
The House Is Burning: Assessment of Habitat Loss Due to Wildfires in Central Mexico

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
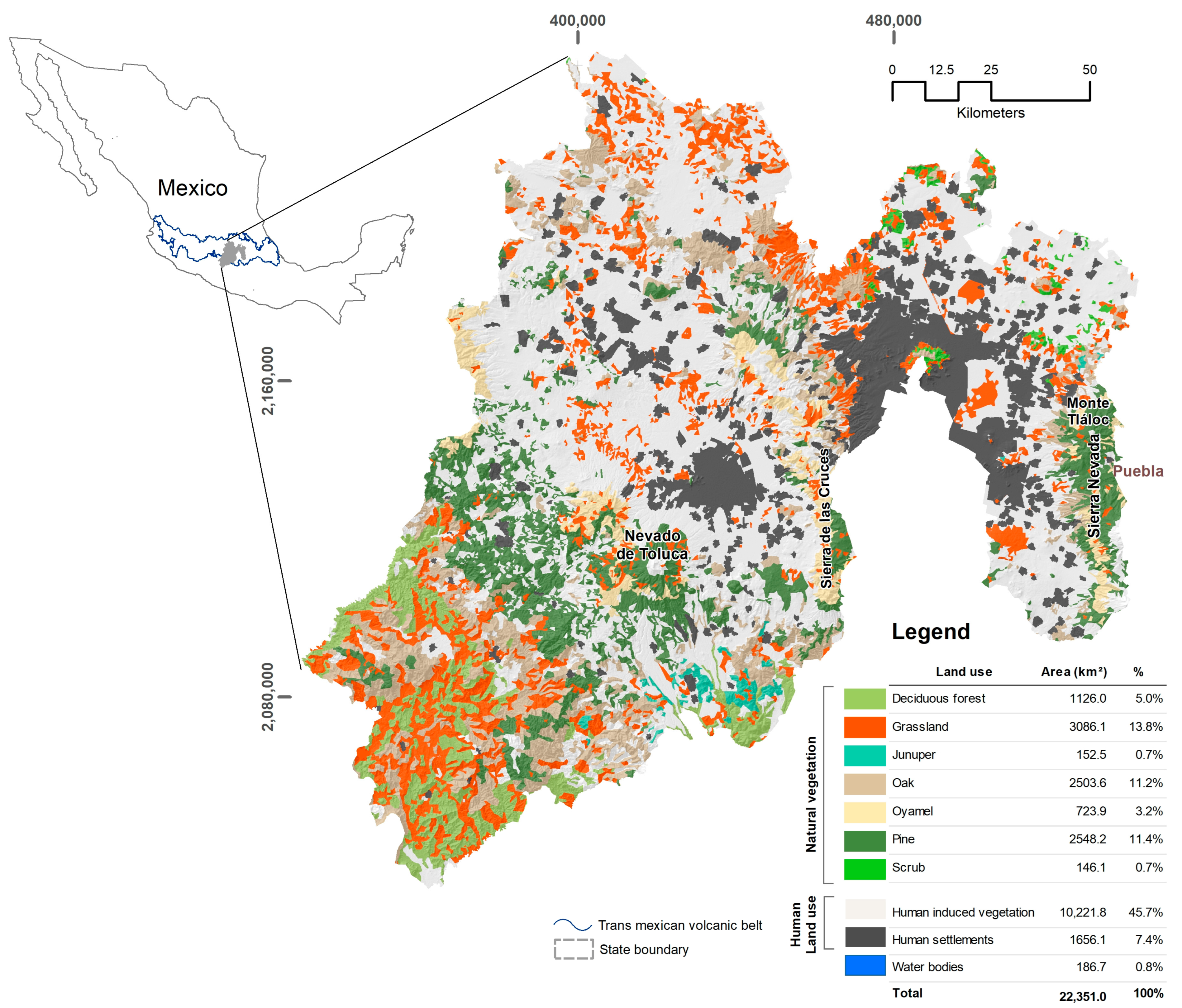
2.1. Study Area
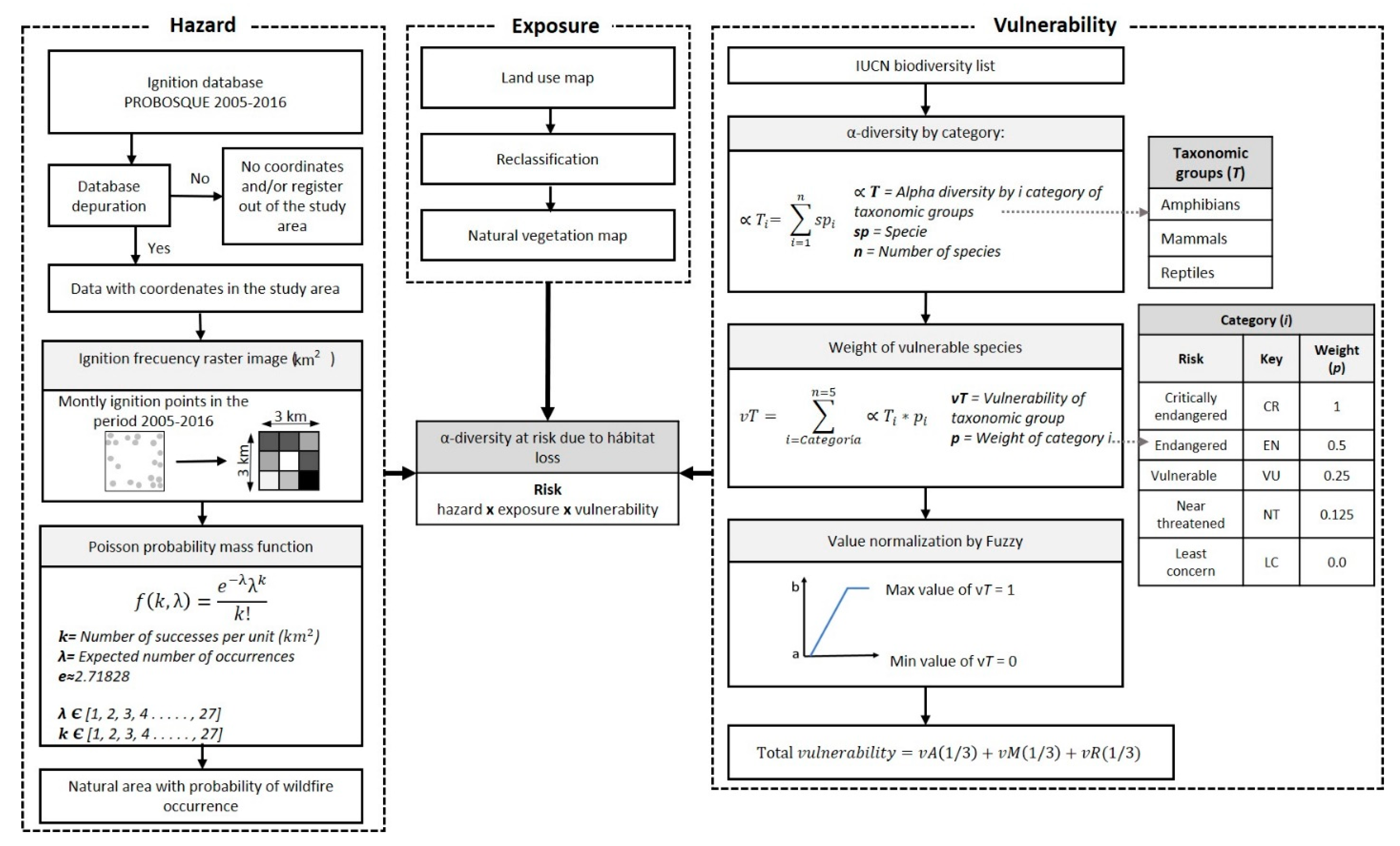
2.2. Methodology to Determine Biodiversity at Risk
2.2.1. Hazard
2.2.2. Exposure
2.2.3. Vulnerability
2.2.4. α-Diversity at Risk due to Wildfires Habitat Loss
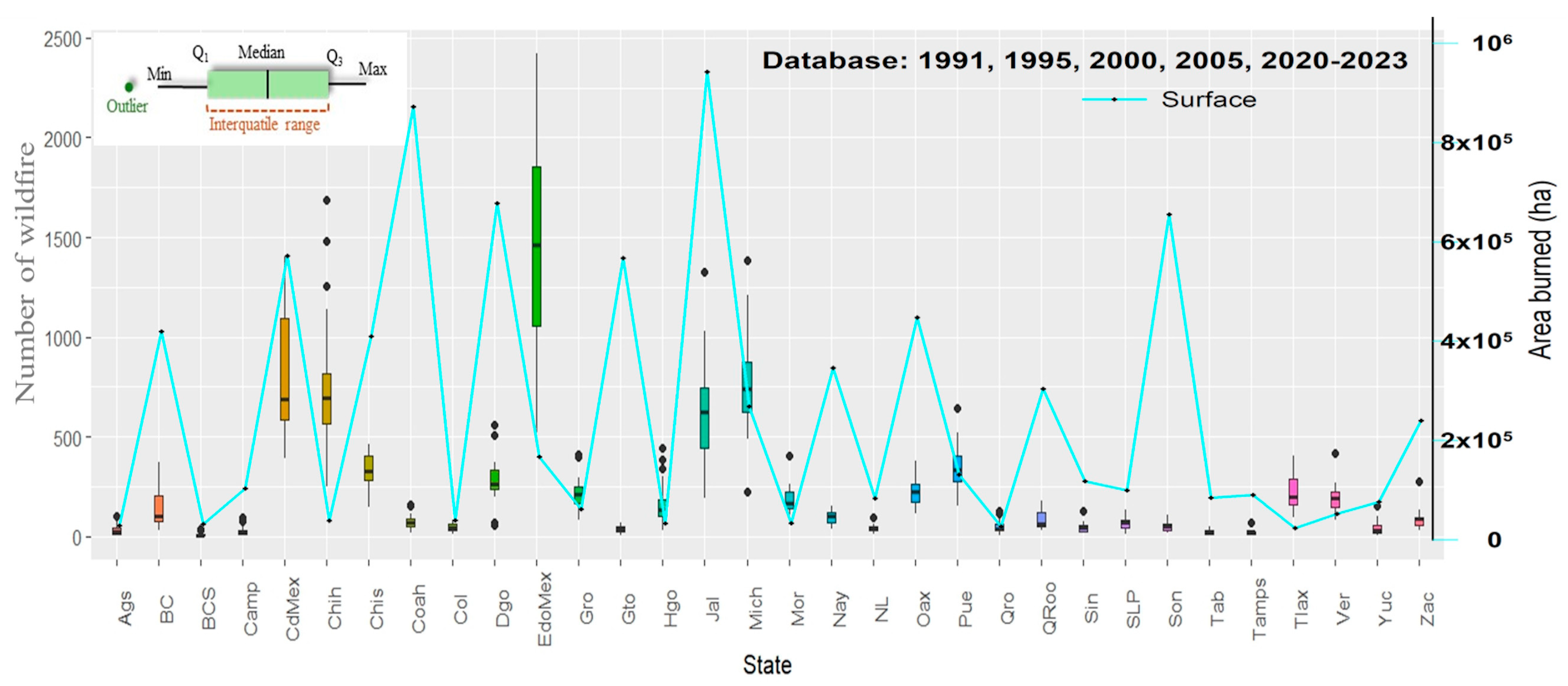
3. Results
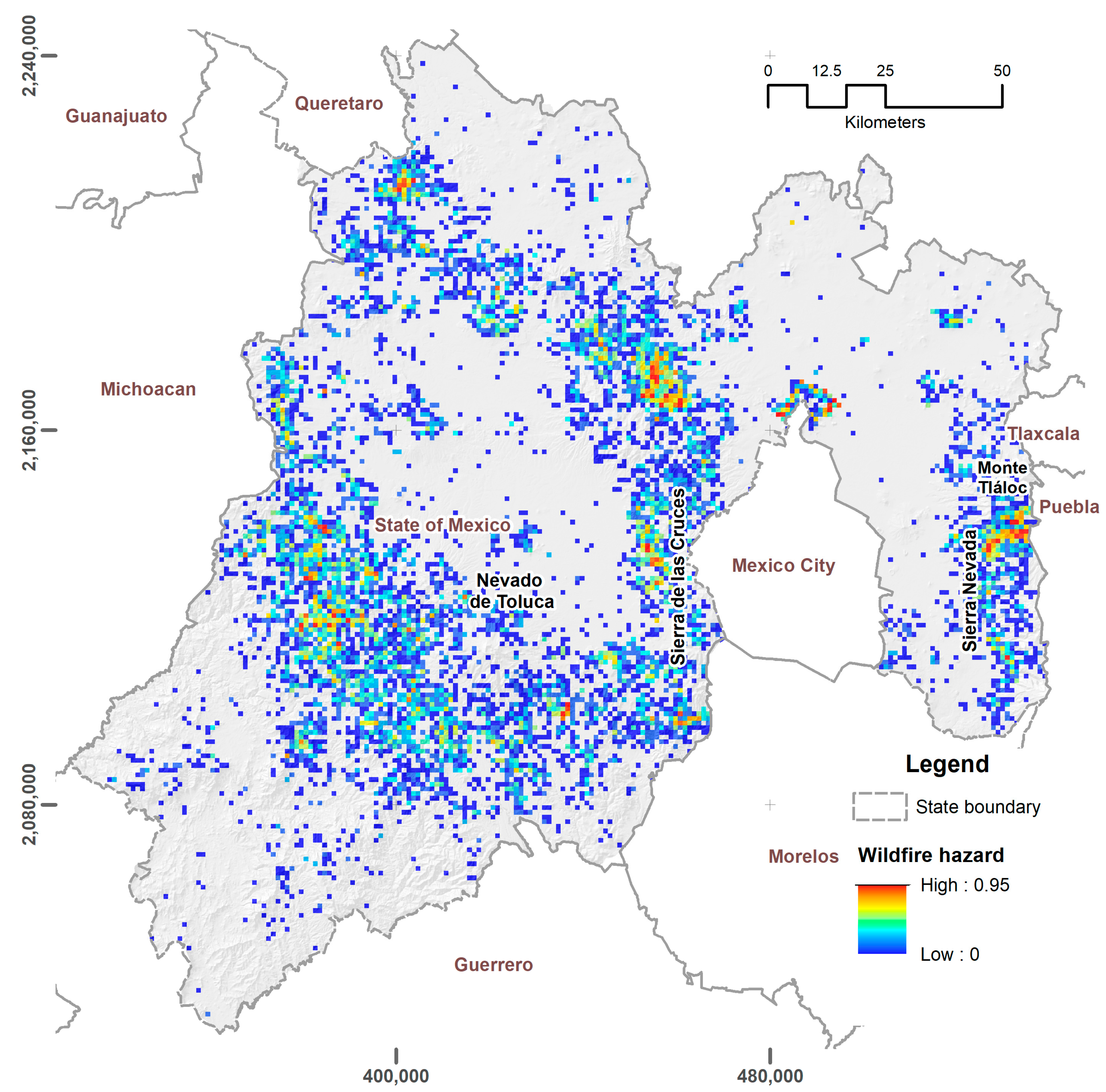
3.1. Exposure Zones and Hazard Analysis
3.2. Vulnerability of Amphibians, Mammals and Reptiles
3.3. α-Diversity at Risk due to Wildfires Habitat Degradation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Harrison, S.P.; Marlon, J.R.; Bartlein, P.J. Fire in the Earth System; Springer: Berlin/Heidelberg, Germany, 2010; ISBN 90-481-8715-X. [Google Scholar]
- Haque, M.K.; Azad, M.A.K.; Hossain, M.Y.; Ahmed, T.; Uddin, M.; Hossain, M.M. Wildfire in Australia during 2019–2020, Its Impact on Health, Biodiversity and Environment with Some Proposals for Risk Management: A Review. J. Environ. Prot. 2021, 12, 391–414. [Google Scholar] [CrossRef]
