
The Sardinian Mammoth’s Evolutionary History: Lights and Shadows


Abstract
1. Introduction
2. Methods
3. Historical Background
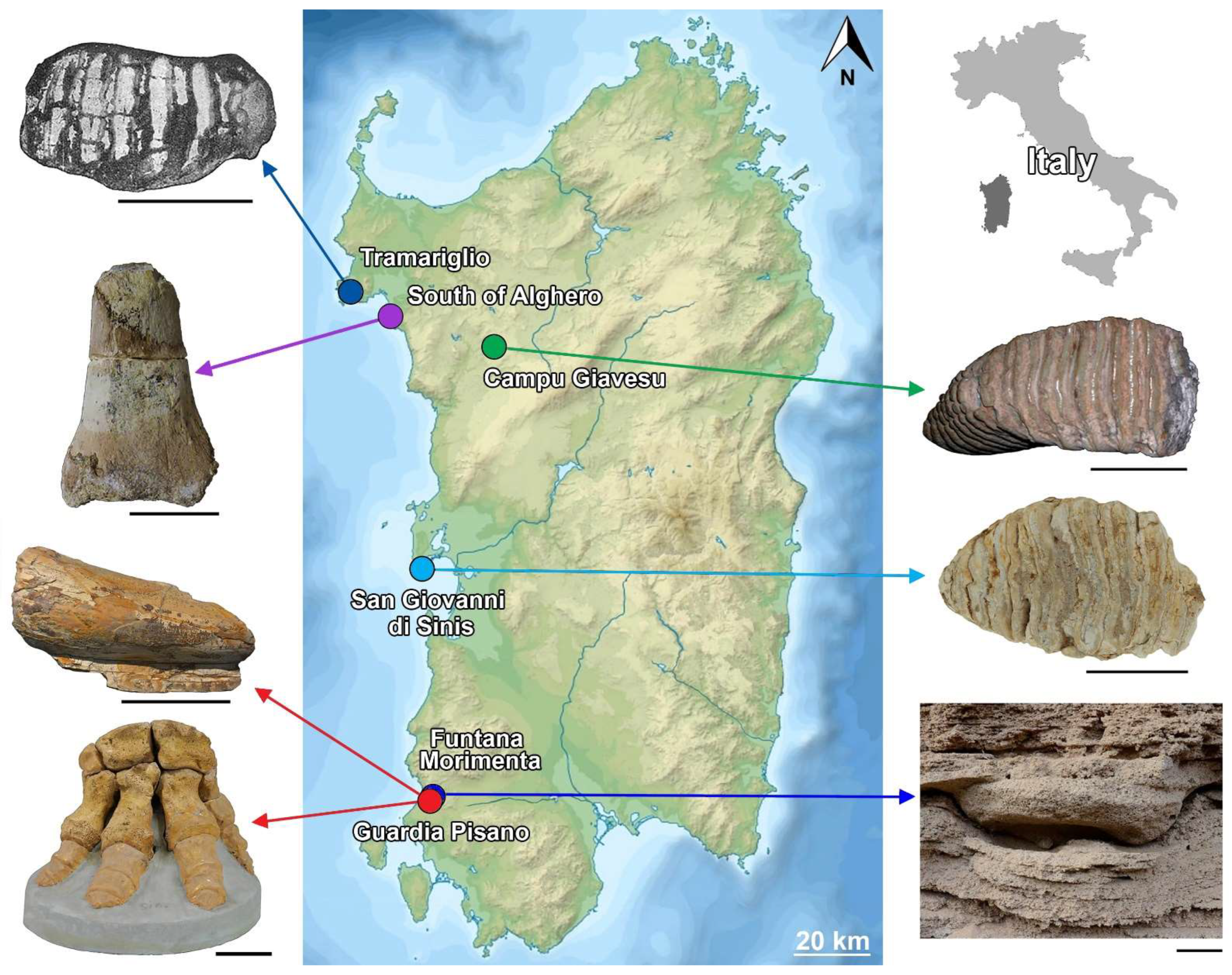
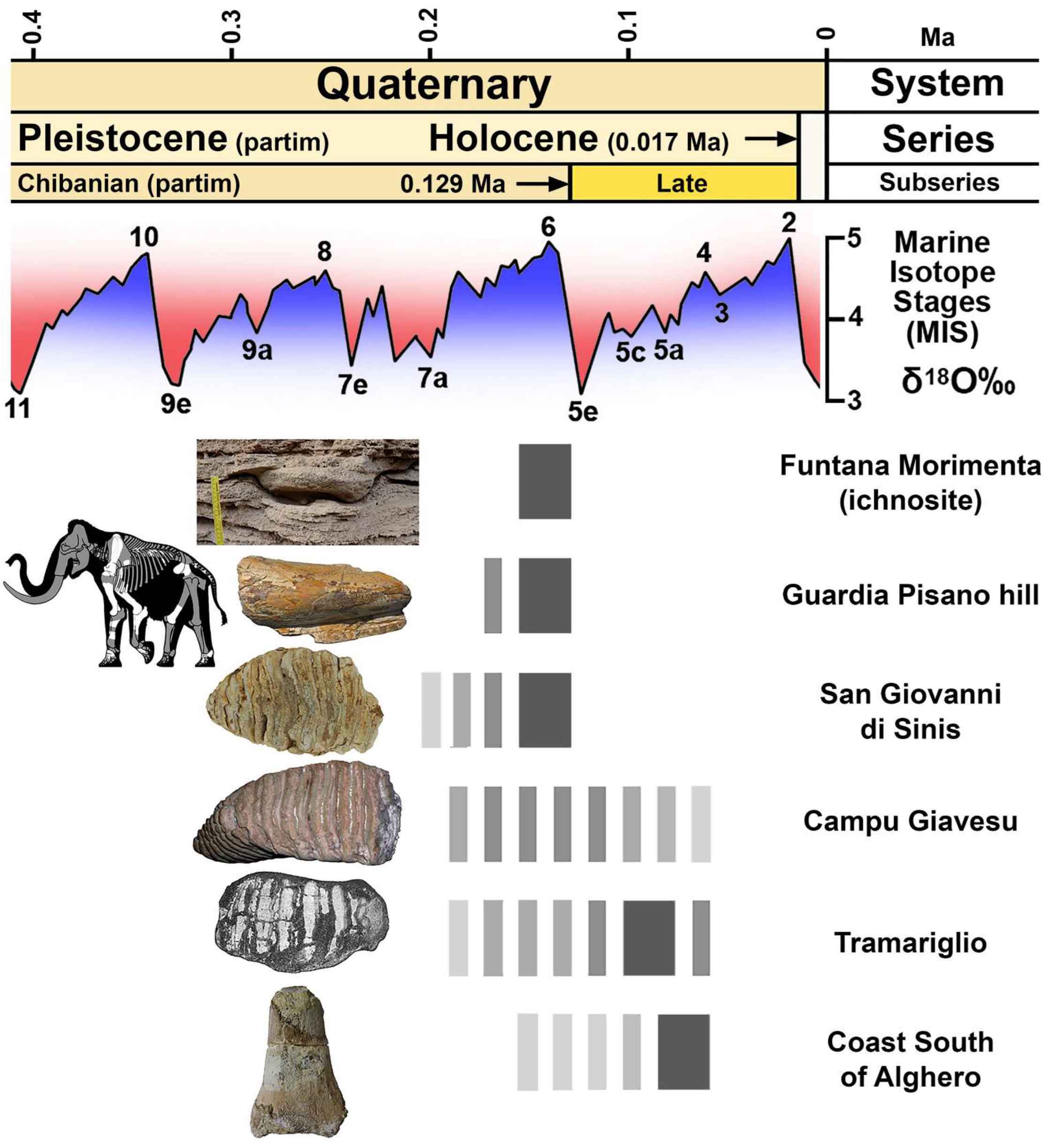
4. The Sardinian Mammoth Remains: A Critical Synopsis
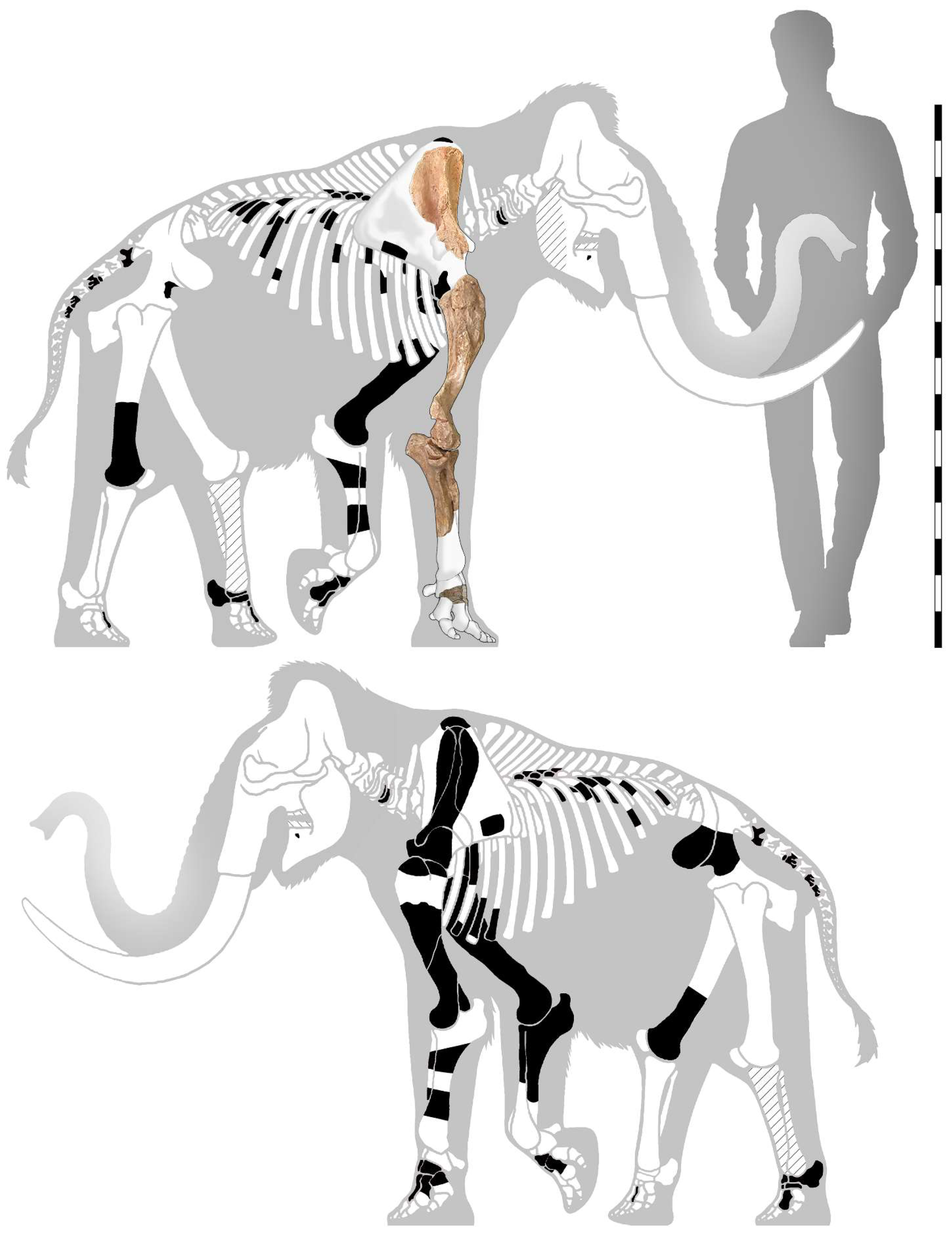
4.1. The Mammoth Skeleton and Other Remains from Guardia Pisano Hill, Funtana Morimenta, and the Surrounding Area
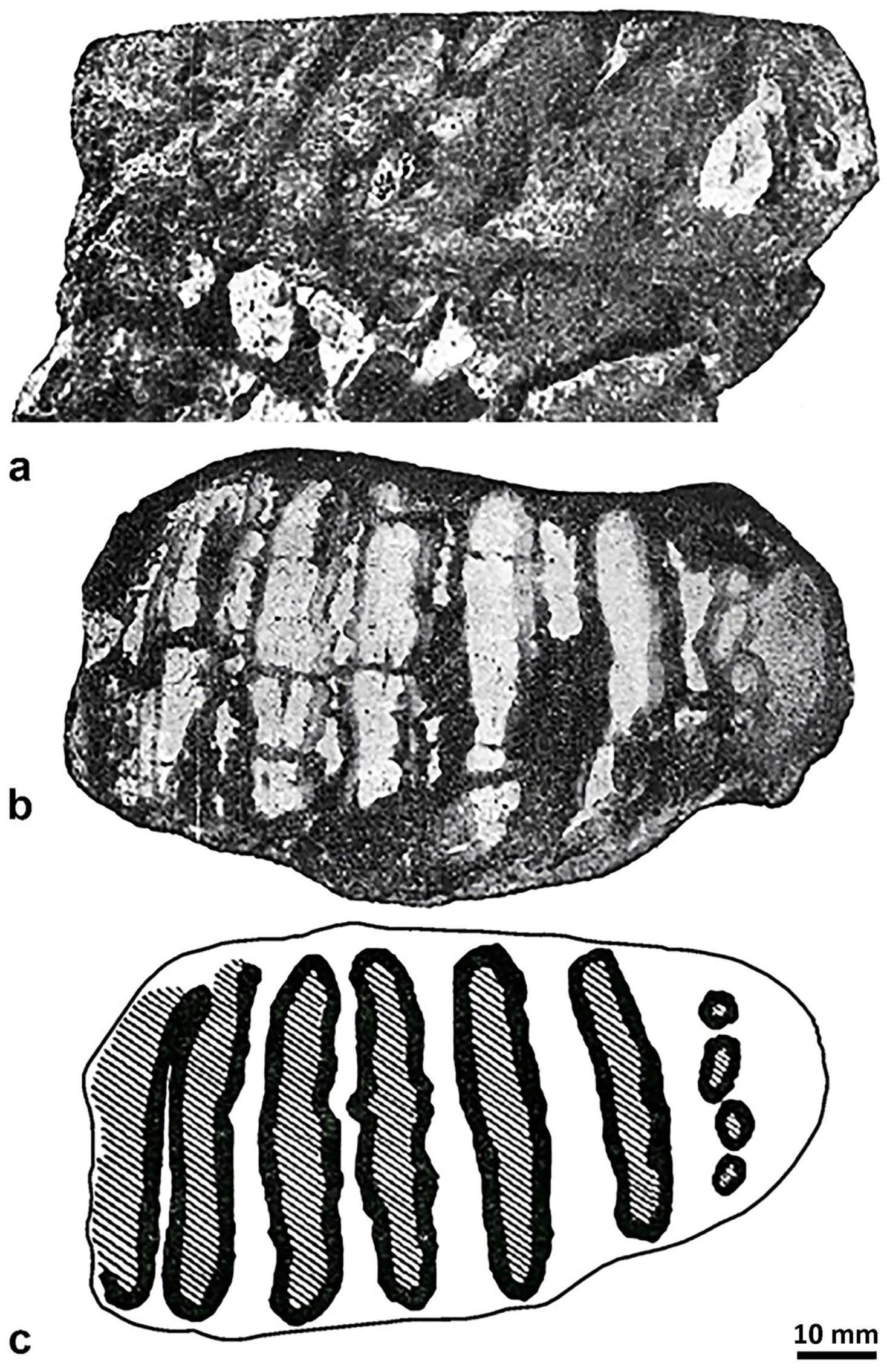
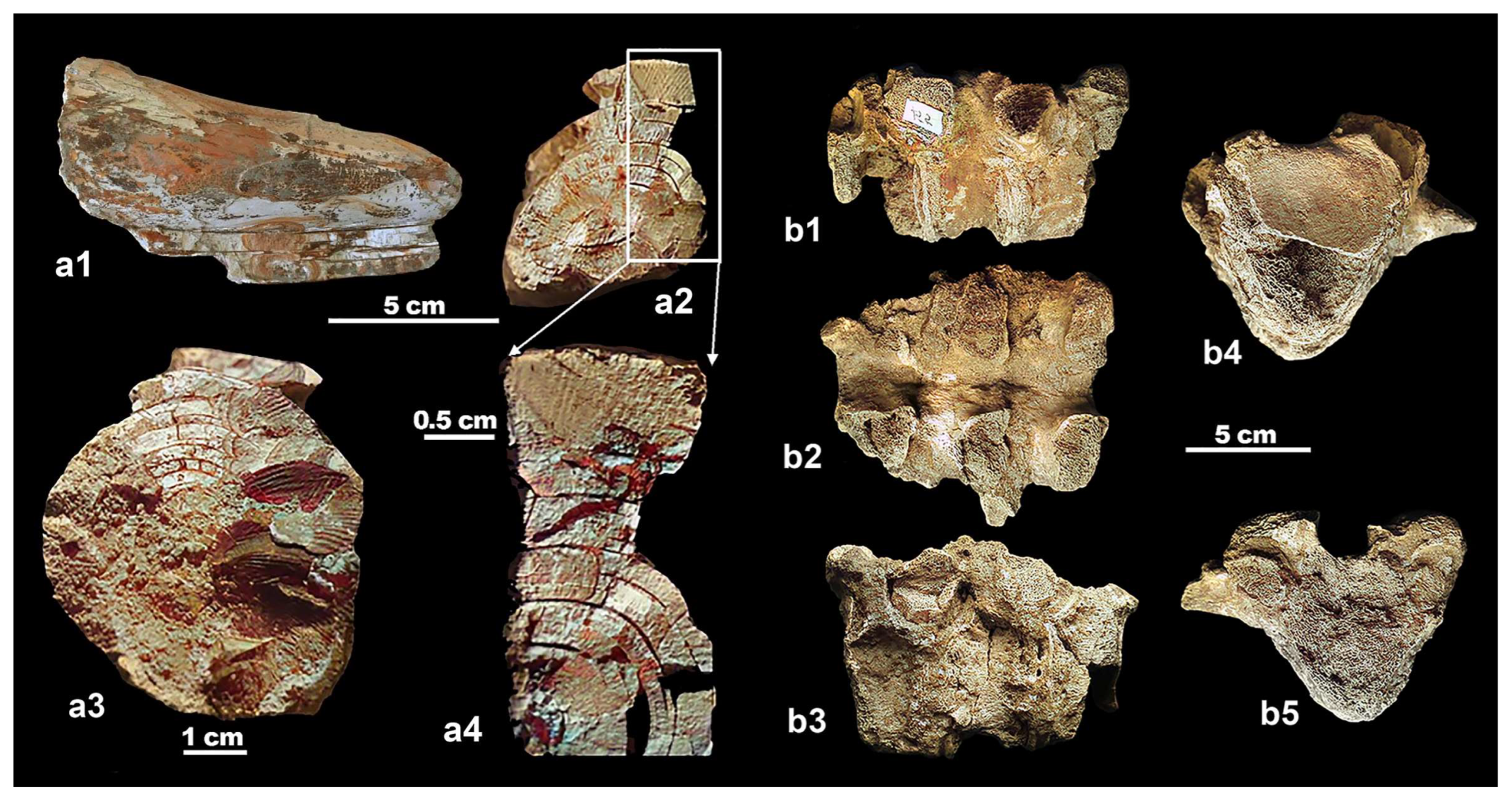
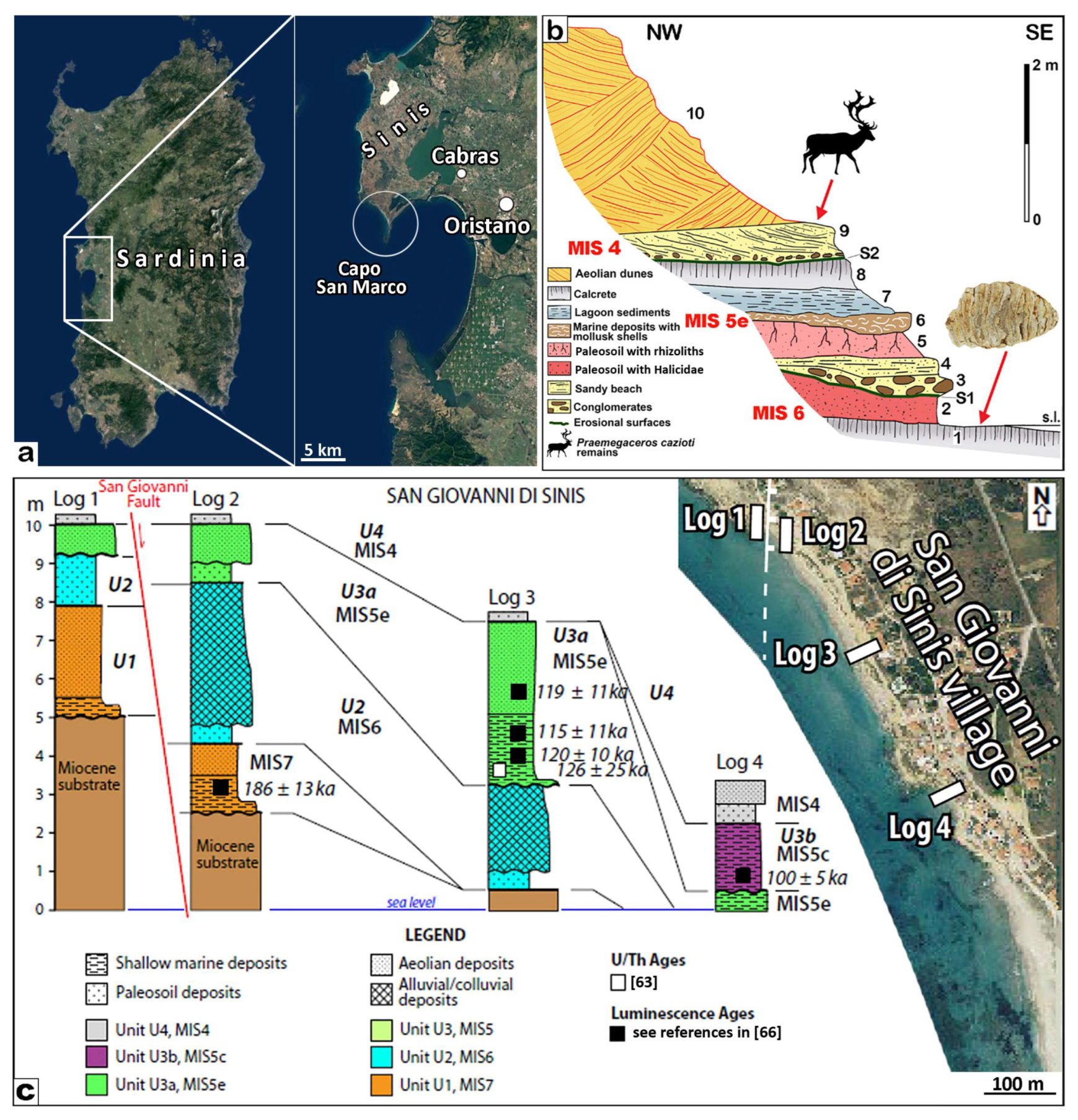
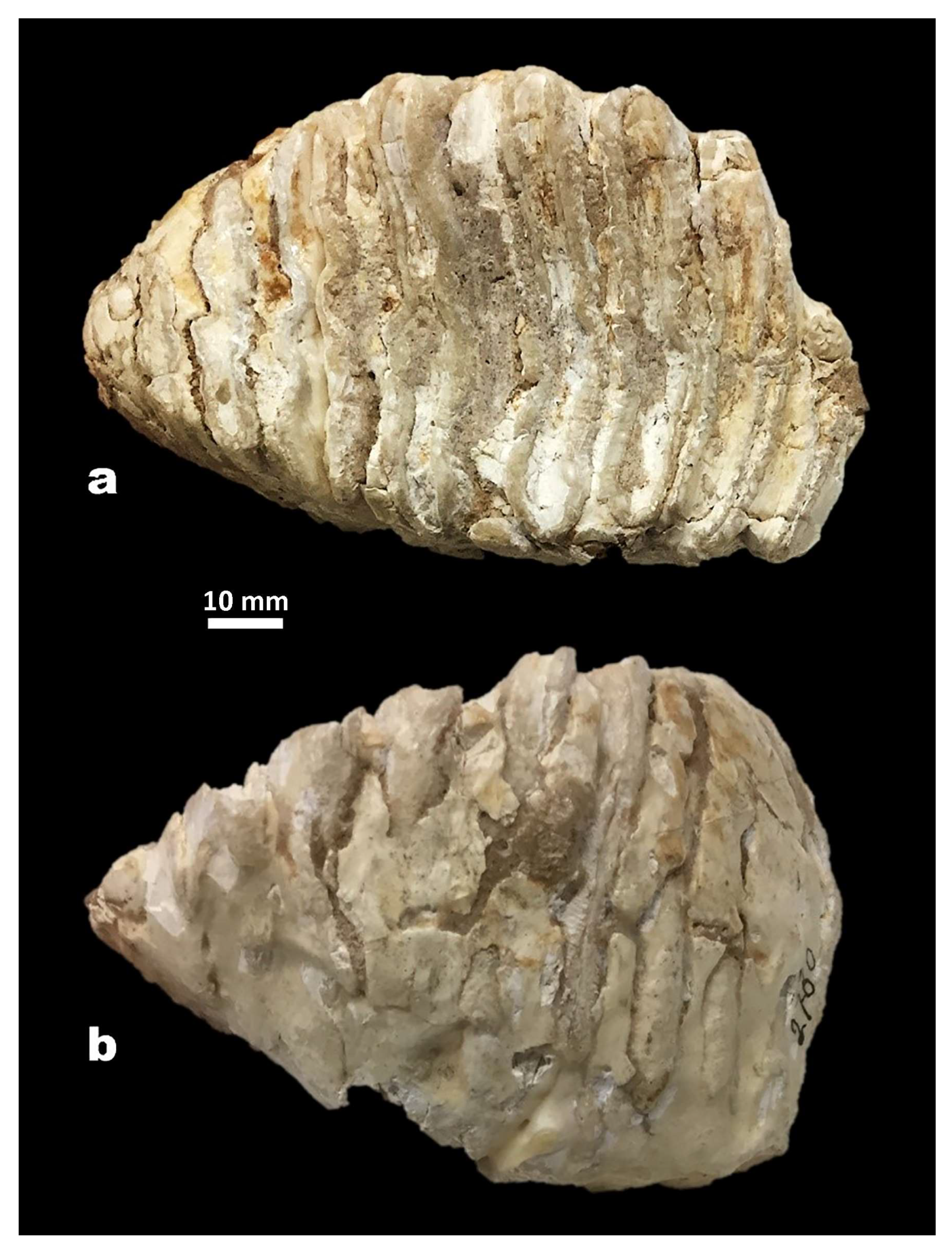
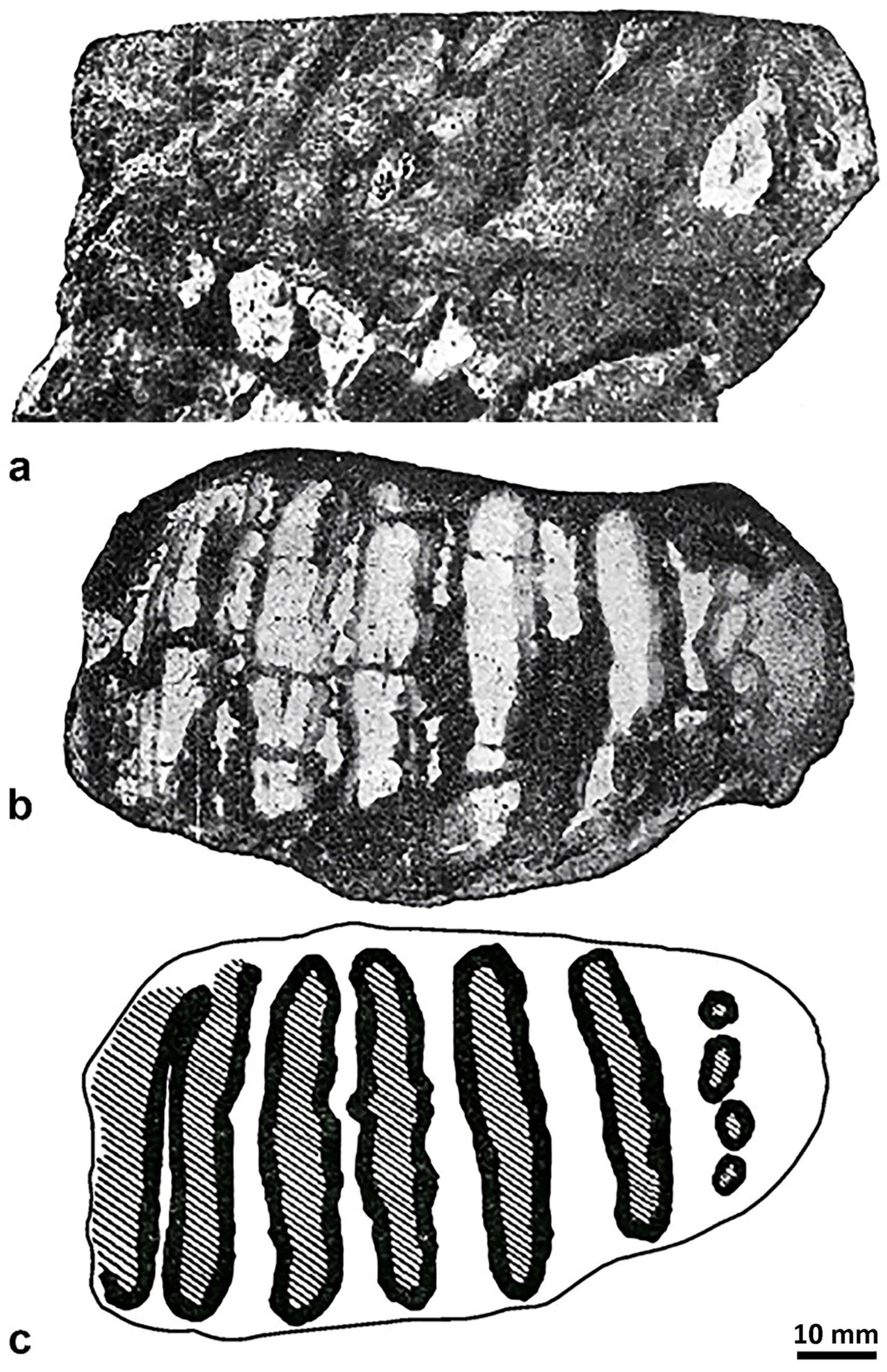
4.2. Molar from San Giovanni di Sinis
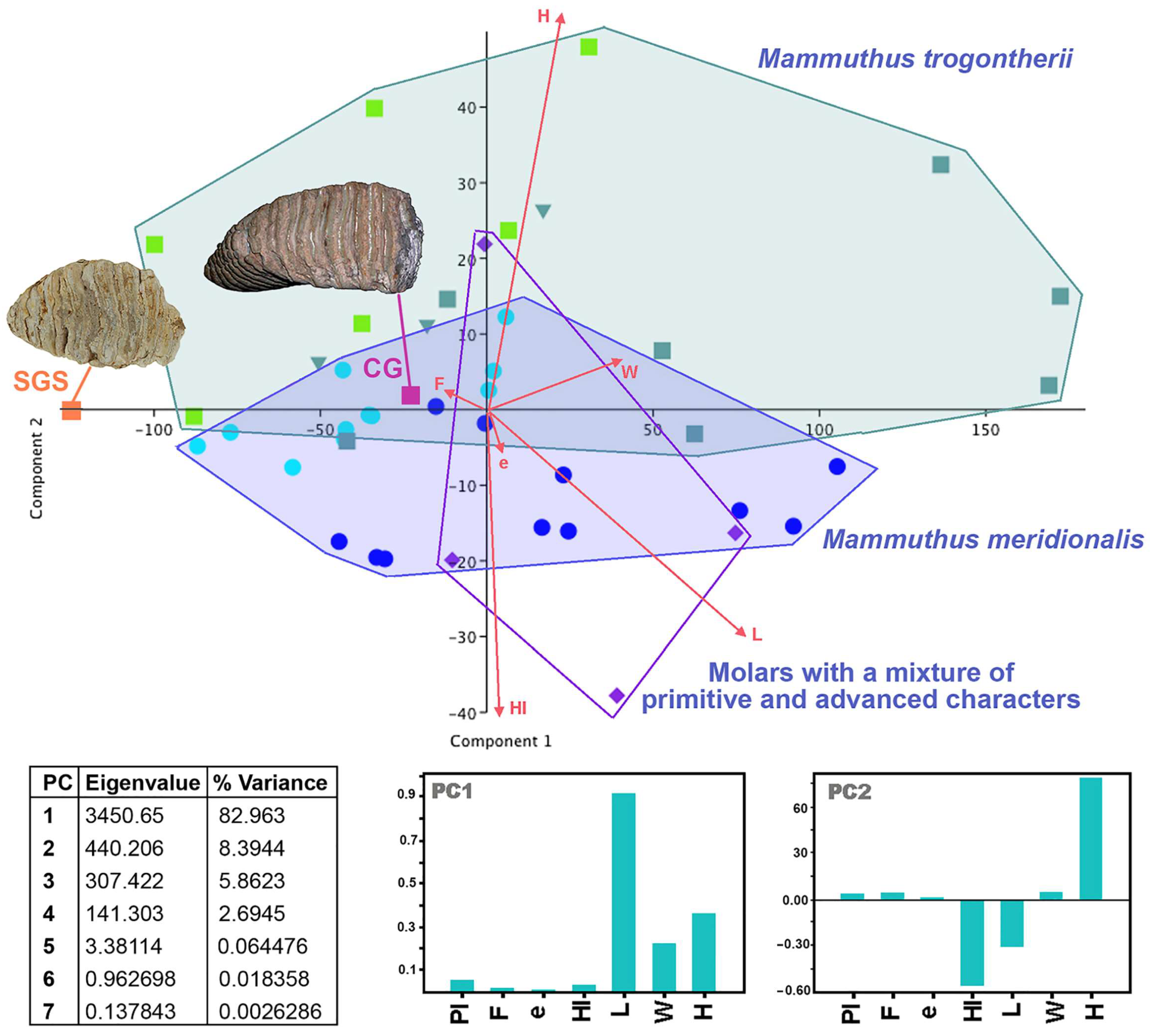
4.3. The Molars from Campu Giavesu
4.4. The Problematic Lost Molar from the Tramariglio Bone Breccia
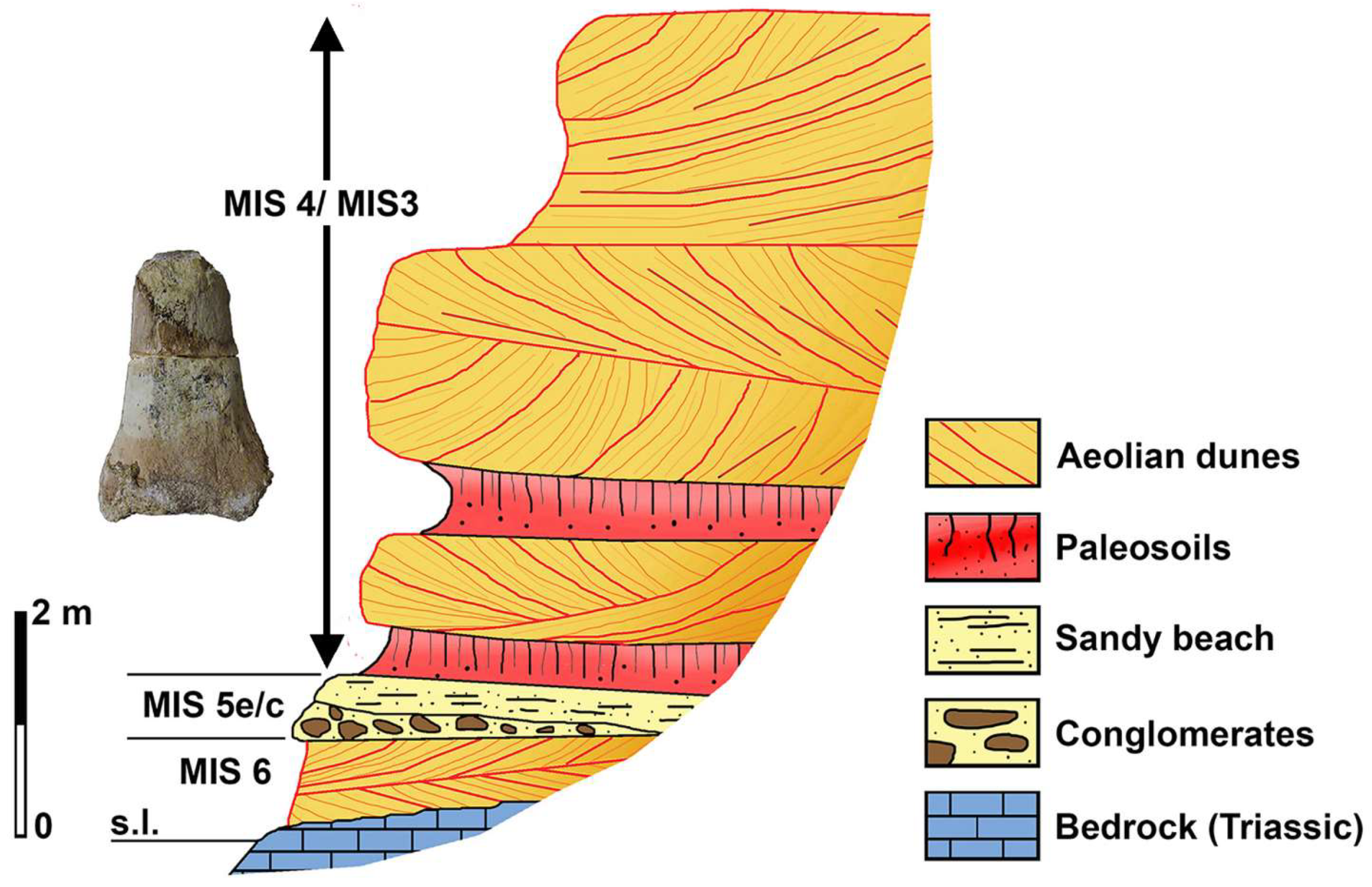
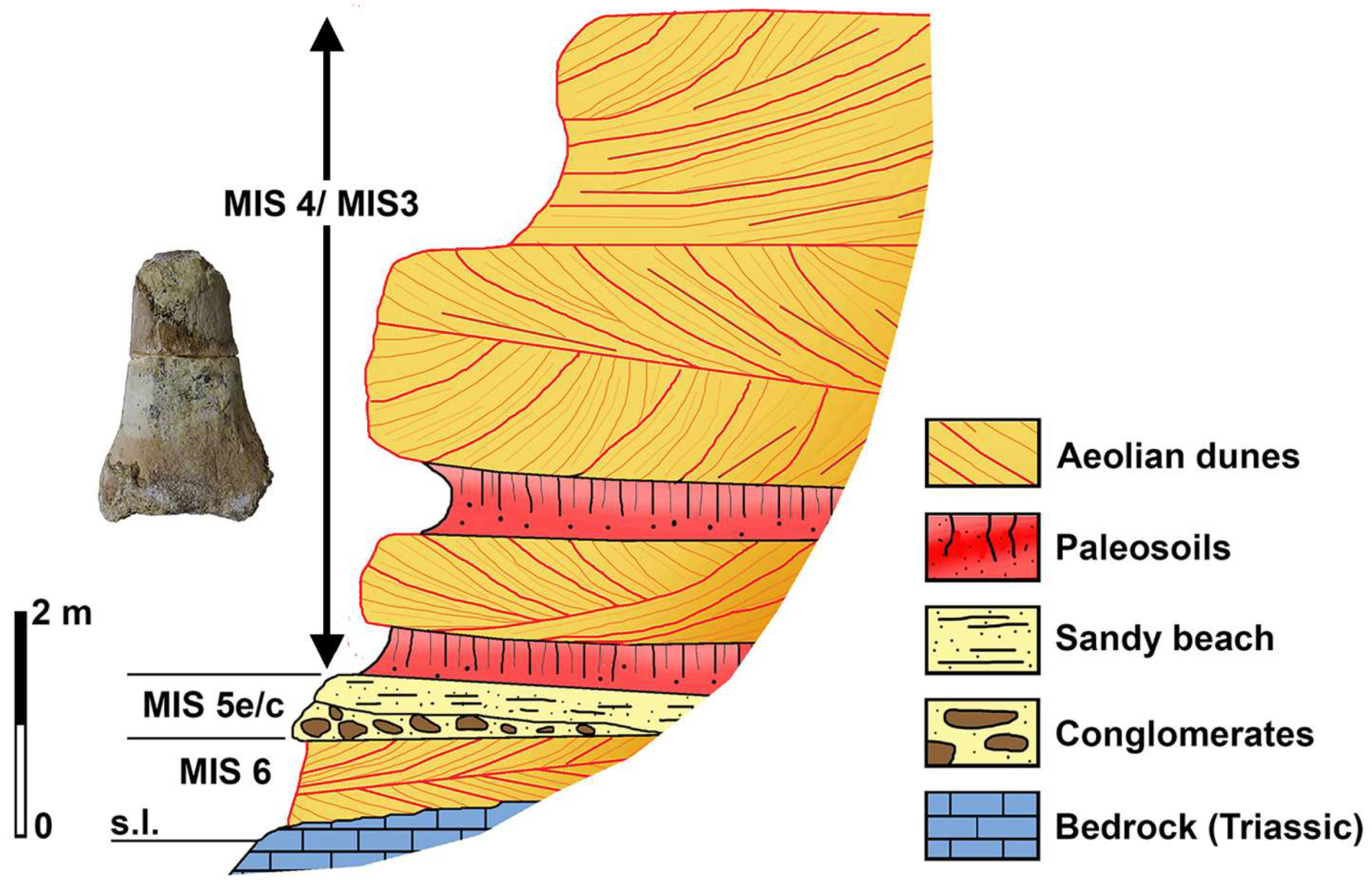
4.5. The Mammoth Remains from the Alghero Aeolian Deposits
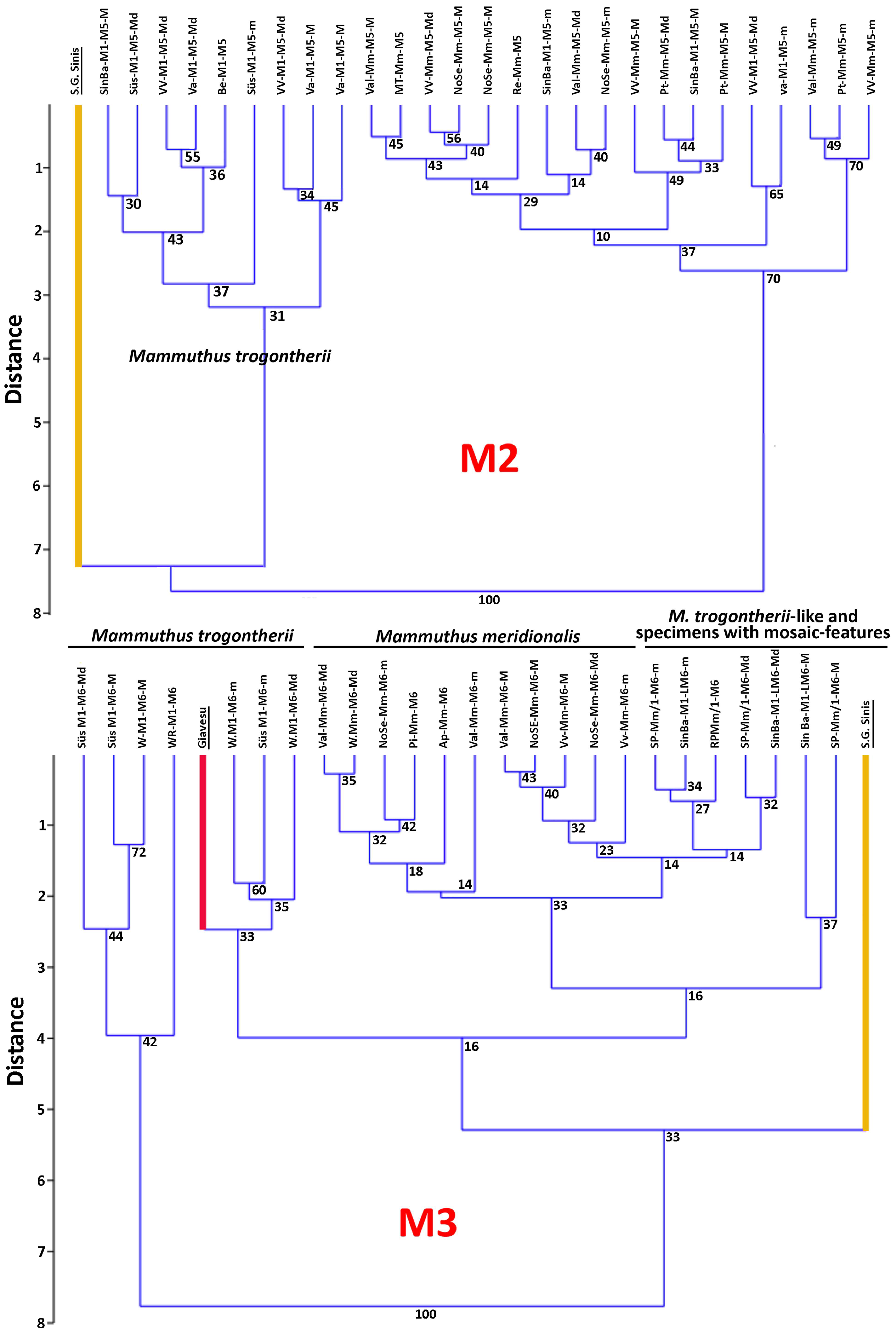
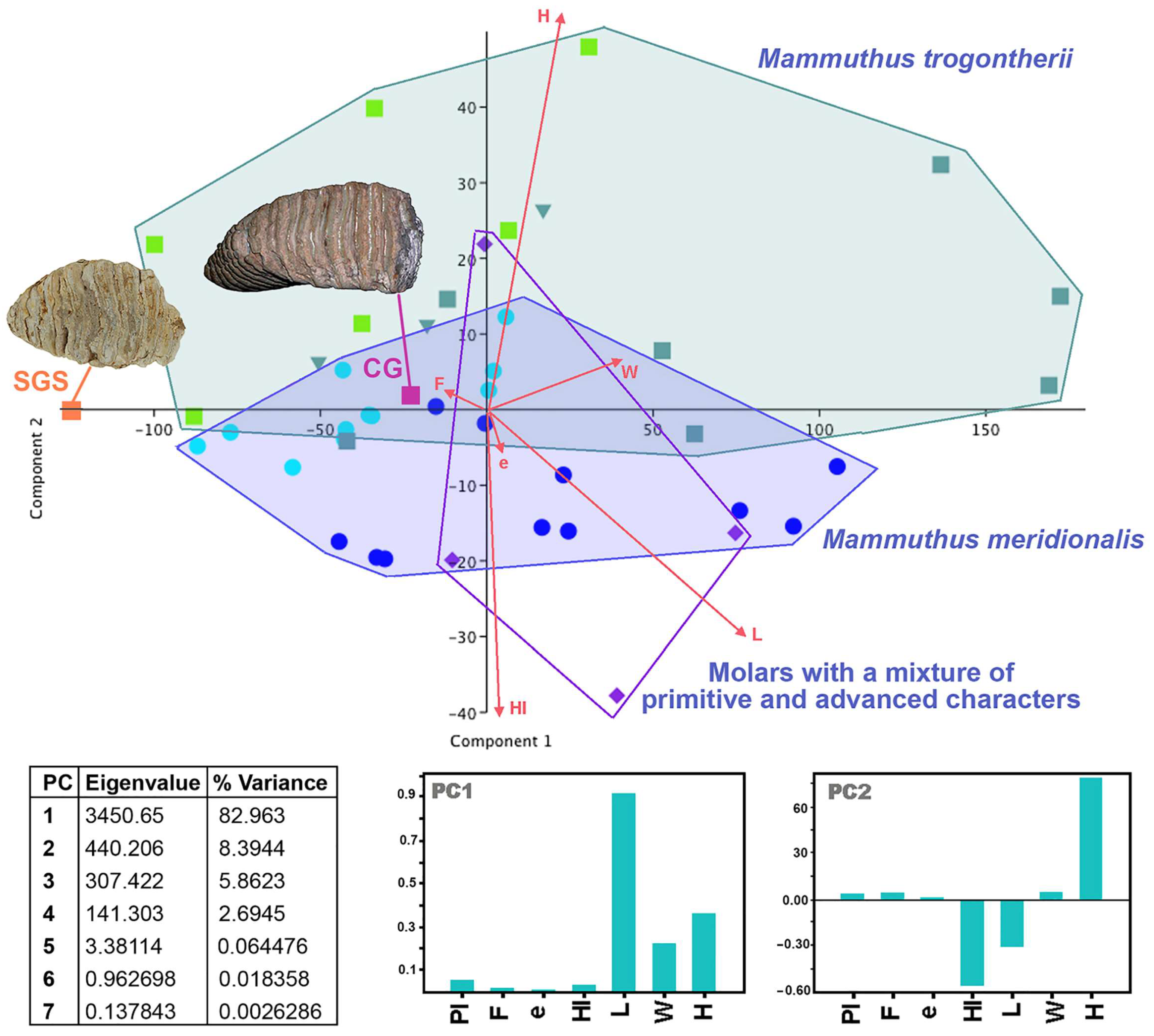
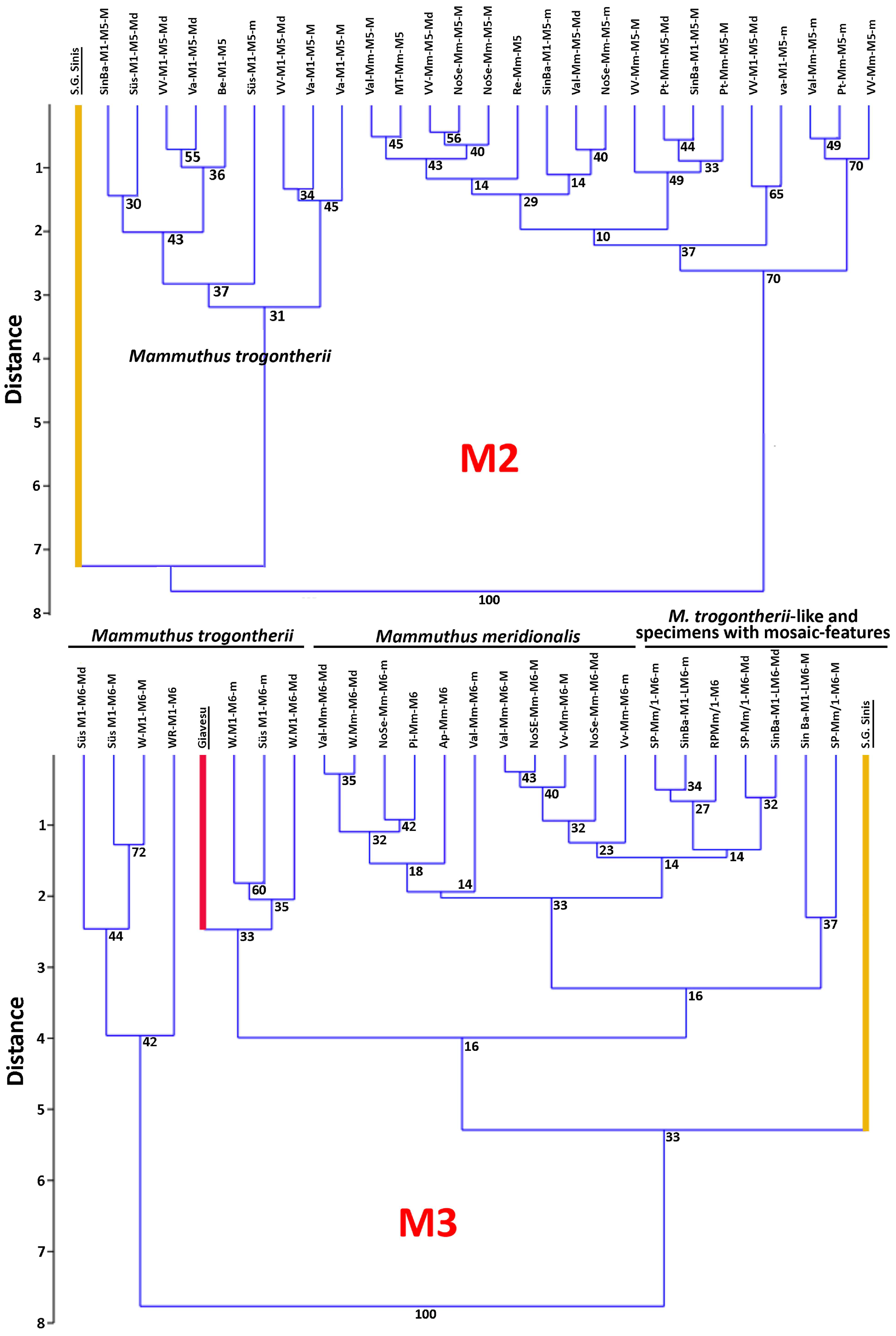
4.6. A Few Taxonomic Notes
5. Ongoing and Planned Research
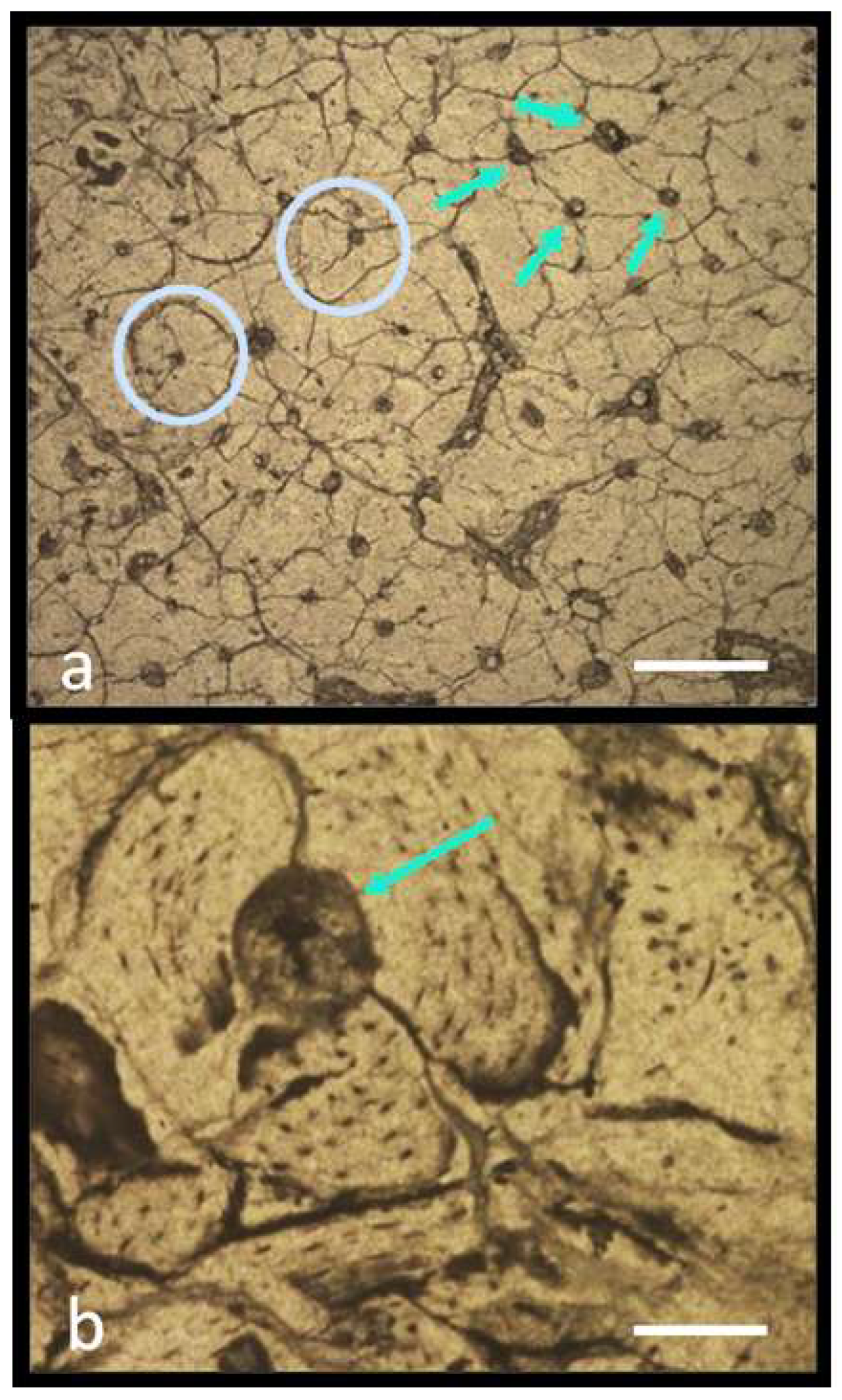
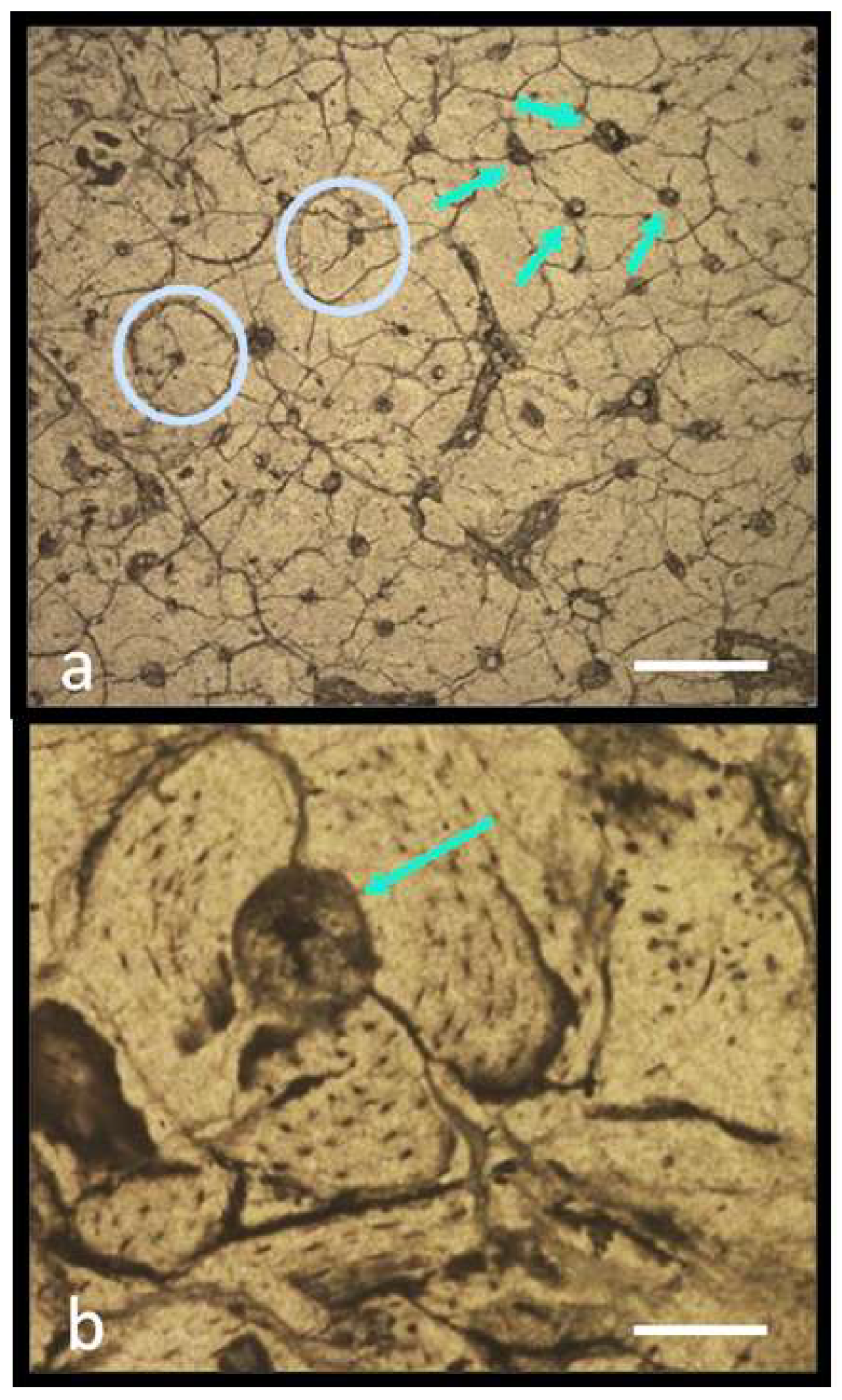
5.1. Histological Analysis
5.1.1. Preliminary Results
5.1.2. Notes
5.2. Planned Research
5.2.1. Geochronology
5.2.2. Paleoecology
6. Open Questions
7. Concluding Remarks
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CG | Campu Giavesu (Giave, SS) |
| FM | Funtana Morimenta spring (Gonnesa, SU) |
| GPH | Guardia Pisano Hill (Gonnesa, SU) |
| MPAS | Museo dei Paleoambienti Sulcitani—“E.A. Martel” (Carbonia) |
| MDLCA | Museo Sardo di Geologia e Paleontologia “D. Lovisato” (Cagliari) |
| NHMP | Natural History Museum of Pisa University (Calci, Pisa) |
| SGS | San Giovanni di Sinis (Cabras, OR) |
| HC.Dm | Haversian canal diameter |
| On.Dm | Osteon diameter |
References and Notes
- Christy, C. The African Elephant: Part II. J. R. Afr. Soc. 1922, 21, 187–198. [Google Scholar] [CrossRef]
- Sondaar, P.Y. Insularity and Its Effect on Mammal Evolution. In Major Patterns in Vertebrate Evolution; Hecht, M., Goody, P.L., Hecht, B.M., Eds.; Plenum Press: New York, NY, USA, 1977; pp. 671–707. [Google Scholar]
- Johnson, D.L. Problems in the land vertebrate zoogeography of certain islands and the swimming powers of elephants. J. Biogeogr. 1980, 7, 383–398. [Google Scholar] [CrossRef]
- Van Hoven, W.; Prins, R.A.; Lankhorst, A. Fermentative digestion in the African elephant. S. Afr. J. Wildl. Res. 1981, 11, 78–88. [Google Scholar]
- Plotnik, J.M.; Shaw, R.C.; Brubaker, D.L.; Tiller, L.N.; Clayton, N.S. Thinking with their trunks: Elephants use smell but not sound to locate food and exclude nonrewarding alternatives. Anim. Behav. 2014, 88, 91–98. [Google Scholar] [CrossRef]
- Palombo, M.R. How can endemic proboscideans help us understand the “island rule”? A case study of Mediterranean islands. Quat. Int. 2007, 169, 105–124. [Google Scholar] [CrossRef]
