
Stage or Sub-Stage: The Contribution of Small Mammals to the Characterization of Middle Pleistocene Local Climate Variation



Abstract
:1. Introduction
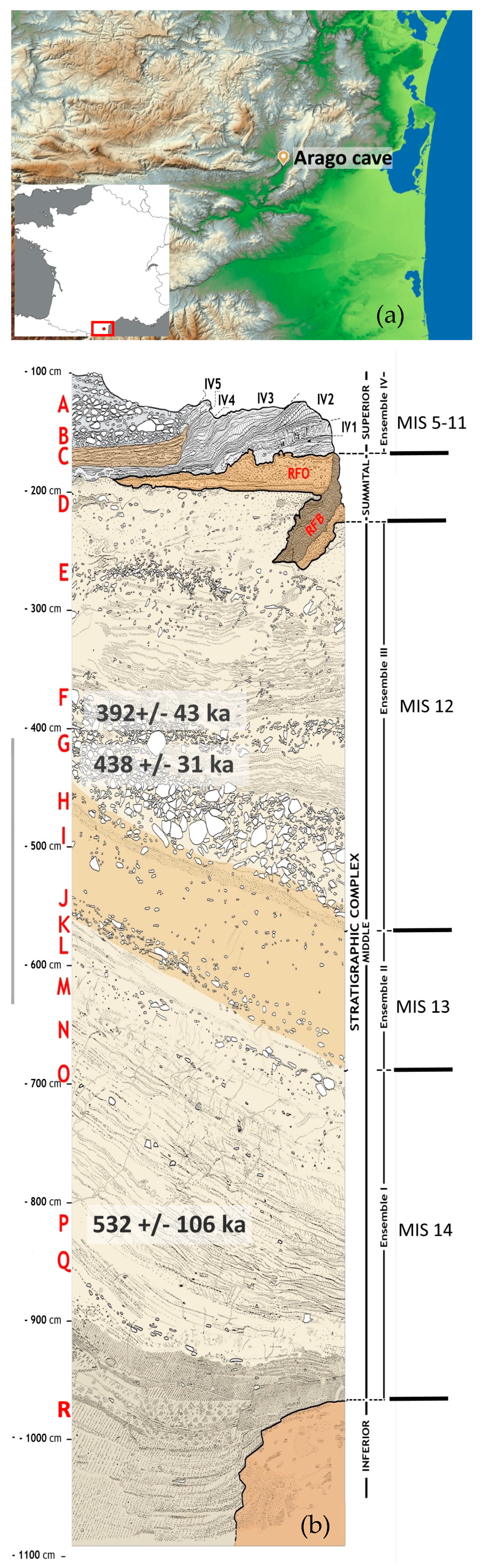
2. Site Presentation
3. Materials and Methods
4. Results
4.1. Correspondence Analysis
4.2. Climatic Parameters
4.3. Synthesis
5. Discussion
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Berger, W.H.; Jansen, E. Mid-pleistocene climate shift—The Nansen Connection. In The Polar Oceans and Their Role in Shaping the Global Environment; Johannessen, O.M., Muench, R.D., Overland, J.E., Eds.; American Geophysical Union: Washington, DC, USA, 2013; Volume 85, pp. 295–311. [Google Scholar]
- Head, M.J.; Gibbard, P.L. Early-Middle Pleistocene transitions: An Overview and Recommendation for the Defining Boundary. In Early-Middle Pleistocene Transitions: The Land-Ocean Evidence; Head, M.J., Gibbard, P.L., Eds.; Geological Society of London: London, UK, 2005; pp. 1–18. [Google Scholar]
- Discamps, E.; Royer, A. Reconstructing palaeoenvironmental conditions faced by Mousterian hunters during MIS 5 to 3 in southwestern France: A multi-scale approach using data from large and small mammal communities. Quat. Int. 2017, 433, 64–87. [Google Scholar]
- Denys, C. Des référentiels en taphonomie des petits vertébrés: Bilan et perspectives. In Taphonomie des Petits Vertébrés: Référentiels et Transferts Aux Fossiles; Laroulandie, V., Mallye, J.-B., Denys, C., Eds.; British Archaeological Reports; International Series 2269; Archaeopress: Oxford, UK, 2011; pp. 7–22. [Google Scholar]
- Puzachenko, A.Y.; Markova, A.K. The Scandinavian ice sheet against the Atlantic ocean: How the scandinavian ice sheet affected European small mammal assemblage during the Greenland stadial GS-2.1. Quat. Sci. Rev. 2023, 305, 108013. [Google Scholar]
- Lumley, H.; Fournier, A.; Park, Y.C.; Yokoyama, Y.; Demouy, A. Stratigraphie du remplissage Pléistocène moyen de la Caune de l’Arago à Tautavel. Etude de huit carottages effectués de 1981 à 1983. L’Anthropologie 1984, 88, 5–18. [Google Scholar]
- Barsky, D.; Lumley, H. Early European Mode 2 and the stone industry from the Caune de l’Arago’s archeostratigraphical levels “P”. Quat. Int. 2010, 223, 71–86. [Google Scholar]
- Barsky, D.; Moigne, A.M.; Pois, V. The shift from typical Western European Late Acheulian to microproduction in unit ‘D’of the late Middle Pleistocene deposits of the Caune de l’Arago (Pyrénées-Orientales, France). J. Hum. Evol. 2019, 135, 102650. [Google Scholar] [PubMed]
- Viallet, C.; Grégoire, S.; Perrenoud, C. Break to Rebuild—The First European Evidence of a Fragmented Chaine Opératoire for Handaxe Production (OIS 14, Caune de l’Arago, France). J. Paleolit. Archaeol. 2022, 5, 1. [Google Scholar]
- Blain, H.A.; Fagoaga, A.; Ruiz-Sánchez, F.J.; Garcia-Medrano, P.; Ollé, A.; Jiménez-Arenas, J.M. Coping with arid environments: A critical threshold for human expansion in Europe at the Marine Isotope Stage 12/11 transition? The case of the Iberian Peninsula. J. Hum. Evol. 2021, 153, 102950. [Google Scholar] [PubMed]
