
Old World Fossil Equus (Perissodactyla, Mammalia), Extant Wild Relatives and Incertae Sedis Forms
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Material and Methods
2.1. Material
2.2. Methods
2.2.1. System of Measurements
2.2.2. Simpson’s Diagrams
3. Taxonomy
3.1. Diagnosis of Genus Equus
3.2. Affinities between Extant Subgenera
3.3. Affinities between Sussemiones and Pseudohydruntines
4. Descriptions and Discussions
4.1. Hemiones
4.1.1. Extant Taxa

4.1.2. E. (Hemionus) hydruntinus Regalia, 1907






4.1.3. Other Fossil Hemiones
4.2. Asses
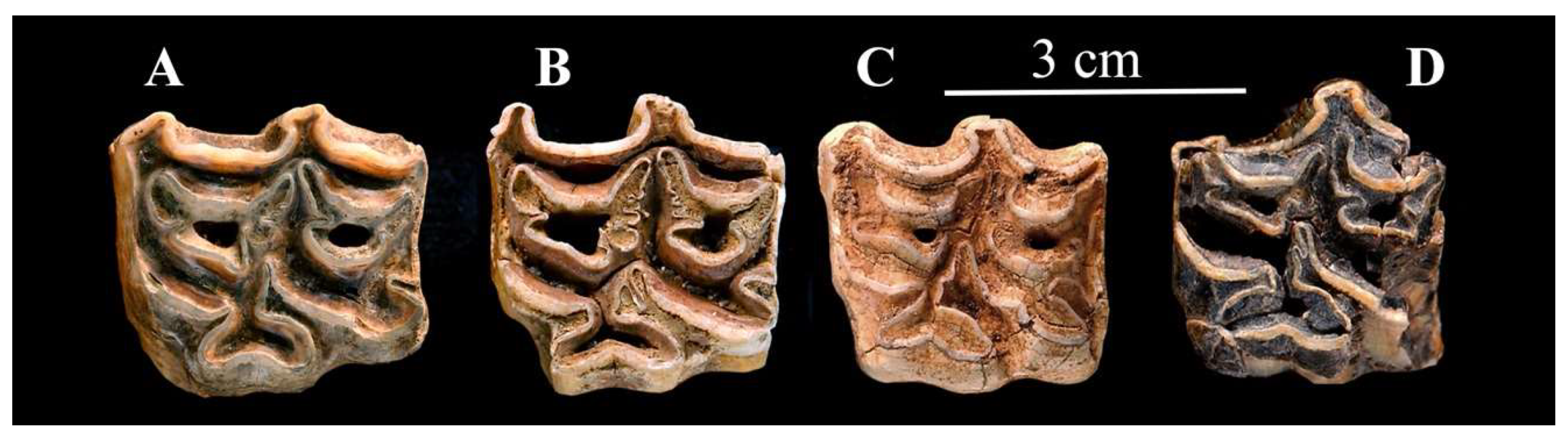
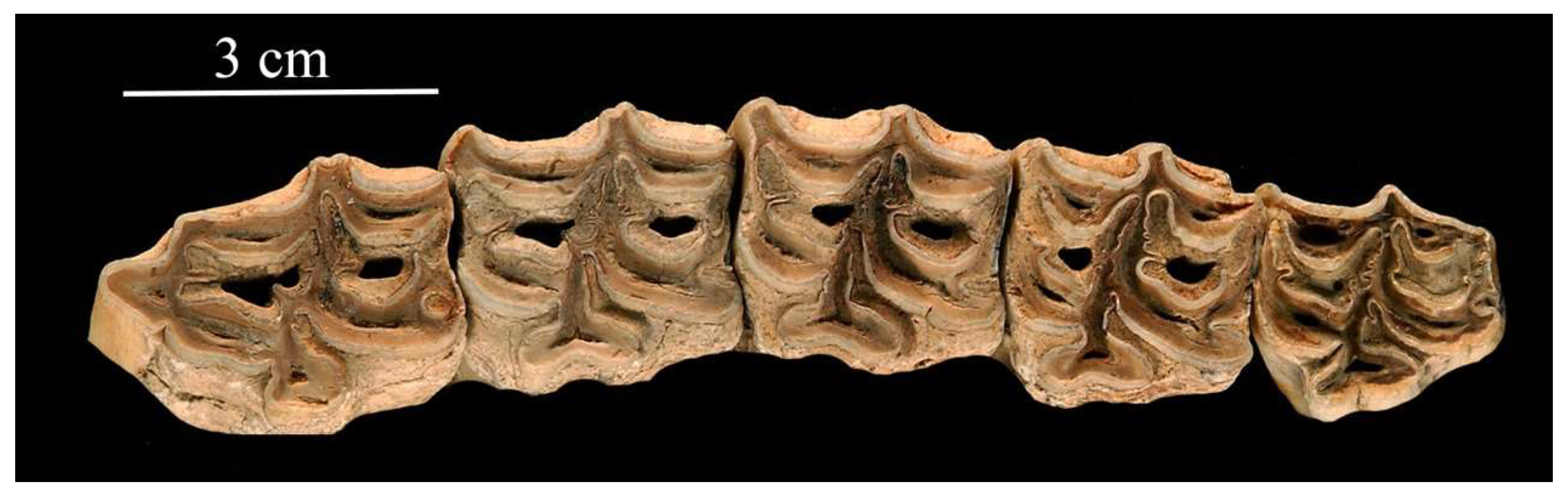
