
New Diatom and Sedimentary Data Confirm the Existence of the Northern Paleo-Outlet from Lake Ladoga to the Baltic Sea
 , ,
, ,
Abstract
1. Introduction
Methodological Background
2. Materials and Methods
3. Results
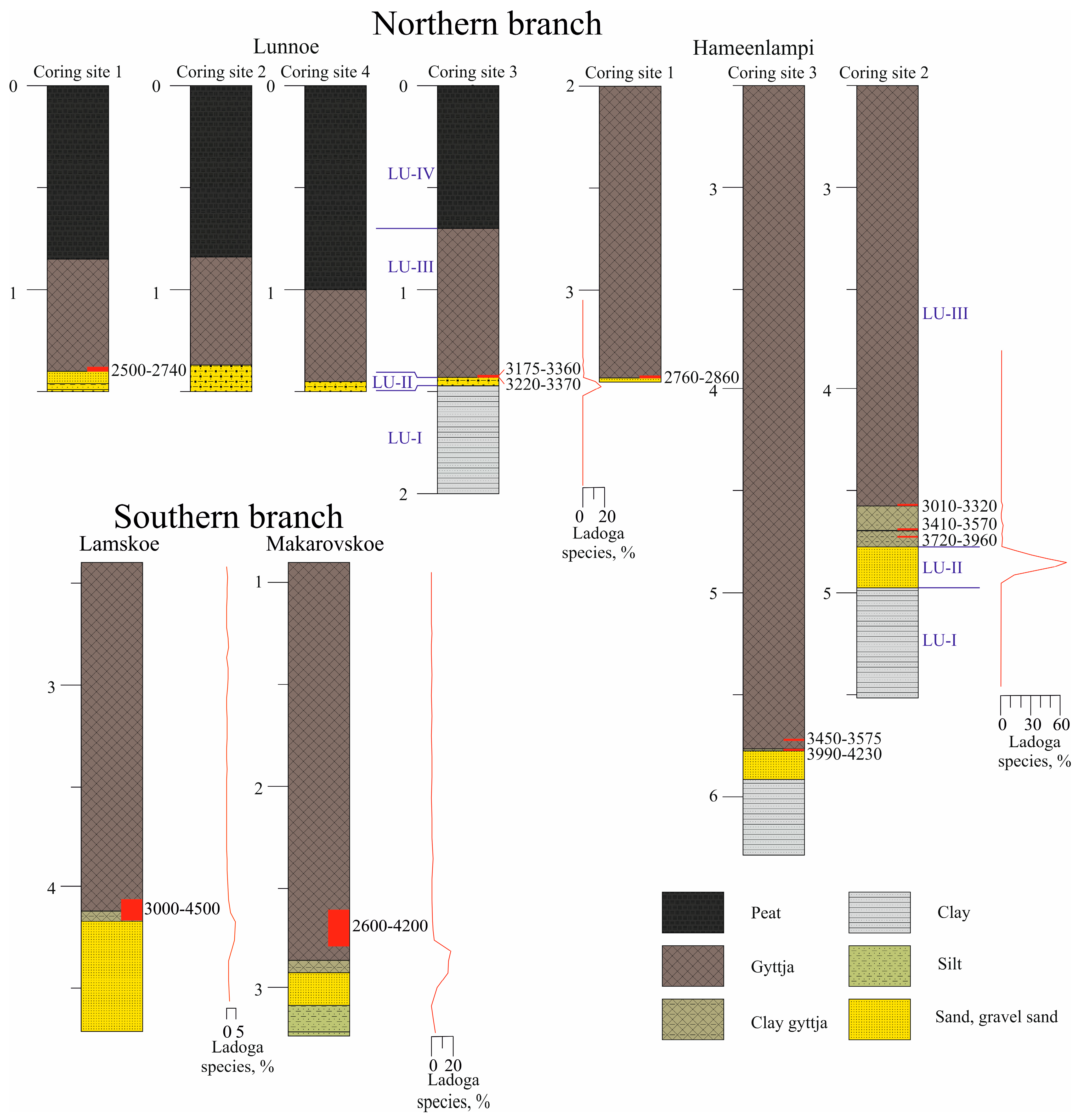
3.1. Age and Lithology
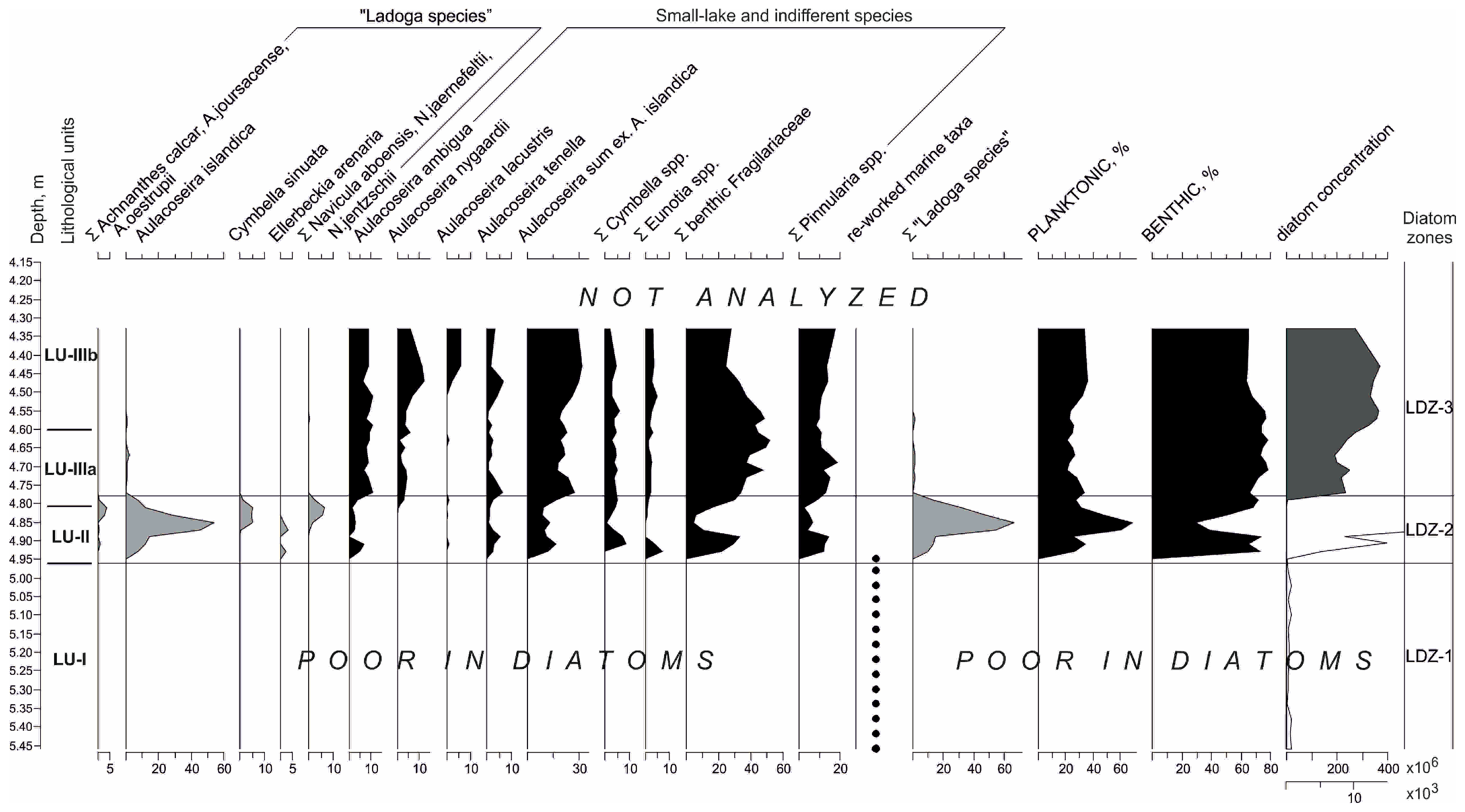
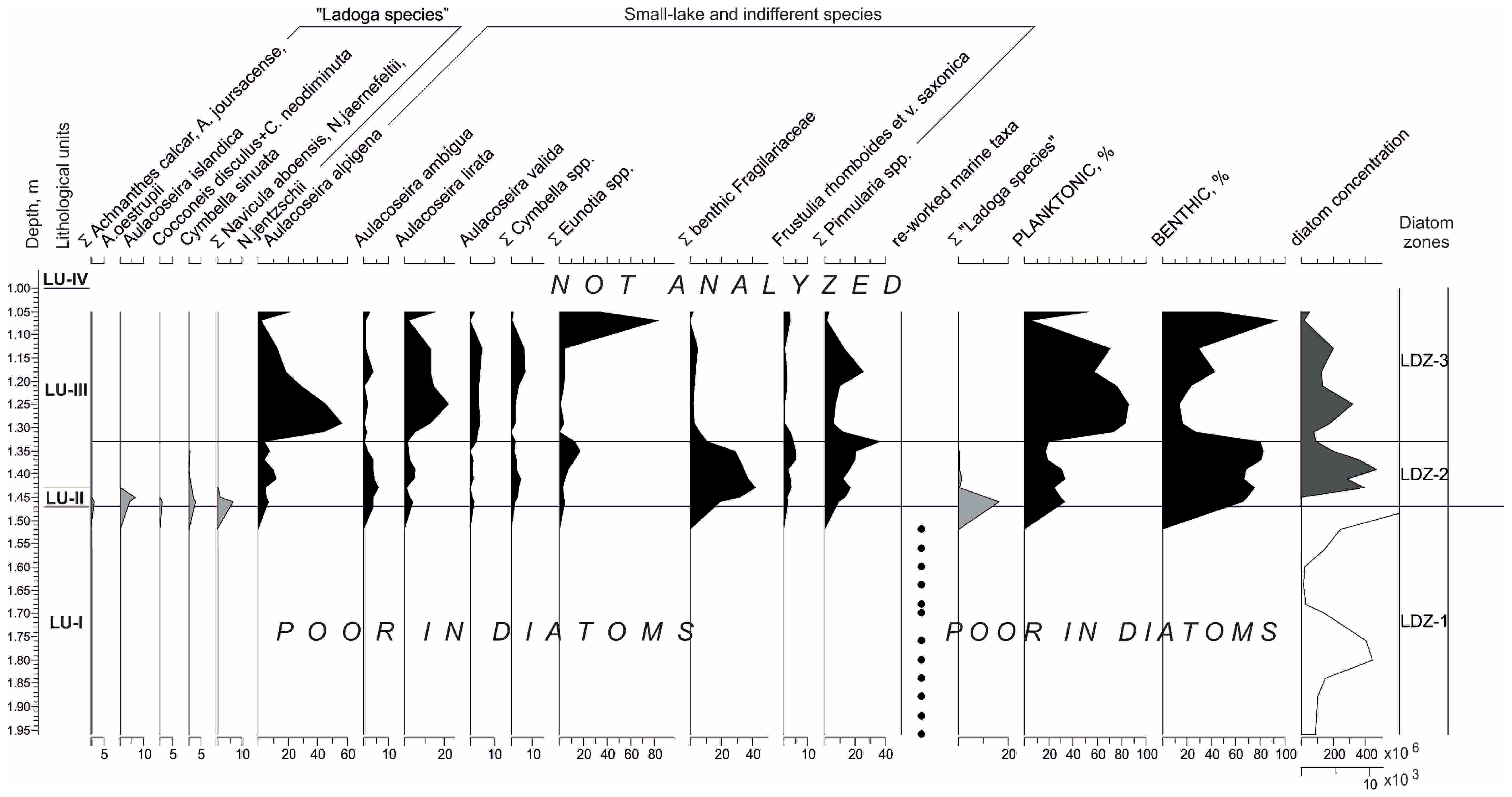
3.2. Diatoms
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bergman, J.; Plikk, A.; Heimdahl, J.; Hagberg, L.; Hallgren, F.; Risberg, J.; Molin, F. When the River Began—The Formation of River Motala Ström and Human Presence in the Early Holocene, Sweden. Quaternary 2020, 3, 25. [Google Scholar] [CrossRef]
- Demidov, I.N. On a maximum stage in the evolution of periglacial Lake Onego, variations in its water level and glacioisostatic shore uplift of the shore in the Late Glacial Period. Geol. Polezn. Iskopayemye Karelii 2006, 9, 171–182. [Google Scholar]
- Kvasov, D.D. Late Quaternary History of Large Lakes and Inland Seas of the Eastern Europe; Nauka: Leningrad, Russia, 1975. [Google Scholar]
- Kvasov, D.D.; Martinson, G.G.; Raukas, A.V. (Eds.) History of Ladoga, Onega, Peipsi, Baikal and Khanka Lakes; Nauka: Leningrad, Russia, 1990. [Google Scholar]
- Larson, G.; Schaetzl, R. Origin and Evolution of the Great Lakes. J. Great Lakes Res. 2001, 27, 518–546. [Google Scholar] [CrossRef]
- Lewis, C.F.M. Holocene lake levels and climate, Lakes Winnipeg, Erie, and Ontario. In Proceedings of the Great Lakes Paleo-Levels Workshop: The Last 4000 Years; Sellinger, C.E., Quinn, F.H., Eds.; NOAA, Great Lakes Environmental Research Laboratory: Ann Arbor, MI, USA, 1999; pp. 6–19. [Google Scholar]
- Saarnisto, M. The late Weichselian and Flandrian history of the Saimaa Lake complex. Soc. Sci. Fenn. Comment. Phys.-Math. 1970, 37, 1–107. [Google Scholar]
- Saarnisto, M. Stratigraphical Studies on the Shoreline Displacement of Lake Superior. Can. J. Earth Sci. 1975, 12, 300–319. [Google Scholar] [CrossRef]
- Seppä, H.; Tikkanen, M.; Mäkiaho, J.-P. Tilting of Lake Pielinen, eastern Finland—An example of extreme transgressions and regressions caused by differential post-glacial isostatic uplift. Est. J. Earth Sci. 2012, 61, 149–161. [Google Scholar] [CrossRef]