- Connet, N.; Soriano, S.; Bertran, P.; Lhomme, V.; Debenham, N. A 400,000 years old milestone of the Acheulian technocomplex in Central-Western France at Londigny (Charente). J. Archaeol. Sci. Rep. 2020, 30, 102225. [Google Scholar]
- Moncel, M.H.; Despriée, J.; Voinchet, P.; Tissoux, H.; Moreno, D.; Bahain, J.J.; Courcimault, G.; Falguères, C. Early evidence of Acheulean settlement in northwestern Europe-La Noira Site, a 700 000 year-old occupation in the center of France. PLoS ONE 2013, 8, e75529. [Google Scholar]
- Moncel, M.H.; Ashton, N. From 800 to 500 ka in Western Europe. The oldest evidence of Acheuleans in their technological, chronological, and geographical framework. In The Emergence of the Acheulean in East Africa, 1st ed.; Mussi, M., Gallotti, R., Eds.; Springer: New York, NY, USA, 2018; pp. 215–235. [Google Scholar]
- Nicoud, E.; Aureli, D.; Pagli, M.; Villa, V.; Chaussé, C.; Agostini, S.; Zupancich, A. Preliminary data from Valle Giumentina Pleistocene site (Abruzzo, Central Italy): A new approach to a Clactonian and Acheulian sequence. Quat. Int. 2016, 409, 182–194. [Google Scholar]
- Ollé, A.; Lombao, D.; Asryan, L.; García-Medrano, P.; Arroyo, A.; Fernández-Marchena, J.L.; Vallverdú, J. The earliest European Acheulean: New insights into the large shaped tools from the late Early Pleistocene site of Barranc de la Boella (Tarragona, Spain). Front. Earth Sci. 2023, 11, 1188663. [Google Scholar]
- Falguères, C.; Shao, Q.; Han, F.; Bahain, J.J.; Richard, M.; Perrenoud, C.; Moigne, A.M.; Lumley, H. New ESR and U-series dating at Caune de l’Arago, France: A key-site for European Middle Pleistocene. Quat. Geochronol. 2015, 30, 547–553. [Google Scholar]
- Moigne, A.M.; Palombo, M.R.; Belda, V.; Heriech-Briki, D.; Kacimi, S.; Lacombat, F.; Lumley, M.A.; Moutoussamy, J.; Rivals, F.; Quilès, J.; et al. Les faunes de grands mammifères de la Caune de l’Arago (Tautavel) dans le cadre biochronologique des faunes du Pléistocène moyen italien. L’Anthropologie 2006, 110, 788–831. [Google Scholar]
- Magniez, P.; Moigne, A.M.; Testu, A.; Lumley, H. Biochronologie des mammifères quaternaires. Apport des cervidae du site pléistocène moyen de la Caune de l’Arago (Tautavel, Pyrénées-Orientales, France). Quaternaire 2013, 24, 477–502. [Google Scholar]
- Lumley, H. Caune de l’Arago, Tautavel-en-Roussillon, Pyrénées-Orientales, France: Individualisation des Unités Archéostratigraphiques; CNRS Editions: Paris, France, 2015; p. 642. [Google Scholar]
- Lumley, M.A. L’homme de Tautavel. Un Homo erectus européen évolué. Homo erectus tautavelensis. L’Anthropologie 2015, 119, 303–348. [Google Scholar]
- Lebreton, L.; Desclaux, E.; Hanquet, C.; Moigne, A.M.; Perrenoud, C. Environmental context of the Caune de l’Arago Acheulean occupations (Tautavel, France), new insights from microvertebrates in Q-R levels. Quat. Int. 2016, 411, 182–192. [Google Scholar]