- Bowman, D.M.; Williamson, G.J.; Abatzoglou, J.T.; Kolden, C.A.; Cochrane, M.A.; Smith, A.M. Human Exposure and Sensitivity to Globally Extreme Wildfire Events. Nat. Ecol. Evol. 2017, 1, 0058. [Google Scholar] [CrossRef]
- Moretti, M.; Obrist, M.K.; Duelli, P. Arthropod Biodiversity after Forest Fires: Winners and Losers in the Winter Fire Regime of the Southern Alps. Ecography 2004, 27, 173–186. [Google Scholar] [CrossRef]
- Foster, C.; Barton, P.; Robinson, N.; MacGregor, C.; Lindenmayer, D.B. Effects of a Large Wildfire on Vegetation Structure in a Variable Fire Mosaic. Ecol. Appl. 2017, 27, 2369–2381. [Google Scholar] [CrossRef] [PubMed]
- He, T.; Belcher, C.M.; Lamont, B.B.; Lim, S.L. A 350-million-year Legacy of Fire Adaptation among Conifers. J. Ecol. 2016, 104, 352–363. [Google Scholar] [CrossRef]
- Brooks, M.L.; D’antonio, C.M.; Richardson, D.M.; Grace, J.B.; Keeley, J.E.; DiTomaso, J.M.; Hobbs, R.J.; Pellant, M.; Pyke, D. Effects of Invasive Alien Plants on Fire Regimes. BioScience 2004, 54, 677–688. [Google Scholar] [CrossRef]
- Di Virgilio, G.; Evans, J.P.; Blake, S.A.; Armstrong, M.; Dowdy, A.J.; Sharples, J.; McRae, R. Climate Change Increases the Potential for Extreme Wildfires. Geophys. Res. Lett. 2019, 46, 8517–8526. [Google Scholar] [CrossRef]
