1. Introduction
The Gulf Coast Prairies and Marshes ecoregion in Texas consists of productive rangelands that provide cover and forage to wildlife and livestock [
1,
2]. This ecoregion produces a nearly continuous supply of forage because winters are relatively mild (average low and high temperatures, December through February, are 9–22 °C) [
3], and growing seasons are 289 to 300 days long [
4]. Gulf cordgrass (
Spartina spartinae [Trin.] Hitchc.) and seacoast bluestem (
Schizachyrium scoparium [Michx.] Nash var.
littorale [Nash] Gould) are native, perennial bunchgrasses that are principal components of this ecoregion [
5].
Gulf cordgrass is well adapted to soils high in salinity; it also thrives in a variety of soil textures ranging from sandy loams to clays [
6,
7]. The greatest herbage yield for Gulf cordgrass occurs during the rainy months of spring and during September and October, with its greatest standing crop yield occurring in fertile clay loams and clays because these soil textures have higher moisture-holding capabilities [
8,
9]. Although Gulf cordgrass is highly productive and can maintain green tissue year-round, livestock do not graze the mature foliage to an appreciable extent if other forages are available [
10]. Mature growth produces stiff and spine-like leaf blades of low nutritional quality [
8,
11,
12] (crude protein generally < 5%).
Seacoast bluestem nutritive value is fair to good in spring and early summer [
13,
14,
15] and provides forage for livestock throughout its wide geographic range [
15] (and references therein). With long growing seasons (plants can remain green throughout the winter), seacoast bluestem also provides forage throughout much of the year [
16,
17] and is a preferred forage in the Coastal Prairies [
2,
18,
19].
Management practices to improve forage quality of mid- and tall grasses such as Gulf cordgrass and seacoast bluestem are based on basic principles of plant growth and morphology of grasses [
20,
21,
22,
23]: the removal of fibrous mature material during strategic growth stages can promote nutritious regrowth, increase its abundance, and prolong its period of availability. Although prescribed burning often is used to implement these improvements, more information is needed on responses of these grasses to different seasons of burning. In particular, most landowners in this ecoregion who incorporate livestock grazing into their management enterprises graze cattle year-round: generally, there is no need to rest pastures in good condition to accumulate fuel loads prior to burning because of the productivity of these grasslands. Additionally, because units typically are large (>150 ha), livestock remain in burn units not only during prescribed burning but also post-burning. Thus, whereas plants experience fire and grazing simultaneously in these settings, managers have an option of when to burn—and this is the focus of our work. In particular, our objective is to document changes in forage quality [crude protein (CP) and neutral detergent fiber (NDF)] after prescribed fire in grazed pastures that were burned in winter (early February) or late summer (late July). Our goal was to develop prescribed fire recommendations regarding season of burning for improving nutritional value and utilization of Gulf cordgrass and seacoast bluestem rangelands using a patch-burn grazing design [
24,
25] where “spatially distinct patches [are] burned within larger areas” [
24] (p. 132). We hypothesized that nutritive value would improve following burning in both seasons, and that this response would differ between seasons because plants would be burned in different growth stages.
2. Materials and Methods
2.1. Study Area
The East Foundation’s El Sauz Ranch is located primarily in Willacy County, Texas, with a small portion in Kenedy County, Texas, USA. El Sauz Ranch (26.5577° N/97.4263° W) is 11,082 ha, primarily in the Gulf Coast Prairies and Marshes Ecoregion. There are numerous ecological site types in the study area including sandy flats, active sandhills, low coastal sands, and coastal sands, with coastal sands being the primary site type on the property [
26]. Soil series within the study area are: Arrada sandy clay loam, Dune land, Falfurrias fine sand, Galveston fine sand, (gently undulating), Galveston-Mustang complex, Incell clay, Lalinda sandy clay loam, Mustang fine sand, Sauz-Saucel sandy loam, and Sauz loamy fine sand [
26]. The site has a humid subtropical climate with 658 mm mean annual rainfall and an average temperature fluctuation between 18.9 and 26.7 °C [
27].
