Abstract
Street trees provide ecosystem services such as heat mitigation, improved community well-being, and biodiversity conservation. At the wildland–urban interface (WUI), high-flammability street trees also provide a conflicting ecosystem disservice, heightening risks of wildfire spread into urban areas. We addressed this service–disservice conflict by assessing shoot flammability patterns in 10 street tree species, to identify low-flammability species that can potentially mitigate wildfire risks at the WUI. We found significant differences among species in flammability attributes including time-to-flame (TTF), flame duration (FD), number of flaming events (nF), and flame temperature (FT), and identified low-flammability species for each attribute. Overall, species’ rankings from least to most flammable differed considerably across the four attributes. For example, native water gum (Tristaniopsis laurina) had the slowest TTF, but had the longest FD. Among nine shoot traits, we found that high leafing intensity was the most frequent trait correlated with flammability. In particular, high leafing intensity was significantly related to fast TTF and high FT. Lack of coordination among flammability attributes suggests that, in general, selection of low-flammability street tree species should consider how each flammability attribute differentially contributes to wildfire spread risk. Nonetheless, native Tuckeroo (Cupaniopsis anacardioides) emerged as a potential candidate for further exploration as a low-flammability street tree as it had comparatively long TTF, short FD, and low nF. We found no consistent evidence that exotic species were less flammable than native species, and suggest that native trees be the focus of further research to identify low-flammability street trees.
1. Introduction
At the wildland–urban interface (WUI), urbanized landscapes sit within areas of undeveloped wildland vegetation [1]. Wildfires that spread from wildland to urban areas pose a serious risk for residents living at the WUI [2]. While houses and infrastructure can propagate wildfires into urban areas [3], vegetation in recreational parks and home gardens, and trees planted along streets, can also provide fuel for wildfire spread [4]. In many regions of the world, urban greening strategies are increasing tree canopy cover to combat urban heat island (UHI) effects [5,6,7]. As a result, efforts to increase street tree coverage, which also increase vegetative fuel load, may lead to urban areas at the WUI becoming more vulnerable to the impacts of wildfires. Indeed, previous research has found that areas of the WUI with low residential housing density and broader tree coverage are at greater risk from wildfires [8,9,10].
Urban tree plantings are vital for communities as they provide residents with a range of ecosystem services [11]. In addition to mitigating UHI effects, tree ecosystem services include positive impacts on the health of residents [12], opportunities for urban permaculture that contribute to sustainability and strengthening of community ties [13,14], and biodiversity conservation [12,15]. However, tree ecosystem services can be offset by a range of disservices [16], including safety risks to residents from falling branches, and damage to infrastructure from tree roots [17,18]. The increase in vegetative fuel load from street tree plantings can also provide an ecosystem disservice, because high-flammability tree species contribute to risks of wildfire spread [19]. In contrast, despite increasing vegetative cover, plantings of low-flammability tree species could reduce wildfire risks, providing an ecosystem service in the form of opportunistic green firebreaks that help to slow or stop the spread of wildfires in urban regions [20,21,22,23,24,25,26,27]. As a first step to address this service–disservice tree conflict, flammability assessments of street tree species are needed to distinguish low-flammability from high-flammability species. In this way, evidence-based selection of tree species is prioritized to maximise the potential for the beneficial service of green firebreaks.
In this study, we assessed shoot flammability patterns in five native and five exotic species widely used as street trees in the Central Coast region of New South Wales (NSW), Australia. The aims of this study were (1) to compare and rank the ten species based on their relative flammability to distinguish low-flammability from high-flammability species; (2) to quantify relationships between a range of shoot traits and shoot flammability; and (3) to determine whether shoot flammability differs between native and exotic tree species. There is a growing body of research exploring shoot flammability patterns among species in non-WUI areas (e.g., Argentina, [28]; Australia, [19,29]; New Zealand, [30,31,32]; South Africa, [33,34,35]). However, there have been no studies that specifically examine patterns of shoot flammability among species that are commonly planted as street trees in areas of the WUI, despite such information being of critical importance for understanding wildfire risks posed by street trees. Our identification of plant traits defining low-flammability species can be used to suggest other species worthy of further exploration as low-flammability species based on the traits that they possess [19,36].
2. Materials and Methods
2.1. Study Region and Species
Our study region was the Central Coast Local Government Area (LGA), on the east coast of NSW (33°31′55″ S, 151°10′51″ E). The Central Coast LGA is one of eastern Australia’s largest urban population centres (Figure 1). The LGA covers 1681 km2 and has a population of 346,596 people, which is predicted to increase to 400,000 by 2036 [37,38]. The region is characterised by an urban to peri-urban landscape primarily composed of separate low-density houses (78.4% of dwellings in the LGA) [38] and National Parks covering over half of the LGA [39]. Risks of wildfire incursion into urban areas are posed from surrounding fire-prone dry sclerophyll forest, which is the dominant wildland vegetation at the region’s WUI [40]. The region was heavily impacted by the 2019–2020 ‘Black Summer’ bushfires along eastern Australia, when 459 km2 (27%) of the LGA was burnt [41]. The region has a fire season of six months over spring and summer (September to February), when the average daily maximum temperatures are 21 °C and 33 °C, respectively, with humidity averaging between 70–90% throughout the year [42].
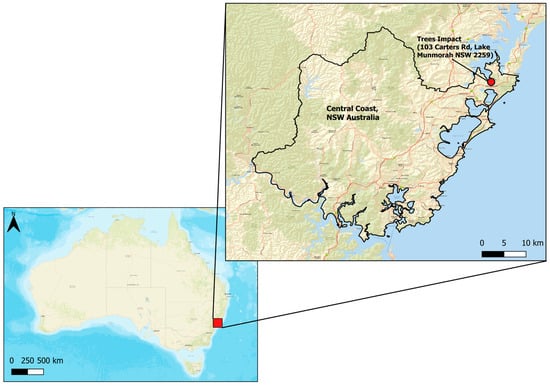
Figure 1.
Map of the Central Coast study region in New South Wales in eastern Australia.
Approximately 70% of the land surface of the LGA is covered by plants, with nearly 20% of the vegetation found in home gardens, recreational parks, and as street tree plantings in residential and city-centre areas [39]. Mature trees are often commissioned by Central Coast Council for street plantings from specialised tree nurseries as part of urban greening projects [27]. In consultation with the council, we selected five native and five exotic street tree species commonly occurring in the LGA as street trees (Table 1). All 10 study species grow taller than 5 m, with branches concentrated high on the trunk, allowing ease of passage for pedestrians and cars, while providing shade and offering shelter for wildlife. The species are all broad-leaved, which aids in the creation of shade to mitigate urban heat island effects [6] and the provision of shelter to support native bird biodiversity [43]. We collected shoot samples of the study species for our flammability experiments from Trees Impact (Lake Munmorah, Central Coast NSW) (Figure 1), a large wholesale tree nursery that is the only dedicated containerised large-tree grower in eastern Australia. The widespread use of the study species as street trees in the region was confirmed by Trees Impact, with the 10 species among those most commonly commissioned for large-scale plantation projects.

Table 1.
The ten street tree species assessed for shoot flammability with their common name, taxonomic family, and status as either native or exotic in Australia.