- McWethy, D.B.; Schoennagel, T.; Higuera, P.E.; Krawchuk, M.; Harvey, B.J.; Metcalf, E.C.; Schultz, C.; Miller, C.; Metcalf, A.L.; Buma, B. Rethinking Resilience to Wildfire. Nat. Sustain. 2019, 2, 797–804. [Google Scholar] [CrossRef]
- Littell, J.S.; Peterson, D.L.; Riley, K.L.; Liu, Y.; Luce, C.H. A Review of the Relationships between Drought and Forest Fire in the United States. Glob. Chang. Biol. 2016, 22, 2353–2369. [Google Scholar] [CrossRef]
- Ruffault, J.; Curt, T.; St-Paul, N.M.; Moron, V.; Trigo, R.M. Extreme Wildfire Occurrence in Response to Global Change Type Droughts in the Northern Mediterranean. Nat. Hazards Earth Syst. Sci. 2017, 1–21. [Google Scholar] [CrossRef]
- Pausas, J.G.; Keeley, J.E. Evolutionary Ecology of Resprouting and Seeding in Fire-prone Ecosystems. New Phytol. 2014, 204, 55–65. [Google Scholar] [CrossRef]
- Le Breton, T.D.; Lyons, M.B.; Nolan, R.H.; Penman, T.; Williamson, G.J.; Ooi, M.K. Megafire-induced Interval Squeeze Threatens Vegetation at Landscape Scales. Front. Ecol. Environ. 2022, 20, 327–334. [Google Scholar] [CrossRef]
- EM-DAT; CRED; UCLouvain, Brussels EM-DAT CRED. Available online: https://www.emdat.be (accessed on 20 January 2024).