- Subetto, D.A.; Potakhin, M.S.; Zobkov, M.B.; Tarasov, A.Y.; Shelekhova, T.S.; Gurbich, V.A. Lake Onego development in the Late Glacial assessed with the use of GIS technologies. Geomorfologia 2019, 3, 83–90. [Google Scholar] [CrossRef]
- Tikkanen, M. Long-term changes in lake and river systems in Finland. Fennia 2002, 180, 31–42. [Google Scholar]
- Subetto, D.A. (Ed.) Paleolimnology of Lake Onego: From the Onego Ice Lake to the Present State; Karelian Research Centre of the Russian Academy of Sciences: Petrozavodsk, Russia, 2022; ISBN 978-5-9274-0955-6. [Google Scholar]
- Påsse, T. Lake-tilting, a method for estimation of glacio-isostatic uplift. Boreas 1998, 27, 69–80. [Google Scholar] [CrossRef]
- Seppä, H.; Tikkanen, M. Land uplift-driven shift of the outlet of Lake Ähtärinjärvi, western Finland. Bull. Geol. Soc. Finl. 2006, 78, 5–18. [Google Scholar] [CrossRef]
- Zobkov, M.; Potakhin, M.; Subetto, D.; Tarasov, A. Reconstructing Lake Onego evolution during and after the Late Weichselian glaciation with special reference to water volume and area estimations. J. Paleolimnol. 2019, 62, 53–71. [Google Scholar] [CrossRef]
- Rumyantsev, V.A. (Ed.) Lake Ladoga and Remarkable Sights on Its Shores. Atlas; Nestor-Istoriya: Saint Petersburg, Russia, 2015. [Google Scholar]
- Koshechkin, B.I.; Ekman, I.M. Holocene transgressions of Lake Ladoga. In Evolution of Natural Environments and the Current State of the Geosystem of Lake Ladoga; Davydova, N.N., Koshechkin, B.I., Eds.; Russian Academy of Sciences, Russian Geographical Society: St. Petersburg, Russia, 1993; pp. 49–60. [Google Scholar]
- Malakhovskiy, D.B.; Arslanov, K.A.; Gej, N.A.; Dzhinoridze, R.N.; Kozyreva, M.G. New data on the Holocene history of Lake Ladoga. In Evolution of Natural Environments and the Current State of the Geosystem of Lake Ladoga; Davydova, N.N., Koshechkin, B.I., Eds.; Russian Academy of Sciences, Russian Geographical Society: St. Petersburg, Russia, 1993; pp. 61–73. [Google Scholar]
