
Effects of Bioturbation by Earthworms on Litter Flammability in Young and Mature Afforested Stands



Abstract
1. Introduction
2. Materials and Methods
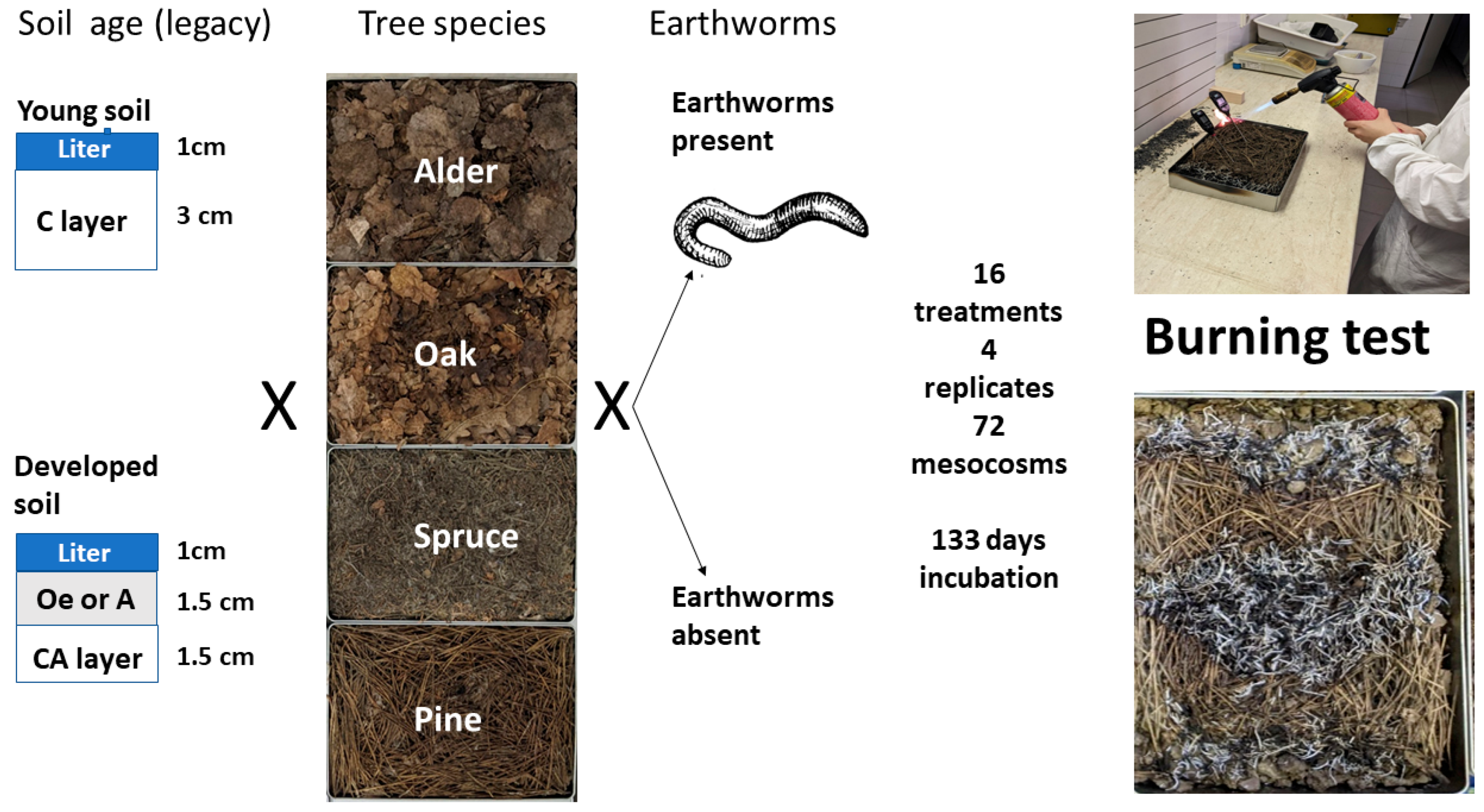
2.1. The Material
2.2. Mesocosm Establishment
2.3. The Burning Test
2.4. The Statistical Analysis
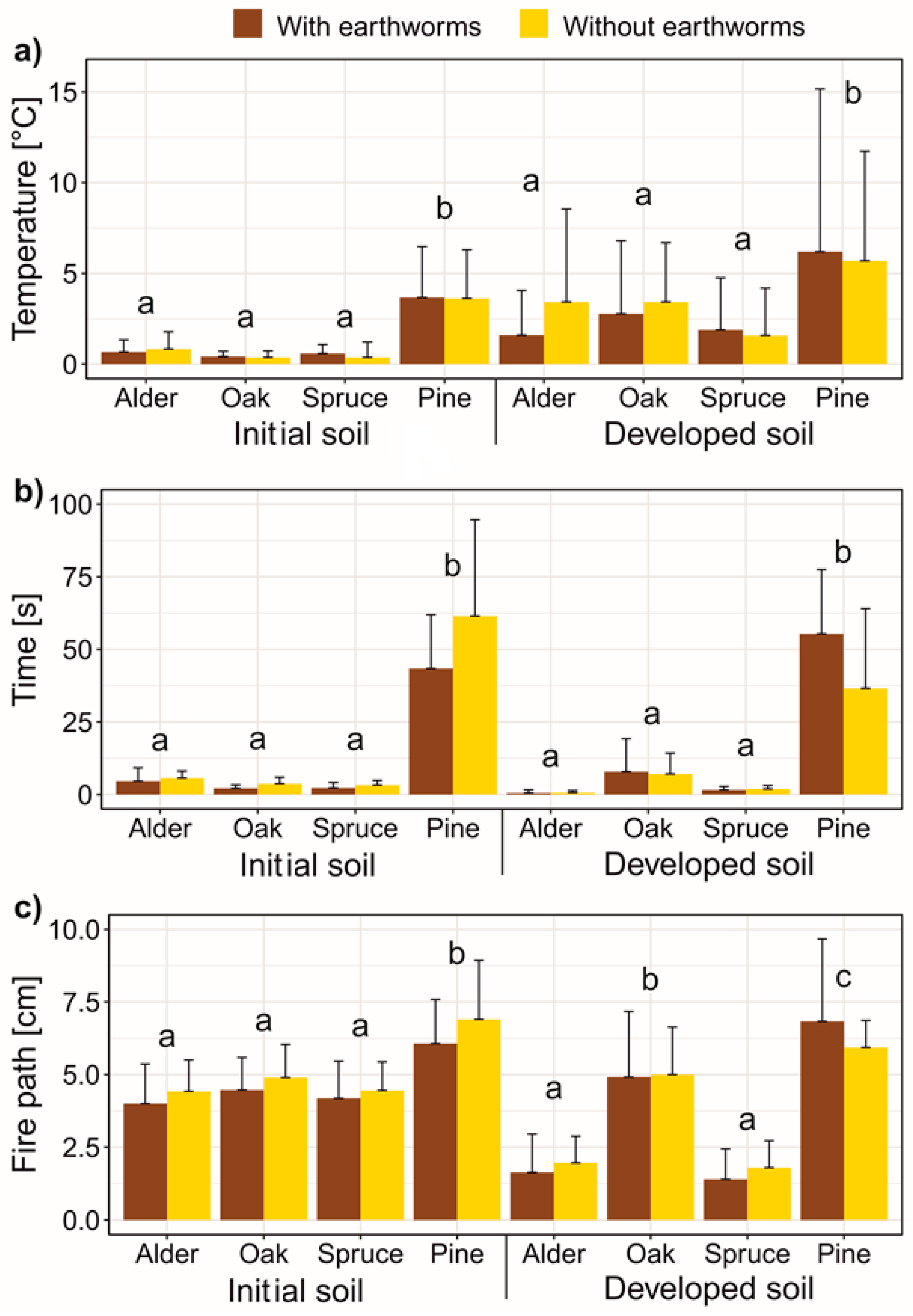
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pausas, J.G.; Keeley, J.E. A Burning Story: The Role of Fire in the History of Life. BioScience 2009, 59, 593–601. [Google Scholar] [CrossRef]
- Halofsky, J.E.; Peterson, D.L.; Harvey, B.J. Changing wildfire, changing forests: The effects of climate change on fire regimes and vegetation in the Pacific Northwest, USA. Fire Ecol. 2020, 16, 4. [Google Scholar] [CrossRef]
- Lasslop, G.; Coppola, A.I.; Voulgarakis, A.; Yue, C.; Veraverbeke, S. Influence of Fire on the Carbon Cycle and Climate. Curr. Clim. Change Rep. 2019, 5, 112–123. [Google Scholar] [CrossRef]
- Noss, R.F. Fire Ecology of Florida and the Southeastern Coastal Plain; University Press of Florida: Gainesville, FL, USA, 2018. [Google Scholar]
- de Magalhães, R.M.; Schwilk, D.W. Leaf traits and litter flammability: Evidence for non-additive mixture effects in a temperate forest. J. Ecol. 2012, 100, 1153–1163. [Google Scholar] [CrossRef]
- Bond, W.J.; Keeley, J.E. Fire as a global ‘herbivore’: The ecology and evolution of flammable ecosystems. Trends Ecol. Evol. 2005, 20, 387–394. [Google Scholar] [CrossRef]
- Murphy, B.P.; Bradstock, R.A.; Boer, M.M.; Carter, J.; Cary, G.J.; Cochrane, M.A.; Fensham, R.J.; Russell-Smith, J.; Williamson, G.J.; Bowman, D.M.J.S. Fire regimes of Australia: A pyrogeographic model system. J. Biogeogr. 2013, 40, 1048–1058. [Google Scholar] [CrossRef]
- Flannigan, M.D.; Stocks, B.J.; Wotton, B.M. Climate change and forest fires. Sci. Tot. Environ. 2000, 262, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Flannigan, M.D.; Krawchuk, M.A.; De Groot, W.J.; Wotton, B.M.; Gowman, L.M. Implications of changing climate for global wildland fire. Int. J. Wildland Fire 2009, 18, 483–507. [Google Scholar] [CrossRef]
- Liu, Y.; Stanturf, J.; Goodrick, S. Trends in global wildfire potential in a changing climate. For. Ecol. Manag. 2010, 259, 685–697. [Google Scholar] [CrossRef]
- Bowman, D.M.; Panton, W.J. Decline of Callitris intratropica R. T. Baker & H. G. Smith in the Northern Territory: Implications for pre- and post-European colonization fire regimes. J. Biogeogr. 1993, 20, 373–381. [Google Scholar] [CrossRef]