- Herridge, V.L. Dwarf Elephants on Mediterranean Islands: A Natural Experiment in Parallel Evolution. Ph.D. Thesis, London University College, London, UK, 2010; pp. 1–480. [Google Scholar]
- Van der Geer, A.; Lyras, G.; de Vos, J. History of Island. In Evolution of Island Mammals: Adaptation and Extinction of Placental Mammals on Islands; Wiley-Blackwell: Oxford, UK, 2021. [Google Scholar]
- Palombo, M.R.; Moncunill-Solé, B. Dwarfing and Gigantism in Quaternary Vertebrates. In Encyclopedia of Quaternary Science, 3rd ed.; Elias, S.A., Mock, C., Eds.; Elsevier: Oxford, UK, 2023; pp. 1–25. [Google Scholar] [CrossRef]
- Herridge, V.L.; Lister, A.M. Extreme insular dwarfism evolved in a mammoth. Proc. R. Soc. B 2012, 279, 3193–3200. [Google Scholar] [CrossRef] [PubMed]
- Rick, T.C.; Hofman, C.A.; Braje, T.J.; Maldonado, J.E.; Sillett, T.S.; Danchisko, K.; Erlandson, J.M. Flightless ducks, giant mice and pygmy mammoths: Late Quaternary extinctions on California’s Channel Islands. World Archaeol. 2012, 44, 3–20. [Google Scholar] [CrossRef]
- Palombo, M.R.; Zedda, M.; Melis, R.T. A new elephant fossil from the late Pleistocene of Alghero: The puzzling question of Sardinian dwarf elephants. C. R. Palevol 2017, 16, 841–849. [Google Scholar] [CrossRef]
- Agenbroad, L.D. Giants and pygmies: Mammoths of Santa Rosa island, California (USA). Quat. Int. 2012, 255, 2–8. [Google Scholar] [CrossRef]
- Athanassiou, A.; Van der Geer, A.A.; Lyras, G.A. Pleistocene insular Proboscidea of the Eastern Mediterranean: A review and update. Quat. Sci. Rev. 2019, 218, 306–321. [Google Scholar] [CrossRef]
- Palombo, M.R.; Antonioli, F.; Di Patti, C.; Valeria, L.P.; Scarborough, M.E. Was the dwarfed Palaeoloxodon from Favignana Island the last endemic Pleistocene elephant from the western Mediterranean islands? Hist. Biol. 2021, 33, 2116–2134. [Google Scholar] [CrossRef]
- Garutt, V.E.; Avenanov, A.O.; Vartanyan, S.L. On the systematic position of Holocene Dwarf Mammoths, Mommuthus primigenius (Blumenbach, 1799) from Wrangel Island (North East Siberia). Dokl. Akad. Nauk 1993, 332, 799–801. (In Russian) [Google Scholar] [PubMed]
- Vartanyan, S.L.; Garutt, V.E.; Sher, A.V. Holocene dwarf mammoths from Wrangel Island in the Siberian. Arct. Nat. 1993, 362, 337–340. [Google Scholar] [CrossRef] [PubMed]
- Vartanyan, S.L.; Arslanov, K.A.; Karhu, J.A.; Possnert, G.; Sulerzhitsky, L.D. Collection of radiocarbon dates on the mammoths (Mammuthus primigenius) and other genera of Wrangel Island, northeast Siberia, Russia. Quat. Res. 2008, 70, 51–59. [Google Scholar] [CrossRef]
- Acconci, L. Sopra alcune ossa fossili di elefanti rinvenute nel Quaternario di Morimenta in Sardegna. Atti Soc. Sci. Nat. Processi Verbali 1881, 2, 266–267. [Google Scholar]
- Major, F.C.J. Die Tyrrhenis: Studien über geographische Verbreitung von Tieren und Pflanzen im westlich Mittelmeergebiet. Kosmos 1883, 13, 81–106. [Google Scholar]