- Malakhovskiy, D.B.; Arslanov, K.A.; Gej, N.A.; Dzhinoridze, R.N. New data on the history of the Neva. In Evolution of Natural Environments and the Current State of the Geosystem of Lake Ladoga; Davydova, N.N., Koshechkin, B.I., Eds.; Russian Academy of Sciences, Russian Geographical Society: St. Petersburg, Russia, 1993; pp. 74–84. [Google Scholar]
- Saarnisto, M. Emergence history of the Karelian Isthmus. Iskos 2008, 16, 128–139. [Google Scholar]
- Subetto, D.A. History of the formation of Lake Ladoga and its connection with the Baltic Sea. Terra Humana 2007, 1, 111–120. [Google Scholar]
- Dolukhanov, P.M.; Subetto, D.A.; Arslanov, K.A.; Davydova, N.N.; Zaitseva, G.I.; Kuznetsov, D.D.; Ludikova, A.V.; Sapelko, T.V.; Savelieva, L.A. Holocene oscillations of the Baltic Sea and Lake Ladoga levels and early human movements. Quat. Int. 2010, 220, 102–111. [Google Scholar] [CrossRef]
- Kuznetsov, D.D.; Subetto, D.A.; Sapelko, T.V.; Ludikova, A.V. Hydrographic network of northern part of the Karelian Isthmus in the Holocene according to sediments of the small lakes. Geomorfologiya 2015, 1, 54–69. [Google Scholar] [CrossRef]
- Subetto, D.A. Lake Sediments: Palaeolimnological Reconstructions; Publishing House of Herzen State Pedagogical University: St. Petersburg, Russia, 2009; ISBN 978-5-8064-1444-2. [Google Scholar]
- Björck, S. The late Quaternary development of the Baltic Sea basin. In Assessment of Climate Change for the Baltic Sea Basin; von Storch, H., Ed.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 398–407. ISBN 978-3-540-72785-9. [Google Scholar]
- Amantov, A.V.; Amantova, M.G. Modeling of postglacial development of Lake Ladoga and eastern part of the Gulf of Finland. Reg. Geol. Metallog. 2017, 69, 5–14. [Google Scholar]
- Saarnisto, M.; Grönlund, T. Shoreline displacement of Lake Ladoga—New data from Kilpolansaari. Hydrobiologia 1996, 322, 205–215. [Google Scholar] [CrossRef]
- Virtasalo, J.J.; Ryabchuk, D.; Kotilainen, A.T.; Zhamoida, V.; Grigoriev, A.; Sivkov, V.; Dorokhova, E. Middle Holocene to present sedimentary environment in the Easternmost Gulf of Finland (Baltic Sea) and the birth of the Neva River. Mar. Geol. 2014, 350, 84–96. [Google Scholar] [CrossRef]
- Malakhovskii, D.B.; Markov, K.K. (Eds.) Geomorphology and Quaternary Deposits of the NW of the European Part of the USSR; Nauka: Leningrad, Russia, 1969. [Google Scholar]