2.2. Shoot Collection
Trees Impact provided access to their nursery for collecting shoots of the 10 species in the summer (January–February) of 2023. We collected shoot samples from the exposed terminal branches of healthy, mature, and well-watered individuals, consistent with previous assessments of shoot flammability [29,30,44]. Shoots were sampled from nine replicate individual plants of each species [19,45]. Two representative and adjacent 50 cm shoot samples were collected from each plant. One shoot was the ‘burn’ shoot and used in flammability assessments, while the other shoot was the ‘trait’ shoot and used to measure a range of shoot traits. All shoots possessed multiple leaves, twigs, and small branches to include key factors influencing the flammability of vegetation such as fuel arrangement, continuity, and quantity [19,46,47]. Immediately after sampling, the cut ends of shoots were wrapped in dampened cloth, and shoots were placed in sealed plastic bags. Shoot samples were stored at room temperature overnight before experimental assessment of shoot flammability the following day. Previous studies have allowed shoot samples to air-dry overnight to increase the likelihood that shoots of all species ignite under experimental conditions [29,30,48,49]. This drying procedure was unnecessary in the present study as initial pilot studies showed that all 10 study species ignited within minutes of exposure to fire without any prior drying treatment.
2.3. Measurement of Shoot Flammability and Shoot Traits
Shoot flammability experiments were performed using a portable device (see Supplementary File S1) following the standardised design of Jaureguiberry et al. (2011) [44] and modified by Wyse et al. (2016) [30]. We measured four flammability attributes including the time taken for a shoot to begin flaming combustion (time-to-flame, TTF); the length of time a shoot spent in flaming combustion (flame duration, FD); the number of times a shoot entered flaming combustion (number of flaming events, nF), which included the initial flaming event and any other times the shoot entered flaming combustion after having ceased flaming; and the maximum heat of combustion of a shoot (flame temperature, FT). Observations ceased when a shoot did not enter flaming combustion again after two minutes. Our assessments focused on shoot flammability as it was not feasible to burn replicates of whole large trees of the study species [44]. While flammability testing of entire plants can provide a direct assessment of whole-plant flammability [50], recent research has demonstrated that flammability assessments of shoots from plant canopies provide an efficient and robust assessment of whole plant flammability [30]. In this context, Alam et al. (2020) [47] compared a ranking of shoot flammability among 42 indigenous New Zealand plant species to a ranking of the same species derived from elicitation of expert opinion of fire managers (based on field observations by 59 fire managers of the burning characteristics of species during wildfires or prescribed burns across New Zealand; [51]), and found a good correlation between the two rankings.
The LPG-powered flammability device provided heat when lit to radiate from below and through each burn shoot, with shoots burned one at a time. Each of a species’ nine replicate burn shoots was placed horizontally on a mesh grill in the device set 20 cm above the flames, ensuring that overall shoot architecture was kept intact. The grill temperature was maintained at approximately 185 °C. As soon as a shoot was placed on the BBQ grill above the flames, a timer was started to measure TTF and FD. We recorded nF for each shoot, with FD calculated as a summed value of time across all separate flaming events. A handheld infrared laser thermometer (Digitech QM 7226) was used at a distance of 50 cm from the shoot sample to measure the maximum temperature (FT) reached by the external flame plume [19,30,34,45,52].
We weighed each burn shoot prior to flammability assessment using a portable scale spring balance to determine shoot mass (g), and measured shoot length, width, and height (cm) to estimate shoot volume (cm3) as an elliptical cylinder (π × shoot length × shoot width × 0.5 × shoot height × 0.5). We also measured minimum and maximum stem thickness (cm) and counted the number of leaves (leafing intensity) and branches (branching intensity) on each burn shoot. The nine replicate trait shoots collected for each species were weighed (using a Sartorius A 120 S Analytical Balance) to determine fresh mass (g), placed in a drying oven at 80 °C for 48 h, then weighed again to determine shoot dry mass (g). Shoot fuel moisture content was expressed as a percentage and calculated as the ratio of the difference between air-dried and oven-dried mass to air-dried dry mass ([air-dried mass–oven-dried mass]/air-dried mass). Shoot bulk density (g.cm−3) was calculated as shoot air-dried mass divided by shoot volume. Leaf area (mm2) was measured on five representative leaves from each trait shoot using a portable leaf area meter (LI-3000A).
2.4. Data Analysis
We examined relationships between all pairwise combinations of the four flammability attributes using species’ mean data (all sqrt transformed) (Figure 2). With a critical value of p for significance of 0.05, five of the six bivariate correlations in the matrix were non-significant (p > 0.05), while the sixth between TTF and FT was only marginally significant (p = 0.048). Consequently, we elected to explore shoot flammability patterns separately for each flammability attribute rather than use a principal components analysis to reduce the attributes into a smaller subset. We fitted a linear model on each flammability attribute, using species as a categorical factor (ten-level fixed factor), to determine whether there was significant variation among species in shoot flammability. For these models, TTF was ln transformed and nF was sqrt transformed to meet the assumptions of normality and homogeneity of variances in linear models, FD and FT were used without transformation, and all replicate observations were used (nine per species). Where the species term was significant, Tukey’s pairwise tests (with correction for family-wise error rate) were used to determine patterns of differences between species. Paired-sample Wilcoxon tests were performed on modelled values for species’ means to determine if flammability rankings were consistent between all pairs of flammability attributes. For this test, FD, nF, and FT were made negative and TTF left as is, so that a rank closer to one (out of ten) indicated faster TTF, longer FD, more flaming events (nF), and higher temperature.
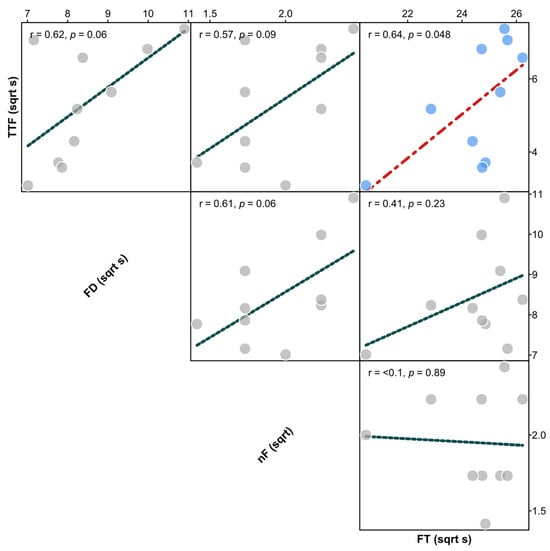
Figure 2.
Correlation coefficients (r) with p values of bivariate relationships between time-to-flame (TTF), flame duration (FD), number of flaming events (nF), and flame temperature (FT). A red line with light blue points shows a significant relationship, while a blue line with grey points shows a non-significant relationship.