- Linley, G.D.; Jolly, C.J.; Doherty, T.S.; Geary, W.L.; Armenteras, D.; Belcher, C.M.; Bliege Bird, R.; Duane, A.; Fletcher, M.; Giorgis, M.A. What Do You Mean,‘Megafire’? Glob. Ecol. Biogeogr. 2022, 31, 1906–1922. [Google Scholar] [CrossRef]
- Singleton, M.P.; Thode, A.E.; Meador, A.J.S.; Iniguez, J.M. Increasing Trends in High-Severity Fire in the Southwestern USA from 1984 to 2015. For. Ecol. Manag. 2019, 433, 709–719. [Google Scholar] [CrossRef]
- Weber, K.T.; Yadav, R. Spatiotemporal Trends in Wildfires across the Western United States (1950–2019). Remote Sens. 2020, 12, 2959. [Google Scholar] [CrossRef]
- Collins, L.; Bradstock, R.A.; Clarke, H.; Clarke, M.F.; Nolan, R.H.; Penman, T.D. The 2019/2020 Mega-Fires Exposed Australian Ecosystems to an Unprecedented Extent of High-Severity Fire. Environ. Res. Lett. 2021, 16, 044029. [Google Scholar] [CrossRef]
- Castellnou, M.; Guiomar, N.; Rego, F.; Fernandes, P.M. Fire Growth Patterns in the 2017 Mega Fire Episode of October 15, Central Portugal. Adv. For. Fire Res. 2018, 447–453. [Google Scholar]
- Pliscoff, P.; Folchi, M.; Aliste, E.; Cea, D.; Simonetti, J.A. Chile Mega-Fire 2017: An Analysis of Social Representation of Forest Plantation Territory. Appl. Geogr. 2020, 119, 102226. [Google Scholar] [CrossRef]
- Troumbis, A.Y.; Kalabokidis, K.; Palaiologou, P. Diverging Rationalities between Forest Fire Management Services and the General Public after the 21st-Century Mega-Fires in Greece. J. For. Res. 2022, 33, 553–564. [Google Scholar] [CrossRef]
- Fidelis, A.; Alvarado, S.T.; Barradas, A.C.S.; Pivello, V.R. The Year 2017: Megafires and Management in the Cerrado. Fire 2018, 1, 49. [Google Scholar] [CrossRef]
- Pereira, M.G.; Fernandes, L.S.; Carvalho, S.; Santos, R.B.; Caramelo, L.; Alencoao, A. Modelling the Impacts of Wildfires on Runoff at the River Basin Ecological Scale in a Changing Mediterranean Environment. Environ. Earth Sci. 2016, 75, 392. [Google Scholar] [CrossRef]
- Pastor, A.V.; Nunes, J.P.; Ciampalini, R.; Koopmans, M.; Baartman, J.; Huard, F.; Calheiros, T.; Le-Bissonnais, Y.; Keizer, J.J.; Raclot, D. Projecting Future Impacts of Global Change Including Fires on Soil Erosion to Anticipate Better Land Management in the Forests of NW Portugal. Water 2019, 11, 2617. [Google Scholar] [CrossRef]
- Kastridis, A.; Margiorou, S.; Sapountzis, M. Check-Dams and Silt Fences: Cost-Effective Methods to Monitor Soil Erosion under Various Disturbances in Forest Ecosystems. Land 2022, 11, 2129. [Google Scholar] [CrossRef]
- Pingree, M.R.; Kobziar, L.N. The Myth of the Biological Threshold: A Review of Biological Responses to Soil Heating Associated with Wildland Fire. For. Ecol. Manag. 2019, 432, 1022–1029. [Google Scholar] [CrossRef]
- Neary, D.G.; Ryan, K.C.; DeBano, L.F. Wildland Fire in Ecosystems: Effects of Fire on Soils and Water; Gen. Tech. Rep. RMRS-GTR-42-vol. 4; US Department of Agriculture, Forest Service, Rocky Mountain Research Station: Ogden, UT, USA, 2005; Volume 42, 250p. [Google Scholar]
- Bond, W.J.; Woodward, F.I.; Midgley, G.F. The Global Distribution of Ecosystems in a World without Fire. New Phytol. 2005, 165, 525–538. [Google Scholar] [CrossRef] [PubMed]
- Silveira, J.M.; Louzada, J.; Barlow, J.; Andrade, R.; Mestre, L.; Solar, R.; Lacau, S.; Cochrane, M.A. A Multi-taxa Assessment of Biodiversity Change after Single and Recurrent Wildfires in a Brazilian Amazon Forest. Biotropica 2016, 48, 170–180. [Google Scholar] [CrossRef]
- Adams, M.A. Mega-Fires, Tipping Points and Ecosystem Services: Managing Forests and Woodlands in an Uncertain Future. For. Ecol. Manag. 2013, 294, 250–261. [Google Scholar] [CrossRef]
- Caon, L.; Vallejo, V.R.; Ritsema, C.J.; Geissen, V. Effects of Wildfire on Soil Nutrients in Mediterranean Ecosystems. Earth-Sci. Rev. 2014, 139, 47–58. [Google Scholar] [CrossRef]
- Slingsby, J.A.; Moncrieff, G.R.; Rogers, A.J.; February, E.C. Altered Ignition Catchments Threaten a Hyperdiverse Fire-dependent Ecosystem. Glob. Chang. Biol. 2020, 26, 616–628. [Google Scholar] [CrossRef]
- DeBano, L.F.; Neary, D.G.; Ffolliott, P.F. Fire Effects on Ecosystems; John Wiley & Sons: Hoboken, NJ, USA, 1998. [Google Scholar]
- Southwell, D.; Legge, S.; Woinarski, J.; Lindenmayer, D.; Lavery, T.; Wintle, B. Design Considerations for Rapid Biodiversity Reconnaissance Surveys and Long-term Monitoring to Assess the Impact of Wildfire. Divers. Distrib. 2022, 28, 559–570. [Google Scholar] [CrossRef]
- Gade, M.R.; Gould, P.R.; Peterman, W.E. Habitat-Dependent Responses of Terrestrial Salamanders to Wildfire in the Short-Term. For. Ecol. Manag. 2019, 449, 117479. [Google Scholar] [CrossRef]
- Bradstock, R.A. Effects of Large Fires on Biodiversity in South-Eastern Australia: Disaster or Template for Diversity? Int. J. Wildland Fire 2008, 17, 809. [Google Scholar] [CrossRef]
- Legge, S.; Rumpff, L.; Woinarski, J.C.; Whiterod, N.S.; Ward, M.; Southwell, D.G.; Scheele, B.C.; Nimmo, D.G.; Lintermans, M.; Geyle, H.M. The Conservation Impacts of Ecological Disturbance: Time-bound Estimates of Population Loss and Recovery for Fauna Affected by the 2019–2020 Australian Megafires. Glob. Ecol. Biogeogr. 2022, 31, 2085–2104. [Google Scholar] [CrossRef]
- Jolly, C.J.; Dickman, C.R.; Doherty, T.S.; van Eeden, L.M.; Geary, W.L.; Legge, S.M.; Woinarski, J.C.; Nimmo, D.G. Animal Mortality during Fire. Glob. Chang. Biol. 2022, 28, 2053–2065. [Google Scholar] [CrossRef] [PubMed]