- Ailio, J. Die geographische Entwicklund des Ladogasees in postglacialer Zeit. Bull. Comm. Géol. Finl. 1915, 45, 1–159. [Google Scholar]
- Ludikova, A.V.; Subetto, D.A.; Davydova, N.N.; Sapelko, T.V.; Arslanov, K.A. Holocene Lake Ladoga level fluctuations inferred from sediments of the lake of St. Sergey (Putsari Island). Izv. Russ. Geogr. O-va 2005, 137, 34–41. [Google Scholar]
- Ludikova, A.V.; Shatalova, A.E.; Subetto, D.A.; Kublitskiy; Yu, A.; Rosentau, A.; Hang, T. Diatom-inferred palaeolimnological changes in a small lake in the context of the Holocene Baltic Sea transgressions: A case study of Lake Goluboye, Karelian Isthmus (NW Russia). IOP Conf. Ser. Earth Environ. Sci. 2020, 438, 012014. [Google Scholar] [CrossRef]
- Sapelko, T.V.; Kuznetsov, D.D.; Korneenkova, N.Y.; Denisenkov, V.P.; Ludikova, A.V. Paleolimnology of inland lakes of Putsari Island (Lake Ladoga). Izv. Russ. Geogr. O-va 2014, 146, 29–40. [Google Scholar]
- Shatalova, A.E.; Ludikova, A.V.; Subetto, D.A.; Kublitskij, Y.A.; Demidionov, M.Y. Reconstruction of the transgressive-regressive stages of the Baltic Sea in the Holocene based on diatom analysis of bottom sediments of the isolation basin on the Karelian Isthmus. Astrakhan Bull. Ecol. Educ. 2022, 6, 58–68. [Google Scholar] [CrossRef]
- Ludikova, A.V.; Kuznetsov, D.D.; Subetto, D.A.; Sapelko, T.V. The use of isolation basins approach in reconstructing Lake Ladoga level changes. In All-Russian Conference on Large Inland Basins (V Ladoga Symposium); Rumyantsev, V.A., Pozdnyakov, S.R., Eds.; Lema: St. Petersburg, Russia, 2016; pp. 247–253. [Google Scholar]
- Hedenström, A.; Risberg, J. Early Holocene shore-displacement in southern central Sweden as recorded in elevated isolated basins. Boreas 1999, 28, 490–504. [Google Scholar] [CrossRef]
- Jouse, A.P. Diatoms in Holocene marine and continental sediments. In Holocene Questions; Neystadt, M.Y., Gudelis, V., Eds.; Vilnus, Lithuania, 1961; pp. 115–139. [Google Scholar]