- Palombo, M.R.; Ferretti, M.P.; Pillola, G.L.; Chiappini, L. A reappraisal of the dwarfed mammoth Mammuthus lamarmorai (Major, 1883) from Gonnesa (south-western Sardinia, Italy). Quat. Int. 2012, 255, 15–170. [Google Scholar] [CrossRef]
- Zoboli, D.; Pillola, G.L.; Palombo, M.R. The remains of Mammuthus lamarmorai (Major, 1883) housed in the Naturhistorisches Museum of Basel (Switzerland) and the complete “Skeleton-Puzzle”. Boll. Soc. Paleontol. Ital. 2018, 57, 45–57. [Google Scholar]
- Lister, A.M.; Sher, A.V.; van Essen, H.; Wei, G. The pattern and process of mammoth evolution in Eurasia. Quat. Int. 2005, 126, 49–64. [Google Scholar] [CrossRef]
- Aguirre, E. Revision Sistematica de los Elephantidae, por su morfologia y morfometria dentaria. Estud. Geol. 1968, 24, 109–167. [Google Scholar]
- Maglio, V.J. Origin and evolution of the Elephantidae. Trans. Am. Philos. Soc. 1973, 63, 1–149. [Google Scholar] [CrossRef]
- Lister, A.M. Sexual Dimorphism in the Mammoth Pelvis: An Aid to Gender Determination. In The Proboscidea; Shoshani, J., Tassy, P., Eds.; Oxford University Press: Oxford, UK, 1966; pp. 254–259. [Google Scholar]
- Roth, V.L. Insular Dwarf Elephants: A case Study in Body Mass Estimation and Ecological Inference. In Body Size in Mammalian Paleobiology: Estimation and Biological Implications; Damuth, J., MacFadden, B., Eds.; Cambridge University Press: Cambridge, UK, 1990; pp. 151–179. [Google Scholar]
- Christiansen, P. Body size in proboscideans, with notes on elephant metabolism. Zool. J. Linn. Soc. 2004, 140, 523–549. [Google Scholar] [CrossRef]
- Campione, N.E.; Evans, D.C. A universal scaling relationship between body mass and proximal limb bone dimensions in quadrupedal terrestrial tetrapods. BMC Biol. 2012, 10, 60. [Google Scholar] [CrossRef] [PubMed]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- La Marmora, A. Voyage en Sardaigne. Troisième Partie. Description Géologique et Paléontologique; Delaforest: Paris, France, 1857; pp. 651–704. [Google Scholar]
- Comaschi Caria, I. L’elefante nano del Quaternario di Gonnesa (Sardegna sudoccidentale). Rend. Sem. Fac. Sci. Univ. Cagliari 1965, 35, 1–11. [Google Scholar]
- De Stefani, C. Cenni preliminari sui terreni cenozoici della Sardegna. Rend. Accad. Lincei 1891, 7, 464–467. [Google Scholar]
- Caterini, F. Catalogo dei proboscidiani Pliocenici e Quaternari conservati nel Museo di Geologia dell’Università di Pisa. Mem. Soc. Toscana Sci. Nat. 1923, 35, 195–210. [Google Scholar]
- Vaufrey, R. Les éléphants nains des ȋles méditerranéennes et la question des isthmes Pléistocènes. Arch. L’Inst. Paléontol. Hum. Mémoire 1929, 6, 1–220. [Google Scholar]
- Joleaud, L. Le peuplement de la Corse: Les mammifères. Bull. Soc. Sci. Hist. Nat. Corse 1926, 45, 35–107. [Google Scholar]
- Malatesta, A. Primo dente di elefante fossile rinvenuto in Sardegna. Quaternaria 1954, 1, 97–105. [Google Scholar]