Vegetation within the study area consisted of native grasses such as Gulf cordgrass, seacoast bluestem, switchgrass (
Panicum virgatum L.), hooded windmill grass (
Chloris cucullata Bisch.), red lovegrass (
Eragrostis secundiflora J. Presl.), tumble lovegrass (
E. sessilispica Buckley), brownseed paspalum (
Paspalum plicatulum Michx.), and coastal sandbur (
Cenchrus spinifex Cav.). Forbs include partridge pea (
Chamaecrista fasciculata (Michx.) Greene), Gulf croton (
Croton punctatus Jacq.), littleleaf sensitive-briar (
Mimosa microphylla Dryand.), lavender thrift (
Limonium carolinianum [Walter] Britton), and American snoutbean (
Rhynchosia americana [Houst. ex Mill.] M.C. Metz) [
26]. Mottes of native woody species include primarily honey mesquite (
Prosopis glandulosa Torr.) and live oak (
Quercus virginiana Mill.) [
26].
El Sauz was using a moderate stocking density of cattle close to 1 AU per 14 ha that was managed as a continuous grazing system. The cow-calf operation had roughly 660 breeding cows and 27 bulls that were not provided any supplemental feed. Breeding cows were a cross between Santa Gertrudis and Beefmasters (Bos taurus) breeds.
2.2. Field Methods
In June 2015 as part of a larger patch-burn and grazing study, we delineated 10 study patches (size range: 200–305 ha) in areas dominated by Gulf cordgrass and seacoast bluestem using existing roads as firebreaks wherever possible. We used a completely randomized design, with three treatments, (1) non-burn control, (2) winter burn, and (3) summer burn, randomly assigned to patches with 2, 4, and 4 replications, respectively. We defined winter treatments as any burn conducted in January or February, whereas summer treatments were defined as any burn conducted in July or August. Control treatments were not used for this aspect of our study; we compared effects of winter burning to those of summer burning. For nutritional analyses we collected data from the two patches burned in winter 2016 and two patches burned in summer 2016. Each patch had two 60 m transects that traversed a Gulf cordgrass community and an adjacent “other” vegetative community where seacoast bluestem dominated with roughly half of each transect in each vegetative community.
We estimated fuel load (kg ha
−1) and fuel moisture (%) by gathering all aboveground biomass (standing live, standing dead, and litter) to the ground level in two, 0.25 m
2 frames randomly placed on the ground within each vegetation community near the sampling transects. We placed fuel load samples in paper bags, weighed them while wet, and transported them to a drying trailer where they dried at 40 °C until all moisture was lost. Samples were then re-weighed. Fuel moisture was calculated using the following formula:
We placed a HOBO U30/RX3000 (Onset® Computer Corporation, Bourne, MA, USA) weather station at the center of the study area to acquire daily rainfall (mm) data following burning. On burn days, we recorded weather variables of air temperature (°C), wind speed (mps), wind direction, and relative humidity (%) near the burn patch approximately every 30 min during the burn (Kestrel® 4500 Weather Meter, Nielsen-Kellerman, Boothwyn, PA, USA). We measured fine fuel load (kg ha−1), fuel moisture (%), and fire temperature (°C) in each burn patch. We recorded fire temperatures using HOBO Type J, K, T, E, R, S, B, N Thermocouple Data Loggers (Onset® Computer Corporation, Bourne, MA) with High Temperature Inconel Overbraided Ceramic Fiber Insulated Thermocouples (XCIB-K-1-2-10, OMEGA® Engineering, Norwalk, CT). We placed each data logger inside a PVC pipe section capped at both ends with the thermocouple extended through a slot in the pipe. We buried the PVC with the data logger in the ground for protection from extreme heat immediately prior to burning. We used two data loggers near each vegetation transect in each burn patch, one near the Gulf cordgrass section and one near the seacoast bluestem section of the transect. The thermocouple was placed at the ground level near the base of a Gulf cordgrass plant on that end of each transect and near the base of a seacoast bluestem plant on the other end.
Fuel load samples were kept for pre-burn nutritional analyses after dry weights were obtained. To evaluate the influence of days since burning on nutritional value after winter and summer burning treatments, Gulf cordgrass and seacoast bluestem forage samples were collected in recently burned patches every third d for 36 d, and then once weekly for another 49 d. Forage samples were obtained by randomly placing four 1 m2 quadrats in both the Gulf cordgrass and seacoast bluestem communities near each transect, and hand-clipping all Gulf cordgrass or seacoast bluestem in each quadrat to a stubble height of ~2.5 cm. All forage clipped from a quadrat was placed into a labeled paper bag. Forage samples were kept in a drying trailer at 40 °C until no further weight loss occurred and were weighed for utilization estimation prior to nutritional analyses. After weighing, the four samples of Gulf cordgrass and seacoast bluestem from each transect were combined by species to create a composite sample. Composite samples were then ground in a THOMAS® Wiley mill (Thomas Scientific, Swedesboro, NJ, USA). Ground material was mixed and placed into labeled 113 g WHIRL-PAK® bags (Nasco, Fort Atkinson, WI, USA) for nutritional analyses.