- Alenius, T.; Gerasimov, D.; Sapelko, T.; Ludikova, A.; Kuznetsov, D.; Golyeva, A.; Nordqvist, K. Human-environment interaction during the Holocene along the shoreline of the Ancient Lake Ladoga: A case study based on palaeoecological and archaeological material from the Karelian Isthmus, Russia. Holocene 2020, 30, 1622–1636. [Google Scholar] [CrossRef]
- Miettinen, A.; Savelieva, L.; Subetto, D.A.; Djinoridze, R.; Arslanov, K.; Hyvärinen, H. Palaeoenvironment of the Karelian Isthmus, the easternmost part of the Gulf of Finland, during the Litorina Sea stage of the Baltic Sea history. Boreas 2007, 36, 441–458. [Google Scholar] [CrossRef]
- Znamenskaya, O.M.; Vishnevskaya, E.M.; Kleimenova, G.I.; Malakhovsky, D.B. Problems of Environmental Protection; Leningrad State University Press: Leningrad, Russia, 1980; pp. 133–155. [Google Scholar]
- Ludikova, A.V. Diatom evidences for the mid-holocene Lake Ladoga transgression. Izv. Russ. Geogr. O-va 2015, 4, 38–51. [Google Scholar]
- Kuznetsov, D.D.; Subetto, D.A.; Ludikova, A.V. The Connection between Lake Ladoga and the Baltic Sea in the Late Holocene: New Paleolimnological Data. Dokl. Earth Sci. 2022, 506, 693–697. [Google Scholar] [CrossRef]
- Davydova, N.N. Diatoms as Indicators of the Environmental Conditions of Water-Bodies in the Holocene; Nauka: Leningrad, Russia, 1985. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae 1. Teil: Naviculaceae. In Süsswasserflora von Mitteleuropa; Ettl, H., Gerloff, J., Heynig, H., Mollenhauer, D., Eds.; G. Fischer: Stuttgart, Germany; New York, NY, USA, 1986; Volume 2/1. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae. 2. Teil: Bacillariaceae, Epithemiaceae, Surirellaceae. In Süsswasserflora von Mitteleuropa; Ettl, H., Gerloff, J., Heynig, H., Mollenhauer, D., Eds.; Gustav Fischer Verlag: Jena, Germany, 1988; Volume 2/2. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae. 3. Teil: Centrales, Fragilariaceae, Eunotiaceae. In Süsswasserflora von Mitteleuropa; Ettl, H., Gerloff, J., Heynig, H., Mollenhauer, D., Eds.; Gustav Fischer Verlag: Stuttgart, Germany; Jena, Germany, 1991; Volume 2/3. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae. 4. Teil: Achnanthaceae, Kritische Ergänzungen zu Navicula (Lineolatae) und Gomphonema, Gesamtliteraturverzeichnis. In Süsswasserflora von Mitteleuropa; Ettl, H., Gerloff, J., Heynig, H., Mollenhauer, D., Eds.; Gustav Fischer Verlag: Stuttgart, Germany; Jena, Germany, 1991; Volume 2/4. [Google Scholar]
