] [PubMed]
- Ruppe, V.; Dilsiz, P.; Reiss, C.S.; Carlson, C.; Devinsky, O.; Zagzag, D.; Weiner, H.L.; Talos, D.M. Developmental brain abnormalities in tuberous sclerosis complex: A comparative tissue analysis of cortical tubers and perituberal cortex. Epilepsia 2014, 55, 539–550. [Google Scholar] [CrossRef]
- Mizuguchi, M.; Takashima, S. Neuropathology of tuberous sclerosis. Brain Dev. 2001, 23, 508–515. [Google Scholar] [CrossRef]
- Grajkowska, W.; Kotulska, K.; Jurkiewicz, E.; Matyja, E. Brain lesions in tuberous sclerosis complex. Review. Folia Neuropathol. 2010, 48, 139–149. [Google Scholar] [PubMed]
- Talos, D.M.; Kwiatkowski, D.J.; Cordero, K.; Black, P.M.; Jensen, F.E. Cell-specific alterations of glutamate receptor expression in tuberous sclerosis complex cortical tubers. Ann. Neurol. 2008, 63, 454–465. [Google Scholar] [CrossRef] [PubMed]
- Orlova, K.A.; Crino, P.B. The tuberous sclerosis complex. Ann. New York Acad. Sci. 2010, 1184, 87–105. [Google Scholar] [CrossRef]
- Lampada, A.; Taylor, V. Notch signaling as a master regulator of adult neurogenesis. Front. Neurosci. 2023, 17, 1179011. [Google Scholar] [CrossRef]
- Zhao, N.; Guo, Y.; Zhang, M.; Lin, L.; Zheng, Z. Akt-mTOR signaling is involved in Notch-1-mediated glioma cell survival and proliferation. Oncol. Rep. 2010, 23, 1443–1447. [Google Scholar] [CrossRef]
- Xu, P.; Qiu, M.; Zhang, Z.; Kang, C.; Jiang, R.; Jia, Z.; Wang, G.; Jiang, H.; Pu, P. The oncogenic roles of Notch1 in astrocytic gliomas in vitro and in vivo. J. Neurooncol. 2010, 97, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Hales, E.C.; Taub, J.W.; Matherly, L.H. New insights into Notch1 regulation of the PI3K-AKT-mTOR1 signaling axis: Targeted therapy of gamma-secretase inhibitor resistant T-cell acute lymphoblastic leukemia. Cell Signal. 2014, 26, 149–161. [Google Scholar] [CrossRef]
- Yi, L.; Zhou, X.; Li, T.; Liu, P.; Hai, L.; Tong, L.; Ma, H.; Tao, Z.; Xie, Y.; Zhang, C.; et al. Notch1 signaling pathway promotes invasion, self-renewal and growth of glioma initiating cells via modulating chemokine system CXCL12/CXCR4. J. Exp. Clin. Cancer Res. 2019, 38, 339. [Google Scholar] [CrossRef] [PubMed]
- Kunnimalaiyaan, M.; Chen, H. Tumor suppressor role of Notch-1 signaling in neuroendocrine tumors. Oncologist 2007, 12, 535–542. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Wu, S.; Wu, H.; Liu, L.; Zhou, J. Effect of the Notch1-mediated PI3K-Akt-mTOR pathway in human osteosarcoma. Aging 2021, 13, 21090–21101. [Google Scholar] [CrossRef]
- Sibbe, M.; Häussler, U.; Dieni, S.; Althof, D.; Haas, C.A.; Frotscher, M. Experimental epilepsy affects Notch1 signalling and the stem cell pool in the dentate gyrus. Eur. J. Neurosci. 2012, 36, 3643–3652. [Google Scholar] [CrossRef]
- Liu, X.; Yang, Z.; Yin, Y.; Deng, X. Increased expression of Notch1 in temporal lobe epilepsy: Animal models and clinical evidence. Neural Regen. Res. 2014, 9, 526–533. [Google Scholar] [CrossRef] [PubMed]