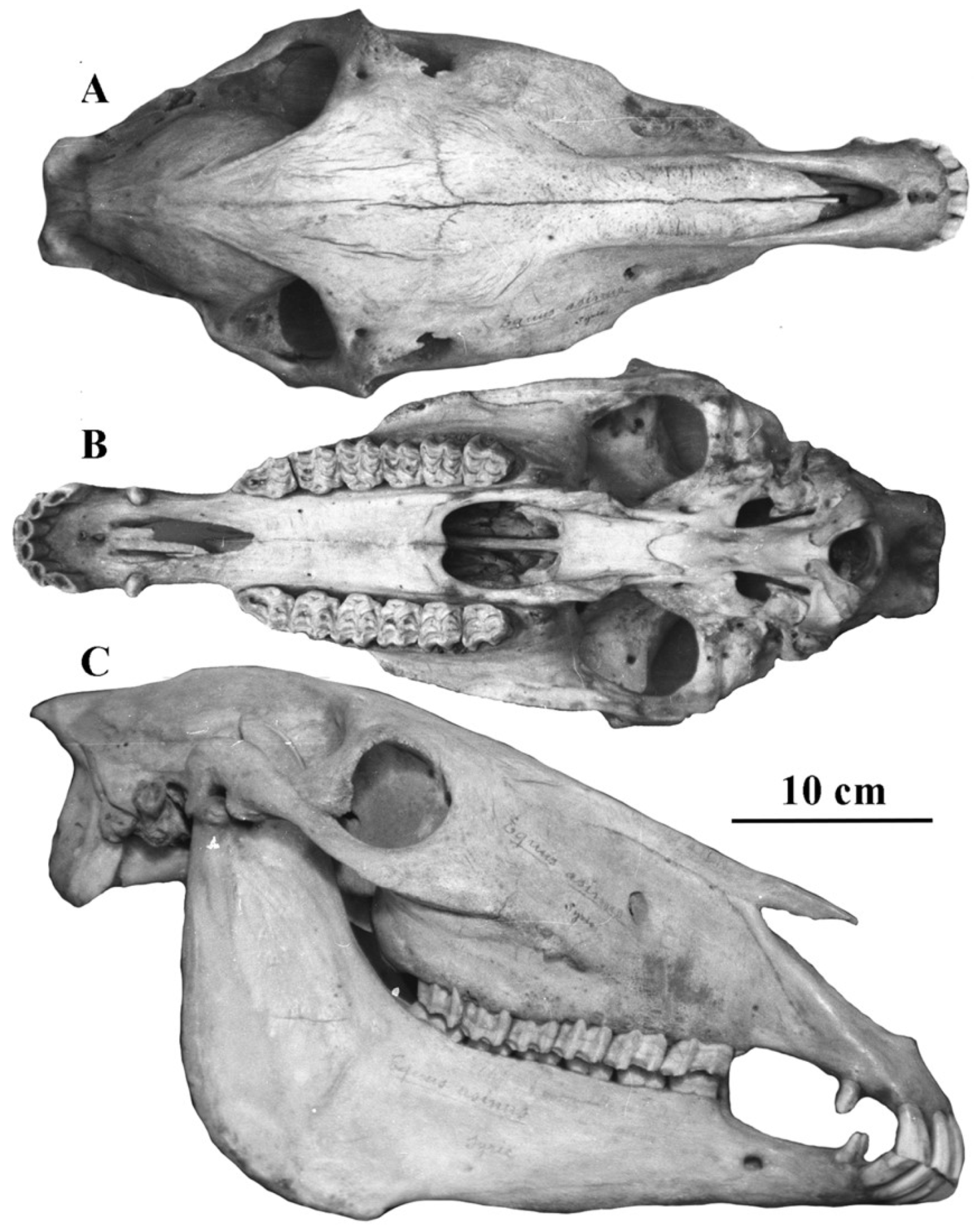
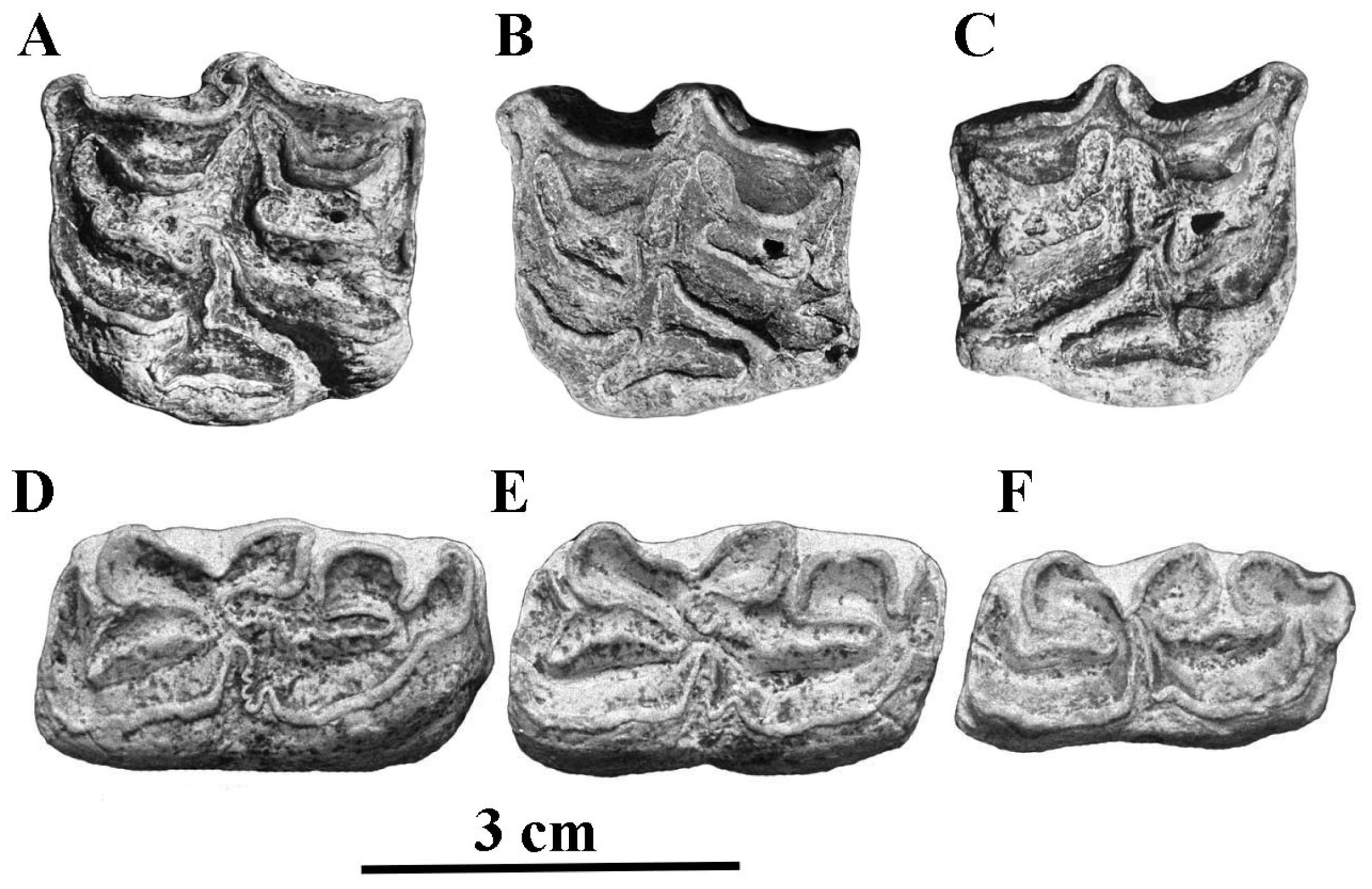
4.2.1. E. (Asinus) africanus Heuglin and Fitzinger, 1866 (Figure 11)




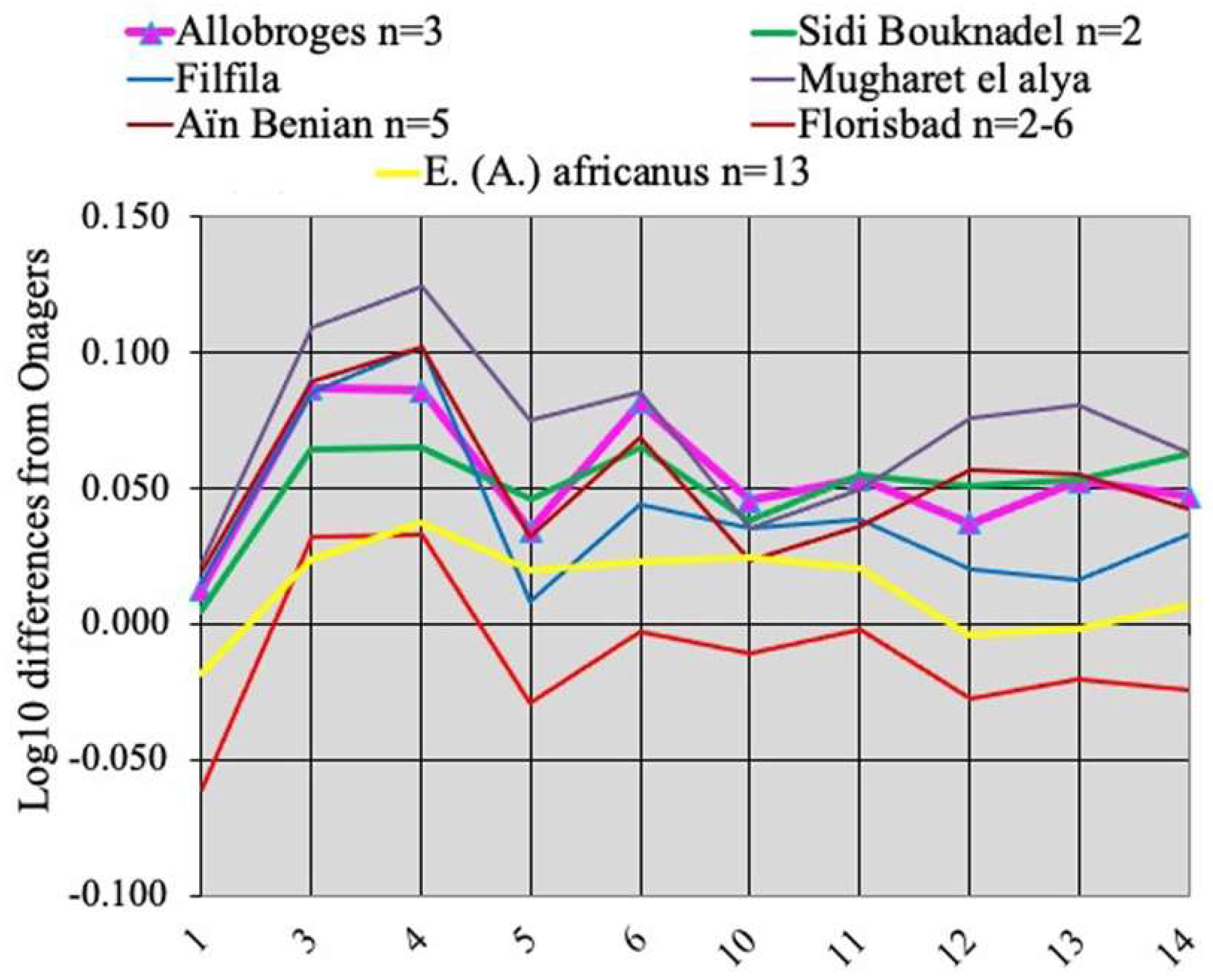