- Juggins, S. C2 Vesrion 1.5 User Guide. Software for Ecological and Palaeoecological Data Analysis and Visualisation; Newcastle University: Newcastle upon Tyne, UK, 2007. [Google Scholar]
- Bronk Ramsey, C. Bayesian analysis of radiocarbon dates. Radiocarbon 2009, 51, 337–360. [Google Scholar] [CrossRef]
- Reimer, P.J.; Austin, W.E.N.; Bard, E.; Bayliss, A.; Blackwell, P.G.; Bronk Ramsey, C.; Butzin, M.; Cheng, H.; Edwards, R.L.; Friedrich, M.; et al. The IntCal20 Northern Hemisphere Radiocarbon Age Calibration Curve (0–55 cal kBP). Radiocarbon 2020, 62, 725–757. [Google Scholar] [CrossRef]
- Kuznetsov, D.D.; Subetto, D.A. Stratigraphy of Lake Sediments of the Karelian Isthmus; GEOS: Moscow, Russia, 2019; ISBN 978-5-89118-801-3. [Google Scholar]
- Krasnov, I.I.; Zarrina, E.P. (Eds.) Leningrad and Leningrad Region: Guide to Excursions A–15, C–15; Moskow, Russia, 1982. [Google Scholar]
- Cheremisinova, E.A. Marine diatom flora of Quaternary sediments from Ladoga basin. Byullyuten Kom. Izucheniyu Chetvertichnykh Otlozhenii 1957, 21, 105–112. [Google Scholar]
- Miettinen, A.; Rinne, K.; Haila, H.; Hyvarinen, H.; Eronen, M.; Delusina, I.; Kadastik, E.; Kalm, V.; Gibbard, P. The marine Eemian of the Baltic: New pollen and diatom data from Peski, Russia, and Põhja-Uhtju, Estonia. J. Quat. Sci. 2002, 17, 445–458. [Google Scholar] [CrossRef]
- Weckström, K.; Juggins, S. Coastal diatom-environment relationships from the Gulf of Finland, Baltic Sea. J. Phycol. 2005, 42, 21–35. [Google Scholar] [CrossRef]
- Saarnisto, M. Late Holocene land uplift/neotectonics on the island of Valamo (Valaam), Lake Ladoga, NW Russia. Quat. Int. 2012, 260, 143–152. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| № IGANAMS | Lake, Coring Site, Core № | Sampling Depth (m) below the Water Surface | Sediment Composition | Dated Material | 14C Date, BP | Calibrated Age, Mean ± σ (95.4% Probability) | Data Source |
|---|---|---|---|---|---|---|---|
| 8931 | Lunnoe, 1,1 | 1.38–1.40 | gyttja | TOC | 2540 ± 25 | 2500–2740, 2635 ± 80 | [42] |