To identify trait drivers of species-level patterns in shoot flammability, we first built a correlation matrix (Table 2) of the shoot traits that we measured using species’ mean data for shoot mass; bulk density; branching and leafing intensity; fuel moisture content (ln transformed); volume, and minimum and maximum stem thickness (all sqrt transformed); and untransformed values for leaf area. Variables uncorrelated with other variables (i.e., r < 0.70) were then selected for further analysis (bulk density, leafing intensity, volume, fuel moisture content, minimum stem thickness, and LA). Shoot mass, branching intensity, and maximum stem thickness were omitted from further analyses as they were highly correlated with other traits (Table 2). As the number of candidate traits (six) used in our analyses was relatively high given the number of species (ten), we used an information theoretic approach to identify the most explanatory candidate model containing four or fewer shoot traits and a term for species (all observations were used). A suite of models were then fitted to each flammability attribute using all unique four, three, and two term combinations of the selected shoot traits, as well as models using individual shoot traits, with all models including a controlling term for species. The most explanatory model was then selected on the basis of having the lowest Bayesian information criterion (BIC, chosen for its strong penalisation for number of model terms relative to other metrics, e.g., Akaike’s information criterion). This approach allowed identification of the subset of traits driving shoot flammability while also favouring more parsimonious models. The trait models identified using the information theoretic approach were then tested for statistical significance using multiple regressions. We then fitted a linear model on each of these shoot traits, using species as a categorical factor (ten-level fixed factor), to determine whether there was significant variation among species in the traits. The same transformations of shoot traits were used as described above.
Table 2.
Correlation matrix of all pairwise relationships between shoot traits. Correlation coefficients (r) are presented above the line of equivalence and p values below. BD = bulk density, BI = branching intensity, LI = leafing intensity, FMC = fuel moisture content, MinST = minimum stem thickness, MaxST = maximum stem thickness, LA = leaf area.
To examine the relationship of plant native or exotic status to each of the four shoot flammability attributes, we fitted a linear mixed model on each flammability attribute separately with a fixed term for status (two-level factor; exotic or native species) and a random term for species (nine observations per species). We then fitted the same linear mixed model structure (fixed term for status, random term for species) to each shoot trait separately, to determine native–exotic differences in shoot traits, that underpinned any significant native–exotic differences in shoot flammability. For these two sets of analyses exploring native–exotic patterns, we used the same transformations as in the previous analyses.
All analyses were performed in R 4.2.1 [53], using the packages car 3.1-2 ([54]; ANOVA for both linear and random effects models), emmeans 1.8.7 ([55]; estimates of marginal effects and pairwise testing), and lme4 1.1-34 ([56]; random model fitting).
3. Results
3.1. Comparison of Shoot Flammability Attributes among Species
There was a significant difference in TTF among species (F9,80 = 16.63, p < 0.001; Figure 3a). The fastest species to ignite were the exotics J. mimosifolia (median = 9.52 s) and Q. palustris (median = 12.78 s), which demonstrated significantly (p < 0.05) faster TTF than all native species (T. laurina median = 54.27 s; C. anacardioides median = 49.86 s; B. integrifolia median = 46.45 s; L. confertus median = 43.26 s; C. maculata median = 31.79 s). A larger variance in TTF occurred among species than within species (65% vs. 35%).
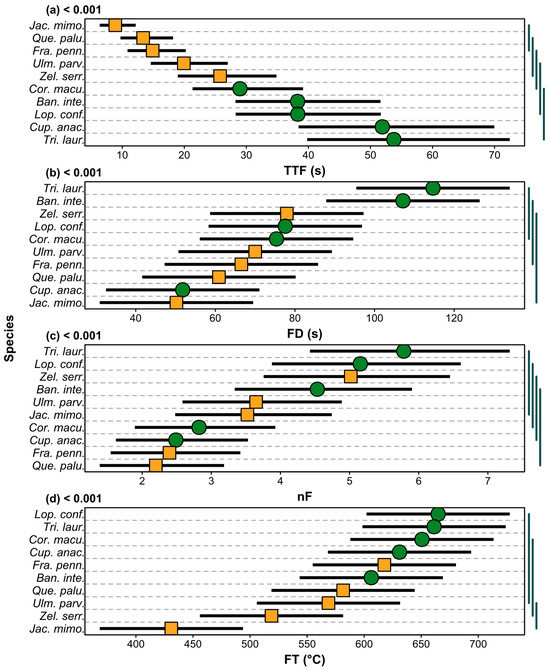
Figure 3.
Comparisons among species of (a) time-to-flame (TTF), (b) flame duration (FD), (c) number of flaming events (nF), and (d) flame temperature (FT). Exotic species are represented in orange, native species in green. Squares and circles show mean values as estimated by models with 95% confidence intervals (black lines). Presented are p values from linear models. Lines on the right-hand side group together species that are not significantly different (p > 0.05) based on pairwise tests (with Tukey correction).
A significant difference in FD was found among species (F9,80 = 5.01, p < 0.001; Figure 3b). Here, a larger variance in FD occurred within species than across species (64% vs. 36%). Nevertheless, two native species T. laurina (median = 118.88 s) and B. integrifolia (median = 99.69 s) sustained significantly (p < 0.05) longer flaming combustion than the three species that flamed for the shortest time, which were the exotics J. mimosifolia (median = 49.17 s) and Q. palustris (median = 61.78 s), and the native C. anacardioides (median = 51.22 s).
There was a significant difference in nF among species (F9,80 = 5.08, p < 0.001; Figure 3c). As with FD, a larger variance in FD occurred within species than across species (64% vs. 36%). However, two native species T. laurina (median = six flame events) and L. confertus (median = five flame events) had significantly (p < 0.05) higher nF than the exotics Q. palustris (median = three flame events) and F. pennsylvanica (median = three flame events), as well as the native C. anacardioides (median = three flame events).
We found a significant difference in FT among species (F9,80 = 5.57, p < 0.001; Figure 3d). A larger variance in FT occurred within species than across species (61% vs. 39%). Indeed, all but one species had highly overlapping median flame temperatures between 500 °C and 700 °C. The exception was J. mimosifolia which had the lowest FT (median = 419 °C), overlapping with the exotics Z. serrata (median = 522 °C) and U. parvifolia (median = 594 °C), but differing significantly (p < 0.05) from the other seven species of Q. palustris (median = 607 °C), B. integrifolia (median = 611 °C), F. pennsylvanica (median = 618 °C), C. maculata (median = 646 °C), T. laurina (median = 653 °C), C. anacardioides (median = 659 °C), and L. confertus (median = 688 °C).
Overall, species’ rankings from most to least flammable differed significantly between all pairs of flammability attributes, with little evidence of concordance in the ranking of species between all possible pairs of TTF, FD, nF, and FT (Figure 4). For example, Q. palustris ranked second most flammable for TTF but eighth for FD; T. laurina ranked the least flammable (tenth) of all species for TTF yet most flammable (first) for nF; J. mimosifolia ranked most flammable for TTF yet least flammable for FT; C. maculata ranked fifth most flammable for FD but seventh for nF; C. anacardioides ranked fourth most flammable for FT but ninth for FD; and U. parvifolia ranked fifth most flammable for nF and eighth for FT.