4.2.2. E. (Asinus) atlanticus Thomas, 1884

4.2.3. E. (Asinus) melkiensis Bagtache, Hadjouis, Eisenmann, 1984
4.2.4. E. (Asinus) asinus Linné, 1758 and E. (Asinus) sp.

4.2.5. E. graziosii Azzaroli, 1966
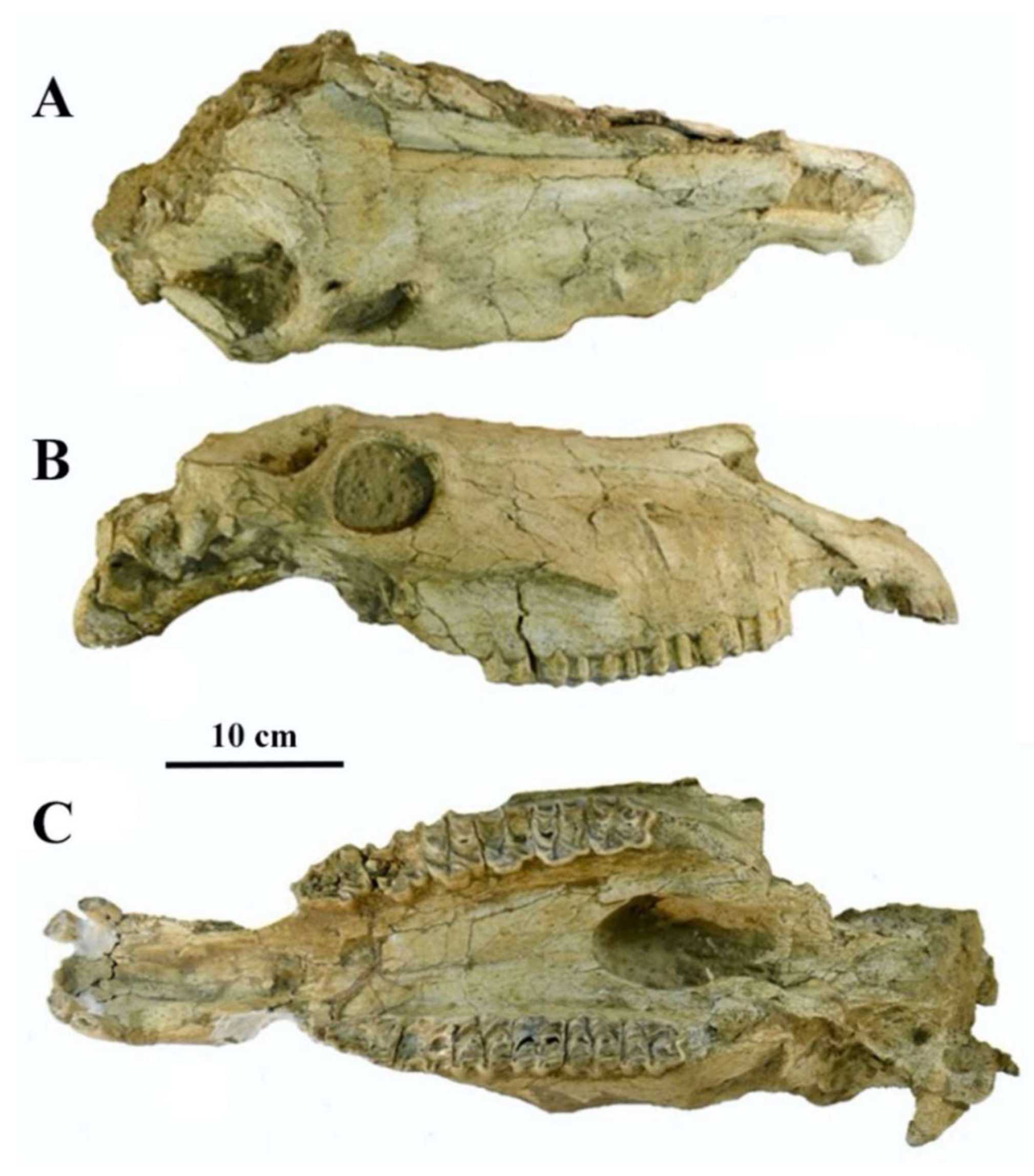
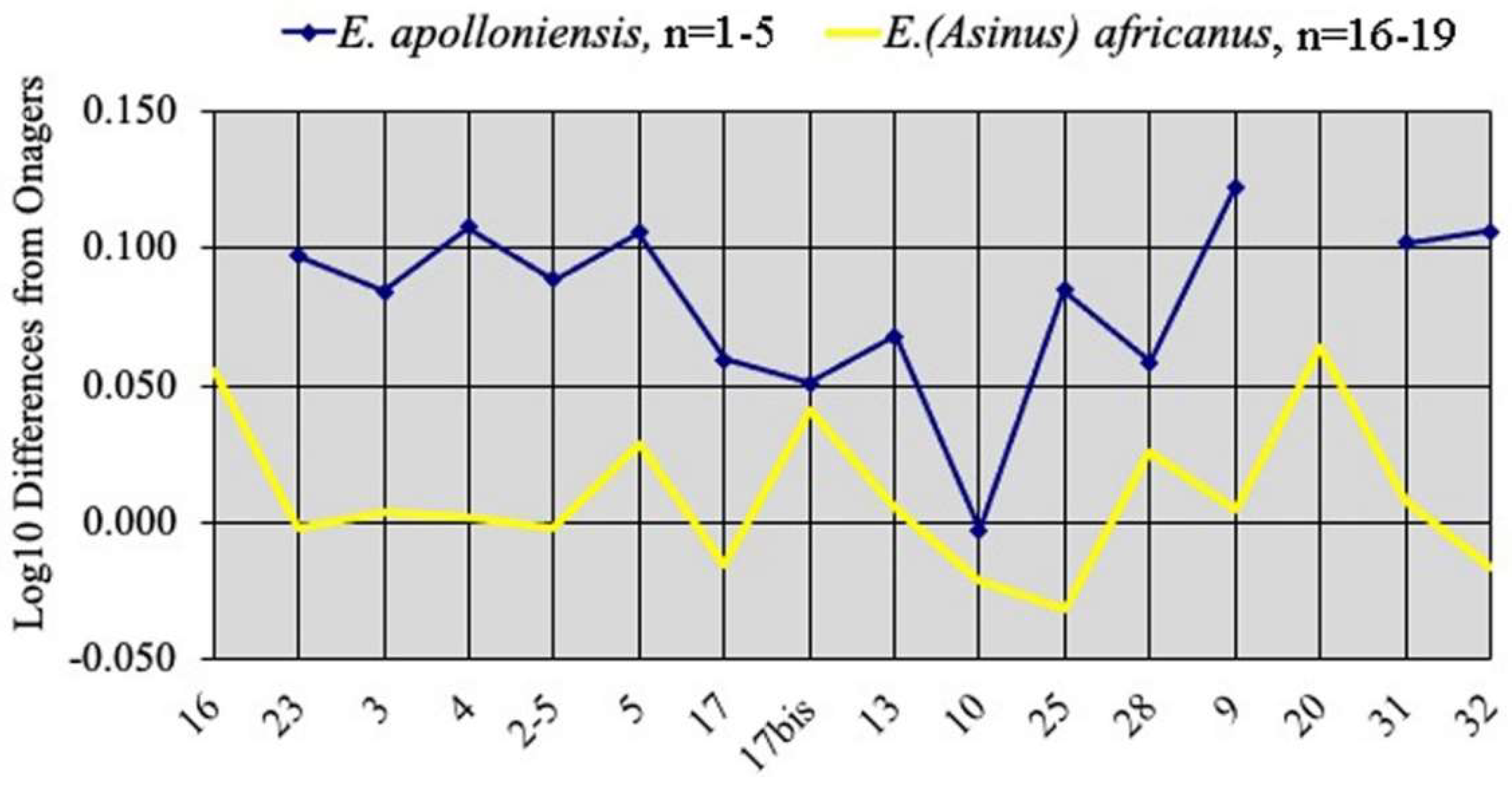
4.2.6. E. (Asinus) apolloniensis Koufos et al., 1997
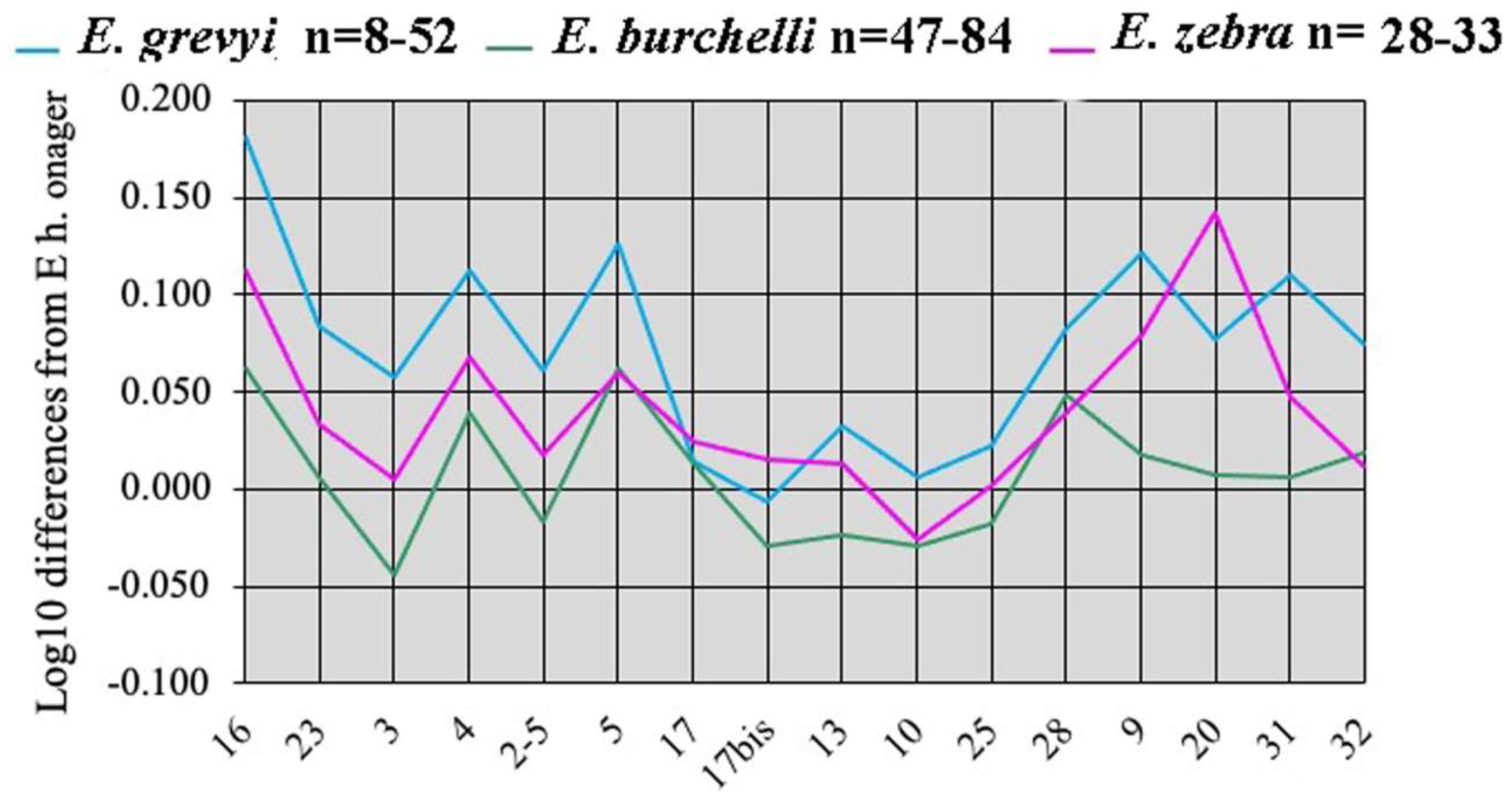
4.3. E. (Dolichohippus) grevyi Oustalet, 1882
