

Holocene Climatic Changes in the Negev Desert (Israel): Pollen and Stable Isotopes (13C/12C and 15N/14N) Records from Atzmaut and Ramon I Zoogenic Deposits


Abstract
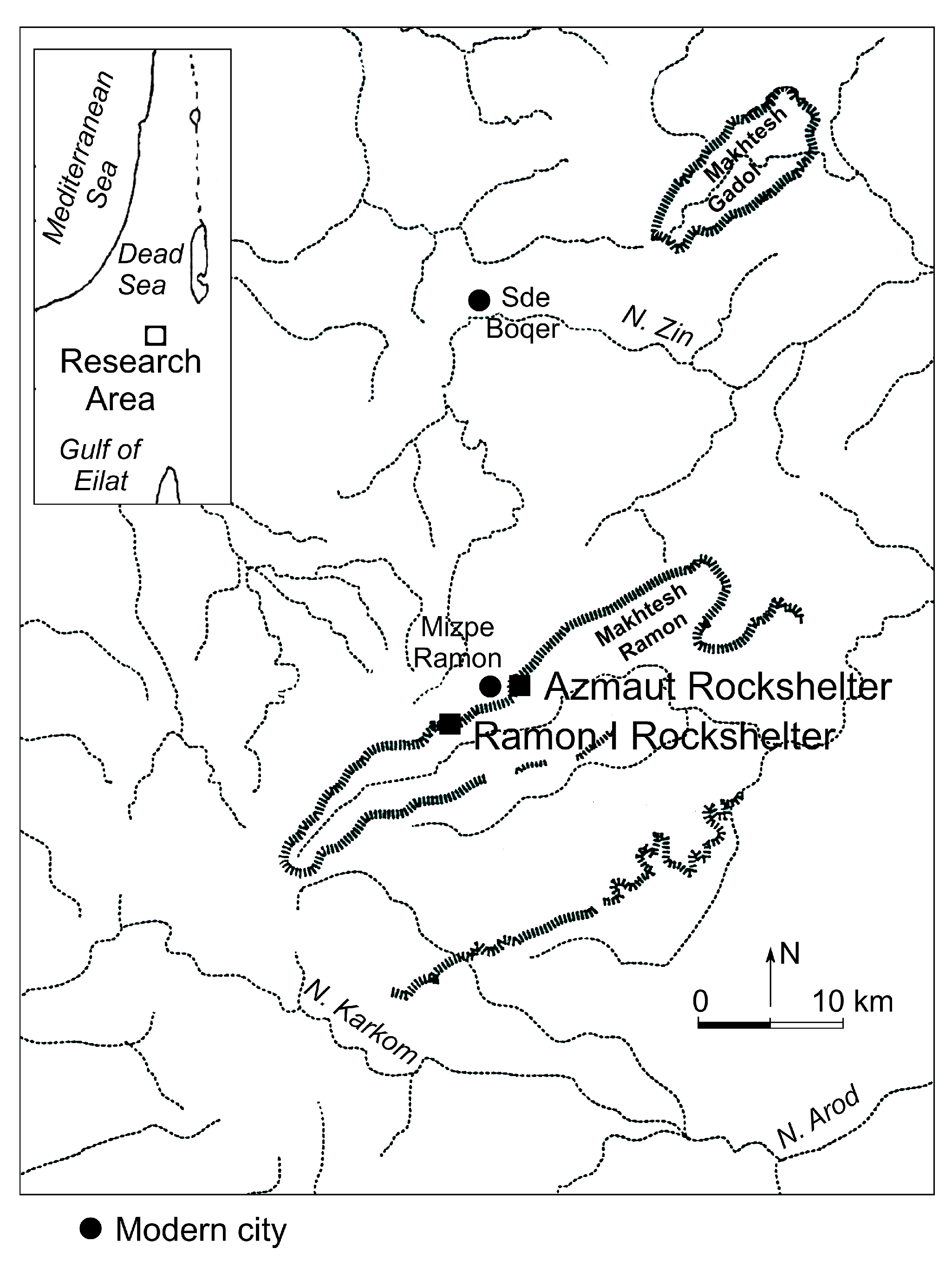
1. Introduction
2. Materials and Methods
3. Results
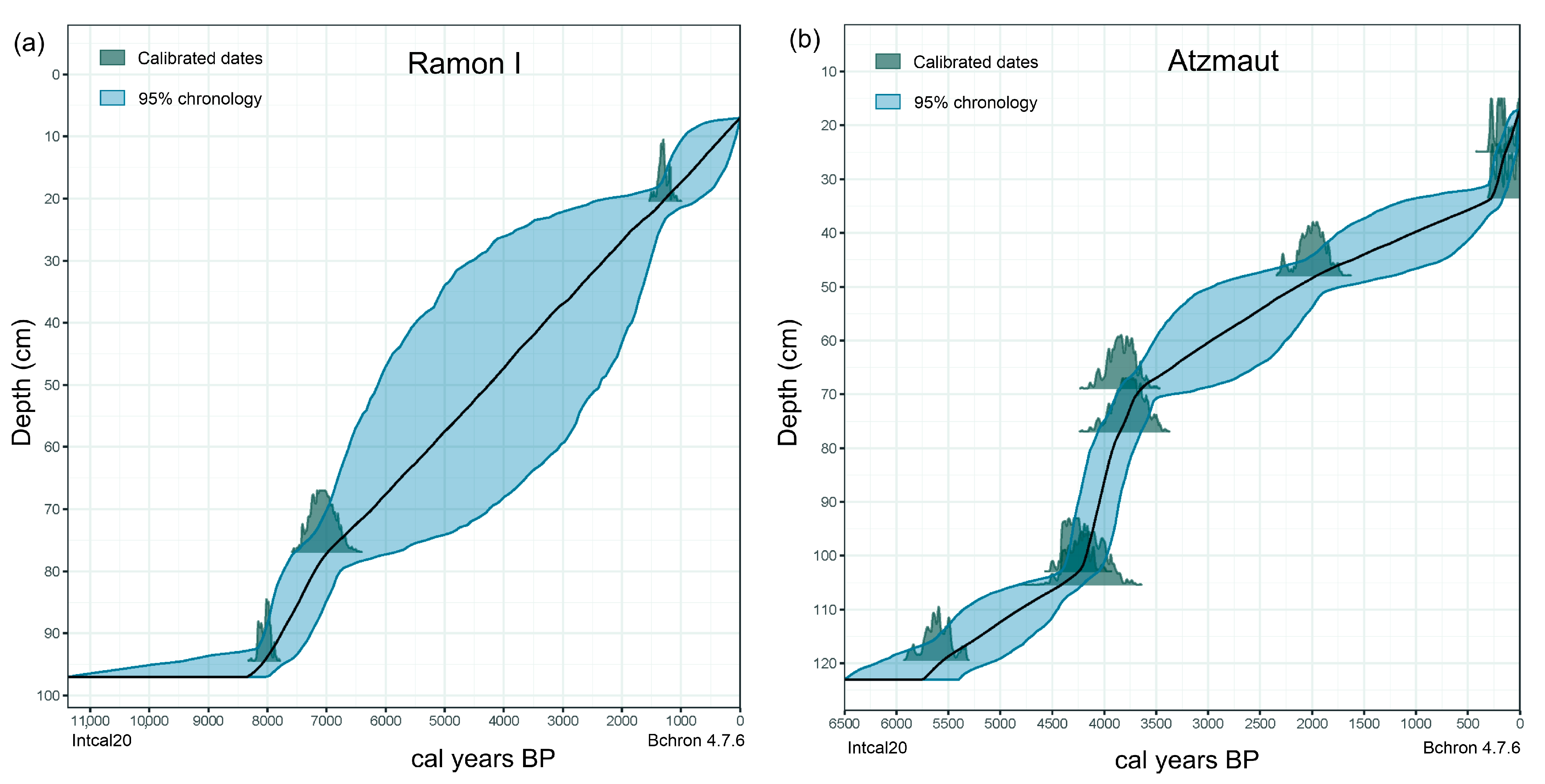
3.1. Chronology
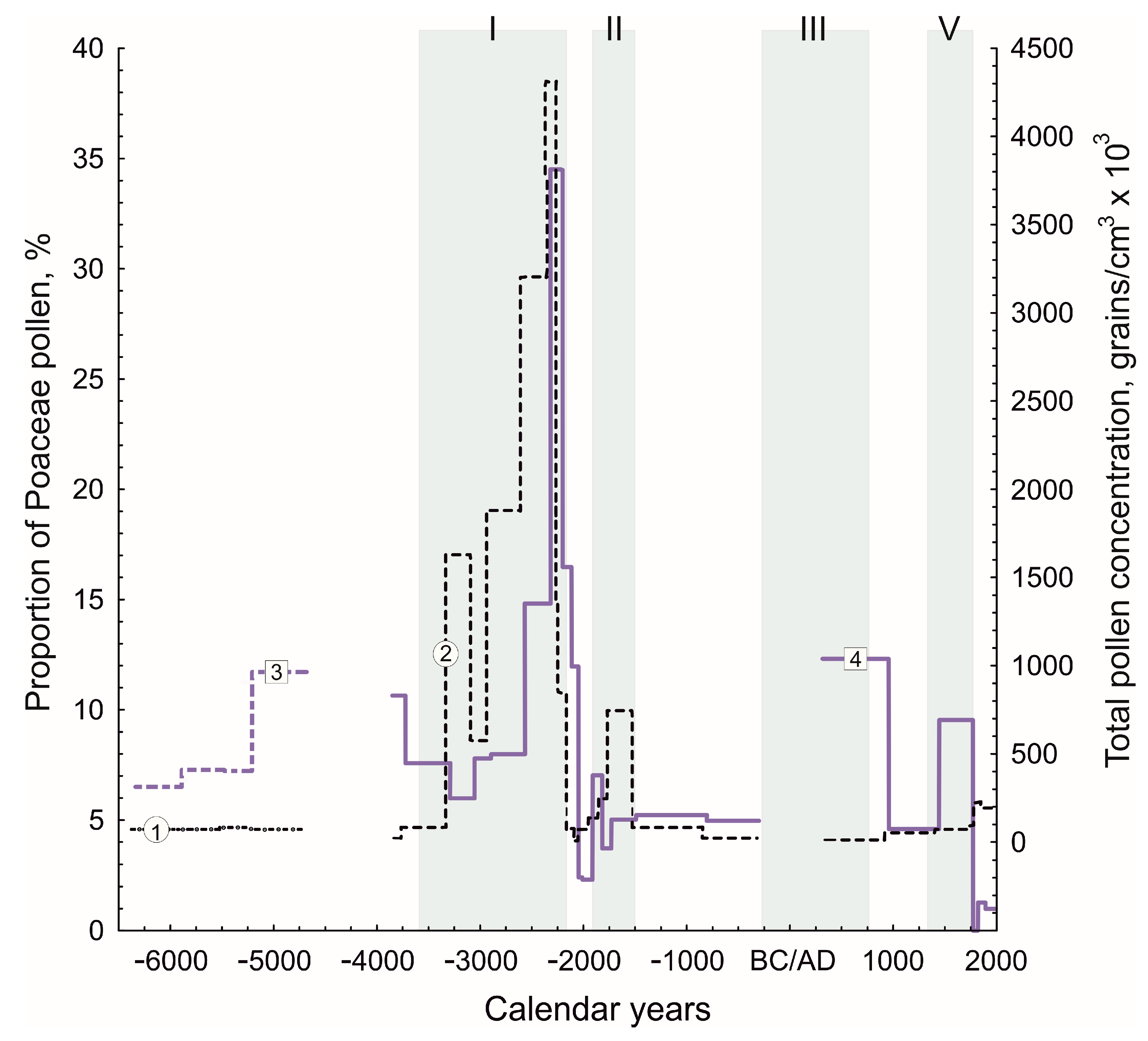
3.2. Pollen Analysis
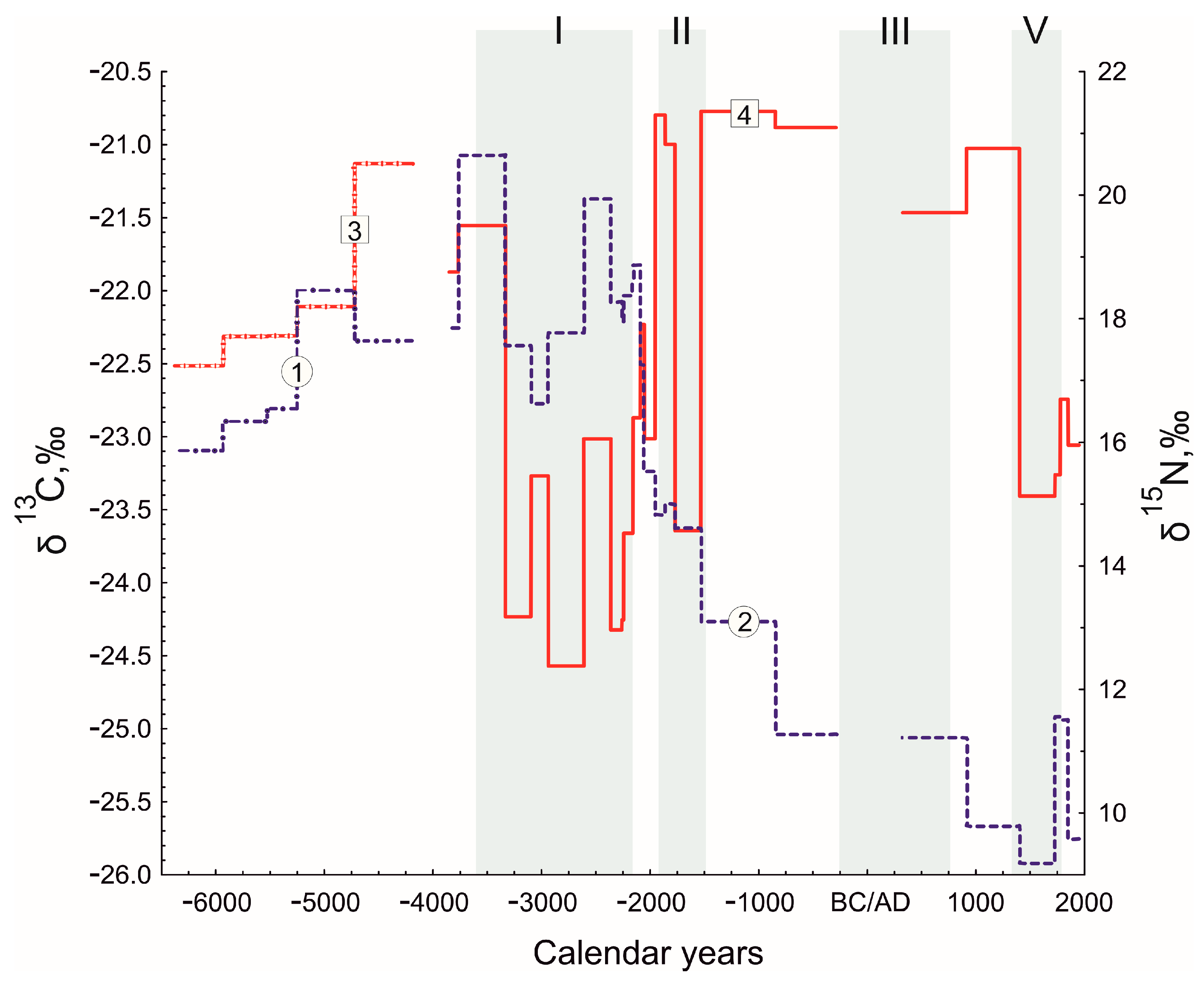
3.3. Stable Isotope Analysis
4. Discussion
5. Conclusions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pons, A.; Quézel, P. Premieres remarques sur l’etude palynologique d’un guano fossile du Hoggar. Comptes-Rendus Seances L’Academie Sci. 1958, 246, 2290–2292. [Google Scholar]
- Potter, L.D.; Rowley, J. Pollen Rain and Vegetation, San Augustin Plains, New Mexico. Bot. Gaz. 1960, 122, 1–25. [Google Scholar] [CrossRef]
- Horowitz, A. Palynology of Arid Lands; Elsevier: Amsterdam, The Netherlands, 1992; p. 546. [Google Scholar]
- Scott, L.; Vogel, J.C. Evidence for environmental conditions during the last 20 000 years in Southern Africa from 13C in fossil hyrax dung. Glob. Planet. Change 2000, 26, 207–215. [Google Scholar] [CrossRef]
- Choi, W.-J.; Lee, S.-M.; Ro, H.-M.; Kim, K.-C.; Yoo, S.-H. Natural 15N abundances of maize and soil amended with urea and composted pig manure. Plant and Soil. 2002, 245, 223–232. [Google Scholar] [CrossRef]
- Sørensen, P.; Weisbjerg, M.R.; Lund, P. Dietary effects on the composition and plant utilization of nitrogen in dairy cattlemanure. J. Agric. Sci. 2003, 141, 79–91. [Google Scholar] [CrossRef]
- Codron, D.; Codron, J.; Lee-Thorp, J.A.; Sponheimer, M.; de Ruiter, D. Animal diets in the Waterberg based on stable isotopic composition of faeces. S. Afr. J. Wildl. Res. 2005, 35, 43–52. [Google Scholar] [CrossRef]
- Kelly, J.F. Stable isotopes of carbon and nitrogen in the study of avian and mammalian trophic ecology. Can. J. Zool. 2000, 78, 1–27. [Google Scholar] [CrossRef]
- Bocherens, H.; Drucker, D. Trophic level isotopic enrichment of carbon and nitrogen in bone collagen: Case studies from recent and ancient terrestrial ecosystems. Int. J. Osteoarchaeol. 2003, 13, 46–53. [Google Scholar] [CrossRef]
- O`Leary, M.H. Carbon isotopes in photosynthesis. BioScience 1988, 38, 328–336. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Ehleringer, J.R.; Hubick, K.T. Carbon isotope discriminationand photosynthesis. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1989, 40, 503–537. [Google Scholar] [CrossRef]
- Cerling, T.E.; Harris, J.M.; MacFadden, B.J.; Leakey, M.G.; Quade, J.; Eisenmann, V.; Ehleringer, J.R. Global vegetation change through the Miocene/Pliocene boundary. Nature 1997, 389, 153–158. [Google Scholar] [CrossRef]
- Kohn, M.J. Carbon isotope compositions of terrestrial C3 plants as indicators of (paleo)ecology and (paleo)climate. Proc. Natl. Acad. Sci. USA 2010, 107, 19691–19695. [Google Scholar] [CrossRef] [PubMed]
- Cernusak, L.A.; Ubierna, N.; Winter, K.; Holtum, J.A.M.; Marshall, J.D.; Farquhar, G.D. Environmental and physiological determinants of carbon isotope discrimination in terrestrial plants. New Phytol. 2013, 200, 950–965. [Google Scholar] [CrossRef] [PubMed]