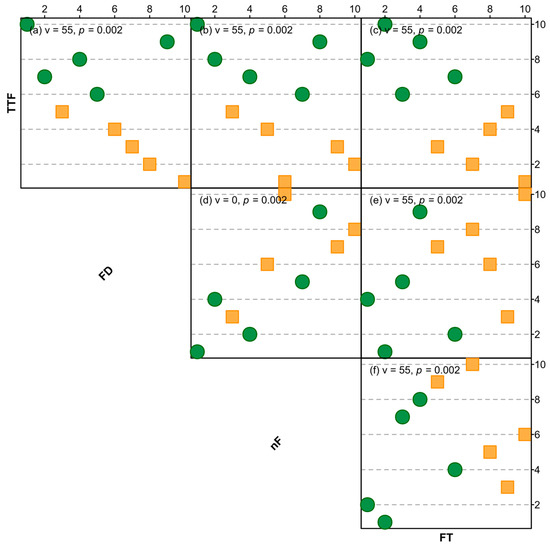
Figure 4.
Rank correlations of bivariate relationships between time-to-flame (TTF), flame duration (FD), number of flaming events (nF), and flame temperature (FT). Exotic species are represented with orange squares, native species with green circles. Each relationship is presented with v and p values from Wilcoxon signed rank tests. Values of FD, nF, and FT were made negative and TTF left as is, so that a rank closer to one (out of ten) indicates faster TTF, longer FD, more flaming events (nF), and higher temperature.
3.2. Relationships between Shoot Traits and Shoot Flammability
The best explanatory model for TTF had a BIC value lower than the saturated trait model and contained three shoot traits (Table 3). Faster TTF was significantly related to lower shoot bulk density, greater leafing intensity, and smaller shoot volume (Figure 5a–c). Only one shoot trait, bulk density, was retained in the best explanatory model for FD (Table 3). While this model indicated a trend for a relationship between longer FD and higher shoot bulk density (Figure 5d), this relationship was not statistically significant (Table 3). The best explanatory model for nF had a BIC value lower than the saturated trait model and contained one shoot trait (Table 3). A larger number of flame events was significantly related to lower leafing intensity (Figure 5e). For FT, the best explanatory model had a BIC value lower than the saturated trait model and contained three shoot traits (Table 3). Higher FT was significantly related to greater leafing intensity, higher fuel moisture content, and larger stem thickness (Figure 5f–h). All shoot traits that were significantly related to shoot flammability were found to differ significantly among species, including bulk density (F9,80 = 3.12, p = 0.003; Figure 6a), leafing intensity (F9,80 = 18.63, p < 0.001; Figure 6b), volume (F9,80 = 6.16, p < 0.001; Figure 6c), FMC (F9,80 = 12.74, p < 0.001; Figure 6d), and stem thickness (F9,80 = 14.66, p < 0.001; Figure 6e).
Table 3.
Multiple regression results of the significance of shoot traits for shoot flammability. The combination of traits included for each attribute are those generating the lowest BIC values from information theoretic models. TTF = time-to-flame, FD = flame duration, nF = number of flaming events, FT = flame temperature.
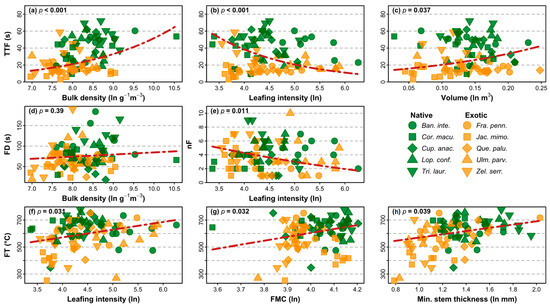
Figure 5.
Relationships between flammability attributes and shoot traits (a–h) with p values from multiple regressions. Exotic species are represented in orange, native species in green. Species are represented by symbols of different shape and colour combinations. The traits included for each attribute are those generating the lowest BIC values in information theoretic models. TTF = time-to-flame, FD = flame duration, nF = number of flaming events, FT = flame temperature, FMC = fuel moisture content.
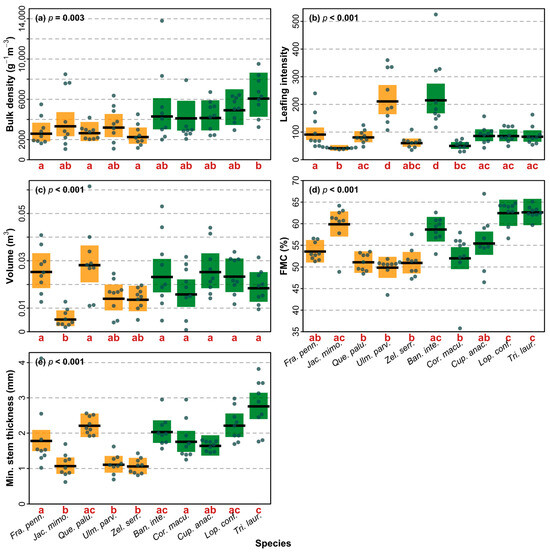
Figure 6.
Shoot trait differences among species for the traits that were significantly related to one or more of the flammability attributes. Exotic species are represented in orange, native species in green. Different letters under species in each graph indicate that the species differ significantly from each other based on pairwise tests (with Tukey correction). The mean (black line) with 95% CI (shading) is shown with observations for species as points, and p values from linear models. FMC = fuel moisture content.
3.3. Shoot Flammability Differences between Native and Exotic Species
We found that TTF was significantly faster in exotics compared to natives (χ2 = 20.75, DF = 1, p < 0.001; Figure 7a), and that FT was significantly higher in natives compared to exotics (χ2 = 8.49, DF = 1, p = 0.004; Figure 7d). There were no significant differences in either FD (χ2 = 2.67, DF = 1, p = 0.10; Figure 7b) or nF (χ2 = 0.89, DF = 1, p = 0.35; Figure 7c) between native and exotic species. Two shoot traits were found to differ significantly between natives and exotics, including bulk density (χ2 = 21.70, DF = 1, p < 0.001; Figure 8a) and stem thickness (χ2 = 4.62, DF = 1, p = 0.03; Figure 8b). None of the other shoot traits differed significantly between natives and exotics (volume χ2 = 1.01, DF = 1, p = 0.31; leafing intensity χ2 = 0.09, DF = 1, p = 0.77; fuel moisture content χ2 = 3.64, DF = 1, p = 0.06; leaf area χ2 = 0.25, DF = 1, p = 0.62).
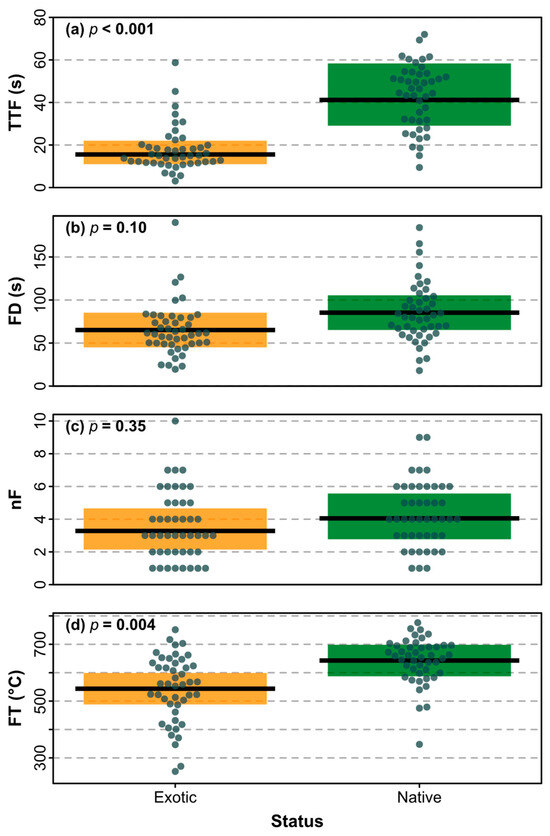
Figure 7.