- Saffarzadeh, F.; Modarres Mousavi, S.M.; Lotfinia, A.A.; Alipour, F.; Hosseini Ravandi, H.; Karimzadeh, F. Discrepancies of Notch 1 receptor during development of chronic seizures. J. Cell Physiol. 2019, 234, 13773–13780. [Google Scholar] [CrossRef] [PubMed]
- Sha, L.; Wu, X.; Yao, Y.; Wen, B.; Feng, J.; Sha, Z.; Wang, X.; Xing, X.; Dou, W.; Jin, L.; et al. Notch signaling activation promotes seizure activity in temporal lobe epilepsy. Mol. Neurobiol. 2014, 49, 633–644. [Google Scholar] [CrossRef]
- Sun, C.; Fu, J.; Qu, Z.; Li, D.; Si, P.; Qiao, Q.; Zhang, W.; Xue, Y.; Zhen, J.; Wang, W. Chronic mild hypoxia promotes hippocampal neurogenesis involving Notch1 signaling in epileptic rats. Brain Res. 2019, 1714, 88–98. [Google Scholar] [CrossRef]
- Ganeva, R.; Parvanov, D.; Vidolova, N.; Ruseva, M.; Handzhiyska, M.; Arsov, K.; Decheva, I.; Metodiev, D.; Moskova-Doumanova, V.; Stamenov, G. Endometrial immune cell ratios and implantation success in patients with recurrent implantation failure. J. Reprod. Immunol. 2023, 156, 103816. [Google Scholar] [CrossRef] [PubMed]
- Rakic, P.; Knyihar-Csillik, E.; Csillik, B. Polarity of microtubule assemblies during neuronal cell migration. Proc. Natl. Acad. Sci. USA 1996, 93, 9218–9222. [Google Scholar] [CrossRef] [PubMed]
- Mathern, G.W.; Cepeda, C.; Hurst, R.S.; Flores-Hernandez, J.; Mendoza, D.; Levine, M.S. Neurons recorded from pediatric epilepsy surgery patients with cortical dysplasia. Epilepsia 2000, 41, S162–S167. [Google Scholar] [CrossRef] [PubMed]
- Cepeda, C.; Hurst, R.S.; Flores-Hernández, J.; Hernández-Echeagaray, E.; Klapstein, G.J.; Boylan, M.K.; Calvert, C.R.; Jocoy, E.L.; Nguyen, O.K.; André, V.M.; et al. Morphological and electrophysiological characterization of abnormal cell types in pediatric cortical dysplasia. J. Neurosci. Res. 2003, 72, 472–486. [Google Scholar] [CrossRef] [PubMed]
- Blümcke, I.; Aronica, E.; Miyata, H.; Sarnat, H.B.; Thom, M.; Roessler, K.; Rydenhag, B.; Jehi, L.; Krsek, P.; Wiebe, S.; et al. International recommendation for a comprehensive neuropathologic workup of epilepsy surgery brain tissue: A consensus Task Force report from the ILAE Commission on Diagnostic Methods. Epilepsia 2016, 57, 348–358. [Google Scholar] [CrossRef] [PubMed]
- Blümcke, I.; Thom, M.; Aronica, E.; Armstrong, D.D.; Vinters, H.V.; Palmini, A.; Jacques, T.S.; Avanzini, G.; Barkovich, A.J.; Battaglia, G.; et al. The clinicopathologic spectrum of focal cortical dysplasias: A consensus classification proposed by an ad hoc Task Force of the ILAE Diagnostic Methods Commission. Epilepsia. 2011, 52, 158–174. [Google Scholar] [CrossRef] [PubMed]
- Cotter, D.; Honavar, M.; Lovestone, S.; Raymond, L.; Kerwin, R.; Anderton, B.; Everall, I. Disturbance of Notch-1 and Wnt signalling proteins in neuroglial balloon cells and abnormal large neurons in focal cortical dysplasia in human cortex. Acta Neuropathol. 1999, 98, 465–472. [Google Scholar] [CrossRef]
- Aizpuru, R.N.; Quencer, R.M.; Norenberg, M.; Altman, N.; Smirniotopoulos, J. Meningioangiomatosis: Clinical, radiologic, and histopathologic correlation. Radiology 1991, 179, 819–821. [Google Scholar] [CrossRef]