- Marshall, J.D.; Brooks, J.R.; Lajtha, K. Sources of variation in the stable isotopic composition of plants. In Stable Isotopes in Ecology and Environmental Science, 2nd ed.; Michener, R., Lajtha, K., Eds.; Blackwell Publishing Ltd.: Malden, MA, USA; Oxford, UK; Carlton, Australia, 2007; pp. 22–60. [Google Scholar]
- Chase, B.M.; Quick, L.J.; Meadows, M.E.; Scott, L.; Thomas, D.S.G.; Reimer, P.J. Late glacial interhemispheric climate dynamics revealed in South African hyrax middens. Geology 2011, 39, 19–22. [Google Scholar] [CrossRef]
- Chase, B.M.; Boom, A.; Carr, A.S.; Carré, M.; Chevalier, M.; Meadows, M.E.; Pedro, J.B.; Stager, J.C.; Reimer, P.J. Evolving southwest African response to abrupt deglacial North Atlantic climate change events. Quat. Sci. Rev. 2015, 121, 132–136. [Google Scholar] [CrossRef]
- Shahack-Gross, R.; Simons, A.; Ambrose, S.H. Identification of pastoral sites using stable nitrogen and carbon isotopes from bulk sediment samples: A case study in modern and archaeological pastoral settlements in Kenya. J. Archaeol. Sci. 2008, 35, 983–990. [Google Scholar] [CrossRef]
- Carr, A.S.; Chase, B.M.; Boom, A.; Medina-Sanchez, J. Stable isotope analyses of rock hyrax faecal pellets, hyraceum and associated vegetation in southern Africa: Implications for dietary ecology and palaeoenvironmental reconstructions. J. Arid. Environ. 2016, 134, 33–48. [Google Scholar] [CrossRef]
- Chase, B.M.; Meadows, M.E.; Carr, A.S.; Reimer, P.J. Evidence for progressive Holocene aridification in southern Africa recorded in Namibian hyrax middens: Implications for African Monsoon dynamics and the “African Humid Period”. Quat. Res. 2010, 74, 36–45. [Google Scholar] [CrossRef]
- Chase, B.M.; Scott, L.; Meadows, M.E.; Gil-Romera, G.; Boom, A.; Carr, A.S.; Reimer, P.J.; Truc, L.; Valsecchi, V.; Quick, L.J. Rock hyrax middens: A palaeoenvironmental archive for southern African drylands. Quat. Sci. Rev. 2012, 56, 107–125. [Google Scholar] [CrossRef]
- Chase, B.M.; Niedermeyer, E.M.; Boom, A.; Carr, A.S.; Chevalier, M.; He, F.; Meadows, M.E.; Ogle, N.; Reimer, P.J. Orbital controls on Namib Desert hydroclimate over the past 50,000 years. Geology 2019, 47, 867–871. [Google Scholar] [CrossRef]
- Chase, B.M.; Boom, A.; Carr, A.S.; Quick, L.J.; Reimer, P.J. High-resolution record of Holocene climate change dynamics from southern Africa’s temperate-tropical boundary, Baviaanskloof, South Africa. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2020, 539, 109518. [Google Scholar] [CrossRef]
- Chase, B.M.; Boom, A.; Carr, A.S.; Reimer, P.J. Climate variability along the margin of the southern African monsoon region at the end of the African Humid Period. Quat. Sci. Rev. 2022, 291, 107663. [Google Scholar] [CrossRef]
- Chase, B.M.; Boom, A.; Carr, A.S.; Meadows, M.E.; Lima, S. A ca. 39,000-year record of vegetation and climate change from the margin of the Namib Sand Sea. Quat. Res. 2023, 116, 1–11. [Google Scholar] [CrossRef]