Winter burning treatments were conducted on 5 and 10 February and summer treatments on 3 and 22 July 2016. Burning began as soon as fuels were dry enough to carry a fire, typically around 1000 (GMT -6). Blacklining was typically completed by 1400, and the burning the interior of the plot by 1800. Burn patches were pasture size and irregularly shaped because existing roads were used as firelines wherever possible. Fuels were unprotected and most closely resembled fuel model GR 8 [
28] in Gulf cordgrass-dominated areas, and GR 6 or GR 7 in areas dominated by seacoast bluestem depending on recent rainfall and soil moisture. With a predominant southeast wind off the Gulf of Mexico, the lighting strategy of each patch began with blacklining occurring on the downwind west and north sides inside double mineral lines that were created roughly 30.5 m apart. With four to five torch carriers spread across the width of the area, the blacklines were ignited by walking and lighting into the wind in areas where no Gulf cordgrass occurred. In areas with thick Gulf cordgrass blacklining was completed using a backfire. Blacklining was conducted from 2 days to approximately 1 month before burning the interior of the patch because fire behavior is highly dependent on weather conditions, thus waiting for a suitable burning day is necessary. Flanking fires were used to burn the interior of the patch. Two lines of fire were simultaneously ignited using drip torches, starting in the southwest and northeast corners of the patch, lighting toward the southeast corner until they met. This created two flanking fires that pulled together in the middle of the patch.
2.2.1. Forage Chemical Analyses
Forage samples were analyzed at Texas A&M University-Kingsville for crude protein (CP) and sequential detergent fiber analysis. Van Soest et al. [
29] detergent fiber analysis of neutral detergent fiber (NDF) was determined with an ANKOM 200 Fiber Analyzer (ANKOM
® Technology Corp., Macedon, NY, USA). Total nitrogen (N) for the CP analysis (N × 6.25) was calculated using a vario MACRO analyzer in the carbon-nitrogen (CN) mode (Elementar Analysensysteme GmBH, Hanau, Germany).
2.2.2. Statistical Analyses
We used graphs [
30,
31,
32] of CP and NDF for a 90-day period following fire and locally weighted regression (loess; R Core Team, 2020, v. 4.0.30) as an “exploratory graphical tool” to gain “insight into the behavior of the data [to] help us choose parametric models” [
33] (p. 596). Preliminary analyses suggested that, for the most part, CP and NDF changed approximately linearly over time but not monotonically, with periods of rapid change until thresholds were reached, followed by subsequent period(s) of change in different directions and at different rates (
Supplemental Equations).
Based on these results, we chose as a class of parametric models piecewise linear regression [
34,
35,
36] with days after burning as an explanatory variable to describe changes in CP and NDF following fire and to test hypotheses related to rates of change within a season as well as between seasons of burning. Regression models with 0, 1, or 2 thresholds were fit for each season of burning. Our set of candidate models, which regressed CP or NDF as the response variable,
Y, on days after burning,
D, included:
where
; so that the slope of the first segment is
, and the slope of the second segment is
;
where
and
; so that the slopes of the first, second and third segments are
,
, and
, respectively (notation follows) [
34,
37]. Equations 1, 2, and 3 are for simple linear regression (no thresholds), a continuous function with one threshold, and a continuous function with two thresholds, respectively. Regressions were analyzed with the segmented package [
38] (R Core Team 2020, v. 4.0.3). We used BIC and AICc criteria for model selection [
39]. Slopes of models were compared between seasons following [
40] with the lm package and esticon statements using the doBy package [
41] in R Core Team (2020, v. 4.0.3). Influence of outliers was assessed with
dffit statistics [
34] (base package, R Core Team, 2020, v. 4.0.3). For model validation we calculated the prediction coefficient of determination (
) [
34]. Initial (pre-burn) forage values are presented graphically but were not included in the regression analyses. For data presentation, estimated slopes for a given species are denoted as, for example,
where the subscript “
W” refers to season of burn (“winter” in this example) and the sub-subscript “2” refers to the line segment (the second segment in this example).
Nutritional values estimated at 90 days after burning were compared to samples collected prior to each burn using a one-sample t test when n = 1 for pre-burn data (cordgrass NDF and seacoast bluestem NDF in summer) and a two-sample
t-test [
42] when n > 1.