- Chaline, J. L’âge des Hominiens de la Caune de l’Arago à Tautavel (Pyrénées-Orientales), d’après l’étude des Rongeurs. Comptes Rendus Hebd. Des Séances De L’académie Des Sci. 1971, 272, 1743–1746. [Google Scholar]
- Brunet-Lecomte, P. Statut des campagnols souterrains (Rodentia, Arvicolidae) du gisement du Pléistocène moyen de l’Arago à Tautavel (Pyrénées-Orientales, France). Bull. Mens. De La Société Linnéenne De Lyon 1990, 59, 100–104. [Google Scholar]
- Desclaux, E. Les petits vertébrés de la Caune de l’Arago (Tautavel, Pyrénées-orientales): Paléontologie, paléoécologie et taphonomie. Ph.D. Thesis, University of Perpignan, Perpignan, France, 1992. [Google Scholar]
- Paunescu, A.C. Les Rongeurs du Pléistocène Inférieur et Moyen de trois grottes du sud-est de la France (Vallonnet, Caune de l’Arago, Baume Bonne). Implications systématiques, biostratigraphiques et paléoenvironnementales. Ph.D. Thesis, University of Perpignan, Perpignan, France, 2001. [Google Scholar]
- Brunet-Lecomte, P.; Paunescu, A. Morphologie comparée de la première molaire inférieure du campagnol Microtus (Terricola) vaufreyi tautavelensis (Rodentia, Arvicolidae) du gisement pléistocène moyen de l’Arago (Pyrénées, France) et inférences paléoclimatiques. Quaternaire 2004, 15, 263–268. [Google Scholar] [CrossRef]
- Hanquet, C. Evolution des paléoenvironnements et des paléoclimats au Pléistocène moyen, en Europe méridionale, d’après les faunes de micromammifères. Ph.D. Thesis, University of Perpignan, Perpignan, France, 2011. [Google Scholar]
- Lebreton, L.; Desclaux, E.; Hanquet, C.; Cuenca-Bescós, G.; Moigne, A.M.; Perrenoud, C.; Grégoire, S. Variations paléoenvironnementales au sein de l’Unité Archéostratigraphique G (UA G) de la Caune de l’Arago (Tautavel, France): Apport des paléocommunautés de rongeurs. Quaternaire 2017, 28, 313–321. [Google Scholar] [CrossRef]
- Lebreton, L.; Moigne, A.M.; Filoux, A.; Perrenoud, C. A specific small game exploitation for Lower Paleolithic: The beaver (Castor fiber) exploitation at the Caune de l’Arago (Pyrénées-Orientales, France). J. Archaeol. Sci. Rep. 2017, 11, 53–58. [Google Scholar]
- Manzano, A. Les Amphibiens et les Reptiles des Sites du Pléistocène Moyen et Supérieur du Pourtour Méditérranéen (Caune de l’Arago, Grotte du Lazaret, Baume Moula-Guercy). Etude D’herpétofaunes et Reconstitutions Paléoclimatiques et Paléoenvironnementales. Ph.D. Thesis, University of Perpignan, Perpignan, France, 2015. [Google Scholar]
- Lebreton, L. Approche taphonomique multi-taxons des accumulations de petits vertébrés, implication pour les reconstitutions paléoenvironnementales au Pléistocène. Ph.D. Thesis, University of Perpignan, Perpignan, France, 2018. [Google Scholar]
- Marquet, J.C. Paléoenvironnement et Chronologie des Sites du Domaine Atlantique Français d’âge Pléistocène Moyen et Supérieur D’après L’étude des Rongeurs. Ph.D. Thesis, University of Dijon, Dijon, France, 1989. [Google Scholar]