- Ambrosetti, P. L’elefante fossile della Sardegna. Boll. Soc. Geol. Ital. 1972, 91, 127–131. [Google Scholar]
- Osborn, H.F. Proboscidea. A monograph of the discovery, evolution, migration and extinction of the mastodonts and elephants of the world. Vol. II. Stegodontoidea, Elephantoidea. Am. Mus. Press 1942, 27, 805–1676. [Google Scholar]
- Melis, R.T.; Palombo, M.R.; Mussi, M. Mammuthus lamarmorae (Major, 1883) Remains in the Pre-Tyrrhenian Deposits of S. Giovanni di Sinis (Western Sardinia, Italy). In The World of Elephants; Cavarretta, G., Gioia, P., Mussi, M., Palombo, M.R., Eds.; CNR: Rome, Italy, 2001; pp. 481–485. [Google Scholar]
- Palombo, M.R.; Ginesu, S.; Sias, S. Mammuthus lamarmorai (Major, 1883) Remains in the Middle Pleistocene Alluvial Deposits of Campu Giavesu Plain (North-Western Sardinia; Italy). In AAVV, Occasional Paper in Earth Science, Abstracts, Proceedings of the 3rd International Mammoth Conference, Dawson City, YT, Canada, 25–29 May 2003; Paleontology Program, Government of Yukon: Whitehorse, YT, Canada, 2003; pp. 122–125. [Google Scholar]
- Palombo, M.R. Biochronology of the Plio-Pleistocene terrestrial mammals of Sardinia: The state of the art. Hell. J. Geosci. 2006, 41, 47–66. [Google Scholar]
- Maxia, C.; Pecorini, G. Il Quaternario della Sardegna. In Proceedings of the Atti del X Congresso Internazionale di Studi Sardi–Simposio sul Quaternario della Sardegna (1968), Cagliari, Italy, 29 December 1966–6 January 1967; pp. 59–69. [Google Scholar]
- Palombo, M.R.; Ginesu, S.; Melis, R.T.; Sias, S. The endemic elephants from Sardinia: An unsolved problem. Monogr. Soc. Hist. Nat. Balear. 2005, 12, 245–254. [Google Scholar]
- Pillola, G.L.; Zoboli, D. Dwarf mammoth footprints from the Pleistocene of Gonnesa (southwestern Sardinia, Italy). Boll. Soc. Paleontol. Ital. 2017, 56, 57–64. [Google Scholar]
- Pillola, G.L.; Palombo, M.R.; Panarello, A.; Zoboli, D. Tetrapod ichnology in Italy: The state of the art. J. Mediterr. Earth Sci. 2020, 12, 193–212. [Google Scholar]
- Zoboli, D.; Pillola, G.L. New evidences of mammal tracks from the Pleistocene of Gonnesa area (southwestern Sardinia, Italy). J. Mediterr. Earth Sci. 2018, 10, 173–175. [Google Scholar]
- Zoboli, D.; Pillola, G.L. The Funtana Morimenta ichnosite (Sardinia, Italy): A potential geotourist attraction. Geoheritage 2021, 13, 30. [Google Scholar] [CrossRef]
- Zoboli, D. The rich palaeontological heritage of SW Sardinia (Italy), a possible resource for geotourism development. Geoheritage 2023, 15, 41. [Google Scholar] [CrossRef]
- Orrù, P.; Ulzega, A. Geomorfologia costiera e sottomarina della Baia di Funtanamare (Sardegna sud-occidentale). Geogr. Fis. Din. Quat. 1986, 9, 56–67. [Google Scholar]
- Melis, R.; Palombo, M.R.; Mussi, M. The stratigraphic sequence of Gonnesa (SW Sardinia): Palaeoenvironmental, palaeontological and archaeological evidence. BAR Int. Ser. 2002, 1095, 445–453. [Google Scholar]