3. Results
3.1. Burn Conditions
Ambient air temperature range was approximately 10 °C cooler during winter burning than summer burning (
Table 1). Wind speeds were similar during both seasons of burning. Relative humidity was lower and had a greater range during winter burns than during summer burns. Mean fuel loads and mean high fire temperature were greater in the Gulf cordgrass community than the seacoast bluestem community in both seasons.
3.2. Precipitation
Rainfall throughout the 90 d following the winter burns was 50.3 mm, and 116.3 mm following summer burn treatments (
Figure 1). Precipitation totaled 48.3 mm in the month prior to the winter burn treatments, and only 2.5 mm during the month prior to summer burn treatments.
3.3. Gulf Cordgrass CP
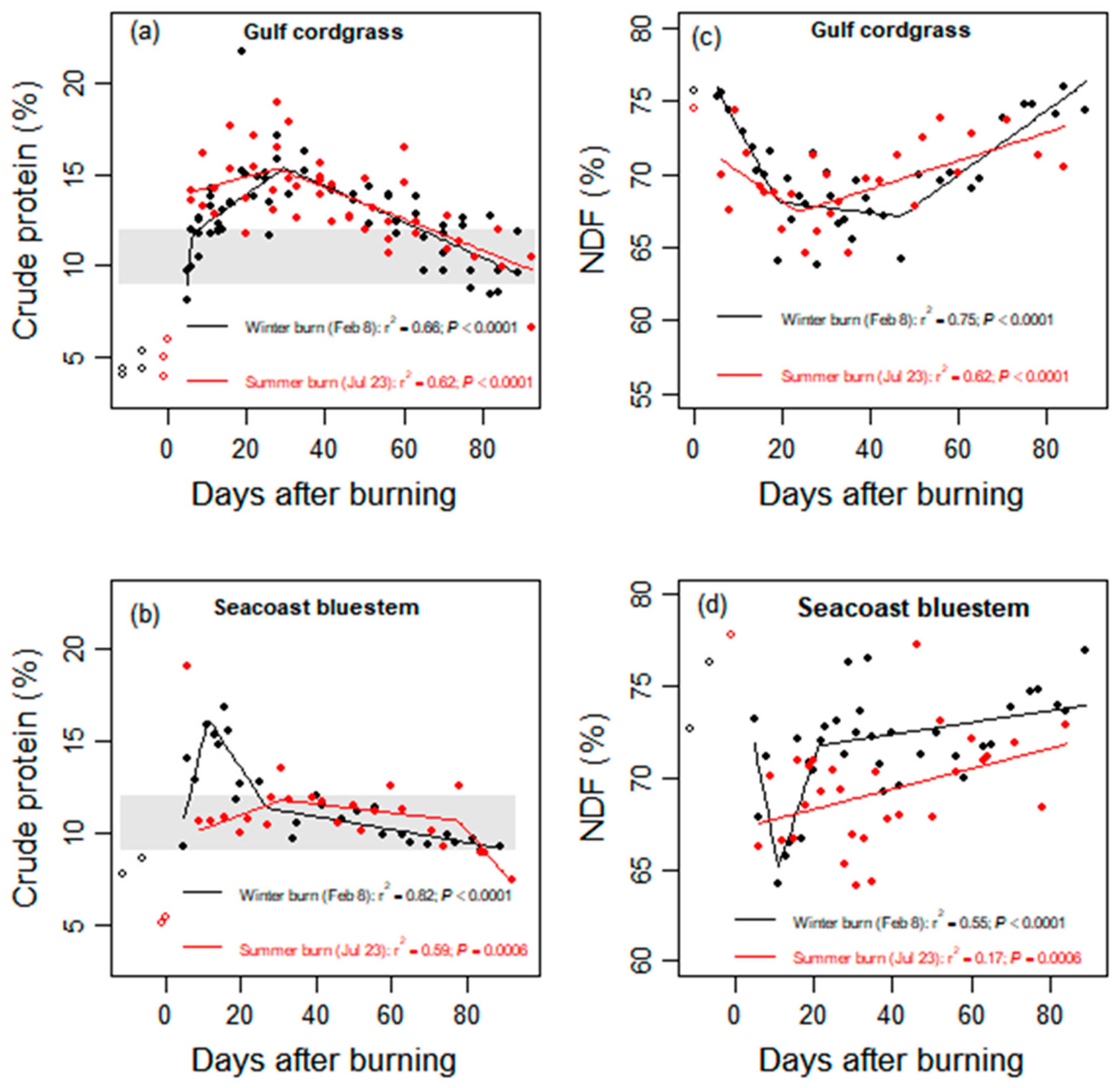
Pre-burn Gulf cordgrass mean CP levels were 4.55% (±0.29%) and 4.97% (±0.61%) for winter- and summer-burned patches, respectively (
Figure 2). Mean CP was 12.68% (±0.30%) throughout the 90-day period following winter burns (range: 8.1–21.8%); mean CP was 13.43% (±0.34%) throughout the 90-day period after summer burning (range: 6.6–18.9%;
Figure 2a).