- Kyle, S.C.; Block, W.M. Effects of Wildfire Severity on Small Mammals in Northern Arizona Ponderosa Pine Forests. In Fire and Forest Ecology: Innovative Silviculture and Vegetation Management. Tall Timbers Fire EcologyConference Proceedings, No. 21; Tall Timbers Research Station: Tallahassee, FL, USA, 2000; Volume 21, pp. 163–168. [Google Scholar]
- Pastro, L.A.; Dickman, C.R.; Letnic, M. Burning for Biodiversity or Burning Biodiversity? Prescribed Burn vs. Wildfire Impacts on Plants, Lizards, and Mammals. Ecol. Appl. 2011, 21, 3238–3253. [Google Scholar] [CrossRef]
- Driscoll, D.A.; Armenteras, D.; Bennett, A.F.; Brotons, L.; Clarke, M.F.; Doherty, T.S.; Haslem, A.; Kelly, L.T.; Sato, C.F.; Sitters, H. How Fire Interacts with Habitat Loss and Fragmentation. Biol. Rev. 2021, 96, 976–998. [Google Scholar] [CrossRef] [PubMed]
- Cunillera-Montcusí, D.; Gascón, S.; Tornero, I.; Sala, J.; Àvila, N.; Quintana, X.D.; Boix, D. Direct and Indirect Impacts of Wildfire on Faunal Communities of Mediterranean Temporary Ponds. Freshw. Biol. 2019, 64, 323–334. [Google Scholar] [CrossRef]
- Ayars, J.; Kramer, H.A.; Jones, G.M. The 2020 to 2021 California Megafires and Their Impacts on Wildlife Habitat. Proc. Natl. Acad. Sci. USA 2023, 120, e2312909120. [Google Scholar] [CrossRef] [PubMed]
- González, T.M.; González-Trujillo, J.D.; Muñoz, A.; Armenteras, D. Effects of Fire History on Animal Communities: A Systematic Review. Ecol. Process. 2022, 11, 11. [Google Scholar] [CrossRef]
- Hossack, B.R.; Pilliod, D.S. Amphibian Responses to Wildfire in the Western United States: Emerging Patterns from Short-Term Studies. Fire Ecol. 2011, 7, 129–144. [Google Scholar] [CrossRef]
- Chia, E.K.; Bassett, M.; Nimmo, D.G.; Leonard, S.W.; Ritchie, E.G.; Clarke, M.F.; Bennett, A.F. Fire Severity and Fire-Induced Landscape Heterogeneity Affect Arboreal Mammals in Fire-Prone Forests. Ecosphere 2015, 6, 1–14. [Google Scholar] [CrossRef]
- Roberts, S.L.; Kelt, D.A.; Van Wagtendonk, J.W.; Miles, A.K.; Meyer, M.D. Effects of Fire on Small Mammal Communities in Frequent-Fire Forests in California. J. Mammal. 2015, 96, 107–119. [Google Scholar] [CrossRef]
- Bagne, K.E.; Finch, D.M. Response of Small Mammal Populations to Fuel Treatment and Precipitation in a Ponderosa Pine Forest, New Mexico. Restor. Ecol. 2010, 18, 409–417. [Google Scholar] [CrossRef]
- Converse, S.J.; White, G.C.; Block, W.M. Small Mammal Responses to Thinning and Wildfire in Ponderosa Pine–Dominated Forests of the Southwestern United States. J. Wildl. Manag. 2006, 70, 1711–1722. [Google Scholar] [CrossRef]
- Otto, C.R.V.; Kroll, A.J.; McKenny, H.C. Amphibian Response to Downed Wood Retention in Managed Forests: A Prospectus for Future Biomass Harvest in North America. For. Ecol. Manag. 2013, 304, 275–285. [Google Scholar] [CrossRef]
- Todd, B.D.; Luhring, T.M.; Rothermel, B.B.; Gibbons, J.W. Effects of Forest Removal on Amphibian Migrations: Implications for Habitat and Landscape Connectivity. J. Appl. Ecol. 2009, 46, 554–561. [Google Scholar] [CrossRef]
- Beranek, C.T.; Hamer, A.J.; Mahony, S.V.; Stauber, A.; Ryan, S.A.; Gould, J.; Wallace, S.; Stock, S.; Kelly, O.; Parkin, T. Severe Wildfires Promoted by Climate Change Negatively Impact Forest Amphibian Metacommunities. Divers. Distrib. 2023, 29, 785–800. [Google Scholar] [CrossRef]
- Hossack, B.R.; Eby, L.A.; Guscio, C.G.; Corn, P.S. Thermal Characteristics of Amphibian Microhabitats in a Fire-Disturbed Landscape. For. Ecol. Manag. 2009, 258, 1414–1421. [Google Scholar] [CrossRef]
- Santos, X.; Belliure, J.; Gonçalves, J.F.; Pausas, J.G. Resilience of Reptiles to Megafires. Ecol. Appl. 2022, 32, e2518. [Google Scholar] [CrossRef]
- Wilgers, D.J.; Horne, E.A. Effects of Different Burn Regimes on Tallgrass Prairie Herpetofaunal Species Diversity and Community Composition in the Flint Hills, Kansas. J. Herpetol. 2006, 40, 73–84. [Google Scholar] [CrossRef]
- Barrile, G.M.; Chalfoun, A.D.; Estes-Zumpf, W.A.; Walters, A.W. Wildfire Influences Individual Growth and Breeding Dispersal, but Not Survival and Recruitment in a Montane Amphibian. Ecosphere 2022, 13, e4212. [Google Scholar] [CrossRef]
- INEGI. Censo Población y Vivienda 2020; INEGI: Aguascalientes, Mexico, 2020.
- INEGI. Producto Interno Bruto por Entidad Federativa (PIBE); INEGI: Aguascalientes, Mexico, 2023; p. 12.
- Godinez-Tovar, A.G.; Lopez-Gutierrez, M.; Becerril-Piña, R.; Mastachi-Loza, C.A. Influencia Del Cambio Del Uso de Suelo Sobre La Dinámica de La Precipitación. Caso de Estudio: Curso Alto de La Cuenca Alta Del Río Lerma, México. In Geología Ambiental y Recursos Hídricos; Cromberger Editores e Impresores, S.A. de C.V.: Mexico City, Mexico, 2023; Volume 37, pp. 349–364. ISBN 978-607-589-210-8. [Google Scholar]
- Monroy-Vilchis, O.; Luna-Gil, A.A.; Endara-Agramont, A.R.; Zarco-González, M.M.; González-Desales, G.A. Nevado de Toluca: Habitat for Romerolagus Diazi? Anim. Biodivers. Conserv. 2020, 43, 115–121. [Google Scholar] [CrossRef]
- Aguilar, X.; Casas, G. Secretaría del Medio Ambiente Anfibios y Reptiles. In Biodiversidad del Estado de México: Estudio de Estado; Secretaría del Medio Ambiente, Ed.; Gobierno del Estado de México y Comisión para el Conocimiento y Uso de la Biodiversidad: Toluca, Mexico, 2009; pp. 125–130. [Google Scholar]
- INEGI. Conjunto de Datos Vectoriales de Uso de Suelo y Vegetación Escala 1:250 000, Serie V; NEGI: Aguascalientes, Mexico, 2013.