4.4. E. (Quagga) Shortridge, 1934 (Plain’s Zebras)
4.4.1. Extant Quaggas
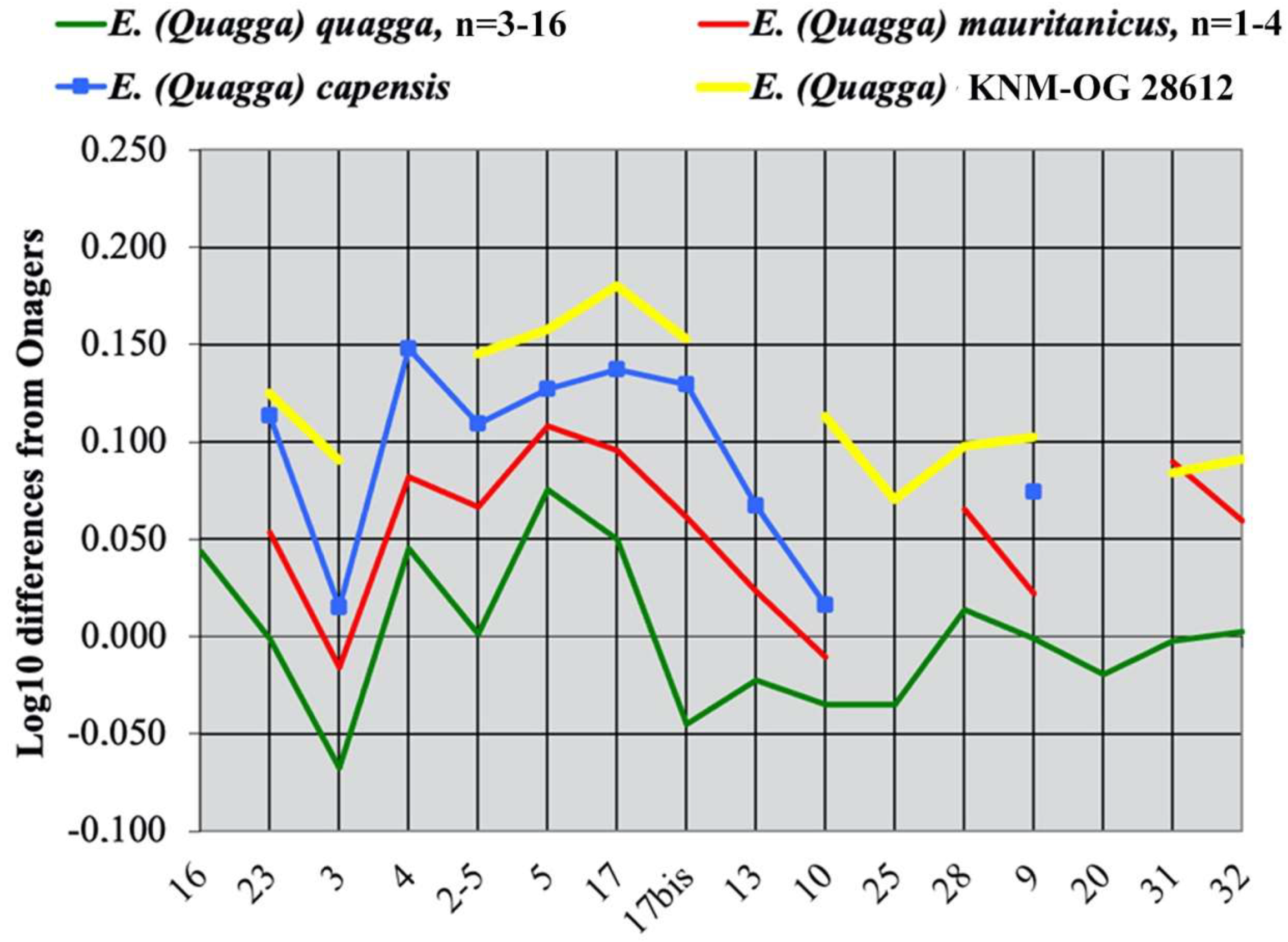
4.4.2. E. (Quagga) mauritanicus Pomel, 1897
4.4.3. E. (Quagga) capensis Broom, 1909