A single datum for the winter burn with CP > 20% (for which the
dffit statistic was an order of magnitude larger than for any other observation) was removed from the data set for analysis (but is shown in
Figure 2a). The top model (
Table S1) was a two-threshold model that explained 66%
of the variation in CP. For this model, CP increased rapidly
3,
P = 0.0035) until day 6.4 (±0.07) when estimated CP reached 11.8% (±0.38). After 6.4 days, CP increased more slowly (
5.9,
P < 0.0001) until day 29.7 (±2.9) when it peaked at 15.4% (±0.36), after which CP declined (
−9.4,
P < 0.0001) to 9.5% (±0.39) at 90 days post-burning. Estimated CP at 5 DAB (8.9% ± 0.82) did not differ (
) from estimated CP at 90 DAB. A one-threshold model failed to describe the initial rapid increase in CP but modeled the decline after reaching peak CP very similarly to the two-threshold model (
Table S1).
A piecewise regression model with one threshold fit CP patterns in Gulf cordgrass following summer burns (
Figure 2a). Days after burning explained 45% of the variation in CP (
;
Table S1). Although peak CP was estimated at 15.3% (±0.43%) at 28 (±6.9) DAB, CP did not change significantly immediately following burning up to the threshold
(=0.058 ± 0.041,
t43 = 1.4,
P > 0.1621). After 28 days, however, CP decreased
= −0.086 ± 0.015,
t43 = −5.7,
P < 0.0001;
Figure 2a), reaching 10% (±0.66) at 90 DAB, which was lower (
) than estimated CP at 6 DAB (14.05 ± 0.69).
Top models for winter (two thresholds) and summer (one threshold) burning differed in form (
F3,100 = 29.7, P < 0.0001;
Figure 2a). The primary differences between seasons involved responses immediately after burning. In particular, CP increased more rapidly whether the first or the second segment of the winter response was compared to the first segment of the summer response: that is,
t100 = 2.57,
P = 0.0115, and
,
t100 = 2.10,
P = 0.0379. However, peak CP following winter burning (15.38% at 29.7 DAB) and peak CP following summer burning (15.32% at 28 DAB) did not differ (
).
Importantly, however, responses of CP following these two seasons of burning were similar in four respects. First, CP peaked at similar levels (15.4% and 15.3% following winter and summer burning, respectively). Second, peak CP was reached at similar times after burning (~30 and ~28 DAB for winter and summer, respectively). Third, after reaching peak levels, CP declined at similar rates (, t100 = −0.67, P = 0.4994) until 90 DAB. Fourth, estimated CP at 90 DAB did not differ () between winter (9.47% ± 0.44) and summer (9.99% ± 0.57) burns.
3.4. Seacoast Bluestem CP
Pre-burn seacoast bluestem mean CP levels were 8.18% (±0.51%) and 5.21% (±0.13%) in winter- and summer-burned patches, respectively. Mean CP was 11.66% (±0.23%) throughout the 90-day period following winter burns (range: 8.22–21.63%); after summer burns mean CP was 11.23% (±0.31%) throughout the 90-day period after burning (range: 1.72–32.88%;
Figure 2b).
A single datum for the winter burn with CP > 19% (for which the
dffit statistic was an order of magnitude larger than for any other observation) was removed from the data set for analysis (but is shown in
Figure 2b). The top model (
Table S1) was a two-threshold model that explained 82%
of the variation in CP. For this model, CP increased rapidly (
5.3,
P < 0.0001) until day 11.3 (±1.2) when estimated CP reached 16.2% (±0.56), followed by a decline (
−5.7,
P < 0.0001) until 27.1 (±4.7) DAB when CP was estimated at 11.36% (±0.5). Thereafter, CP declined gradually (
−2.6,
P = 0.0171) until it reached 9.12% (±0.507) at 90 DAB, a value only marginally lower (
) than estimated CP at 5 DAB (10.86% ± 0.77).