- Swetnam, T.W. Fire history and climate change in giant sequoia groves. Science 1993, 262, 885–889. [Google Scholar] [CrossRef] [PubMed]
- Bradstock, R.A. Flammable Australia: The Fire Regimes and Biodiversity of a Continent; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar]
- Curt, T.; Schaffhauser, A.; Borgniet, L.; Dumas, C.; Estève, R.; Ganteaume, A.; Jappiot, M.; Martin, W.; N’Diaye, A.; Poilvet, B. Litter flammability in oak woodlands and shrublands of southeastern France. For. Ecol. Manag. 2011, 261, 2214–2222. [Google Scholar] [CrossRef]
- Ganteaume, A.; Guijarro, M.; Jappiot, M.; Hernando, C.; Lampin-Maillet, C.; Pérez-Gorostiaga, P.; Vega, J.A. Laboratory characterization of firebrands involved in spot fires. Ann. For. Sci. 2011, 68, 531–541. [Google Scholar] [CrossRef]
- Hodgkinson, K.C. Fire regimes in Acacia wooded landscapes: Effects on functional processes and biological diversity. In Flammable Australia: The Fire Regimes and Biodiversity of a Continent; Bradstock, R.A., Williams, J.E., Gill, A.M., Eds.; Cambridge University Press: Cambridge, UK, 2002; pp. 259–277. [Google Scholar]
- Archibald, S.; Lehmann, C.E.; Gómez-Dans, J.L.; Bradstock, R.A. Defining pyromes and global syndromes of fire regimes. Proc. Natl. Acad. Sci. USA 2013, 110, 6442–6447. [Google Scholar] [CrossRef]
- Costes, L.; Laoutid, S.; Brohez, P. Dubois, Bio-based flame retardants: When nature meets fire protection. Mat. Sci. Eng. R 2017, 117, 1–25. [Google Scholar] [CrossRef]
- Owens, M.K.; Lin, C.-D.; Taylor, C.A., Jr.; Whisenant, S.G. Seasonal Patterns of Plant Flammability and Monoterpenoid Content in Juniperus ashei. J. Chem. Ecol. 1998, 24, 2115–2129. [Google Scholar] [CrossRef]
- Ormeño, E.; Céspedes, B.; Sánchez, I.A.; Velasco-García, A.; Moreno, J.M.; Fernandez, C.; Baldy, V. The relationship between terpenes and flammability of leaf litter. For. Ecol. Manag. 2009, 257, 471–482. [Google Scholar] [CrossRef]
- Küçük, Ö.; Aktepe, N. Effect of phenolic compounds on the flammability in forest fires. Int. J. Eng. Sci. Res. Technol. 2017, 6, 320–327. [Google Scholar] [CrossRef]
- Cornwell, W.K.; Elvira, A.; van Kempen, L.; van Logtestijn, R.S.P.; Aptroot, A.; Cornelissen, J.H.C. Flammability across the gymnosperm phylogeny: The importance of litter particle size. New Phytol. 2015, 206, 672–681. [Google Scholar] [CrossRef]