Comparisons of flammability attributes (a) time-to-flame (TTF), (b) flame duration (FD), (c) number of flaming events (nF), and (d) flame temperature (FT) between exotic (orange) and native (green) species. The mean (black line) with 95% CI (shading) is shown with observations for species as points, and p values from linear mixed models.
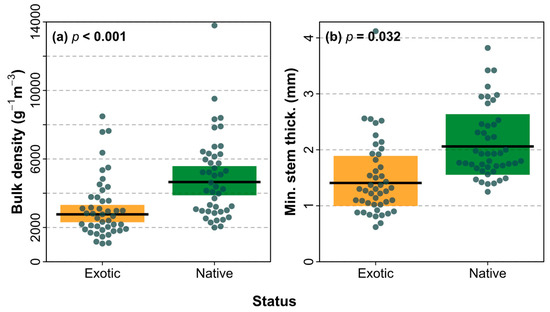
Figure 8.
Comparisons of shoot traits (a) bulk density and (b) minimum stem thickness between exotic (orange) and native (green) species. The mean (black line) with 95% CI (shading) is shown with observations for species as points, and p values from linear mixed models.
4. Discussion
4.1. Shoot Flammability Differences between Native and Exotic Species
Our results demonstrate that shoot flammability differs among common street tree species of eastern Australia. This finding contributes to a relatively small but growing body of evidence showing that species vary considerably in the intrinsic flammability of their shoots [19,28,29,30,31,32,33,34,35,44]. The differences that we observed in all four flammability attributes among species provide an important first step for evidence-based selection of low-flammability street tree species at the WUI to minimize the risk of wildfire incursion into urban areas. At the same time, our experiments have distinguished high-flammability street tree species that may increase fire incursion risks. Importantly, we found that rankings of species from low-flammability to high-flammability were not consistent across the four flammability attributes. For example, native water gum (T. laurina) took the longest time to ignite (i.e., a low-flammability characteristic), yet burned for the longest period of time (i.e., a high-flammability characteristic). Our finding for a lack of coordination among the four flammability attributes effectively means that selection of low-flammability street tree species needs to consider carefully how each flammability attribute may differentially contribute to fire risk at the WUI [24].
Time to flame is a critical flammability attribute that provides a relative measure of ignition delay times of among species [30,57]. There are two advantages of street trees that have relatively long ignition delay times in the context of preventing the spread of wildfires into urban areas. First, fire conditions might change during a longer delay period, which means that some slow-igniting trees might not ignite, thus, reducing the risk of localised fires. Second, if trees do ignite, slow-igniting trees provide more time for organised efforts to cover large areas to eliminate spot fires (i.e., fires started by flying sparks or embers at a distance from the main fire). Street trees identified in the present study as having the longest ignition delay times (median TTF > 50 s) were the native species water gum (T. laurina) and tuckeroo (C. anacardioides). Coastal banksia (B. integrifolia) and brush box (L. confertus) also had relatively long ignition delay times (median TTF just under 40 s). In contrast, the shortest ignition delay times (median TTI < 15 s) were observed in the exotics jacaranda (J. mimosifolia) and pin oak (Q. palustris), which suggests that these species may pose immediate fire risks at the WUI. Jacaranda in particular is often planted or retained in urban settings in the east coast of Australia based in part on its favoured ornamental features and cultural values [58]. One potential way to mitigate its fire risk could be to ensure that jacaranda canopies do not overlap with other trees to minimise the risk of rapid fire spread. In addition, previous work has demonstrated that the proximity of vegetation to houses influences fire impact, with homes situated less than 40 m to vegetation more prone to fire damage [10]. We suggest that further research on plant flammability in the context of opportunistic green fire-breaks should focus on how and where to plant low-flammability species in this context.
The length of time over which flaming combustion occurs, which is a measure of sustainability [57], describes the ability of a fuel to keep burning and sustain the spread of fire [46]. The primary advantage of street trees that have relatively short flaming times is reducing the likelihood that wildfires will spread to other vegetation, homes, and infrastructure within urban areas. Despite being identified as low-flammability species in terms of ignitibility in our study, water gum (T. laurina) and coastal banksia (B. integrifolia) emerged as relatively high-flammability species in terms of both sustainability (median FD > 100 s) as well as the number of flaming events (median nF > 4). In a similar vein, brush box (L. confertus) was also a relatively high-flammability species for sustainability (median FD just under 80 s) and number of flaming events (median nF > 5). Interestingly, however, the native species tuckeroo (C. anacardioides) emerged as a relatively low-flammability species for sustainability (median FD just over 50 s) and number of flaming events (median nF < 3). When considered together with its low ignitibility, the observed low-flammability characteristics of these three flammability attributes suggest that tuckeroo could be targeted for further study with respect to whole-plant flammability (e.g., assessments of bark flammability, ember production) as an appropriate candidate for planting as a low-flammability tree along streets. Interestingly, tuckeroo was also highlighted as a fire-retardant plant in the list of Sands et al. (2005) [59].
Measurements of flame temperatures provide an assessment of combustibility [44]. Although we found that maximum flame temperatures reached by shoots differed significantly among species, these differences were primarily generated by one major difference between eight species with relatively high maximum flame temperatures and two species with significantly lower maximum flame temperatures (Figure 3d). Because of the slightly higher level of overlap among species in FT, it may be that FT is not as useful an attribute (compared with ignitibility for example, which had much less overlap among species) to use in assessments of street tree flammability, given that nearly all species burned with median temperatures exceeding 550 °C. As such, it becomes difficult to identify low-flammability species in terms of FT.
4.2. The Role of Shoot Traits in Shaping Shoot Flammability Patterns
Our study has identified important relationships among street tree species between shoot flammability and some of the shoot traits. We found that bulk density and leafing intensity in particular had the most explanatory power of all shoot traits that were measured, with these two traits explaining variation across the largest number of flammability attributes (Figure 5). Higher shoot bulk density was associated with longer TTF and longer FD, a result supported by the findings of previous studies [19,48]. These two flammability attributes work together antagonistically, with high bulk density (especially in high-volume shoots, Figure 5c) producing low-flammability shoots in terms of ignitibility, yet high-flammability shoots in terms of sustainability. A similar antagonistic role of the leaf trait, leaf mass per area (LMA), has been observed in previous work on leaf-level flammability [60]. In that study, high LMA was associated with slow ignitibility but long sustainability. In our study, we suggest that a larger shoot mass within a given area provides for less aeration for flames, which makes for longer ignition delay times. However, once the shoot starts to burn, flames can spread more easily to other close parts of the shoot, with the greater mass available to burn leading to longer and hotter fires [19].