- Horisk, K.; Ivory, S.; McCorriston, J.; McHale, M.; Al Mehri, A.; Anderson, A.; Scott, R.A.; Al Kathiri, A. Vegetation dynamics in Dhofar, Oman, from the Late Holocene to present inferred from rock hyrax middens. Quat. Res. 2023, 116, 12–29. [Google Scholar] [CrossRef]
- Ivory, S.J.; Cole, K.L.; Anderson, R.S.; Anderson, A.; McCorriston, J.; Williams, J. Human landscape modification and expansion of tropical woodland in southern Arabia during the mid-Holocene from rock hyrax middens. J. Biogeogr. 2021, 48, 2588–2603. [Google Scholar] [CrossRef]
- Meadows, M.E.; Seliane, M.; Chase, B.M. Holocene palaeoenvironments of the Cederberg and Swartruggens mountains, Western Cape, South Africa: Pollen and stable isotope evidence from hyrax dung middens. J. Arid. Environ. 2010, 74, 786–793. [Google Scholar] [CrossRef]
- Handley, L.L.; Austin, A.T.; Robinson, D.; Scrimgeour, C.M.; Raven, J.A.; Heaton, T.H.E.; Schmidt, S.; Stewart, G.R. The 15N natural abundance (δ15N) of ecosystem samples reflects measures of water availability. Australian J. Plant Physiol. 1999, 26, 185–199. [Google Scholar] [CrossRef]
- Robinson, D. δ15N as an integrator of the nitrogen cycle. Trends Ecol. Evol. 2001, 16, 153–162. [Google Scholar] [CrossRef]
- Amundson, R.; Austin, A.T.; Schuur, E.A.G.; Yoo, K.; Matzek, V.; Kendall, C.; Uebersax, A.; Brenner, D.; Baisden, W.T. Global patterns of the isotopic composition of soil and plant nitrogen. Glob. Biogeochem. Cycles 2003, 17, 1031. [Google Scholar] [CrossRef]
- Craine, J.M.; Elmore, A.J.; Aidar, M.P.M.; Bustamante, M.; Dawson, T.E.; Hobbie, E.A.; Kahmen, A.; Mack, M.C.; McLauchlan, K.K.; Michelsen, A.; et al. Global patterns of foliar nitrogen isotopes and their relationships with climate, mycorrhizal fungi, foliar nutrient concentrations, and nitrogen availability. New Phytol. 2009, 183, 980–992. [Google Scholar] [CrossRef]
- Heaton, T.H.E. The 15N/14N ratios of plants in South Africa and Namibia: Relationship to climate and coastal/saline environments. Oecologia 1987, 74, 236–246. [Google Scholar] [CrossRef] [PubMed]
- Swap, R.J.; Aranibar, J.N.; Dowty, P.R.; Gilhooly, W.P.; Macko, S.A. Natural abundance of 13C and 15N in C3 and C4 vegetation of southern Africa: Patterns and implications. Glob. Change Biol. 2004, 10, 350–358. [Google Scholar] [CrossRef]
- Aranibar, J.N.; Otter, L.; Macko, S.A.; Feral, C.J.W.; Epstein, H.E.; Dowty, P.R.; Eckardt, F.; Shugart, H.H.; Swap, R.J. Nitrogen cycling in the soil–plant system along a precipitation gradient in the Kalahari sands. Glob. Change Biol. 2004, 10, 359–373. [Google Scholar] [CrossRef]
- Murphy, B.P.; Bowman, D.M.J.S. The carbon and nitrogen isotope composition of Australian grasses in relation to climate. Funct. Ecol. 2009, 23, 1040–1049. [Google Scholar] [CrossRef]
- Hartman, G.; Danin, A. Isotopic values of plants in relation to water availability in the Eastern Mediterranean region. Oecologia 2010, 162, 837–852. [Google Scholar] [CrossRef]
- Hartman, G. Are elevated δ15N values in herbivores in hot and arid environments caused by diet or animal physiology? Funct. Ecol. 2011, 25, 122–131. [Google Scholar] [CrossRef]