- Gonzalez-Fernandez, A.; Segarra, J.; Sunny, A.; Couturier, S. Forest Cover Loss in the Nevado de Toluca Volcano Protected Area (Mexico) after the Change to a Less Restrictive Category in 2013. Biodivers. Conserv. 2022, 31, 871–894. [Google Scholar] [CrossRef]
- SEMARNAT. Sistema Nacional de Información Ambiental y de Recursos Naturales; SNIARN: Mexico City, Mexico, 2024. [Google Scholar]
- Crichton, D. The Risk Triangle in Natural Disaster Management; Tudor Rose: London, UK, 1999. [Google Scholar]
- Crichton, D. UK and Global Insurance Responses to Flood Hazard. Water Int. 2002, 27, 119–131. [Google Scholar] [CrossRef]
- Katti, S.K.; Rao, A.V. Handbook of the Poisson Distribution; Taylor & Francis Group: Abingdon, UK, 1968. [Google Scholar]
- CONABIO. Sistema Nacional de Información Sobre Biodiversidad (SNIB); Registros de Ejemplares; CONABIO: Mexico City, Mexico, 2024. [Google Scholar]
- Lange, H.J.D.; Lahr, J.; Van der Pol, J.J.; Wessels, Y.; Faber, J.H. Ecological Vulnerability in Wildlife: An Expert Judgment and Multicriteria Analysis Tool Using Ecological Traits to Assess Relative Impact of Pollutants. Environ. Toxicol. Chem. Int. J. 2009, 28, 2233–2240. [Google Scholar] [CrossRef]
- Leverington, F.; Costa, K.L.; Pavese, H.; Lisle, A.; Hockings, M. A Global Analysis of Protected Area Management Effectiveness. Environ. Manag. 2010, 46, 685–698. [Google Scholar] [CrossRef] [PubMed]
- IUCN. IUCN Red List Categories and Criteria, Version 3.1, 2nd ed.; IUCN: Gland, Switzerland, 2012; ISBN 978-2-8317-1435-6. [Google Scholar]
- CONABIO; CONANP; PNUD. Corredores Bioclimáticos Para La Conservación de La Biodiversidad. 2019. Mexico City, Mexico. Available online: http://www.conabio.gob.mx/informacion/gis/?vns=gis_root/region/prioridad/clccrecgw (accessed on 3 April 2024).
- Jung, M. LecoS—A Python Plugin for Automated Landscape Ecology Analysis. Ecol. Inform. 2016, 31, 18–21. [Google Scholar] [CrossRef]
- QGIS Development Team QGIS Geographic Information System. Open source Geospatial Foundation Project. Available online: http://qgis.org (accessed on 25 March 2020).
- Mastachi-Loza, C.A.; Becerril-Piña, R.; Gómez-Albores, M.A.; Díaz-Delgado, C.; Romero-Contreras, A.T.; Garcia-Aragon, J.A.; Vizcarra-Bordi, I. Regional Analysis of Climate Variability at Three Time Scales and Its Effect on Rainfed Maize Production in the Upper Lerma River Basin, Mexico. Agric. Ecosyst. Environ. 2016, 225, 1–11. [Google Scholar] [CrossRef]
- Luna-Vega, I.; Alcántara-Ayala, O.; García-Morales, L.J.; Espinosa, D.; Ramírez-Martínez, J.C.; Contreras-Medina, R. Threatened Trees Characteristic of Mexican Tropical Montane Cloud Forests. Diversity 2022, 15, 42. [Google Scholar] [CrossRef]
- Strachinis, I. The Herpetofauna of the Peri-Urban Forest Seich Sou (Kedrinos Lofos), Thessaloniki, Greece. Ecol. Balk. 2023, 15, 1–7. [Google Scholar]
- Glickman, D.; Babbitt, B. Urban Wildland Interface Communities within the Vicinity of Federal Lands That Are at High Risk from Wildfire. Fed. Regist. 2001, 66, 751–777. [Google Scholar]
- Cobos, E.P. Zona Metropolitana Del Valle de México: Neoliberalismo y Contradicciones Urbanas. Sociologias 2016, 18, 54–89. [Google Scholar] [CrossRef]
- CITES. Estado de Conservación, Uso, Gestión y Comercio de Las Especies Del Género Abronia Que Se Distribuyen En México; Vigésimo Séptima Reunión Del Comité de Fauna Veracruz: Veracruz, Mexico, 2014; Volume 23. [Google Scholar]