A one-threshold model for CP following summer burning had a slightly smaller AICc value than a two-threshold model, but a two-threshold model explained more variability (59%) in CP ). The two-threshold model described an initial gradual increase (2.6, P = 0.0529) in CP until 31 DAB (±8.7) when CP reached 11.8% (±0.40), followed by a long period of only slightly-declining CP (−1.59, P = 0.1293) until 78 (±5) DAB when CP reached 10.7% (±0.47). CP abruptly declined ( −3.3, P = 0.0034) and was estimated at 7.77% (±0.68) at 90 days post-burning.
The two-threshold models differed (F4,42 = 17.9, P < 0.0001) between seasons of burning. The initial increase following burning was not only shorter but also more rapid (, t42 = 4.9, P < 0.0001) for plants burned in the winter than for plants burned in the summer. Furthermore, CP peaked at higher levels () following winter burns (11.8%) than summer burns (4.39%). Winter-burned plants also experienced a shorter and more rapid (, t42 = −5.26, P < 0.0001) period of decline following peak seasonal CP than plants burned in the summer. In fact, the decline in CP during the 3rd period for winter burned plants (from 27 to 90 DAB) was similar (, t42 = −0.54, P = 0.5918) to the decline during the second period for summer burned plants (from 27 to 78 DAB). CP was only slightly higher () in summer-burned plants (10.7%) than in winter-burned plants (9.5%) 78 DAB and did not differ () at 90 DAB between winter (9.1%) and summer (7.8%) burns.
3.5. Gulf Cordgrass NDF
Gulf cordgrass mean NDF was 75.7% and 74.5% prior to winter and summer burning, respectively. Gulf cordgrass mean NDF was 70.1% (±0.57%) after winter burning (range: 63.8–76.0%); after summer burning mean NDF was 69.6% (±0.51%, range: 64.5–74.3%) (
Figure 2c).
A two-threshold model (
Table S1) that explained 75% (
F3,32 = 32.5,
P < 0.0001,
) of the variation in Gulf cordgrass NDF following winter burns estimated thresholds at 19 (±3.1) and 47 (±6.1) DAB (
Figure 2c). NDF decreased (
= −0.559 ± 0.097,
t32 = −5.7,
P < 0.0001) immediately after burning until 19 DAB, reaching an estimated 68.1% (±0.67) after which it stabilized (
= −0.035 ± 0.04,
t32 = −0.85,
P = 0.4014) until 47 DAB at an estimated 67.1% (±0.72); NDF then increased (
= 0.218 ± 0.032,
t32 = 6.7,
P < 0.0001) until 90 DAB with an estimated 76.5% (±0.96).
Gulf cordgrass NDF also decreased (= −0.204 ± 0.09, t24 = −2.3, P = 0.0334) following summer burning until 23.6 ± 6.0 DAB when it was estimated at 67.4% (±0.75). In contrast to winter burning, however, NDF after summer burning increased thereafter (= 0.096 ± 0.027, t24 = 3.6, P = 0.0016) until 90 DAB, reaching an estimated 73.8% (±1.31).
Models describing NDF responses differed (F3,56 = 11.8, P < 0.0001) between seasons of burning. First, the decrease in NDF after burning was more rapid (, t42 = −2.63, P = 0.0110) following winter burning than summer burning. However, despite a sharper decline in NDF following winter burning, minimum NDF values did not differ () between winter (67.13% at 47 DAB) and summer (67.43% at 23.6 DAB) burning. Additionally, despite the fact that NDF increased following the final threshold identified for each season of burning, this threshold occurred later following winter burning (47 DAB) than following summer burning (23.6 DAB). Finally, although the increase in NDF following minimum levels was more rapid following winter burning t56 = 2.83, P = 0.0065), NDF levels did not differ () at 90 DAB between winter (76.5% ± 1.06) and summer (73.8% ± 1.18) burning.
3.6. Seacoast Bluestem NDF
Seacoast bluestem mean NDF was 74.4% (±0.91%) and 76.8% (±0.75%) before winter and summer burning, respectively. Mean NDF was 71.4% (±0.32%) after winter burning (range: 45.3–85.7%), and 69.7% (±0.33%) after summer burning (range: 58.3–79.4%;
Figure 2d).