We found that high leafing intensity in shoots was related to faster TTF, lower nF, and higher FT, which broadly supports the findings of [34]. However, the role of leafing intensity in driving variation in shoot flammability has generally not been explored to any great extent, even though leaf density has been correlated with patterns of other measures of flammability (e.g., [61,62,63]). We suggest that all else being equal, shoots with more leaves have more opportunity for a leaf to begin flaming, which would explain faster TTF in shoots with high leafing intensity. The lower nF observed in shoots with high leafing intensity likely emerged as a result of faster fire spread in these shoots among more leaves, leading to one large flame event instead of several small and unrelated flame commencements. Higher leafing intensity also likely leads to higher FT, with the burning of more leaves leading to hotter flame temperatures.
4.3. Native–Exotic Differences and Similarities in Shoot Flammability
Shoot flammability was found to differ between native and exotic tree species for the flammability attributes of TTF and FD (Figure 7). Exotic species ignited significantly faster than native species, while, in contrast, natives were found be more flammable than exotics for the FT attribute. This means that when deciding whether to plant exotic or native species based on their flammability, each flammability attribute should be looked at individually in response to the location being planted. At the WUI, species that ignite quickly should be avoided as this reduces their capability of acting as a green firebreak in being able to withstand radiant heat and embers [21]. Out of the species studied, bulk density was found to be higher in natives than exotics (Figure 8); and a high bulk density was found to help explain a longer TTF and FD in Figure 5, showing that the natives are more likely to have a later ignition and longer burn. Whilst exotics species were found to have a shorter FD, they were determined to have significantly faster ignition than natives. Indigenous cultural fire management aims for frequent, small scale, slow burning, and low-intensity fires [64]. The native species reflect these ideal burning characteristics in terms of the TTF and FD. However, native species were found to have a higher FT and a significantly thicker minimum stem thickness than exotics (Figure 8). A wider minimum stem thickness was also found to help explain hotter FT (Figure 5), which supports the results of natives having a higher FT than exotics.
5. Conclusions
Selection of plant species for use as street trees at the WUI should be based on a wholistic assessment of a range criteria, including the ability of the plants to be biodiversity-promoting, climate-change-resistant, health-safe, and to encompass aesthetic and functional community values [19,23]. It will also be important for future studies to explore other aspects of plant flammability, such as the flammability of bark and leaf litter of species, in addition to intrinsic flammability attributes as assessed in our study when selecting low-flammability species. In terms of the functionality of street trees in shaping opportunistic green fire-breaks of low-flammability species, we have shown that a one-size fits all approach to selecting individual trees in relation to their relative contribution to increased bushfire risk is not optimal, at least for our study species. Street tree plantings within new estate developments, for example, should consider both the interactive effects of flammability among species, with the use of ‘early igniters’ (i.e., high-flammability species in terms of ignitibility) and ‘late igniters’ (i.e., low-flammability species in terms of ignitibility) in combination, increasing the possibility of introduction of wildland fire into the urban matrix. Further research on the interactive effect of species on carrying fire is, however, required. In addition, for species commonly used at the WUI, like our ten study species, the basic assumption that exotic species are overall less flammable is not correct. Our advice is to focus on the selection of low-flammability native species, with further studies exploring which of these species are good value in terms of promoting the persistence of native biodiversity in urban landscapes.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/fire6110440/s1, File S1: Diagram of the typical set-up for the flammability testing device.
Author Contributions
Conceptualization, N.K.H.-S. and B.R.M.; writing—original draft, N.K.H.-S., B.R.M., and D.W.K.; methodology, N.K.H.-S., E.S.M., B.R.M., and D.W.K.; data collection, N.K.H.-S. and E.S.M.; data analysis, D.W.K., N.K.H.-S., and B.R.M.; writing—review and editing, N.K.H.-S., E.S.M., D.W.K., M.L.M., J.K.W., L.J.M., K.Y., C.M.M., M.C.H., and B.R.M.; supervision, B.R.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by an Australian Government Research Training Program Scholarship to N.K.H-S. and by the Local Government NSW Research and Innovation Fund, NSW, Australia to Central Coast Council.
Data Availability Statement
Data are available from the authors on request.
Acknowledgments
We thank Trees Impact for their enormous generosity and help in providing shoot samples for our experiments.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Radeloff, V.C.; Hammer, R.B.; Stewart, S.I.; Fried, J.S.; Holcomb, S.S.; McKeefry, J.F. The wildland–urban interface in the United States. Ecol. Appl. 2005, 15, 799–805. [Google Scholar] [CrossRef]
- McWethy, D.B.; Schoennagel, T.; Higuera, P.E.; Krawchuk, M.; Harvey, B.J.; Metcalf, E.C.; Schultz, C.; Miller, C.; Metcalf, A.L.; Buma, B. Rethinking resilience to wildfire. Nat. Sustain. 2019, 2, 797–804. [Google Scholar] [CrossRef]
- Haight, R.G.; Cleland, D.T.; Hammer, R.B.; Radeloff, V.C.; Rupp, T.S. Assessing fire risk in the wildland-urban interface. J. For. 2004, 102, 41–48. [Google Scholar]
- Caton, S.E.; Hakes, R.S.P.; Gorham, D.J.; Zhou, A.; Gollner, M.J. Review of pathways for building fire spread in the wildland urban interface part I: Exposure conditions. Fire Technol. 2017, 53, 429–473. [Google Scholar] [CrossRef]
- Pincetl, S.; Gillespie, T.; Pataki, D.E.; Saatchi, S.; Saphores, J.-D. Urban tree planting programs, function or fashion? Los Angeles and urban tree planting campaigns. Geo J. 2013, 78, 475–493. [Google Scholar] [CrossRef]
- Wang, Y.; Akbari, H. The effects of street tree planting on Urban Heat Island mitigation in Montreal. Sustain. Cities Soc. 2016, 27, 122–128. [Google Scholar] [CrossRef]
- Nguyen, V.D.; Roman, L.A.; Locke, D.H.; Mincey, S.K.; Sanders, J.R.; Fichman, E.S.; Duran-Mitchell, M.; Tobing, S.L. Branching out to residential lands: Missions and strategies of five tree distribution programs in the US. Urban For. Urban Green. 2017, 22, 24–35. [Google Scholar] [CrossRef]