- Palombo, M.R.; Villa, P. Schreger Lines as Support in the Elephantinae Identification. In The World of Elephants; Cavaretta, G., Gioia, P., Mussi, M., Palombo, M.R., Eds.; Consiglio Nazionale Ricerche: Rome, Italy, 2001; pp. 656–660. [Google Scholar]
- Palombo, M.R.; Villa, P. L’utilità Dell’analisi delle Linee di Schreger nello Studio dei Proboscidati. Atti 3° Conv. Intern. Archeozoologia. In Atti 3° Convegno Nazionale di Archeozoologia (Siracusa, 2000) Collana Boll. Paleontol. It.; Fiore, I., Malerba, G., Chilardi, S., Eds.; Studi di Paleontologia: Rome, Italy, 2007; pp. 35–44. [Google Scholar]
- Scarborough, M.E. Insular Adaptations in the Appendicular Skeleton of Sicilian and Maltese Dwarf Elephants. Unpublished Ph.D. Dissertation, University of Cape Town, Cape Town, South Africa, 2020; 232p. [Google Scholar]
- Agenbroad, L.D. Channel Island (USA) Pygmy Mammoths (Mammuthus exilis) Compared and Contrasted with M. columbi, Their Continental Stock. In The World of Elephants; Cavaretta, G., Gioia, P., Mussi, M., Palombo, M.R., Eds.; Consiglio Nazionale Richerche: Rome, Italy, 2001; pp. 473–475. [Google Scholar]
- Palombo, M.R. Elephas? Mammuthus? Loxodonta? The question of the true ancestor of the smallest dwarfed elephant of Sicily. Deinsea 2003, 9, 273–291. [Google Scholar]
- Larramendi, A. Skeleton of a Late Pleistocene steppe mammoth (Mammuthus trogontherii) from Zhalainuoer, Inner Mongolian Autonomous Region, China. Paläontol. Z. 2015, 89, 229–250. [Google Scholar] [CrossRef]
- Larramendi, A. Shoulder height, body mass, and shape proboscideans. Acta Palaeontol. Pol. 2016, 61, 537–574. [Google Scholar] [CrossRef]
- Romano, M.; Manucci, F.; Antonelli, M.; Rossi, M.A.; Agostini, S.; Palombo, M.R. In vivo restoration and volumetric body mass estimate of Mammuthus meridionalis from Madonna della Strada (Scoppito, Aquila). Riv. Ital. Paleontol. Stratigr. 2022, 128, 559–573. [Google Scholar]
- Palombo, M.R.; Giovinazzo, C. Elephas falconeri from Spinagallo Cave (South-Eastern Sicily, Hyblean Plateau, Siracusa): A preliminary report on brain to body weight comparison. Monogr. Soc. Hist. Nat. Balear. 2005, 12, 255–264. [Google Scholar]
- Palombo, M.R.; Ibba, A.; Fanelli, F. Porto Conte Bay. In Fossil Mammalian Biotas of Sardinia Fieldtrip Guide-Book; Palombo, M.R., Pillola, G.L., Kotsakis, T., Eds.; EuroMam; PubliedilService, Pirri (CA): Cagliari, Italy, 2008; pp. 47–57. [Google Scholar]
- Roth, V.L. Dwarfism and Variability in the Santa Rosa Island Mammoth: An Interspecific Comparison of Limb Bone Sizes and Shapes in Elephants. In Recent Advances in Research on the California Islands; Hochberg, F.G., Ed.; 3rd California Islands Symp.; Santa Barbara Museum of Natural History: Santa Barbara, CA, USA, 1993; pp. 433–442. [Google Scholar]
- Carboni, S.; Lecca, L.; Hillaire-Marcel, C.; Ghaleb, B. MIS 5e at San Giovanni di Sinis (Sardinia, Italy): Stratigraphy, U/Th dating and “eustatic” inferences. Quat. Int. 2014, 328, 21–30. [Google Scholar] [CrossRef]