- Chaline, J.; Brunet-Lecomte, P.; Campy, M. The last glacial/interglacial record of rodent remains from the Gigny karst sequence in the French Jura used for palaeoclimatic and palaeoecological reconstructions. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1995, 117, 229–252. [Google Scholar]
- Foury, Y.; Desclaux, E.; Daujeard, C.; Defleur, A.; Moncel, M.H.; Raynal, J.P. Evolution des faunes de rongeurs en moyenne vallée du Rhône (rive droite, Ardèche, France) au cours du pléistocène moyen final et du pléistocène supérieur ancien, du mis 6 au mis 4. Quaternaire 2016, 27, 55–79. [Google Scholar]
- Hernández Fernández, M. Bioclimatic discriminant capacity of terrestrial mammal faunas. Glob. Ecol. Biogeogr. 2001, 10, 189–204. [Google Scholar]
- Hernández Fernández, M.; Peláez-Campomanes, P. The bioclimatic model: A method of palaeoclimatic qualitative inference based on mammal associations. Glob. Ecol. Biogeogr. 2003, 12, 507–517. [Google Scholar]
- Hernández Fernández, M.; Peláez-Campomanes, P. Quantitative palaeoclimatic inference based on terrestrial mammal faunas. Glob. Ecol. Biogeogr. 2005, 14, 39–56. [Google Scholar]
- Fernández, M.H.; Sierra, M.Á.; Peláez-Campomanes, P. Bioclimatic analysis of rodent palaeofaunas reveals severe climatic changes in Southwestern Europe during the Plio-Pleistocene. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2007, 251, 500–526. [Google Scholar]
- Royer, A.; Yelo, B.A.G.; Laffont, R.; Fernández, M.H. New bioclimatic models for the quaternary palaearctic based on insectivore and rodent communities. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2020, 560, 110040. [Google Scholar]
- Jeannet, M. L’écologie quantifiée. Essai de description de l’environnement continental à l’aide des microvertébrés. Préhistoires méditerranéennes 2010, 1, 1–26. [Google Scholar]
- Gamisch, A. Oscillayers: A dataset for the study of climatic oscillations over Plio-Pleistocene time-scales at high spatial-temporal resolution. Glob. Ecol. Biogeogr. 2019, 28, 1552–1560. [Google Scholar]
- Lebreton, L.; Bailon, S.; Guillaud, E.; Testu, A.; Perrenoud, C. Multi-taxa referential of a modern Eurasian Eagle-Owl (Bubo bubo) aerie. J. Archaeol. Sci. Rep. 2020, 32, 102417. [Google Scholar]
- Hanquet, C.; Desclaux, E. Analyse paléoécologique des communautés de micromammifères de la Caune de l’Arago (Tautavel, France) dans le contexte des migrations de faunes en Europe méridionale au cours du Pléistocène moyen. Quaternaire 2011, 22, 34–45. [Google Scholar]
- Guthrie, D.; van Kolfschoten, T. Neither warm and moist, nor cold and arid: The ecology of the Mid Upper Palaeolithic. In Hunters of the Golden Age; Roebroeks, W., Mussi, M., Svoboda, J., Fennema, K., Eds.; Leiden University: Leiden, The Netherlands, 2000; pp. 3–20. [Google Scholar]
- Bennett, K.D.; Provan, J. What do we mean by ‘refugia’? Quat. Sci. Rev. 2008, 27, 2449–2455. [Google Scholar]
- Stewart, J.R.; Lister, A.M.; Barnes, I.; Dalén, L. Refugia revisited: Individualistic responses of species in space and time. Proc. Royal Soc. B 2010, 277, 661–671. [Google Scholar]