| 8932 | Lunnoe, 1,1 | 1.45–1.46 | sand | Plant macrofossil | 245 ± 30 | Present | This publication |
| 8933 | Lunnoe, 3,3 | 1.46–1.47 | gyttja | TOC | 3085 ± 30 | 3220–3370, 3295 ± 40 | [42] |
| 8934 | Lunnoe, 3,4 | 1.46–1.47 | sand | Plant macrofossil | 127 ± 0.366 | Present | This publication |
| 8935 | Lunnoe, 3,4 | 1.42–1.43 | gyttja | TOC | 3055 ± 30 | 3175–3360, 3270 ± 50 | [42] |
| 8942 | Hameenlampi, 1,1 | 3.42–3.43 | gyttja | TOC | 2720 ± 25 | 2760–2870, 2810 ± 30 | [42] |
| 8943 | Hameenlampi, 2,2 | 4.705–4.715 | clayey gyttja | TOC | 3555 ± 30 | 3720–3960, 3840 ± 60 | [42] |
| 8944 | Hameenlampi, 2,2 | 4.67–4.68 | clayey gyttja | TOC | 3275 ± 30 | 3410–3570, 3490 ± 40 | This publication |
| 8945 | Hameenlampi, 2,2 | 4.55–4.56 | gyttja | TOC | 2980 ± 30 | 3010–3320, 3150 ± 60 | This publication |
| 8948 | Hameenlampi, 3,8 | 5.75–5.76 | clayey gyttja | TOC | 3750 ± 25 | 3990–4230, 4100 ± 60 | [42] |
| 8949 | Hameenlampi, 3,8 | 5.70–5.71 | gyttja | TOC | 3300 ± 30 | 3450–3575, 3520 ± 40 | This publication |
| Lake Hameenlampi, Coring Site 2 | ||||
| Lithological Unit (LU) | Depth, m | Lithological Description/Contact with the Overlaying Unit | LOI, % Range (Mean) | Local Diatom Zone (LDZ) |
| IIIb | 4.6–2.5 | Brown homogenous gyttja | 23–39 (36) | 3 (upper 3 m n.a.) |
| IIIa | 4.81–4.6 | Light-brown to gray clayey gyttja/gradual, indistinct | 15–24 (19) | 3 |
| II | 4.96–4.81 | Sand with gravel/sharp | 0.3–0.6 (0.4) | 2 |
| I | 5.5–4.96 | Dense gray clay/sharp | 0.4–1.1 (0.7) | 1 |
| Lake Lunnoe, coring site 3 | ||||
| Lithological Unit (LU) | Depth, m | Lithological Description/Contact with the Overlaying Unit | LOI, % Range (Mean) | Local Diatom Zone (LDZ) |
| IV | 0.7–0.0 | Poorly decomposed peat | n.a. | n.a. |
| III | 1.43–0.7 | Brown homogenous gyttja/distinct | 26–64 (46) | 3 |
| II | 1.47–1.43 | Sand with gravel/sharp | 1.0–1.3 (1.2) | 2 |
| I | 2.0–1.47 | Dense gray clay/sharp | 0.4–3.0 (1.1) | 1 |
| “Ladoga Species” | Hameenlampi | Lunnoe | Makarovskoe | Lamskoe |
|---|---|---|---|---|
| Achnanthes calcar Cleve | + | + | + | + |
| A. clevei Grunow | + | + | ||
| A. joursacense Héribaud | + | |||