- Chesi, F.; Delfino, M.; Abbazzi, L.; Carboni, S.; Lecca, L.; Rook, L. New Fossil Vertebrate remains from San Giovanni di Sinis (Late Pleistocene, Sardinia): The last Mauremys (Reptilia, Testudines) in the Central Mediterranean. Riv. Ital. Paleontol. Stratigr. 2007, 113, 287–297. [Google Scholar]
- Lecca, L.; Carboni, S. The Tyrrhenian section of San Giovanni di Sinis (Sardinia): Stratigraphic record, of an irregular single high stand. Riv. Ital. Paleontol. Stratigr. 2007, 113, 509–523. [Google Scholar]
- Cocco, F.; Andreucci, S.; Sechi, D.; Cossu, G.A. Upper Pleistocene tectonics in western Sardinia (Italy): Insights from the Sinis peninsula structural high. Terra Nova 2019, 31, 485–493. [Google Scholar] [CrossRef]
- Sher, A.V.; Garutt, V.E. New data on the morphology of elephant molars. Trans. USSR Acad. Sci. Earth Sci. Sect. 1987, 285, 195–199. [Google Scholar]
- Agenbroad, L.D. New absolute dates and comparisons for California’s Mammuthus exilis. Deinsea 2003, 9, 1–16. [Google Scholar]
- Palombo, M.R.; Ferretti, M.P. Elephant fossil record from Italy: Knowledge, problems, and perspectives. Quat. Int. 2005, 126–128, 107–136. [Google Scholar] [CrossRef]
- Ferretti, M.P. Mammuthus meridionalis (Mammalia, Proboscidea, Elephantidae) from the ‘‘Sabbie Gialle’’ of Oriolo (Cava La Salita, Faenza, Northern Italy) and other European late populations of southern mammoth. Eclogae Geol. Helv. 1999, 92, 503–515. [Google Scholar]
- Maccagno, A.M. L’Elephas meridionalis Nesti di Contrada “Madonna della Strada” Scoppito (L’Aquila). Atti Accad. Sci. Fis. E Mat. Napoli Mem. 1962, 4, 1–132. [Google Scholar]
- van Essen, J.A. Tracing Transitions: An Overview of the Evolution and Migrations of the Genus Mammuthus Brookes, 1828 (Mammalia, Proboscidea). Ph.D. Dissertation, Leiden University, Leiden, The Netherlands, 2011. [Google Scholar]
- Konidaris, G.E.; Kostopoulos, D.S.; Koufos, G.D. Mammuthus meridionalis (Nesti, 1825) from Apollonia-1 (Mygdonia Basin, Northern Greece) and its importance within the Early Pleistocene mammoth evolution in Europe. Geodiversitas 2020, 42, 69–91. [Google Scholar] [CrossRef]
- Albayrak, E.; Lister, A.M. Dental remains of fossil elephants from Turkey. Quat. Int. 2012, 276, 198–211. [Google Scholar] [CrossRef]
- Agostini, S.; Palombo, M.R.; Rossi, M.A.; Di Canzio, E.; Tallini, M. Mammuthus meridionalis (Nesti, 1825) from Campo di Pile (L’Aquila, Abruzzo, Central Italy). Quat. Int. 2012, 276, 42–52. [Google Scholar] [CrossRef]
- Coppens, Y.; Beden, M.; Piveteau, J. Mammuthus meridionalis depereti n. subsp. (mammalia, elephantidae): Nouveau mammouth du Pléistocene inférieur d’Europe occidentale. C. R. Acad. Sci. 1982, 294, 291–294. [Google Scholar]
- Dubrovo, I.A.; Palombo, M.R. Personal unpublished data collected in the last decade of the 20th century by Dubrovo, I.A. and Palombo, M.R. during their collaboration founded by CNR and Russian Academy of Sciences.