- Royer, A.; Montuire, S.; Legendre, S.; Discamps, E.; Jeannet, M.; Lécuyer, C. Investigating the influence of climate changes on rodent communities at a regional-scale (MIS 1-3, Southwestern France). PLoS ONE 2016, 11, e0145600. [Google Scholar]
- Fürsich, F.T.; Aberhan, M. Significance of time-averaging for palaeocommunity analysis. Lethaia 1990, 23, 143–152. [Google Scholar]
- Denys, C. Rodent faunal lists in karstic and open-air sites of Africa: An attempt to evaluate predation and fossilization biases on paleodiversity. Cuad. De Geol. Ibérica 1997, 23, 73–94. [Google Scholar]
- Moigne, A.M.; Grégoire, S.; Lumley, H.D. Les territoires de chasse et d’exploitation des matières premières des hommes préhistoriques de la Caune de l’Arago entre 600 000 ans et 400 000 ans. Actes Des Congrès Natx. Des Sociétés Hist. Et Sci. 2005, 126, 17–31. [Google Scholar]
- Lisiecki, L.E.; Raymo, M.E. A Pliocene-Pleistocene stack of 57 globally distributed benthic δ18O records. Paleoceanography 2005, 20, 1–17. [Google Scholar]
- López-García, J.M.; Fagoaga, A.; Nabais, M.; Póvoas, L.; Zilhão, J. Late Quaternary (MIS 5a-5b) climate and environments of western Iberia inferred from the small-mammal assemblage of Gruta da Oliveira, Torres Novas, Portugal. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2022, 603, 111194. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxa | D | E | F | G1 | G2 | G3 | G4 | H | I | J | K | L | M | N | O | P | R |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Crocidura sp. | 7 | 1 | 1 | 5 | 14 | ||||||||||||
| Ericaneus davidi | 1 | 2 | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||
| Sorex cf. araneus | 2 | ||||||||||||||||
| Sorex sp. | 1 | ||||||||||||||||
| Talpa europaea | 1 | 1 | 2 | 1 | 1 | 1 | 3 | 6 | 1 | 3 | 1 | 1 | |||||
| Arvicola mosbachensis | 2 | 2 | 1 | 2 | 1 | 2 | 2 | 2 | 3 | 5 | 2 | 3 | 8 | 6 | |||
| Allocricetus bursae | 23 | 39 | 59 | 2 | 4 | 9 | 15 | 63 | 9 | 30 | 20 | 22 | 22 | 20 | 30 | 39 | 27 |
| Apodemus sp. | 29 | 16 | 21 | 5 | 6 | 8 | 26 | 62 | 7 | 23 | 7 | 12 | 2 | 6 | 6 | 13 | 14 |
| Castor fiber | 2 | 1 | 1 | 1 | 2 | 3 | 5 | 1 | 7 | 3 | 1 | 1 | |||||
| Chionomys nivalis | 3 | 2 | 1 | 4 | 1 | 2 | |||||||||||
| Clethrionomys glareolus | 1 | 1 | 1 | ||||||||||||||
| Dicrostonyx sp. | 3 | 3 | 4 | 1 | 1 | 2 | |||||||||||
| Eliomys quercinus | 3 | 5 | 11 | 2 | 2 | 5 | 9 | 19 | 3 | 7 | 2 | 1 | 2 | 1 | 3 | 3 | |
| Glis glis | 2 | ||||||||||||||||
| Hystrix sp. | 3 | 1 | 1 | 1 | |||||||||||||
| M. (Iberomys) brecciensis | 39 | 32 | 38 | 4 | 10 | 22 | 62 | 190 | 8 | 55 | 7 | 3 | 8 | 3 | 8 | 58 | 3 |
| Marmota sp. | 1 | 6 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 2 | 2 | |||
| Microtus agrestis/arvalis | 25 | 28 | 71 | 3 | 4 | 5 | 6 | 37 | 7 | 30 | 15 | 10 | 17 | 9 | 17 | 35 | 10 |
| Lasiopodomys gregalis | 93 | 141 | 213 | 5 | 12 | 6 | 6 | 69 | 13 | 84 | 99 | 106 | 186 | 189 | 420 | 253 | |
| Alexandromys oeconomus | 1 | 1 | 4 | 1 | 7 | 6 | 9 | 18 | 14 | 41 | 21 | ||||||
| Microtus vaufreyi | 73 | 133 | 220 | 13 | 19 | 19 | 27 | 38 | 6 | 15 | 7 | 10 | 10 | 10 | 23 | 47 | 3 |