- Cochrane, M.A.; Ryan, K.C. Fire and Fire Ecology: Concepts and Principles. In Tropical Fire Ecology: Climate Change, Land Use, and Ecosystem Dynamics; Cochrane, M.A., Ed.; Springer Praxis Books: Berlin/Heidelberg, Germany, 2009; pp. 24–62. [Google Scholar] [CrossRef]
- Zhao, W.; Blauw, L.G.; van Logtestijn, R.S.P.; Cornwell, W.K.; Cornelissen, J.H.C. Interactions between fine wood decomposition and flammability. Forests 2014, 4, 827–846. [Google Scholar] [CrossRef]
- Cornelissen, J.H.C.; Grootemaat, S.; Verheijen, L.M.; Cornwell, W.K.; van Bodegom, P.M.; van der Wal, R.; Aerts, R. Are litter decomposition and fire linked through plant species traits? New Phytol. 2017, 216, 653–669. [Google Scholar] [CrossRef] [PubMed]
- Kurz, C.; Coûteaux, M.-M.; Thiéry, J. Residence time and decomposition rate of Pinus pinaster needles in a forest floor from direct field measurements under a Mediterranean climate. Soil Biol. Biochem. 2000, 32, 1197–1206. [Google Scholar] [CrossRef]
- Dos Santos, A.R.; Nelson, B.W. Leaf Decomposition and Fine Fuels in Floodplain Forests of the Rio Negro in the Brazilian Amazon. J. Trop. Ecol. 2013, 29, 455–458. [Google Scholar] [CrossRef]
- Hobbie, S.E. Effects of plant-species on nutrient cycling. Trends Ecol. Evol. 1992, 7, 336–339. [Google Scholar] [CrossRef] [PubMed]
- Cornwell, W.K.; Cornelissen, J.H.C.; Amatangelo, K.; Dorrepaal, E.; Eviner, V.T.; Godoy, O.; Hobbie, S.E.; Hoorens, B.; Kurokawa, H.; Pérez-Harguindeguy, N.; et al. Plant species traits are the predominant control on litter decomposition rates within biomes worldwide. Ecol. Lett. 2008, 11, 1065–1071. [Google Scholar] [CrossRef]
- Ponge, J.F. Plant–soil feedbacks mediated by humus forms: A review. Soil Biol. Biochem. 2013, 57, 1048–1060. [Google Scholar] [CrossRef]
- Frouz, J. Effects of soil macro- and mesofauna on litter decomposition and soil organic matter stabilization. Geoderma 2018, 332, 161–172. [Google Scholar] [CrossRef]
- Heděnec, P.; Jiménez, J.J.; Moradi, J.; Domene, X.; Hackenberger, D.; Barot, S.; Frossard, A.; Oktaba, L.; Filser, J.; Kindlmann, P.; et al. Global distribution of soil fauna functional groups and their estimated litter consumption across biomes. Sci. Rep. 2022, 12, 17362. [Google Scholar] [CrossRef]