- Lister, A.M.; Dimitrijević, V.; Marković, Z.; Knežević, S.; Mol, D. A skeleton of ‘steppe’ mammoth (Mammuthus trogontherii (Pohlig)) from Drmno, near Kostolac, Serbia. Quat. Int. 2012, 276, 129–144. [Google Scholar] [CrossRef]
- Dubrovo, I.A. A history of elephants of the Archidiskodon-Mammuthus phylogenetic line on the territory of the USSR. J. Palaeontol. Soc. India 1977, 20, 33–40. [Google Scholar]
- Lister, A.M.; Stuart, A.J. The West Runton mammoth (Mammuthus trogontherii) and its evolutionary significance. Quat. Int. 2010, 228, 180–209. [Google Scholar] [CrossRef]
- Athanassiou, A. A skeleton of Mammuthus trogontherii (Proboscidea, Elephantidae) from NW Peloponnese, Greece. Quat. Int. 2012, 255, 9–28. [Google Scholar] [CrossRef]
- Baygusheva, V.S.; Titov, V.V.; Timonina, G.I. Two skeletons of Mammuthus trogontherii from the Sea of Azov Region. Quat. Int. 2012, 276, 242–252. [Google Scholar] [CrossRef]
- Pawłowska, K.; Greenfield, H.; Czubla, P. Steppe’mammoth (Mammuthus trogontherii) remains in their geological and cultural context from Bełchatów (Poland): A consideration of human exploitation in the Middle Pleistocene. Quat. Int. 2014, 326, 448–468. [Google Scholar] [CrossRef]
- Lenardić, J. Description and morphometric analysis of the Mammuthus armeniacus (Falconer) skull from Slavonski Brod (Croatia). Geol. Croat. 1994, 47, 157–166. [Google Scholar]
- Garutt, V.E.; Foronova, A.V. Issledovanie zubov vmershih slonov (metodicheskie rekomendacii). Akacl. Nauk SSSR, ibirskoe otdelenie, lnstitut geologii i geofiziki press. Novosibirsk 1976, 35, 1–93. [Google Scholar]
- Beccaluva, L.; Deriu, M.; Macciotta, G.P.; Savelli, C. Carta Geopetrografica del Vulcanismo Plio-Pleistocenico della Sardegna nordoccidentale, Scala 1:50.000; L.A.C. Firenze: Florence, Italy, 1981. [Google Scholar]
- Malatesta, A. Cynotherium sardous Studiati, an extinct canid from the Pleistocene of Sardinia. Mem. Ist. Ital. Paleontol. Um. 1970, 1, 1–72. [Google Scholar]
- Lister, A.M.; Ioysey, K.A. Scaling Effects in Elephant Dental Evolution—The Example of Eurasian Mammuthus. In Structure, Function and Evolution of Teeth; Smith, P., Tchernov, E., Eds.; Freund Publishing House Ltd.: Jerusalem, Israel; London, UK; Tel Aviv, Israel, 1992; pp. 185–213. [Google Scholar]
- Andreucci, S.; Clemmensen, L.B.; Murray, A.S.; Pascucci, V. Middle to late Pleistocene coastal deposits of Alghero, Northwest Sardinia (Italy): Chronology and evolution. Quat. Int. 2012, 222, 3–16. [Google Scholar] [CrossRef]
- Sechi, D. Reconstruction of the Sardinia the North-West Coast Evolution: A Chronologic and Sedimentologic Approach. Ph.D. Dissertation, University of Sassari, Sassari, Italy, 2013. [Google Scholar]
- Ferretti, M.P. The dwarf elephant Palaeoloxodon mnaidriensis from Puntali Cave, Carini (Sicily; late middle Pleistocene): Anatomy, systematics and phylogenetic relationships. Quat. Int. 2008, 182, 90–108. [Google Scholar] [CrossRef]
- Theodorou, G.E. The Dwarf Elephants of the Charkadio Cave on the Island of Tilos (Dodekanese, Greece). Ph.D. Dissertation, University of Athens, Athens, Greece, 1983. [Google Scholar]
- Theodorou, G.; Symeonidis, N.; Stathopoulou, E. Elephas tiliensis n. sp. from Tilos island (Dodecanese, Greece). Hell. J. Geosci. 2007, 42, 19–32. [Google Scholar]
- Palombo, M.R.; Zedda, M. New evidence for the presence of endemic elephants from the late Pleistocene of Alghero (Nortwestern Sardinia, Italy). Alp. Mediterr. Quat. 2020, 33, 107–114. [Google Scholar]
- Skedros, J.G.; Knight, A.N.; Clark, G.C.; Crowder, C.M.; Dominguez, V.M.; Qiu, S.; Mulhern, D.M.; Donahue, S.W.; Busse, B.; Hulsey, B.I.; et al. Scaling of Haversian canal surface area to secondary osteon bone volume in ribs and limb bones. Am. J. Phys. Anthropol. 2013, 151, 230–244. [Google Scholar] [CrossRef] [PubMed]
- Giua, S.; Farina, V.; Cacchioli, A.; Ravanetti, F.; Carcupino, M.; Novas, M.M.; Zedda, M. Comparative histology of the femur between mouflon (Ovis aries musimon) and sheep (Ovis aries aries). J. Biol. Res. 2014, 87, 74–77. [Google Scholar] [CrossRef]
- Zedda, M.; Brits, D.; Giua, S.; Farina, V. Distinguishing domestic pig femora and tibiae from wild boar through microscopic analyses. Zoomorphology 2019, 138, 159–170. [Google Scholar] [CrossRef]
- Babosová, R.; Zedda, M.; Belica, A.; Golej, M.; Chovancová, G.; Kalaš, M.; Vondráková, M. The enrichment of knowledge about the microstructure of brown bear compact bone tissue. Eur. Zool. J. 2022, 89, 615–624. [Google Scholar] [CrossRef]
- Kolb, C.; Scheyer, T.M.; Veitschegger, K.; Forasiepi, A.M.; Amson, E.; Van deer Geer, A.A.E.; Van den Hoek Ostende, L.W.; Hayashi, S.; Sánchez-Villagra, M.R. Mammalian bone palaeohistology: A survey and new data with emphasis on island forms. PeerJ 2015, 10, e1358. [Google Scholar] [CrossRef] [PubMed]
- Zedda, M.; Sathé, V.; Chakraborty, P.; Palombo, M.R.; Farina, V. A first comparison of bone histomorphometry in extant domestic horses (Equus caballus) and a Pleistocene Indian wild horse (Equus namadicus). Integr. Zool. 2020, 15, 448–460. [Google Scholar] [CrossRef]
- Palombo, M.R.; Zedda, M. The intriguing giant deer from the Bate cave (Crete): Could paleohistological evidence question its taxonomy and nomenclature? Integr. Zool. 2022, 17, 54–77. [Google Scholar] [CrossRef]
- Ezra, H.C.; Cook, S.F. Histology of Mammoth Bone. Science 1959, 20, 465–466. [Google Scholar] [CrossRef]
- Curtin, A.J.; Macdowell, A.A.; Schaible, E.G.; Roth, V.L. Noninvasive histological comparison of bone growth patterns among fossil and extant neonatal elephantids using synchrotron radiation X-ray microtomography. J. Vertebr. Paleontol. 2012, 32, 939–955. [Google Scholar] [CrossRef]
- Rogoz, A.; Sawlowicz, Z.; Wojtal, P. Diagenetic history of wolly mammoth (Mammuthus primigenius) skeletal remains from the archeological site Cracow Spadzista street B, Southern Poland. Palaios 2012, 27, 541–549. [Google Scholar] [CrossRef]
- Papageorgopoulou, C.; Link, K.; Rühli, F.J. Histology of a Woolly Mammoth (Mammuthus primigenius) Preserved in Permafrost, Yamal Peninsula, Northwest Siberia. Anat. Rec. 2015, 298, 1059–1071. [Google Scholar] [CrossRef]
- Köhler, M.; Herridge, V.; Nacarino-Meneses, C.; Fortuny, J.; Moncunill-Solé, B.; Rosso, A.; Sanfilippo, R.; Palombo, M.R.; Moyà-Solà, S. Palaeohistology reveals a slow pace of life for the dwarfed Sicilian elephant. Sci. Rep. 2021, 11, 22862. [Google Scholar] [CrossRef]
- Basilia, P.; Miszkiewicz, J.; Nganvongpanit, K.; Zaim, J.; Rizal, Y.; Aswan, A.; Puspaningrum, M.R.; Trihascaryo, A.; Price, G.J.; van der Geer, A.A.E.; et al. Bone histology in a fossil elephant (Elephas maximus) from Pulau Bangka, Indonesia. Hist. Biol. 2022, 35, 1356–1367. [Google Scholar] [CrossRef]
- Zedda, M. The Arrangement of the Osteons and Kepler’s Conjecture. Appl. Sci. 2023, 13, 5170. [Google Scholar] [CrossRef]
- Cook, S.F.; Brooks, S.T.; Ezra-Cohn, H.E. Histological studies on fossil bones. J. Paleontol. 1962, 36, 483–494. [Google Scholar]
- Nganvongpanit, K.; Siengdee, P.; Buddhachat, K.; Brown, J.L.; Klinhom, S.; Pitakarnnop, T.; Angkawanish, T.; Thitaram, C. Anatomy, histology and elemental profile of long bones and ribs of the Asian elephant (Elephas maximus). Anat. Sci. Int. 2017, 92, 554–568. [Google Scholar] [CrossRef] [PubMed]
- Zedda, M.; Babosovà, R. Does the osteon morphology depend on the body mass? A scaling study on macroscopic and histomorphometric differences between cow (Bos taurus) and sheep (Ovis aries). Zoomorphology 2021, 140, 169–181. [Google Scholar] [CrossRef]
- Pfeiffer, S.; Crowder, C.; Harrington, L.; Brown, M. Secondary osteons and Haversian canals dimensions as behavioral indicators. Am. J. Phys. Anthropol. 2006, 131, 460–468. [Google Scholar] [CrossRef] [PubMed]
- Felder, A.A.; Phillips, C.; Cornish, H.; Cooke, M.; Hutchinson, J.R.; Doube, M. Secondary osteons scale allometrically in mammalian humerus and femur. R. Soc. Open Sci. 2017, 4, 170431. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, Y.; Janis, C.M.; Goodall, R.H.; Purnell, M.A. An examination of feeding ecology in Pleistocene proboscideans from southern China (Sinomastodon, Stegodon, Elephas), by means of dental microwear texture analysis. Quat. Int. 2017, 445, 60–70. [Google Scholar] [CrossRef]
- Mainland, I.L. Dental microwear in grazing and browsing Gotland sheep (Ovis aries) and its implications for dietary reconstruction. J. Archaeol. Sci. 2003, 30, 1513–1527. [Google Scholar] [CrossRef]
- Palombo, M.R.; Albayrak, E.; Marano, F. The Straight-Tusked Elephants from Neumark-Nord, a Glance into a Lost World. In Elefantenreich. Eine Fossilwelt in Europa. Landesamt für Denkmalpflege und Archaologie Sachsen; Meller, H., Ed.; Anhalt Landesmuseum für Vorgeschichte: Halle (Saale), Germany, 2010; pp. 218–251. [Google Scholar]
- Fortelius, M.; Solounias, N. Functional characterization of ungulate molars using the abrasion-attrition wear gradient: A new method for reconstructing paleodiets. Am. Mus. Novit. 2000, 3301, 1–36. [Google Scholar] [CrossRef]
- Bocherens, H.; Drucker, D.G. Carbonate Stable Isotopes. Terr. Teeth Bones 2013, 10, 30–44. [Google Scholar]
- Sponheimer, M.; Lee-Thorp, J.A. Oxygen isotopes in enamel carbonate and their ecological significance. J. Archaeol. Sci. 1999, 26, 723–728. [Google Scholar] [CrossRef]
- Van Essen, H. Tooth morphology of Mammuthus meridionalis from the southern bight of the North Sea and from several localities in the Netherlands. Deinsea 2003, 9, 453–511. [Google Scholar]
- Palombo, M.R.; Milli, S. Mammalian fossil record, depositional setting, and sequence stratigraphy in the Middle and Upper Pleistocene of Roman basin. Quat. Ital. J. Quat. Sci. 2011, 23, 243–248. [Google Scholar]
- Palombo, M.R.; Mussi, M.; Agostini, S.; Barbieri, M.; Di Canzio, E.; Di Rita, F.; Fiore, I.; Iacumin, P.; Magri, D.; Speranza, F.; et al. Human peopling of Italian intramontane basins: The early Middle Pleistocene site of Pagliare di Sassa (L’Aquila, central Italy). Quat. Int. 2010, 223, 170–178. [Google Scholar] [CrossRef]
- Bonifay, M.F. Les Praemegaceros du Pléistocène moyen de la Grotte de ’’Escale à Saint-Estève-Janson (Bouches-du-Rhône). Leur intérêt dans le contexte biostratigraphique européen. Bull. Assoc. Franç. Etude Quat. 1981, 18, 109–120. [Google Scholar]
- Álvarez-Posada, C.; Parés, J.M.; Sala, R.; Viseras, C.; Pla-Pueyo, S. New magnetostratigraphic evidence for the age of Acheulean tools at the archaeo-palaeontological site “Solana del Zamborino” (Guadix–Baza Basin, S Spain). Sci. Rep. 2017, 7, 13495. [Google Scholar] [CrossRef] [PubMed]
- Kouvari, M.; van der Geer, A.A. Biogeography of extinction: The demise of insular mammals from the Late Pleistocene till today. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2018, 505, 295–304. [Google Scholar] [CrossRef]
- Kouvari, M.; van der Geer, A.A. Corrigendum to “Biogeography of Extinction: The demise of insular mammals from the Late Pleistocene till today”[PALAEO 505 (15 September 2018) pages 295–304]. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2019, 518, 232–233. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mammuthus lamarmorai from the Guardia Pisano Hill (Gonnesa) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Estimated Body Mass (Regression Equations from Christiansen, 2004) | ||||||||||||