- Babenko, A.N.; Khassanov, B.F. The absolute chronology of the zoogenic deposits from the Negev Desert (Israel). Geochronometria 2007, 28, 47–53. [Google Scholar] [CrossRef]
- Kahana, R.; Baruch, Z.; Enzel, Y.; Dayan, U. Synoptic climatology of major floods in the Negev desert, Israel. Int. J. Climatol. 2002, 22, 867–882. [Google Scholar] [CrossRef]
- Danin, A. Desert Vegetation of Israel and Sinai; Cana Publishing House: Jerusalem, Israel, 1983; p. 133. [Google Scholar]
- Frumkin, A. The Holocene History of Dead Sea Levels. In The Dead Sea: The Lake and Its Setting; Ben-Avraham, Z., Gat, Y., Niemi, T.M., Eds.; Oxford University Press: Oxford, MS, USA, 1997; pp. 237–248. [Google Scholar]
- Rossignol-Strick, M. The Holocene climatic optimum and pollen records of sapropel 1 in the eastern Mediterranean, 9000–6000 BP. Quat. Sci. Rev. 1999, 18, 515–530. [Google Scholar] [CrossRef]
- Frumkin, A.; Magaritz, M.; Carmi, I.; Zak, I. The Holocene climatic record of the salt caves of Mount Sedom, Israel. Holocene 1991, 1, 191–200. [Google Scholar] [CrossRef]
- Frumkin, A.; Carmi, I.; Zak, I.; Magaritz, M. Middle Holocene environmental change determined from the Salt Caves of Mount Sodom, Israel. In Late Quaternary Chronology and Paleoclimates of the Eastern Mediterranean; Bar-Yosef, O., Kra, R.S., Eds.; RADIOCARBON: Tucson, AZ, USA; American School of Prehistoric Research: Cambridge, MA, USA, 1994; pp. 315–332. [Google Scholar]
- Frumkin, A.; Kadan, G.; Enzel, Y.; Eyal, Y. Radiocarbon Chronology of the Holocene Dead Sea: Attempting a regional Correlation. Radiocarbon 2001, 43, 1179–1189. [Google Scholar] [CrossRef]
- Frumkin, A.; Elitzur, Y. Historic Dead Sea Level Fluctuations Calibrated with Geological and Archaeological Evidence. Quat. Res. 2002, 57, 334–342. [Google Scholar] [CrossRef]
- Bookman (Ken-Tor), R.; Enzel, Y.; Agnon, A.; Stein, M. Late Holocene lake levels of the Dead Sea. GSA Bull. 2004, 116, 555–571. [Google Scholar] [CrossRef]
- Hazan, N.; Stein, M.; Agnon, A.; Marco, S.; Nadel, D.; Negendank, J.F.W.; Schwab, M.J.; Neev, D. The late Quaternary limnological history of Lake Kinneret (Sea of Galilee), Israel. Quat. Res. 2005, 63, 60–77. [Google Scholar] [CrossRef]
- Arz, H.W.; Lamy, F.; Patzold, J.; Muller, P.J.; Prins, M. Mediterranean Moisture Source for an Early-Holocene Humid Period in the Northern Red Sea. Science 2003, 300, 118–121. [Google Scholar] [CrossRef]
- Arz, H.W.; Lamy, F.; Patzold, J. A pronounced dry event recorded around 4.2 ka in brine sediments from the northern Red Sea. Quat. Res. 2006, 66, 432–441. [Google Scholar] [CrossRef]
- Migowski, C.; Stein, M.; Prasad, S.; Negendank, J.F.W.; Agnon, A. Holocene climate variability and cultural evolution in the Near East from the Dead Sea sedimentary record. Quat. Res. 2006, 66, 421–431. [Google Scholar] [CrossRef]
- Gvirtzman, G.; Wieder, M. Climate of the last 53,000 years in the eastern Mediterranian, based on the soil-sequence stratigraphy in the coastal plain of Israel. Quat. Sci. Rev. 2001, 20, 1827–1849. [Google Scholar] [CrossRef]