| Pliomys episcopalis | 2 | ||||||||||||||||
| Pliomys coronensis | 32 | 83 | 192 | 10 | 18 | 21 | 29 | 81 | 14 | 59 | 43 | 39 | 63 | 84 | 177 | 127 | 15 |
| Sciurus sp. | 1 | 1 | 2 | 1 | 1 | 1 | |||||||||||
| Spermophilus sp. | 11 | 31 | 35 | 4 | 1 | 1 | 2 | 1 | 7 | 5 | 2 | 3 | 4 | 6 | 9 | ||
| Ochotona pusilla | 1 | 1 | 4 | 7 | 7 | 2 | 1 | 3 | 2 | 2 | 1 | ||||||
| Oryctolagus cuniculus | 5 | 16 | 48 | 7 | 12 | 13 | 40 | 45 | 9 | 37 | 9 | 16 | 7 | 24 | |||
| Lepus timidus | 1 | 2 | 3 | 1 | 2 | 1 | 1 | 2 | 1 | 6 |
| US | T°max | MAT | T°min | |||
|---|---|---|---|---|---|---|
| BM | QEM | BM | QEM | BM | QEM | |
| D | 20.6 (±2.6) | 21.0 | 6.9 (±3.4) | 7.2 | −5.3 (±7.4) | −6.8 |
| E | 20.1 (±2.5) | 20.8 | 6.3 (±3.3) | 6.3 | −6.1 (±7.4) | −8.5 |
| F | 19.8 (±2.5) | 20.9 | 6.1 (±3.3) | 6.7 | −6.2 (±7.4) | −7.8 |
| G1 | 20.7 (±2.6) | 20.7 | 7.0 (±3.4) | 7.3 | −5.1 (±7.4) | −6.4 |
| G2 | 20.5 (±2.5) | 20.8 | 7.1 (±3.3) | 7.4 | −5.0 (±7.4) | −.6.2 |
| G3 | 21.4 (±2.7) | 21.5 | 7.2 (±3.6) | 8.4 | −5.7 (±8) | −4.9 |
| G4 | 22.1 (±2.7) | 21.4 | 8.1 (±3.6) | 8.3 | −4.5 (±8) | −4.9 |
| H | 21.4 (±2.7) | 21.1 | 7.3 (±3.6) | 7.3 | −5.4 (±7.9) | −6.7 |
| I | 21.8 (±3.6) | 21.4 | 8.1 (±3.5) | 8.1 | −4.4 (±7.8) | −5.4 |
| J | 21.6 (±2.8) | 21.2 | 7.5 (±3.6) | 7.3 | −5.4 (±7.9) | −6.8 |
| K | 20.3 (±2.6) | 20.7 | 6.6 (±3.3) | 6.4 | −5.7 (±7.4) | −8.2 |
| L | 21.6 (±2.8) | 21.2 | 7.5 (±3.6) | 7.2 | −5.4 (±7.9) | −7.1 |
| M | 21.3 (±2.7) | 21.8 | 7.3 (±3.5) | 7.8 | −5.4 (±7.8) | −6.5 |
| N | 21.4 (±2.7) | 22 | 7.8 (±3.5) | 7.7 | −4.5 (±7.8) | −6.9 |
| O | 20.8 (±2.6) | 21.8 | 6.0 (±3.4) | 7.8 | −7.8 (±7.6) | −6.7 |
| P | 21.2 (±2.7) | 21.45 | 7.0 (±3.5) | 7.7 | −5.5 (±7.7) | −6.3 |
| R | 22.6 (±2.9) | 21.9 | 9.5 (±3.7) | 9.5 | −2.3 (±7.7) | −3.0 |
| period | −430 ka | −440 ka | −450 ka | −460 ka | −470 ka | −480 ka |
| MAT | 9.2 | 8.7 | 9.1 | 10 | 9.8 | 11.1 |
| period | −490 ka | −500 ka | −510 ka | −520 ka | −530 ka | −540 ka |
| MAT | 13.2 | 12.1 | 11.6 | 11.2 | 10.7 | 10.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lebreton, L.; López-García, J.M. Stage or Sub-Stage: The Contribution of Small Mammals to the Characterization of Middle Pleistocene Local Climate Variation. Quaternary 2023, 6, 54. https://doi.org/10.3390/quat6040054
Lebreton L, López-García JM. Stage or Sub-Stage: The Contribution of Small Mammals to the Characterization of Middle Pleistocene Local Climate Variation. Quaternary. 2023; 6(4):54. https://doi.org/10.3390/quat6040054
Chicago/Turabian StyleLebreton, Loïc, and Juan Manuel López-García. 2023. "Stage or Sub-Stage: The Contribution of Small Mammals to the Characterization of Middle Pleistocene Local Climate Variation" Quaternary 6, no. 4: 54. https://doi.org/10.3390/quat6040054
APA StyleLebreton, L., & López-García, J. M. (2023). Stage or Sub-Stage: The Contribution of Small Mammals to the Characterization of Middle Pleistocene Local Climate Variation. Quaternary, 6(4), 54. https://doi.org/10.3390/quat6040054