- Cruz-Sáenz, D.; Vázquez, S.G.; Lazcano, D. Notes on the Herpetofauna of Western Mexico 13: Effects of Wildfires on the Reptile Community in the Natural Protected Area “La Primavera,” in Jalisco, Mexico. Bull. Chic. Herp. Soc. 2015, 50, 96–100. [Google Scholar]
- Moreira, F.; Russo, D. Modelling the Impact of Agricultural Abandonment and Wildfires on Vertebrate Diversity in Mediterranean Europe. Landsc. Ecol. 2007, 22, 1461–1476. [Google Scholar] [CrossRef]
- Pianka, E.R.; Goodyear, S.E. Lizard Responses to Wildfire in Arid Interior Australia: Long-Term Experimental Data and Commonalities with Other Studies. Austral Ecol. 2012, 37, 1–11. [Google Scholar] [CrossRef]
- SEMARNAT. Programa de Acción Para La Conservación de Las Especies Abronia (Abronia spp) En México; SNIARN: Mexico City, Mexico, 2018. [Google Scholar]
- Setser, K.; Cavitt, J.F. Effects of Burning on Snakes in Kansas, USA, Tallgrass Prairie. Nat. Areas J. 2003, 23, 315–319. [Google Scholar]
- Woinarski, J.C.; Armstrong, M.; Price, O.; McCartney, J.; Griffiths, A.D.; Fisher, A. The Terrestrial Vertebrate Fauna of Litchfield National Park, Northern Territory: Monitoring over a 6-Year Period and Response to Fire History. Wildl. Res. 2005, 31, 587–596. [Google Scholar] [CrossRef]
- Hossack, B.R.; Lowe, W.H.; Corn, P.S. Rapid Increases and Time-Lagged Declines in Amphibian Occupancy after Wildfire. Conserv. Biol. 2013, 27, 219–228. [Google Scholar] [CrossRef]
- García, S.; Monroy-Vilchis, O.; Fajardo, V.; Aguilera-Reyes, U. Genetic Diversity and Structure of an Endemic and Critically Endangered Stream River Salamander (Caudata: Ambystoma Leorae) in Mexico. Conserv. Genet. 2014, 15, 49–59. [Google Scholar]
- Hossack, B.R.; Lowe, W.H.; Honeycutt, R.K.; Parks, S.A.; Corn, P.S. Interactive Effects of Wildfire, Forest Management, and Isolation on Amphibian and Parasite Abundance. Ecol. Appl. 2013, 23, 479–492. [Google Scholar] [CrossRef]
- Monroy Vilchis, O.; Zarco Gonzalez, M.; Dominguez Vega, H.; Garcia Aguilar, A.S. Ambystoma Leorae (Taylor, 1943). New Records, Natural History Notes and Threat Status. Short Note Herpetozoa 2015, 30, 166–168. [Google Scholar]
- Romo-Vázquez, E.; León-Paniagua, L.; Sánchez, O. A New Species of Habromys (Rodentia: Neotominae) from México. Proc. Biol. Soc. Wash. 2005, 118, 605–618. [Google Scholar] [CrossRef]
- Velázquez, A.; Romero, F.J.; León, L.V.I. Fragmentación Del Hábitat Del Conejo Zacatuche. In Ecología y Conservación Del Conejo Zacatuche y Su Hábitat; Velázquez, A., Romero, F.J., López-Paniagua, Y.J., Eds.; Universidad Nacional Autónoma de México-Fondo de Cultura Económica: Mexico City, Mexico, 1996; pp. 73–86. [Google Scholar]
- Granados, H. Basic Information on the Vokano Rabbit Ln: Proceedings of the World Lagomorph Conference, Guelph 1979 (Fcis K. Myers and CA Maclnnes); University of Guelph: Guelph, ON, Canada, 1981. [Google Scholar]
- Cervantes, F.A.; Barrera, C.B. Estudios Sobre La Biología de Roedores Silvestres Mexicanos; Universidad Nacional Autónoma de México: Ciudad de México, México, 2012. [Google Scholar]
- Rizo-Aguilar, A.; Guerrero, J.A.; Hidalgo-Mihart, M.G.; González-Romero, A. Relationship between the Abundance of the Endangered Volcano Rabbit Romerolagus Diazi and Vegetation Structure in the Sierra Chichinautzin Mountain Range, Mexico. Oryx 2015, 49, 360–365. [Google Scholar] [CrossRef]
- Matthews, J.M. Effects of Wildfire Intensity on Invasives, Stand Structure and Fuel Loading in Shenandoah National Park. Ph.D. Thesis, Virginia Tech, Blacksburg, VA, USA, 2004. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
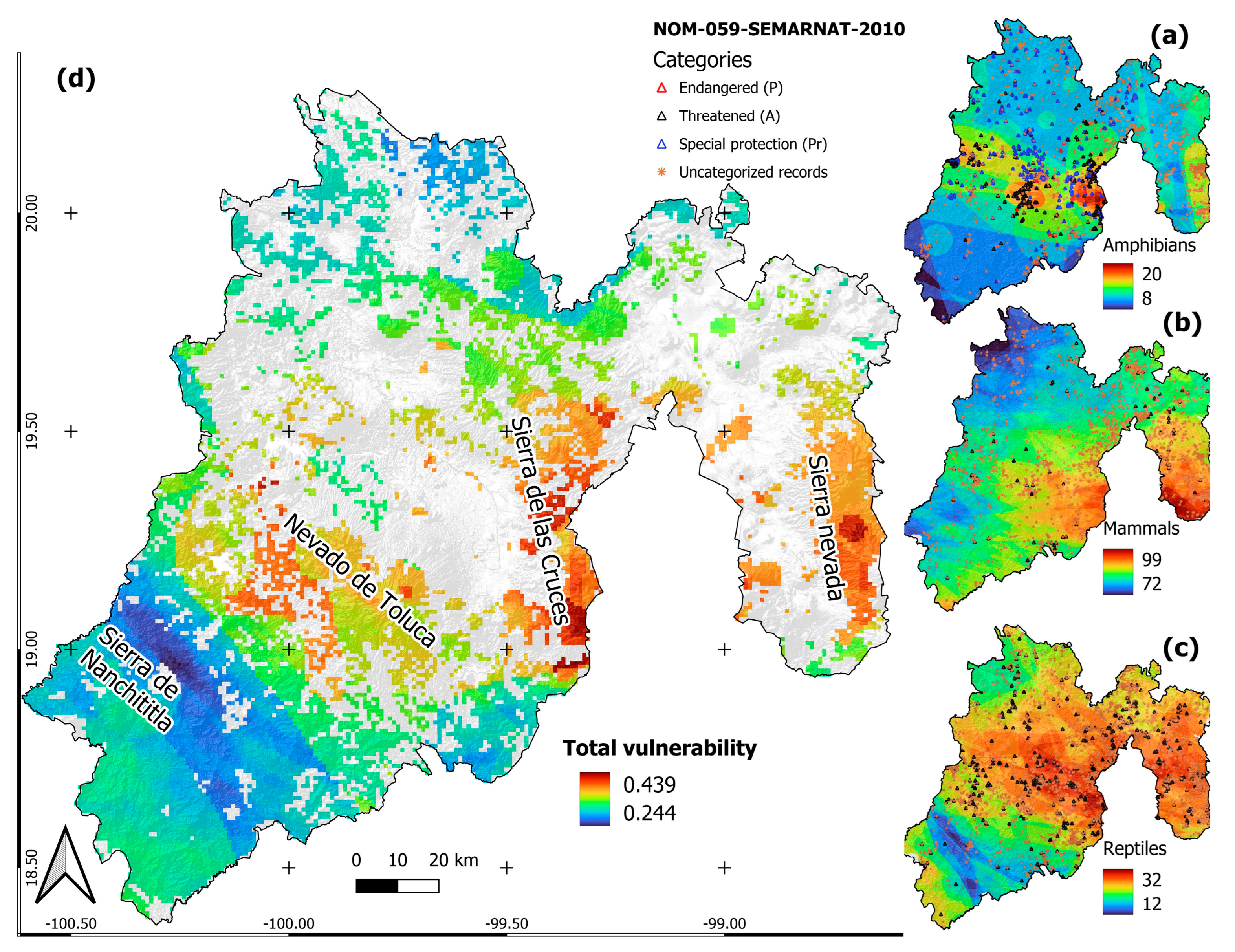{kind=link}