| Variable | a ± 95% CI | b ± 95% CI | r | %SEE | %PE | Bone Measurement (mm) | log(BM) | Estimated Body Mass (Kg) | ||||
| sx | dx | sx | dx | sx | dx | sx + dx/2 | ||||||
| HUMERUS | ||||||||||||
| Maximum length | - | 448 | - | 2.841117567 | - | 694 | 694 | |||||
| Humerus least circumference of diaphysis | −1598 | 2062 | 0.995 | 7.78 | 5.54 | - | 187 | - | 3.0865 | - | 1220 | 1220 |
| Humerus diaphysial diameter in the anteroposterior plane | −0.503 | 2009 | 0.997 | 5.97 | 3.62 | - | 93 | - | 3.4517 | - | 2829 | 2829 |
| Humerus diaphysial diameter in the lateromedial plane | −0.66 | 2124 | 0.989 | 12.21 | 8.56 | - | 76 | - | 3.3348 | - | 2162 | 2162 |
| Humerus width of distal articular surface | −5.29 | 3872 | 0.96 | 24.33 | 16.72 | 115 | 113 | 2.6890 | 2.6595 | 489 | 457 | 473 |
| Humerus medial condyle length | −2554 | 2943 | 0.881 | 43.15 | 28.34 | 90 | 88 | 3.1973 | 3.1686 | 1575 | 1474 | 1525 |
| Humerus medial condyle width | −3202 | 3409 | 0.944 | 29.09 | 18.51 | 58 | 55 | 2.8095 | 2.7309 | 645 | 538 | 592 |
| Humerus lateral condyle length | −2294 | 2867 | 0.904 | 39.34 | 25.43 | 65 | - | 2.9036 | - | 801 | - | 801 |
| Humerus lateral condyle width | −3784 | 3775 | 0.752 | 66.58 | 39.61 | 43 | - | 2.3823 | - | 241 | - | 241 |
| Average BM | 750.20 | 1339.22 | 1286.92 | |||||||||
| Average BM (selected measurements) | 877.45 | 876.65 | 877.05 | |||||||||
| FEMUR | ||||||||||||
| Femur maximum length | −5568 | 3036 | 0.985 | 14.54 | 6.15 | - | - | - | - | - | - | - |
| Femur least esircumference of diaphysis | −1606 | 2073 | 0.976 | 18.46 | 11.52 | - | 189 | - | 3.1131 | - | 1297 | - |
| Femur diaphysial diameter in the anteroposterior plane | −0.912 | 2315 | 0.98 | 16.64 | 11.4 | - | 42 | - | 2.8458 | - | 701 | - |
| Femur diaphysial diameter in the lateromedial plane | −0.342 | 1904 | 0.966 | 22.23 | 14.42 | - | 74 | - | 3.2170 | - | 1648 | - |
| Humerus width of distal articular surface | −4347 | 3502 | 0.928 | 33.49 | 21.71 | - | 101 | - | 2.6721 | - | 470 | - |
| Femur medial condyle length | −0.819 | 2156 | 0.862 | 23.42 | 15.35 | - | 48 | - | 2.8058 | - | 639 | - |
| Femur medial condyle width | −1.55 | 2619 | 0.95 | 27.39 | 17.71 | - | 99 | - | - | - | - | - |
| Femur lateral condyle length | −4.97 | 4345 | 0.833 | 53.49 | 30.98 | - | 53 | - | 2.5220 | - | 333 | - |
| Femur lateral condyle width | −1514 | 2695 | 0.987 | 13.21 | 9.91 | - | 118 | - | - | - | - | - |
| AVERAGE BM | 848.16 | |||||||||||
| AVERAGE BM (selected measurements) | 951.26 | |||||||||||
| Taxon | Site | Tooth | Plates (Pl) | Length | Width | Crown Height | Lamellar Frequency | Enamel Thickness | Hypsodonty Index (H/W) | Refs: | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Formula | Total (Pl) | In Use (PlF) | Total (L) | Occlusal (LF) | Crown (W) | Occlusal (WF) | Crown (H) | Functional (HF) | Average (F) | Average (e) | ||||||||||||
| Mean | Range | Mean | Range | Mean | Range | Mean | Range | Mean | Range | Mean | Range | Mean | Range | |||||||||
| M. lamarmorai | San Givanni di Sinis | M3 (?M2) | ∞11× | >11 | 8 | 130 | 116 | 69 | 55 | >90 | 90 | 8 | 1.8 | >1.55 | [35] | |||||||
| Campu Giavesu | M3 | 15 p | 15 | 7 | 225 | c. 82 | 76.5 | - | 129 | 129 | 7 | 2.8 | 1.69 | [36] | ||||||||
| M. cf. M. lamarmorai | Tramariglio | M1? | ∞7× * | >7 * | 7 * | >97 | - | 57 | - | - | - | 7 * | 2–3 | - | [32] | |||||||
| M. exilis | Santa Rosa Island (California, US) | M3 | - | 23.2 | 20–27 | - | 263.8 | 226–285 | - | 92.5 | 68–113 | - | 168.1 | 135–188.5 | - | 9.0 | 6.5–11-1 | 1.6 | 1.3–2-0 | 2.8 | - | [63] |
| M. meridionalis | Valdarno (IT) | M3 | - | 13 | 11–14 | - | 269 | 226–343 | - | 101 | 83–123 | - | 131 | 101–160 | - | 5 | 4.0–6.0 | 3.2 | 2.6–3.9 | 1.3 | 1.2–1.6 | [64] |
| Various sites | - | 12.8 | 11–14 | - | 273 | 223–317 | - | 104.8 | 86–126 | - | 122.7 | 100–142 | - | 4.9 | 3.7–6.1 | 3.3 | 2.6–4.1 | [20] | ||||
| Montopoli (IGF 1077) (IT) | - | 9 | - | - | 263 | - | - | 109 | - | - | - | - | - | 4 | - | 3.8 | - | - | - | [65] | ||
| Pietrafitta (IT) | - | 14 | 13–16 | - | 245 | - | - | 93 | 84–103 | - | 131 | - | - | 6 | - | 2.9 | 2.6–3.3 | 1.5 | - | [65] | ||
| Pietrafitta (CET80) (IT) | - | 14 | - | - | 318 | - | - | 115 | - | - | 130 | - | - | 5 | - | 3.3 | - | 1.32 | - | [64] | ||
| Pietrafitta (CET1119) (IT) | - | 14 | - | - | 245 | - | - | 103 | - | - | 131 | - | - | 6 | - | 2.6 | - | 1.5 | - | [64] | ||
| Farneta (IGF12434) (IT) | - | 14 | - | - | 301 | - | - | 115 | - | - | 134.5 | - | - | 5 | - | 2.9 | - | 1.17 | - | [64] | ||
| Madonna della Strada (right) (IT) | - | >7 | - | - | 255 | - | - | 98 | - | - | 110 | - | - | 5 | - | - | - | 1.12 | - | [66] | ||
| Madonna della Strada (left) (IT) | - | >8 | - | - | 258 | - | - | 95 | - | - | - | - | - | 5 | - | - | - | - | - | [66] | ||
| North Sea | - | 13.3 | 13–14 | - | 265.8 | 220–335 | - | 101.2 | 840–117.5 | - | 120.8 | 104–147 | - | 5.4 | 4.9–5.8 | 3.96 | 2.3–3.8 | 1.31 | 1.23–1.49 | [67] | ||
| Apollonia (APL-686B) (AL) | - | >13 (?14) | - | - | 231 | - | - | c. 87 | - | - | 128.8 | - | - | 5.5 | - | 2.9 | - | 1.48 | - | [68] | ||
| EskişehireYukarısögütönü (TR) | - | 12 | - | 309.5 | - | 119.5 | - | 134.1 | - | c. 3.9 | 3.67 | 1.2 | [69] | |||||||||
| Valdarno (IT) | M2 | - | 9 | 8–10 | - | 212 | 175–243 | - | 89 | 80–103 | - | 120 | 103–148 | - | 5.0 | 4.5–6.0 | 2.7 | 2.2–3.4 | 1.40 | 1.00–1.60 | [64] | |
| Various sites | - | 9.7 | 8–11 | - | 206.7 | 167–242 | - | 86.8 | 76–106 | - | 113.9 | 98–138 | - | 5.1 | 4.0–5.7 | 2.9 | 1.9–3.6 | [20] | ||||
| Pietrafitta (IT) | - | 11 | 8–12 | - | - | - | - | 84 | 73–95 | - | - | - | - | 5.4 | 4.8–5.8 | 2.7 | 2.6–2.9 | 1.3 | 1.2–1.4 | [65] | ||
| Campo di Pile (SBAAp 320) (IT) | - | 9 | - | - | 206 | - | - | 90 | - | - | 94 | - | - | 4.5 | - | 3.1 | - | 1.04 | - | [70] | ||
| Campo di Pile (SBAAp 321) (IT) | - | 9 | - | - | 197 | - | - | 92 | - | - | 91 | - | - | 4.5 | - | 3.1 | - | 0.99 | - | [70] | ||
| Redicicoli (MR E2) (IT) | - | 9 | - | - | 201 | - | - | 94 | - | - | 123 | - | - | 6.0 | - | 2.6 | - | 1.30 | - | [64]] | ||
| Monte Tenda (MCSN 10688) (IT) | - | 10 | - | - | 206 | - | - | 90 | - | - | 115 | - | - | 6.0 | - | 3.0 | - | 1.28 | - | [63] | ||
| North Sea | - | 9.7 | 9–10 | - | 212.3 | 195–242 | - | 92.2 | 79–104.5 | - | 119.7 | 105–135 | - | 5.5 | 4.5–6.3 | 2.72 | 2.2–3.3 | 1.34 | 1.29–1.51 | [66] | ||
| Konya—Zengen (TR) | - | 8 | - | 202.7 | - | 87.7 | - | c.107.1 | - | c.5.3 | 2.97 | c.1.22 | [68] | |||||||||
| Specimens showing a mosaic of advanced and primitive characters | Rio Pradella (IT) | M3 | - | 14 | - | - | 235 | - | - | 90 | - | - | 158 | - | - | 6 | - | 2.4 | - | 1.76 | - | [64] |
| Saint Prest (FR) | - | 14–15 | - | - | - | 245–312 | - | - | 85–105 | - | - | 108–141 | - | - | 3.8–6.0 | - | 2.4–3.7 | - | 1.11–1.40 | [67,71] | ||
| Saint Prest (FR) | M2 | 10 | - | - | - | 175–215 | - | - | 74–102 | - | 85 | - | - | - | 3.6–5.7 | - | 2.3–3.5 | - | - | [67,71] | ||
| M. trogontherii-like | Sinyaya Balka (RO) | M3 | - | 15 | 13–17 | - | 194.9 | 254–328 | - | 98.5 | 82–122 | - | - | - | - | 4.8 | 4.0–6.0 | 3.1 | 2.5–3.5 | 1.58 | 1.25–1.42 | [72] |
| Sinyaya Balka (RO) | M2 | - | 11.4 | 10–14 | - | 225.7 | 195–245 | - | 77.7 | 87–116 | 140.9 | 122–166 | - | 5.65 | 4.5–6.5 | 2.7 | 2.3–3.0 | 1.6 | 1.2–1.9 | [72] | ||
| M. trogontherii | Süssenborn (DE) | M3 | - | 19.7 | 17–22 | - | 300 | 230–397 | - | 101.6 | 77–123 | - | 122.0 | 147–210 | - | 9.96 | 5.80–8.94 | 2.39 | 1.7–3.0 | 1.8 | 1.3–2.3 | [65,73] |
| Various sites | - | 18.6 | 14–21 | - | 293 | 213–358 | - | 85.2 | 57–108 | - | 162.5 | 118–218 | - | 6.5 | 5.0–8.2 | 2.2 | 1.5–3.0 | - | - | [20] | ||
| Tiraspol (MD) | - | - | 16–22 | - | 337 | 303–370 | - | - | 86–115 | - | - | 160–211 | - | - | 5.0–7.0 | - | 2.0–3.0 | - | - | [74] | ||
| Via Flaminia Km 7.2 (IT) | - | 18 | - | - | 303 | - | - | 119 | - | - | - | - | - | 7 | - | 2.3 | - | - | - | [64] | ||
| West Runton (GB) | - | 21 | - | - | c. 400 | - | - | c. 118 | - | - | 196 | - | - | 5.25 | - | 2.3 | - | c. 1.66 | - | [75] | ||
| Laussikà (EL) | - | >14 | - | - | 350 | - | - | 102 | - | - | - | - | - | 6.3 | - | - | 2.6–2.9 | - | - | [76] | ||
| Kagal’nik (Azov Sea) (right) (RO) | - | 18 | - | - | 354 | - | - | 104 | - | - | 191 | - | - | 5.5 | - | 2.4 | - | - | [77] | |||
| Kagal’nik (Azov Sea) (left) (RO) | - | 20 | - | - | 372 | - | - | 105 | - | - | 189 | - | - | 5.5 | - | 2.3 | 1.84 | - | - | [77] | ||
| Belchatów (PL) | - | >19 | - | - | 289 | - | - | 114.4 | - | - | - | - | - | 6.75 | - | 2.2 | 157 | - | - | [78] | ||
| Amasya-Suluova (TR) | - | 19 | - | - | 323 | - | - | 109 | - | - | 181.8 | - | - | 6.41 | - | 2.79 | - | 1.68 | - | [69] | ||
| Konya-Dursunlu (TR) | - | 18 | - | - | 295 | - | - | 106.5 | - | - | c.165.4 | - | - | 6.13 | - | 2.92 | - | c.1.52 | - | [69] | ||
| Süssenborn (DE) | M2 | - | 15 | 13–16 | - | - | - | - | 85 | 68–103 | - | - | - | - | 5.6 | 4.4–6.8 | 2.5 | 2.0–2.9 | 1.8 | 1.3–2.3 | [65] | |
| Various sites | - | 13.3 | 11–17 | - | 206.7 | 168–240 | - | 77.4 | 55.0–95.0 | - | 134.3 | 105–162 | - | 6.9 | 5.5–7.8 | 1.8 | 1.3–2.8 | - | - | [20] | ||
| Pratola Peligna (IT) | - | ?13 | - | - | >198 | - | - | 87 | - | - | - | - | - | 8.0 | - | 2.8 | - | 2.0 | - | [64] | ||
| Belchatów (PL) | - | 14 | - | - | 187 | - | - | 77.7 | - | - | - | - | - | 7.5 | - | 1.8 | - | 1.78 | - | [78] | ||
| Slavonski (left) (HR) | - | >11 | - | - | 178 | - | - | 83 | - | - | - | - | - | - | - | 1.42 | - | - | - | [79] | ||
| Slavonski (right) (HR) | - | >11 | - | - | >157 | - | - | 81 | - | - | - | - | - | - | - | 1.34 | - | - | - | [79] | ||
| Various sites | - | 13 | 10–16 | - | - | 144–250 | - | 82 | 70–109 | - | 167 | 124–196 | - | 7.5 | 4.8–12.8 | 2.0 | 1.0–2.65 | - | - | [79,80] | ||
| Mammuthus cf. M. lamarmorai (Alghero) | |||
|---|---|---|---|
| Measurements | Average Diameter | Average Perimeter | Average Eccentricity |
| Osteons | 205 ± 28 µm | 684 ± 145 µm | 0.38 |
| Haversian canals | 41 ± 12 µm | 138 ± 36 µm | 0.34 |
| Species | Measurements | References | ||
|---|---|---|---|---|
| On.Dm (Osteon Diameter) | HC.Dm (Haversian Canal Diameter) | On.Dm/HC.Dm | ||
| Mammuthus cf. M. lamarmorai | 205 ± 28 µm | 41 ± 12 µm | 5.1 | |
| Mammuthus primigenius | 294 ± 83 µm | 56 ± 19 µm | 4.9 | [104] |
| Elephas maximus | 235 ± 59 µm | 74 ± 21 µm | 3.2 | [111] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palombo, M.R.; Zedda, M.; Zoboli, D. The Sardinian Mammoth’s Evolutionary History: Lights and Shadows. Quaternary 2024, 7, 10. https://doi.org/10.3390/quat7010010
Palombo MR, Zedda M, Zoboli D. The Sardinian Mammoth’s Evolutionary History: Lights and Shadows. Quaternary. 2024; 7(1):10. https://doi.org/10.3390/quat7010010
Chicago/Turabian StylePalombo, Maria Rita, Marco Zedda, and Daniel Zoboli. 2024. "The Sardinian Mammoth’s Evolutionary History: Lights and Shadows" Quaternary 7, no. 1: 10. https://doi.org/10.3390/quat7010010
APA StylePalombo, M. R., Zedda, M., & Zoboli, D. (2024). The Sardinian Mammoth’s Evolutionary History: Lights and Shadows. Quaternary, 7(1), 10. https://doi.org/10.3390/quat7010010