- Bar-Matthews, M.; Ayalon, A.; Kaufman, A. Middle to Late Holocene (6500 Yr. period) paleoclimate in the Eastern Mediterranian region from stable isotopic composition of speleothems from Soreq Cave, Israel. In Water, Environment and Society in Times of Climatic Change; Issar, A.S., Brown, N., Eds.; Water Science and Technology Library; Springer: Dordrecht, The Netherlands, 1998; Volume 31, pp. 203–214. [Google Scholar] [CrossRef]
- Orland, I.J.; Bar-Matthews, M.; Kita, N.T.; Ayalon, A.; Matthews, A.; Valley, J.W. Climate deterioration in the Eastern Mediterranean as revealed by ion microprobe analysis of a speleothem that grew from 2.2 to 0.9 ka in Soreq Cave, Israel. Quat. Res. 2009, 71, 27–35. [Google Scholar] [CrossRef]
- Bar-Matthews, M.; Ayalon, A. Mid-Holocene climate variations revealed by high-resolution speleothem records from Soreq Cave, Israel and their correlation with cultural changes. Holocene 2011, 21, 163–171. [Google Scholar] [CrossRef]
- Goodfriend, G.A. Terrestrial stable isotope records of Late Quaternary paleoclimates in the eastern Mediterranean region. Quat. Sci. Rev. 1999, 18, 501–513. [Google Scholar] [CrossRef]
- Moustafa, Y.A.; Patzold, J.; Loya, Y.; Wefer, G. Mid-Holocene stable isotope record of corals from the northern Red Sea. Int. J. Earth Sci. 2000, 88, 742–751. [Google Scholar] [CrossRef]
- Lisker, S.; Porat, R.; Davidovich, U.; Eshel, H.; Lauritzen, S.E.; Frumkin, A. Late Quaternary environmental and human events at En Gedi, reflected by the geology and archaeology of the Moringa Cave (Dead Sea area, Israel). Quat. Res. 2007, 68, 203–212. [Google Scholar] [CrossRef]
- Yakir, D.; Issar, A.; Gat, J.; Adar, E.; Trimborn, P.; Lipp, J. 13C and 18O of wood from the Roman siege rampart in Masada, Israel (AD 70-73): Evidence for a less arid climate for the region. Geochim. Cosmochim. Acta 1994, 58, 3535–3539. [Google Scholar] [CrossRef]
- Frumkin, A. Stable isotopes of a subfossil Tamarix tree from the Dead Sea region, Israel, and their implications for the Intermediate Bronze Age environmental crisis. Quat. Res. 2009, 71, 319–328. [Google Scholar] [CrossRef]
- Waisel, Y.; Liphschitz, N. Dendrochronological investigations in Israel: II Juniperus phoenica of West and Central Sinai. La-Yarran 1968, 18, 63–67. [Google Scholar]
- Rosen, S.A.; Savinetsky, A.B.; Plakht, Y.; Kisseleva, N.K.; Khassanov, B.F.; Pereladov, A.M.; Haiman, M. Dung in the Desert: Preliminary Results of the Negev Holocene Ecology Project. Curr. Anthropol. 2005, 46, 317–327. [Google Scholar] [CrossRef]
- Simms, S.R.; Russell, K.W. Tur Imdai Rockshelter: Archaeology of Recent Pastoralists in Jordan. J. Field Archaeol. 1997, 24, 459–472. [Google Scholar] [CrossRef]
- Haslett, J.; Parnell, A. A simple monotone process with application to radiocarbon-dated depth chronologies. J. R. Stat. Soc. Ser. C (Appl. Stat.) 2008, 57, 399–418. [Google Scholar] [CrossRef]
- Reimer, P.; Austin, W.; Bard, E.; Bayliss, A.; Blackwell, P.; Ramsey Bronk, C.; Butzin, M.; Cheng, H.; Edwards, R.L.; Friedrich, M.; et al. The IntCal20 Northern Hemisphere Radiocarbon Age Calibration Curve (0–55 cal kBP). Radiocarbon 2020, 62, 725–757. [Google Scholar] [CrossRef]