- Cameron, E.K.; Shaw, C.H.; Bayne, E.M.; Kurz, W.A.; Kull, S.J. Modelling interacting effects of invasive earthworms and wildfire on forest floor carbon storage in the boreal forest. Soil Biol. Biochem. 2015, 88, 189–196. [Google Scholar] [CrossRef]
- Frouz, J.; Roubíčková, A.; Heděnec, P.; Tajovský, K. Do soil fauna really hasten litter decomposition? A meta-analysis of enclosure studies. Eur. J. Soil Biol. 2015, 68, 18–24. [Google Scholar] [CrossRef]
- Weir, J.R.; Limb, R.F. Seasonal Variation in Flammability Characteristics of Quercus marilandica and Quercus stellata Leaf Litter Burned in the Laboratory. Fire Ecol. 2013, 9, 80–88. [Google Scholar] [CrossRef]
- Varner, J.M.; Kane, J.M.; Kreye, J.K.; Engber, E. The Flammability of Forest and Woodland Litter: A Synthesis. Curr. For. Rep. 2015, 1, 91–99. [Google Scholar] [CrossRef]
- Jačka, L.; Walmsley, A.; Kovář, M.; Frouz, J. Effects of different tree species on infiltration and preferential flow in soils developing at a clayey spoil heap. Geoderma 2021, 403, 115372. [Google Scholar] [CrossRef]
- Frouz, J.; Pižl, V.; Cienciala, E.; Kalčík, J. Carbon storage in post-mining forest soil, the role of tree biomass and soil bioturbation. Biogeochemistry 2009, 94, 111–121. [Google Scholar] [CrossRef]
- Frouz, J.; Livečková, M.; Albrechtová, J.; Chroňáková, A.; Cajthaml, T.; Pižl, V.; Háněl, L.; Starý, J.; Baldrian, P.; Lhotáková, Z.; et al. Is the effect of trees on soil properties mediated by soil fauna? A case study from post-mining sites. For. Ecol. Manag. 2013, 309, 87–95. [Google Scholar] [CrossRef]
- Frouz, J.; Elhottová, D.; Pižl, V.; Tajovský, K.; Šourková, M.; Picek, T.; Malý, S. The effect of litter quality and soil faunal composition on organic matter dynamics in post-mining soil: A laboratory study. Appl. Soil. Ecol. 2007, 37, 72–80. [Google Scholar] [CrossRef]
- Frouz, J. Plant-soil feedback across spatiotemporal scales from immediate effects to legacy. Soil Biol. Biochem. 2024, 189, 109289. [Google Scholar] [CrossRef]
- Bufacchi, P.; Santos, J.C.; de Carvalho, J.A.; Krieger Filho, G.C. Estimation of the surface area-to-volume ratios of litter components of the Brazilian rainforest and their impact on litter fire rate of spread and flammability. J. Braz. Soc. Mech. Sci. Eng. 2020, 42, 266. [Google Scholar] [CrossRef]