| A. oestrupii (A.Cleve) Hustedt | + | + | + | + |
| Aulacoseira islandica (O.Müller) Simonsen | + | + | + | + |
| Cocconeis disculus (Schumann) Cleve | + | + | + | |
| C. neodiminuta Krammer | + | + | + | + |
| Cyclotella cf. iris | + | + | ||
| C. schumannii (Grunow) H.Håkansson | + | + | + | + |
| Cymbella sinuata W.Gregory | + | + | ||
| Didymosphenia geminata (Lyngbye) Mart. Schmidt | + | + | ||
| Diploneis elliptica var. ladogensis Cleve | + | |||
| Ellerbeckia arenaria (D.Moore ex Ralfs) R.M. Crawford | + | + | + | |
| Eunotia clevei Grunow | + | |||
| Gyrosigma attenuatum (Kützing) Rabenhorst | ||||
| Navicula aboensis (Cleve) Hustedt | + | + | + | + |
| N. jaernefeltii Hustedt | + | + | + | + |
| N. jentzschii Grunow | + | + | + | |
| N. scutelloides W.Smith ex W.Gregory | + | + | ||
| N. tuscula Ehrenberg | + | |||
| Opephora martyi Héribaud | + | + | ||
| Rhoicosphenia abbreviata (C.Agardh) Lange-Bertalot | + | + | ||
| Stephanodiscus cf. medius | + | + | + | |
| S. neoastraea Håkansson & Hickel | + | + | + | + |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ludikova, A.V.; Subetto, D.A.; Kuznetsov, D.D.; Orlov, A.V.; Shatalova, A.E. New Diatom and Sedimentary Data Confirm the Existence of the Northern Paleo-Outlet from Lake Ladoga to the Baltic Sea. Quaternary 2024, 7, 31. https://doi.org/10.3390/quat7030031
Ludikova AV, Subetto DA, Kuznetsov DD, Orlov AV, Shatalova AE. New Diatom and Sedimentary Data Confirm the Existence of the Northern Paleo-Outlet from Lake Ladoga to the Baltic Sea. Quaternary. 2024; 7(3):31. https://doi.org/10.3390/quat7030031
Chicago/Turabian StyleLudikova, Anna V., Dmitry A. Subetto, Denis D. Kuznetsov, Alexander V. Orlov, and Angelina E. Shatalova. 2024. "New Diatom and Sedimentary Data Confirm the Existence of the Northern Paleo-Outlet from Lake Ladoga to the Baltic Sea" Quaternary 7, no. 3: 31. https://doi.org/10.3390/quat7030031
APA StyleLudikova, A. V., Subetto, D. A., Kuznetsov, D. D., Orlov, A. V., & Shatalova, A. E. (2024). New Diatom and Sedimentary Data Confirm the Existence of the Northern Paleo-Outlet from Lake Ladoga to the Baltic Sea. Quaternary, 7(3), 31. https://doi.org/10.3390/quat7030031