- Parnell, A.C.; Buck, C.E.; Doan, T.K. A review of statistical chronology models for high-resolution, proxy-based Holocene palaeoenvironmental reconstruction. Quat. Sci. Rev. 2011, 30, 2948–2960. [Google Scholar] [CrossRef]
- Faegri, K.; Iversen, J. Textbook of Pollen Analysis, 4th ed.; John Wiley & Sons: Chichester, UK, 1989; p. 328. [Google Scholar]
- Stockmarr, J. Determination of spore concentration with an electronic particle counter. Dan. Geol. Undersøgelse Årbog 1973, 1972, 87–89. [Google Scholar]
- Grimm, E.C. Computer Software, TILIA 2.6.1 Version; State Museum: Springfield, IL, USA, 2019. [Google Scholar]
- Zar, J.H. Biostatistical Analysis, 5th ed.; Pearson: London, UK, 2010; p. 960. [Google Scholar]
- Brooks, J.; Shaw, G. Geochemistry of sporopollenin. Chem. Geol. 1972, 10, 69–87. [Google Scholar] [CrossRef]
- Babenko, A.N.; Kiseleva, N.K.; Plakht, I.; Rosen, S.; Savinetskii, A.B.; Khasanov, B.F. Reconstruction of the Holocene Vegetation in the Central Negev Desert, Israel, on the Basis of Palynological Data on the Atzmaut Zoogenic Deposit. Russ. J. Ecol. 2007, 38, 388–397. [Google Scholar] [CrossRef]
- Babenko, A.N.; Kuzmicheva, E.A.; Khasanov, B.F.; Rosen, S.; Kiseleva, N.K.; Savinetsky, A.B. Dung deposits as archives of environmental change. In Soils and Sediments as Archives of Environmental Change; Lucke, B., Bäumler, R., Schmidt, M., Eds.; Geoarchaeology and Landscape Change in the Subtropics and Tropics; Chapter 15; Fränkische Geographische Gesellschaft: Erlanger, German, 2015; Volume 42, pp. 201–217. [Google Scholar]
- Moe, D. Palynology of sheep’s faeces: Relationship beetween pollen content, diet and local pollen rain. Grana 1983, 22, 105–113. [Google Scholar] [CrossRef]
- Akeret, O.; Haas, N.J.; Leuzinger, U.; Jacomet, S. Plant macrofossils and pollen in goat/sheep faeces from the Neolithic lake-shore settlement Arbon Bleiche 3, Switzerland. Holocene 1999, 9, 175–182. [Google Scholar] [CrossRef]
- Hunt, C.O.; Rushworth, G.; Gilbertson, D.D.; Mattingly, D.J. Romano-Libyan Dryland Animal Husbandry and Landscape: Pollen and Palynofacies Analyses of Coprolites from a Farm in the Wadi el-Amud, Tripolitania. J. Archaeol. Sci. 2001, 28, 351–363. [Google Scholar] [CrossRef]
- Feinbrun-Dothan, N. Flora Palaestina; Part III; Israel Academy of Sciences and Humanities: Jerusalem, Israel, 1978; p. 481. [Google Scholar]
- Feinbrun-Dothan, N. Flora Palaestina; Part IV; Israel Academy of Sciences and Humanities: Jerusalem, Israel, 1986; p. 463. [Google Scholar]
- Zohary, M. Flora Palaestina; Part 1; Israel Academy of Sciences and Humanities: Jerusalem, Israel, 1981; p. 367. [Google Scholar]
- El-Moslimany, A.P. Ecological significance of common nonarboreal pollen: Examples from drylands of the Middle East. Rev. Palaeobot. Palynol. 1990, 64, 343–350. [Google Scholar] [CrossRef]