- Kim, N.R.; Cho, S.J.; Suh, Y.L. Allelic loss on chromosomes 1p32, 9p21, 13q14, 16q22, 17p, and 22q12 in meningiomas associated with meningioangiomatosis and pure meningioangiomatosis. J. Neurooncol. 2009, 94, 425–430. [Google Scholar] [CrossRef]
- Bonduelle, T.; Hartlieb, T.; Baldassari, S.; Sim, N.S.; Kim, S.H.; Kang, H.C.; Kobow, K.; Coras, R.; Chipaux, M.; Dorfmüller, G.; et al. Frequent SLC35A2 brain mosaicism in mild malformation of cortical development with oligodendroglial hyperplasia in epilepsy (MOGHE). Acta Neuropathol. Commun. 2021, 9, 3. [Google Scholar] [CrossRef] [PubMed]
- Elziny, S.; Crino, P.B.; Winawer, M. SLC35A2 somatic variants in drug resistant epilepsy: FCD and MOGHE. Neurobiol. Dis. 2023, 187, 106299. [Google Scholar] [CrossRef]
- Kobow, K.; Blümcke, I. Epigenetics in epilepsy. Neurosci. Lett. 2018, 667, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Akinsoji, E.O.; Leibovitch, E.; Billioux, B.J.; Abath Neto, O.L.; Ray-Chaudhury, A.; Inati, S.K.; Zaghloul, K.; Heiss, J.; Jacobson, S.; Theodore, W.H. HHV-6 and hippocampal volume in patients with mesial temporal sclerosis. Ann. Clin. Transl. Neurol. 2020, 7, 1674–1680. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Histological Diagnosis | Number of Cases (% of Patients) | M/F | Age at Diagnosis (Mean), Years | Localization (Number of Cases) |
|---|---|---|---|---|
| FCD Type IIa | 26 (66.7) | 13/13 | 2–43 (22) | Frontal—12; temporal—12; parietal—2; occipital—0 |
| FCD Type IIb | 13 (33.3) | 6/7 | 1–16 (6.5) | Frontal—8; temporal—5; parietal—0; occipital—0 |
| Lesion Type | FCD IIa | FCD IIb | TS Fetal | TS Adult | Meningioangiomatosis |
|---|---|---|---|---|---|
| Notch-1+ cell density (cells/mm2) | 0 ± 0 | 42.8 ± 5.3 | 154.2 ± 11.7 | 58.5 ± 7.6 | 11.1 ± 2.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Metodiev, D.; Dimova, P.; Ruseva, M.; Parvanov, D.; Ganeva, R.; Stamenov, G.; Nachev, S.; Ivanova, V.; Marinov, R.; Minkin, K. Notch-1 Immunopositivity in Brain Lesions Associated with Pharmacoresistant Epilepsy. Neuroglia 2025, 6, 7. https://doi.org/10.3390/neuroglia6010007
Metodiev D, Dimova P, Ruseva M, Parvanov D, Ganeva R, Stamenov G, Nachev S, Ivanova V, Marinov R, Minkin K. Notch-1 Immunopositivity in Brain Lesions Associated with Pharmacoresistant Epilepsy. Neuroglia. 2025; 6(1):7. https://doi.org/10.3390/neuroglia6010007
Chicago/Turabian StyleMetodiev, Dimitar, Petia Dimova, Margarita Ruseva, Dimitar Parvanov, Rumiana Ganeva, Georgi Stamenov, Sevdalin Nachev, Vesela Ivanova, Rumen Marinov, and Krassimir Minkin. 2025. "Notch-1 Immunopositivity in Brain Lesions Associated with Pharmacoresistant Epilepsy" Neuroglia 6, no. 1: 7. https://doi.org/10.3390/neuroglia6010007
APA StyleMetodiev, D., Dimova, P., Ruseva, M., Parvanov, D., Ganeva, R., Stamenov, G., Nachev, S., Ivanova, V., Marinov, R., & Minkin, K. (2025). Notch-1 Immunopositivity in Brain Lesions Associated with Pharmacoresistant Epilepsy. Neuroglia, 6(1), 7. https://doi.org/10.3390/neuroglia6010007