- Gibbons, P.; van Bommel, L.; Gill, A.M.; Cary, G.J.; Driscoll, D.A.; Bradstock, R.A.; Knight, E.; Moritz, M.A.; Stephens, S.L.; Lindenmayer, D.B. Land management practices associated with house loss in wildfires. PLoS ONE 2012, 7, e29212. [Google Scholar] [CrossRef] [PubMed]
- Villaseñor, N.R.; Blanchard, W.; Lindenmayer, D.B. Decline of forest structural elements across forest–urban interfaces is stronger with high rather than low residential density. Basic Appl. Ecol. 2016, 17, 418–427. [Google Scholar] [CrossRef]
- Price, O.F.; Whittaker, J.; Gibbons, P.; Bradstock, R. Comprehensive examination of the determinants of damage to houses in two wildfires in eastern Australia in 2013. Fire 2021, 4, 44. [Google Scholar] [CrossRef]
- Willis, K.J.; Petrokofsky, G. The natural capital of city trees. Science 2017, 356, 374–376. [Google Scholar] [CrossRef]
- Turner-Skoff, J.B.; Cavender, N. The benefits of trees for livable and sustainable communities. Plants People Planet 2019, 1, 323–335. [Google Scholar] [CrossRef]
- Fadaee, S. The permaculture movement in India: A social movement with Southern characteristics. Soc. Mov. Stud. 2019, 18, 720–734. [Google Scholar] [CrossRef]
- Holmgren, D. Permaculture: Principles & Pathways beyond Sustainability; Holmgren Design Services: Hepburn, Australia, 2006. [Google Scholar]
- Bodnaruk, E.W.; Kroll, C.N.; Yang, Y.; Hirabayashi, S.; Nowak, D.J.; Endreny, T.A. Where to plant urban trees? A spatially explicit methodology to explore ecosystem service tradeoffs. Landsc. Urban Plan. 2017, 157, 457–467. [Google Scholar] [CrossRef]
- Roman, L.A.; Conway, T.M.; Eisenman, T.S.; Koeser, A.K.; Ordóñez Barona, C.; Locke, D.H.; Jenerette, G.D.; Östberg, J.; Vogt, J. Beyond ‘trees are good’: Disservices, management costs, and tradeoffs in urban forestry. Ambio 2021, 50, 615–630. [Google Scholar] [CrossRef] [PubMed]
- Kirkpatrick, J.B.; Davison, A.; Daniels, G.D. Resident attitudes towards trees influence the planting and removal of different types of trees in eastern Australian cities. Landsc. Urban Plan. 2012, 107, 147–158. [Google Scholar] [CrossRef]
- Mullaney, J.; Lucke, T.; Trueman, S.J. A review of benefits and challenges in growing street trees in paved urban environments. Landsc. Urban Plan. 2015, 134, 157–166. [Google Scholar] [CrossRef]
- Murray, B.R.; Hawthorne, T.; Curran, T.J.; Krix, D.W.; Wallace, M.I.; Young, K.; Murray, M.L.; Morley, E.; Huber-Smith, N.; Webb, J.K. Shoot flammability patterns among plant species of the wildland–urban interface in the fire-prone Greater Blue Mountains World Heritage Area. Int. J. Wildland Fire 2023, 32, 1119–1134. [Google Scholar] [CrossRef]
- Mell, W.E.; Manzello, S.L.; Maranghides, A.; Butry, D.; Rehm, R.G. The wildland–urban interface fire problem–current approaches and research needs. Int. J. Wildland Fire 2010, 19, 238–251. [Google Scholar] [CrossRef]
- Curran, T.J.; Perry, G.L.W.; Wyse, S.V.; Alam, M.A. Managing fire and biodiversity in the wildland-urban interface: A role for green firebreaks. Fire 2018, 1, 3. [Google Scholar] [CrossRef]
- Gibbons, P.; Gill, A.M.; Shore, N.; Moritz, M.A.; Dovers, S.; Cary, G.J. Options for reducing house-losses during wildfires without clearing trees and shrubs. Landsc. Urban Plan. 2018, 174, 10–17. [Google Scholar] [CrossRef]
- Murray, B.R.; Martin, L.J.; Brown, C.; Krix, D.W.; Phillips, M.L. Selecting low-flammability plants as green firebreaks within sustainable urban garden design. Fire 2018, 1, 15. [Google Scholar] [CrossRef]
- Murray, B.R.; Brown, C.; Murray, M.L.; Krix, D.W.; Martin, L.J.; Hawthorne, T.; Wallace, M.I.; Potvin, S.A.; Webb, J.K. An integrated approach to identify low-flammability plant species for green firebreaks. Fire 2020, 3, 9. [Google Scholar] [CrossRef]
- Cui, X.; Alam, M.A.; Perry, G.L.W.; Paterson, A.M.; Wyse, S.V.; Curran, T.J. Green firebreaks as a management tool for wildfires: Lessons from China. J. Environ. Manag. 2019, 233, 329–336. [Google Scholar] [CrossRef] [PubMed]
- Corbett, L. Safer Gardens: Plant Flammability and Planning for Fire; Australian Scholarly Publishing: Victoria, Australia, 2021. [Google Scholar]
- Drew-Smythe, J.J.; Davila, Y.C.; McLean, C.M.; Hingee, M.C.; Murray, M.L.; Webb, J.K.; Krix, D.W.; Murray, B.R. Community perceptions of ecosystem services and disservices linked to urban tree plantings. Urban For. Urban Green. 2023, 82, 127870. [Google Scholar] [CrossRef]
- Santacruz-García, A.C.; Bravo, S.; del Corro, F.; Ojeda, F. A comparative assessment of plant flammability through a functional approach: The case of woody species from Argentine Chaco region: A comparative assessment of plant flammability. Austral Ecol. 2019, 44, 1416–1429. [Google Scholar] [CrossRef]
- Potts, E.; Tng, D.; Apgaua, D.; Curran, T.J.; Engert, J.; Laurance, S.G.W. Growth form and functional traits influence the shoot flammability of tropical rainforest species. For. Ecol. Manag. 2022, 522, 120485. [Google Scholar] [CrossRef]
- Wyse, S.V.; Perry, G.L.W.; O’Connell, D.M.; Holland, P.S.; Wright, M.J.; Hosted, C.L.; Whitelock, S.L.; Geary, I.J.; Maurin, K.J.L.; Curran, T.J. A quantitative assessment of shoot flammability for 60 tree and shrub species supports rankings based on expert opinion. Int. J. Wildland Fire 2016, 25, 466–477. [Google Scholar] [CrossRef]
- Padullés Cubino, J.; Buckley, H.L.; Day, N.J.; Pieper, R.; Curran, T.J. Community-level flammability declines over 25 years of plant invasion in grasslands. J. Ecol. 2018, 106, 1582–1594. [Google Scholar] [CrossRef]
- Cui, X.; Paterson, A.M.; Alam, M.A.; Wyse, S.V.; Marshall, K.; Perry, G.L.W.; Curran, T.J. Shoot-level flammability across the Dracophyllum (Ericaceae) phylogeny: Evidence for flammability being an emergent property in a land with little fire. New Phytol. 2020, 228, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Burger, N.; Bond, W.J. Flammability traits of Cape shrubland species with different post-fire recruitment strategies. S. Afr. J. Bot. 2015, 101, 40–48. [Google Scholar] [CrossRef]
- Calitz, W.; Potts, A.J.; Cowling, R.M. Investigating species-level flammability across five biomes in the Eastern Cape, South Africa. S. Afr. J. Bot. 2015, 101, 32–39. [Google Scholar] [CrossRef]
- Kraaij, T.; Msweli, S.T.; Potts, A.J. Fuel trait effects on flammability of native and invasive alien shrubs in coastal fynbos and thicket (Cape Floristic Region). PeerJ 2022, 10, e13765. [Google Scholar] [CrossRef] [PubMed]
- Krix, D.W.; Murray, B.R. A Predictive Model of Leaf Flammability Using Leaf Traits and Radiant Heat Flux for Plants of Fire-Prone Dry Sclerophyll Forest. Forests 2022, 13, 152. [Google Scholar] [CrossRef]
- Central Coast Council. About Council. Available online: https://www.centralcoast.nsw.gov.au/council/about-council (accessed on 16 March 2023).