- Finne, M.; Holmgren, K.; Sundqvis, H.S.; Weiberg, E.; Lindblom, M. Climate in the eastern Mediterranean, and adjacent regions, during the past 6000 years—A review. J. Archaeol. Sci. 2011, 38, 3153–3173. [Google Scholar] [CrossRef]
- Harrison, S.P.; Yu, G.; Tarasov, P.E. Late Quaternary Lake-Level Record from Northern Eurasia. Quat. Res. 1996, 45, 138–159. [Google Scholar] [CrossRef]
- Staubwasser, M.; Weiss, H. Holocene climate and cultural evolution in late prehistoric-early historic West Asia. Quat. Res. 2006, 66, 372–387. [Google Scholar] [CrossRef]
- Weninger, B.; Alram-Stern, E.; Bauer, E.; Clare, L.; Danzeglocke, U.; Joris, O.; Kubatzki, C.; Rollefson, G.; Todorova, H.; Van Andel, T. Climate forcing due to the 8200 cal yr BP event observed at Early Neolithic sites in the eastern Mediterranean. Quat. Res. 2006, 66, 401–420. [Google Scholar] [CrossRef]
- Verheyden, S.; Nader, F.H.; Cheng, H.J.; Edwards, L.R.; Swennen, R. Paleoclimate reconstruction in the Levant region from the geochemistry of a Holocene stalagmite from the Jeita cave, Lebanon. Quat. Res. 2008, 70, 368–381. [Google Scholar] [CrossRef]
- Gat, J.; Magaritz, M. Climatic variations in the eastern Mediterranean Sea area. Naturwissenschaften 1980, 67, 80–87. [Google Scholar] [CrossRef]
- Roberts, N.; Moreno, A.; Valero-Garces, B.L.; Corella, J.P.; Jones, M.; Allcock, S.; Woodbridge, J.; Morellon, M.; Luterbache, J.; Xoplaki, E.; et al. Palaeolimnological evidence for an east–west climate see-saw in the Mediterranean since AD 900. Glob. Planet. Chang. 2012, 84–85, 23–34. [Google Scholar] [CrossRef]
- Connin, S.L.; Feng, X.; Virginia, R.A. Isotopic discrimination during long-term decomposition in an arid land ecosystem. Soil Bioogy Biochem. 2001, 33, 41–51. [Google Scholar] [CrossRef]
- Tiessen, H.; Stewart, J.W.B.; Hunt, H.W. Concepts of soil organic matter transformations in relation to organo-mineral particle size fractions. Plant Soil 1984, 76, 287–295. [Google Scholar] [CrossRef]
- Huygens, D.; Denef, K.; Vandeweyer, R.; Godoy, R.; Van Cleemput, O.; Boeckx, P. Do nitrogen isotope patterns reflect microbial colonization of soil organic matter fractions? Biol. Fertil. Soils 2008, 44, 955–964. [Google Scholar] [CrossRef]
- Tsoar, H. Desertification in Northern Sinai in the eighteenth century. Clim. Chang. 1995, 29, 429–438. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lab No. | Depth (cm) | 14C Age (BP) | Cal. Ages, 1σ | Dated Material |
|---|---|---|---|---|
| Atzmaut | ||||
| IEMAE-1331 | 17–20 | 101.9 ± 0.9% | 1955–2018 AD | Plant remains |
| IEMAE-1330 | 20–23 | 100.5 ± 0.9% | 1690–… AD | Plant remains |
| IEMAE-1329 | 23–27 | 180 ± 40 | 1660–… AD | Plant remains |
| IEMAE-1348 | 31–36 | 155 ± 40 | 1670–1945 AD | Plant remains |
| IEMAE-1342 | 45–51 | 2040 ± 90 | 170 BC–75 AD | Plant remains |
| IEMAE-1328 | 67–71 | 3545 ± 90 | 2020–1750 BC | Plant remains |
| IEMAE-1327 | 75–79 | 3490 ± 110 | 1945–1645 BC | Plant remains |
| IEMAE-1326 | 102–104 | 3865 ± 70 | 2460–2210 BC | Plant remains |
| IEMAE-1325 | 104–107 | 3780 ± 120 | 2430–2030 BC | Plant remains |
| IEMAE-1298 | 116–123 | 4870 ± 100 | 3770–3530 BC | Humic acid |
| Ramon I | ||||