A two-threshold model (
Table S1) best described changes in seacoast bluestem NDF following winter burns (
F3,32 = 12.9,
P < 0.0001,
). NDF decreased from an initial estimated 72% (±1.48) 5 DAB to 65% (±1.23) at 11 ± 2.7 DAB, after which it increased relatively rapidly to 71.8% (±0.61) at 21.2 ± 2.9 DAB; thereafter, NDF increased more slowly, peaking at 74% (±0.89) at 90 DAB.
A simple linear regression best described changes in seacoast bluestem NDF following summer burns (F1,27 = 5.5, P = 0.0265, ). This model predicted an increase in NDF from an estimated 67.5% (±0.9) at 6 DAB to 72.2% (±1.34) 90 DAB but explained < 20% of the variation in NDF. Although models differed (F2,59 = 4.54, P = 0.0147) between seasons of burning, the slow increase in NDF following summer burning did not differ from the slow increase in NDF following winter burning observed between days 21 and 90 after burning , t59 = −0.79, P = 0.4336). Further, minimum NDF values did not differ () between winter (65% ± 1.43) and summer seasons (67.5% ± 0.79); and NDF did not differ () at 90 DAB following winter (74% ± 1.02) and summer (72.2% ± 1.18) burning.
3.7. Duration of Burning Effects: Pre-Burn vs. 90 DAB Nutritional Values
Cordgrass CP was higher (t3.2 = 32, P < 0.0001) at 90 DAB (9.5% ± 0.39) than prior to winter burning (9.5% ± 0.39); a similar response (t3.4 = 16, P = 0.0002) was observed at 90 days after summer burning (10% ± 0.66) compared to pre-burn values (4.97% ± 0.61). Seacoast bluestem CP also was higher (t1 = 16, P = 0.0396) at 90 days after summer burning (7.77% ± 0.68) compared to pre-burn values (5.21% ± 0.13). In contrast to cordgrass, however, we detected no difference (t1 = 2.5, P = 0.2142) 90 days after winter burning (9.12% ± 0.51) compared to pre-burn values (8.18% ± 0.51). Cordgrass NDF 90 days after winter burning (76.5% ± 0.96) was higher (t36 = 6, P < 0.0001) than prior to burning (75.7%) but lower (t26 = 2.8, P = 0.0101) 90 days after summer burning (73.8% ± 1.31) than prior to burning (74.5%). Seacoast bluestem NDF at 90 days after summer burning (72.2% ± 1.18) was lower (t28 = 21, P < 0.0001) than observed prior to burning (76.8%), but not different (t1 = 0.6, P = 0.6443) 90 days after winter burning (74 ± 0.89) compared to pre-burn values (74.4% ± 0.91).
4. Discussion
This study was not designed to test hypotheses about the interaction between fire and grazing on nutritive value: burned patches were continuously grazed by the cooperating ranch both prior to and following burning. Grazing animals [
44], fire [
45], and weather [
46] affect rangeland vegetation composition and structure as well as nutritive value e.g., [
47,
48,
49]. “Pyric herbivory”—the interaction between fire and grazing—has been documented in grasslands worldwide e.g., [
24,
50], and it is widely recognized that “As fire and herbivory both remove above-ground biomass, they clearly compete—but interactions of herbivores and fire can be more complex than this” [
49] (p. 2). When aboveground biomass of perennial grasses is removed in a single event—whether by fire or by grazing—plant regrowth from reserves stored primarily in stem bases quickly replaces aboveground tissues [
22]. We observed regrowth of Gulf cordgrass and seacoast bluestem within days after prescribed burning, a response that was apparent whether plants were burned in winter or summer and supported by McAtee et al. [
51]. It has been widely documented that this regrowth is high in nutritive value for grazing animals, and its underlying mechanisms are well understood: herbaceous regrowth is high in protein but low in structural components (fiber); as forage matures, however, fiber increases and protein decreases [
49,
52,
53,
54]. Species-specific responses arise because of differences in season of growth (e.g., cool
vs warm season species) and plant morphology (e.g., stoloniferous/rhizomatous
vs caespitose growth form).
We analyzed the nutritional content of Gulf cordgrass and seacoast bluestem following the winter or summer 2016 prescribed burning treatments. We recognized that seasonal changes in nutritional content will occur without burning, and we did not examine that, thus we cannot compare the changes we recorded after burning to non-burning. Although both Gulf cordgrass and seacoast bluestem are dominant native, warm-season bunchgrasses in southern Texas coastal prairies, they differ in growth form. Gulf cordgrass plant basal diameters average 29 cm [
51]; plants can reach 1.5 m tall [
5] and are strictly caespitose. Although basal diameters of seacoast bluestem can be 25–30 cm [
55], plants generally are smaller but also have the ability to produce rhizomes that promote lateral spread [
1].