- Frouz, J.; Pižl, V.; Tajovský, K. The effect of earthworms and other saprophagous macrofauna on soil microstructure in reclaimed and un-reclaimed post-mining sites in Central Europe. Conference Information: 8th International Symposium on Earthworm Ecology (ISEE8), SEP 04-09, 2006, Cracow, Poland. Eur. J. Soil Biol. 2007, 43, S184–S189. [Google Scholar] [CrossRef]
- Bohlen, P.J.; Scheu, S.; Hale, C.M.; McLean, M.A.; Migge, S.; Groffman, P.M.; Parkinson, D. Non-native invasive earthworms as agents of change in northern temperate forests. Front. Ecol. Environ. 2004, 2, 427–435. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Parameter | Alder | Oak | Spruce | Pine |
|---|---|---|---|---|
| Oe depth, mm | 0 | 0 | 10 | 10 |
| A + AC depth, mm | 85 | 55 | 18 | 2 |
| C content, % | 7.1 | 5.9 | 4.8 | 4.6 |
| Earthworm density, ind.m−2 | 288 | 68 | 32 | 50 |
| Parameter | Temperature Difference | Burning Time | Fire Path |
|---|---|---|---|
| Young soil and developed soil together, p values (three-way ANOVA) | |||
| 1 legacy | 0.0002 | 0.3303 | <0.0001 |
| 2 tree species | <0.0001 | <0.0001 | <0.0001 |
| 3 earthworms | 0.8099 | 0.8620 | 0.3148 |
| 1 × 2 | 0.7939 | 0.1940 | <0.0001 |
| 1 × 3 | 0.9343 | 0.0086 | 0.2717 |
| 2 × 3 | 0.7757 | 0.9975 | 0.9243 |
| 1 × 2 × 3 | 0.8323 | 0.0014 | 0.4836 |
| Young soil (no legacy), p values (two-way ANOVA) | |||
| 1 tree species | <0.0001 | <0.0001 | <0.0001 |
| 2 earthworms | 0.7821 | 0.0370 | 0.0585 |
| 1 × 2 | 0.9735 | 0.0504 | 0.8792 |
| Developed soil (legacy), p values (two-way ANOVA) | |||
| 1 tree species | <0.0001 | <0.0001 | <0.0001 |
| 2 earthworms | 0.8796 | 0.0981 | 0.9574 |
| 1 × 2 | 0.8358 | 0.0516 | 0.6552 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martinovská, A.; Mudrák, O.; Frouz, J. Effects of Bioturbation by Earthworms on Litter Flammability in Young and Mature Afforested Stands. Fire 2025, 8, 225. https://doi.org/10.3390/fire8060225
Martinovská A, Mudrák O, Frouz J. Effects of Bioturbation by Earthworms on Litter Flammability in Young and Mature Afforested Stands. Fire. 2025; 8(6):225. https://doi.org/10.3390/fire8060225
Chicago/Turabian StyleMartinovská, Aneta, Ondřej Mudrák, and Jan Frouz. 2025. "Effects of Bioturbation by Earthworms on Litter Flammability in Young and Mature Afforested Stands" Fire 8, no. 6: 225. https://doi.org/10.3390/fire8060225
APA StyleMartinovská, A., Mudrák, O., & Frouz, J. (2025). Effects of Bioturbation by Earthworms on Litter Flammability in Young and Mature Afforested Stands. Fire, 8(6), 225. https://doi.org/10.3390/fire8060225