| IEMAE-1402 | 7–9 | 114.95 ± 2.35% | 1960–1990 AD | Plant remains |
| IEMAE-1352 | 18–23 | 1400 ± 90 | 560–770 AD | Plant remains |
| IEMAE-1401 | 74–80 | 6190 ± 180 | 5320–4905 BC | Plant remains |
| IEMAE-1358 | 92–97 | 7210 ± 80 | 6220–5990 BC | Plant remains |
| Depth (cm) | Cal. Ages, BC/AD * | δ13C, ‰ | δ15N (<0.25 mm Fraction), ‰ | δ15N (>0.25 mm Fraction), ‰ | δ15N (Mean Values), ‰ |
|---|---|---|---|---|---|
| Atzmaut | |||||
| 17–20 | 1950–1900 AD | −22.45 | 9.09 | 8.74 | 8.92 |
| 20–23 | 1900–1850 AD | −23.06 | 10.09 | 9.07 | 9.58 |
| 23–27 | 1850–1780 AD | −22.74 | 11.62 | 11.39 | 11.51 |
| 27–31 | 1780–1730 AD | −23.26 | 11.6 | 11.52 | 11.56 |
| 31–36 | 1730–1400 AD | −23.40 | 9.23 | 9.15 | 9.19 |
| 36–40 | 1400–915 AD | −21.03 | 9.49 | 10.1 | 9.8 |
| 40–45 | 915–330 AD | −21.47 | 10.94 | 11.49 | 11.23 |
| 51–58 | 290–850 BC | −20.88 | 11.9 | 10.65 | 11.28 |
| 58–67 | 850–1530 BC | −20.77 | 11.73 | 15.47 | 13.6 |
| 67–71 | 1530–1770 BC | −23.64 | 14.81 | 14.4 | 14.61 |
| 71–75 | 1770–1860 BC | −20.10 | 15.23 | 14.76 | 15 |
| 75–79 | 1860–1950 BC | −20.80 | 15.34 | 14.33 | 14.83 |
| 79–86 | 1950–2050 BC | −23.01 | 16.02 | 15.02 | 15.52 |
| 86–89 | 2050–2090 BC | −22.23 | 17.85 | 16.67 | 17.26 |
| 89–95 | 2090–2150 BC | −22.87 | 19.11 | 18.61 | 18.86 |
| 95–101 | 2150–2240 BC | −23.66 | 17.54 | 19.2 | 18.37 |
| 101–102 | 2240–2260 BC | −24.25 | 17.85 | 18.05 | 17.95 |
| 102–104 | 2260–2360 BC | −24.32 | 18.23 | 18.33 | 18.28 |
| 104–107 | 2360–2610 BC | −23.02 | 20.1 | 19.8 | 19.95 |
| 107–111 | 2610–2935 BC | −24.57 | 17.91 | 17.63 | 17.77 |
| 111–113 | 2935–3095 BC | −23.27 | 17.45 | 15.79 | 16.62 |
| 113–116 | 3095–3330 BC | −24.23 | 17.37 | 17.75 | 17.56 |
| 116–123 | 3330–3765 BC | −21.55 | 21.02 | 20.29 | 20.65 |
| 123–125 | 3765–3850 BC | −21.87 | 17.86 | - | - |
| Ramon I | |||||
| 69–74 | 4180–4720 BC | −21.13 | 17.65 | - | - |
| 74–80 | 4720–5250 BC | −22.11 | 18.46 | - | - |
| 80–85 | 5250–5520 BC | −22.31 | 16.54 | - | - |
| 85–92 | 5520–5930 BC | −22.31 | 16.34 | - | - |
| 92–97 | 5930–6380 BC | −22.52 | 15.87 | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Babenko, A. Holocene Climatic Changes in the Negev Desert (Israel): Pollen and Stable Isotopes (13C/12C and 15N/14N) Records from Atzmaut and Ramon I Zoogenic Deposits. Quaternary 2024, 7, 45. https://doi.org/10.3390/quat7040045
Babenko A. Holocene Climatic Changes in the Negev Desert (Israel): Pollen and Stable Isotopes (13C/12C and 15N/14N) Records from Atzmaut and Ramon I Zoogenic Deposits. Quaternary. 2024; 7(4):45. https://doi.org/10.3390/quat7040045
Chicago/Turabian StyleBabenko, Anna. 2024. "Holocene Climatic Changes in the Negev Desert (Israel): Pollen and Stable Isotopes (13C/12C and 15N/14N) Records from Atzmaut and Ramon I Zoogenic Deposits" Quaternary 7, no. 4: 45. https://doi.org/10.3390/quat7040045
APA StyleBabenko, A. (2024). Holocene Climatic Changes in the Negev Desert (Israel): Pollen and Stable Isotopes (13C/12C and 15N/14N) Records from Atzmaut and Ramon I Zoogenic Deposits. Quaternary, 7(4), 45. https://doi.org/10.3390/quat7040045