Gulf cordgrass CP increased after burning, and although peak CP following burning did not differ between seasons, the rate at which the peak was reached was faster following winter burning. This response might be attributable to warmer-than-normal soil surface temperatures in spring (because of the removal of vegetation by fire), enhanced soil bacterial growth, and subsequent plant nutrient uptake [
56,
57]. The decline in CP following peak post-burning levels did not differ between seasons of burning, nor did CP levels differ 90 DAB between seasons of burning. Gulf cordgrass NDF showed the opposite pattern following burning, declining more rapidly following summer burning than winter burning but reaching minimum NDF values that did not differ between seasons of burning. Although NDF remained low for a longer period following winter burning (i.e., NDF began to increase nearly 20 days sooner following summer burning than winter burning), the rate of increase in NDF was faster following winter burning. Despite these differences in rates of response, 90-day NDF values did not differ between seasons. During the 90-day period following both summer and winter burning of Gulf cordgrass, crude protein content was greater than 9%, which is the minimum threshold for the maintenance of a lactating beef cow [
58]. Comparing only beginning (soon after burning) and ending (90 DAB) nutritional values between seasons of burning fails to take into account important changes—thresholds (one or two? or none?) and interval length between them—that take place throughout the 90-day monitoring period e.g., [
59]. Taken together, our results suggest that winter burning improved the forage quality of Gulf cordgrass more than summer burning: i.e., following winter burning, CP increased more rapidly, and NDF remained depressed for a longer period of time compared to summer burning. Although scientific literature is replete with general statements about the importance of season of burning on its effects on vegetation e.g., [
60] (p. 51), there has been relatively little research (<1% of 338 papers reviewed by Limb et al. [
61]) conducted on effects of fire on nutritive value. For example, season of burning has been shown to affect the nutritive value of purple threeawn (
Aristida purpurea; [
62]) but had no effect on CP in grasses in South Africa [
63].
Although CP in seacoast bluestem also increased following winter burning, there was relatively little change following summer burning, a result that is difficult to explain given that (1) a common response to defoliation (whether through grazing or burning) of grasses is stimulation of new leaf growth that is higher in CP than older leaf material, and (2) summer-burned plants experienced several ~1-cm rainfall events within ~45 DAB. Buttery and Ehreneich [
64], however, cited and confirmed results of Campbell et al. [
65] that
Andropogon divergens crude protein increased when plants were burned in a young leaf stage but had no effect when plants were burned in full or mature leaf stages. As with Gulf cordgrass, the CP response of seacoast bluestem following winter burning may be attributable to increased soil surface temperatures following burning and enhanced soil microbial activity. The rapid and substantial increase in CP following winter burning, coupled with concomitant low NDF, suggest that, like Gulf cordgrass, seacoast bluestem responded more favorably to winter than to summer burning. Similar to Gulf cordgrass, the differences between seasons of burning on seacoast bluestem nutritive value were most apparent soon after burning; after respective thresholds were reached, declines in CP and increases in NDF were similar between seasons of burning.
Duration of burning effects have important management implications. In our study, Gulf cordgrass CP was ~5% higher at 90 days after burning compared to pre-burn values for both winter- and summer-burned plants. Seacoast bluestem CP was ~2.5% higher 90 days after summer burning but neither higher nor lower 90 days after winter burning. NDF of both Gulf cordgrass and seacoast bluestem was lower 90 days after summer burning compared to pre-burn values; NDF values following winter burning were higher in cordgrass and not different in seacoast bluestem. Although statistical differences in duration of burning effects on CP were large enough to have management implications, it is not likely that duration of burning effects on NDF, even when statistically significant, were large enough to have practical management implications.
Prescribed fire can be used to accomplish a number of different management objectives in coastal grasslands of southern Texas. For example, late summer burns might be more effective than early winter burns in providing wintering habitat and resting stops for migratory birds in the Gulf Coast region of Texas [
66]. However, human health hazards associated with summer prescribed burning in this region (heat exhaustion and heat stroke: [
67]) are also important considerations. These can be mitigated by burning during more tolerable winter conditions.


 and
and 


{kind=link}
{kind=link}