- Australian Bureau of Statistics. Greater Sydney—2021 Census All Persons Quickstats. Available online: https://abs.gov.au/census/find-census-data/quickstats/2021/1GSYD (accessed on 16 June 2023).
- Central Coast Council. Flora and Fauna Guidelines. Available online: https://cdn.centralcoast.nsw.gov.au/sites/default/files/Central_Coast_Council_Flora_and_Fauna_Guidelines_July_2019.pdf (accessed on 16 March 2023).
- Central Coast Bush Fire Management Committee. Bush Fire Risk Management Plan 2020–2025. Available online: https://ccbfmc.org/wp-content/uploads/2021/02/Central-Coast-Bushfire-Risk-Management-Plan-2020-v2-Doc-only.pdf (accessed on 16 March 2023).
- NSW Government. Final Report of the NSW Bushfire Inquiry. NSW Government—Premier and Cabinet. Available online: https://www.dpc.nsw.gov.au/assets/dpc-nsw-gov-au/publications/NSW-Bushfire-Inquiry-1630/Final-Report-of-the-NSW-Bushfire-Inquiry.pdf (accessed on 16 March 2023).
- Australian Bureau of Meteorology. Maps of Average Conditions. Available online: http://www.bom.gov.au/climate/averages/maps.shtml (accessed on 16 June 2023).
- Pena, J.C.C.; Martello, F.; Ribeiro, M.C.; Armitage, R.A.; Young, R.J.; Rodrigues, M. Street trees reduce the negative effects of urbanization on birds. PLoS ONE 2017, 12, e0174484. [Google Scholar] [CrossRef] [PubMed]
- Jaureguiberry, P.; Bertone, G.; Diaz, S. Device for the standard measurement of shoot flammability in the field. Austral Ecol. 2011, 36, 821–829. [Google Scholar] [CrossRef]
- Pérez-Harguindeguy, N.; Diaz, S.; Garnier, E.; Lavorel, S.; Poorter, H.; Jaureguiberry, P.; Bret-Harte, M.S.; Cornwell, W.K.; Craine, J.M.; Gurvich, D.E.; et al. New handbook for standardised measurement of plant functional traits worldwide. Aust. Bot. 2013, 61, 167–234. [Google Scholar] [CrossRef]
- Martin, R.E.; Gordon, D.A.; Gutierrez, M.A.; Lee, D.S.; Molina, D.M.; Schroeder, R.A.; Sapsis, D.B.; Stephens, S.L.; Chambers, M. Assessing the Flammability of Domestic and Wildland Vegetation. In Proceedings of the 12th Conference on Fire and Forest Meteorology, Jekyll Island, Georgia, 26–28 October 1993; Bethesda: Rockville, MD, USA, 1994; pp. 130–137. [Google Scholar]
- Alam, M.A.; Wyse, S.V.; Buckley, H.L.; Perry, G.L.W.; Sullivan, J.J.; Mason, N.W.H.; Buxton, R.; Richardson, S.J.; Curran, T.J. Shoot flammability is decoupled from leaf flammability, but controlled by leaf functional traits. J. Ecol. 2020, 108, 641–653. [Google Scholar] [CrossRef]
- Wyse, S.V.; Perry, G.L.W.; Curran, T.J. Shoot-Level Flammability of Species Mixtures is Driven by the Most Flammable Species: Implications for Vegetation-Fire Feedbacks Favouring Invasive Species. Ecosystems 2018, 21, 886–900. [Google Scholar] [CrossRef]
- Cui, X.; Paterson, A.M.; Perry, G.L.W.; Wyse, S.V.; Alam, M.A.; Huang, C.; Zhou, S.; Xiao, L.; Lai, C.; He, F.; et al. Intraspecific variation in shoot flammability in Dracophyllum rosmarinifolium is not predicted by habitat environmental conditions. For. Ecosyst. 2022, 9, 100017–100185. [Google Scholar] [CrossRef]
- Etlinger, M.G.; Beall, F.C. Development of a laboratory protocol for fire performance of landscape plants. Int. J. Wildland Fire 2004, 13, 479. [Google Scholar] [CrossRef]
- Fogarty, L.G. Flammability guide for some common New Zealand native tree and shrub species. In Forest Research Bulletin 197 Forest and Rural Fire Scientific and Technical Series Report 6; Forest Research Institute in association with the New Zealand Fire Service Commission and National Rural Fire Authority: Wellington, New Zealand, 2001; p. 18. [Google Scholar]
- Jir-Ming, C.; Jun-Hsien, Y. The measurement of open propane flame temperature using infrared technique. J. Quant. Spectrosc. Radiat. Transf. 1996, 56, 133–144. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. Available online: https://www.R-project.org/ (accessed on 8 June 2023).
- Fox, J.; Weisberg, S. An {R} Companion to Applied Regression, 3rd ed.; Sage Publications: Thousand Oaks, CA, USA, 2019. [Google Scholar]
- Lenth, R.V. Emmeans: Estimated Marginal Means, aka Least-Squares Means. Available online: https://CRAN.R-project.org/package=emmeans (accessed on 7 June 2023).
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Anderson, H.E. Forest fuel ignitibility. Fire Technol. 1970, 6, 312–319. [Google Scholar] [CrossRef]
- Boon, M. The Purple Reign: How the Jacaranda Became the Crown Jewel of Springtime in Sydney. Available online: https://www.timeout.com/sydney/things-to-do/the-purple-reign-how-the-jacaranda-became-the-crown-jewel-of-springtime-in-sydney (accessed on 14 June 2023).
- Sands DP, A.; Hosking, C.M. Ecologically Sustainable Fire Management: An Advisory Code for Brisbane’s Western Suburbs; Moggill Creek Catchment Group (MCCG) and Pullen Pullen Catchments Group: Brisbane, Australia, 2005. [Google Scholar]
- Krix, D.W.; Phillips, M.L.; Murray, B.R. Relationships among leaf flammability attributes and identifying low-leaf-flammability species at the wildland–urban interface. Int. J. Wildland Fire 2019, 28, 295. [Google Scholar] [CrossRef]
- Plucinski, M.P.; Anderson, W.R. Laboratory determination of factors influencing successful point ignition in the litter layer of shrubland vegetation. Int. J. Wildland Fire 2008, 17, 628–637. [Google Scholar] [CrossRef]
- Schwilk, D.W.; Caprio, A.C. Scaling from leaf traits to fire behaviour: Community composition predicts fire severity in a temperate forest. J. Ecol. 2011, 99, 970–980. [Google Scholar] [CrossRef]
- de Magalhaes, R.M.Q.; Schwilk, D.W. Leaf traits and litter flammability: Evidence for non-additive mixture effects in a temperate forest. J. Ecol. 2012, 100, 1153–1163. [Google Scholar] [CrossRef]
- McKemey, M.B.; Rangers, B.; Ens, E.J.; Hunter, J.T.; Ridges, M.; Costello, O.; Reid, N.C.H. Co-producing a fire and seasons calendar to support renewed Indigenous cultural fire management. Austral Ecol. 2021, 46, 1011–1029. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).