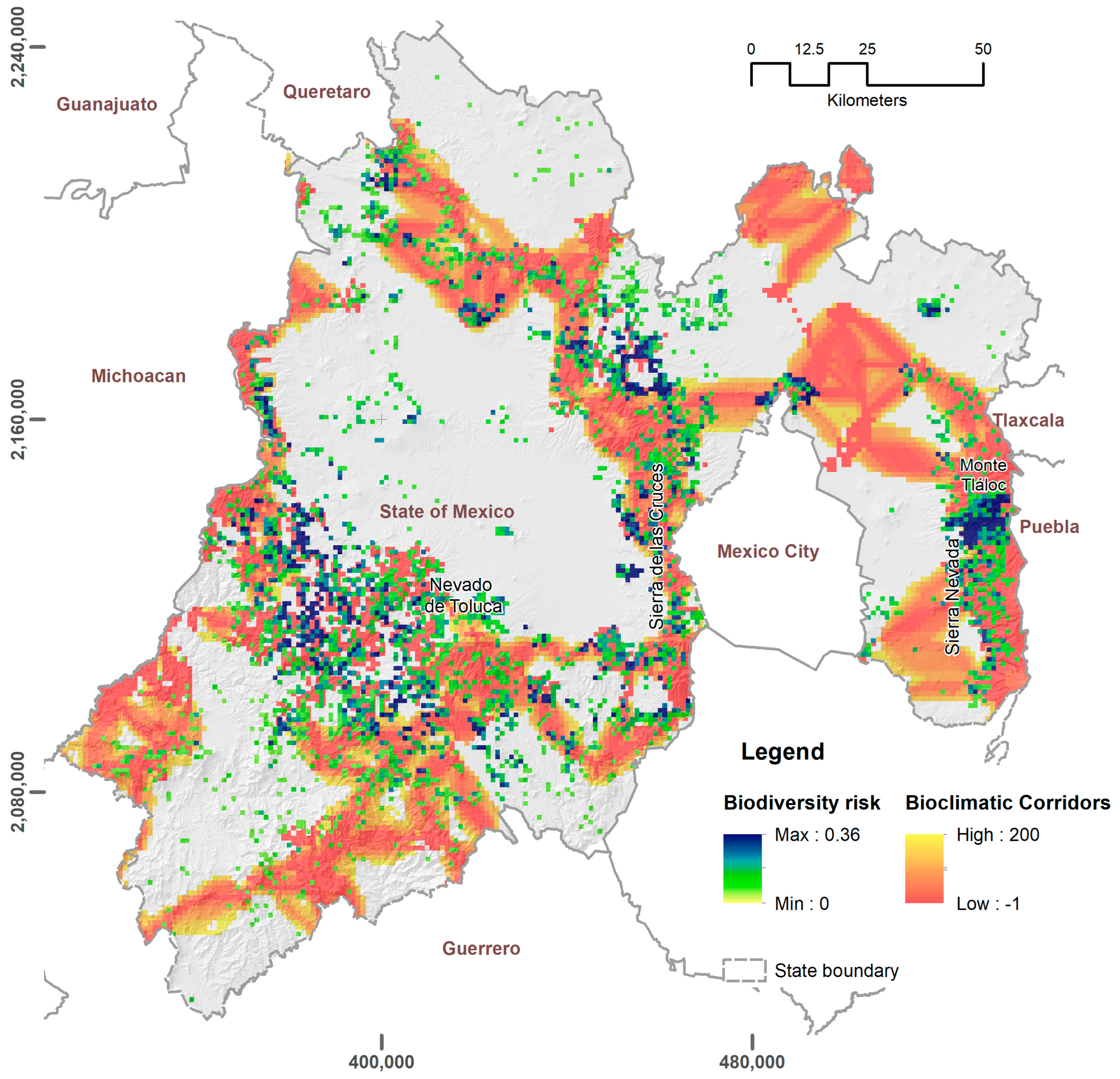{kind=link}
| Category | Name | Last Record | Records in the Last 30 Years | Natural Inhabited Area (km2) | Natural Area Burned (km2) | % Natural Area Burned | |
|---|---|---|---|---|---|---|---|
| Amphibians | CR | Ambystoma bombypellum | 2016 | 3 | 131.24 | 0.69 | 0.52 |
| Ambystoma granulosum | 1997 | 73 | 74.30 | 3.04 | 4.09 | ||
| Ambystoma leorae | 2018 | 6 | 426.57 | 52.79 | 12.37 | ||
| Lithobates tlaloci | -- | -- | 45.70 | 1.84 | 4.02 | ||
| Pseudoeurycea robertsi | 2018 | 20 | 318.43 | 22.70 | 7.13 | ||
| EN | Ambystoma altamirani | 2019 | 57 | 565.18 | 42.69 | 7.55 | |
| Ambystoma lermaense | 2011 | 64 | 8.97 | 0.00 | 0.00 | ||
| Ambystoma ordinarium | -- | -- | 863.92 | 63.12 | 7.31 | ||
| Craugastor hobartsmithi | 1997 | 7 | 94.84 | 3.33 | 3.51 | ||
| Plectrohyla pentheter | -- | -- | 1345.64 | 14.01 | 1.04 | ||
| Pseudoeurycea longicauda | 2014 | 47 | 110.57 | 3.80 | 3.44 | ||
| Pseudoeurycea tlilicxitl | 2015 | 2 | 742.45 | 65.41 | 8.81 | ||
| VU | Chiropterotriton orculus | 2019 | 21 | 1201.87 | 8.89 | 8.89 | |
| Isthmura bellii | 2019 | 7 | 6782.87 | 343.60 | 5.07 | ||
| NT | Aquiloeurycea cephalica | -- | -- | 1221.26 | 95.33 | 7.81 | |
| Lithobates neovolcanicus | 2018 | 7 | 257.67 | 9.96 | 3.87 | ||
| Mammals | CR | Habromys schmidlyi | 2006 | 2 | 4.02 | 0.0 | 0.0 |
| EN | Leptonycteris nivalis | 2000 | 18 | 10,238.87 | 395.51 | 3.86 | |
| Romerolagus diazi | 2018 | 23 | 978.95 | 109.16 | 11.15 | ||
| VU | Leptonycteris yerbabuenae | 2019 | 28 | 10,238.87 | 395.51 | 3.86 | |
| Sigmodon alleni | -- | -- | 220.18 | 0.45 | 0.20 | ||
| NT | Choeronycteris mexicana | 2019 | 1 | 10,238.87 | 395.51 | 3.86 | |
| Corynorhinus mexicanus | 2009 | 11 | 9981.36 | 395.06 | 3.96 | ||
| Leopardus wiedii | -- | -- | 1435.27 | 10.55 | 0.74 | ||
| Lepus callotis | -- | -- | 10,237.84 | 395.51 | 3.86 | ||
| Microtus quasiater | -- | -- | 363.36 | 47.72 | 13.13 | ||
| Reptiles | EN | Abronia deppii | 2019 | 5 | 3093.42 | 174.57 | 5.64 |
| Barisia herrerae | 2016 | 1 | 105.17 | 3.14 | 2.99 | ||
| Barisia rudicollis | 2018 | 12 | 1437.02 | 98.24 | 6.84 | ||
| Thamnophis melanogaster | 2019 | 32 | 3473.83 | 191.94 | 5.53 | ||
| VU | Thamnophis scaliger | 2019 | 56 | 673.97 | 44.45 | 6.60 | |
| NT | Agkistrodon bilineatus | -- | -- | 218.79 | 17.13 | 7.83 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mastachi-Loza, C.A.; Paredes-Tavares, J.; Becerril-Piña, R.; Ruiz-Gómez, M.d.L.; Rangel Patiño, C.A.; Diaz-Delgado, C. The House Is Burning: Assessment of Habitat Loss Due to Wildfires in Central Mexico. Fire 2024, 7, 134. https://doi.org/10.3390/fire7040134
Mastachi-Loza CA, Paredes-Tavares J, Becerril-Piña R, Ruiz-Gómez MdL, Rangel Patiño CA, Diaz-Delgado C. The House Is Burning: Assessment of Habitat Loss Due to Wildfires in Central Mexico. Fire. 2024; 7(4):134. https://doi.org/10.3390/fire7040134
Chicago/Turabian StyleMastachi-Loza, Carlos Alberto, Jorge Paredes-Tavares, Rocio Becerril-Piña, María de Lourdes Ruiz-Gómez, Carlos Alejandro Rangel Patiño, and Carlos Diaz-Delgado. 2024. "The House Is Burning: Assessment of Habitat Loss Due to Wildfires in Central Mexico" Fire 7, no. 4: 134. https://doi.org/10.3390/fire7040134
APA StyleMastachi-Loza, C. A., Paredes-Tavares, J., Becerril-Piña, R., Ruiz-Gómez, M. d. L., Rangel Patiño, C. A., & Diaz-Delgado, C. (2024). The House Is Burning: Assessment of Habitat Loss Due to Wildfires in Central Mexico. Fire, 7(4), 134. https://doi.org/10.3390/fire7040134