4.4.4. E. (Quagga) oldowayensis Hopwood, 1937
4.5. E. (Hippotigris) Smith, 1841 (Mountain Zebras)
4.6. E. (Equus) Linnaeus, 1758
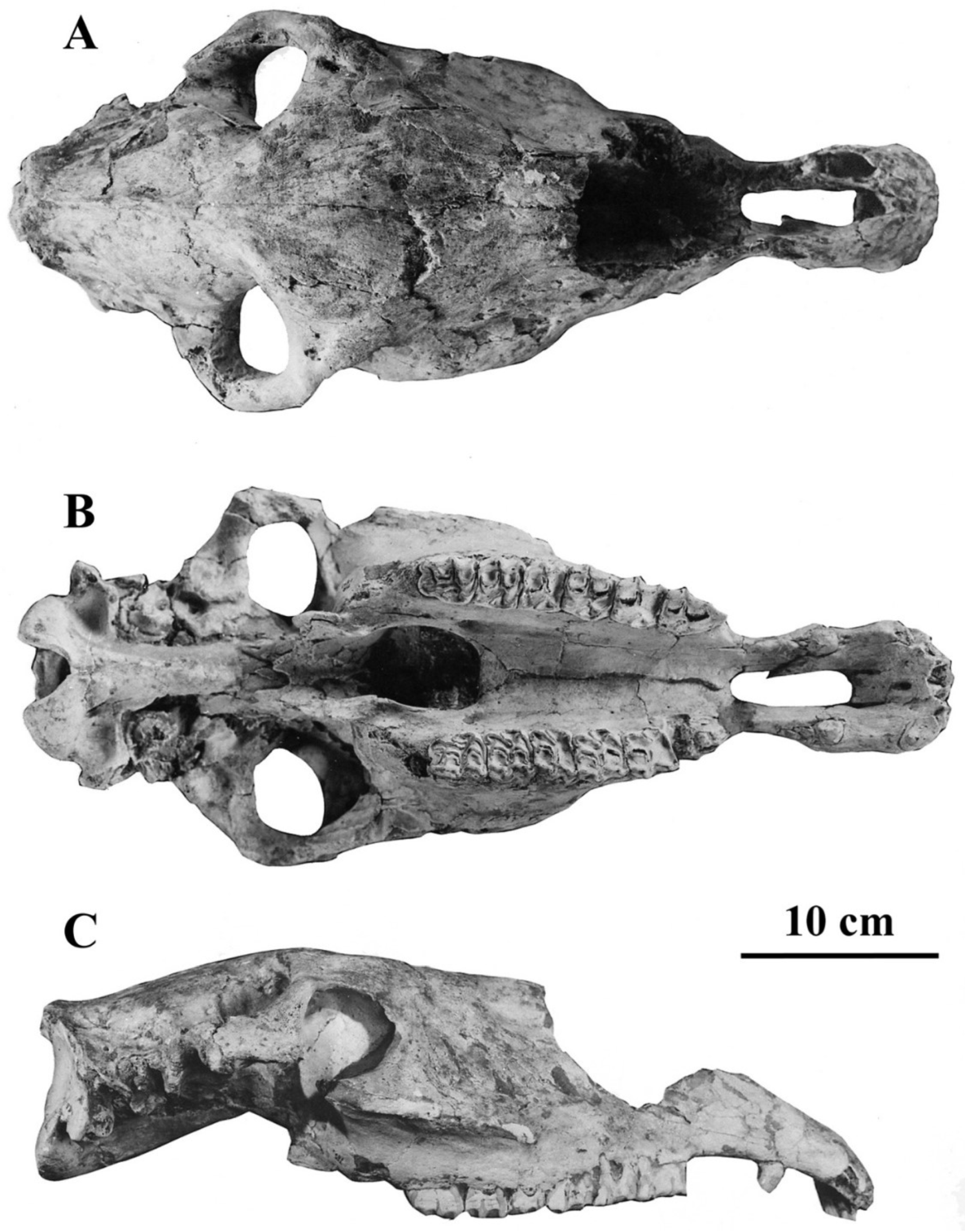
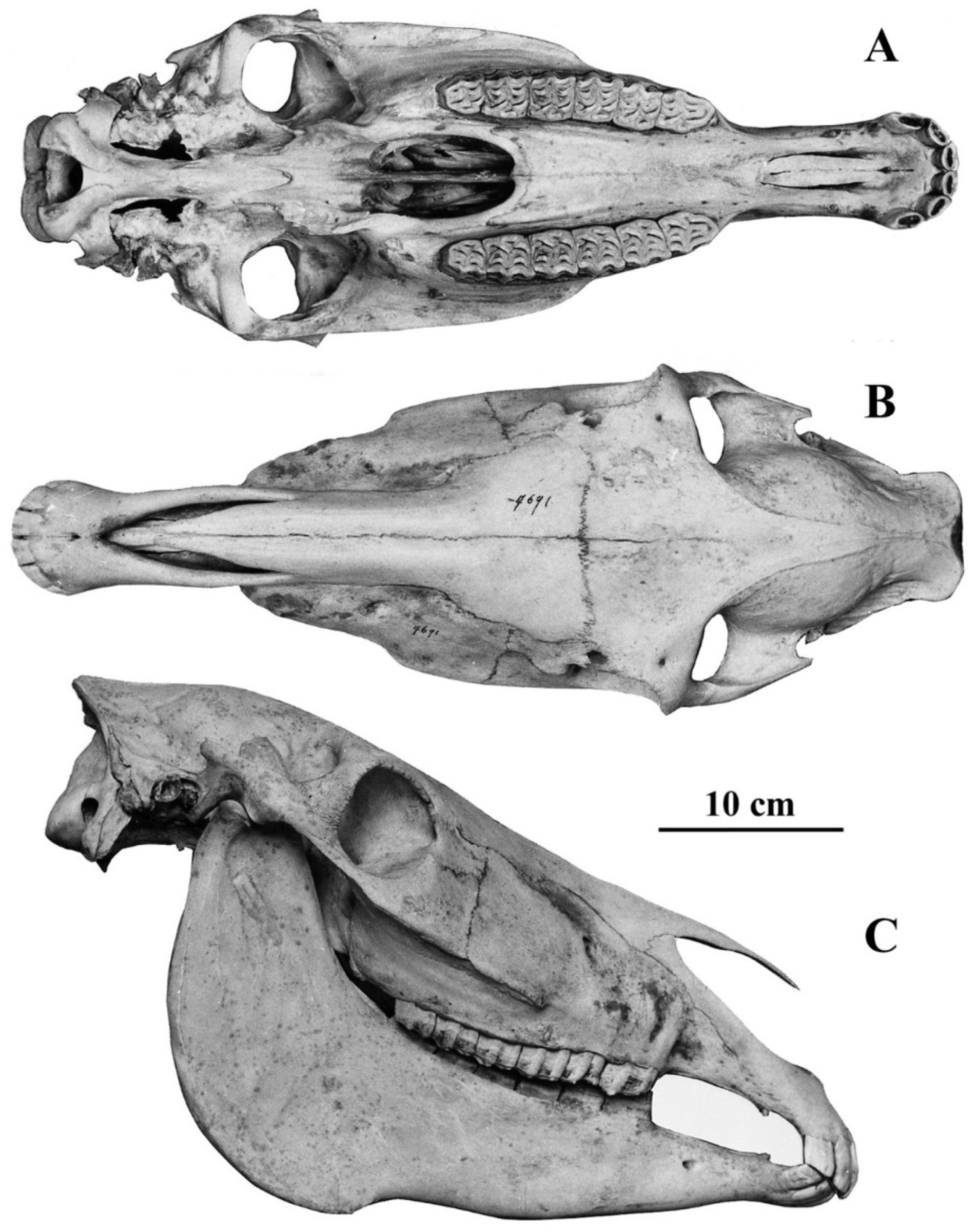
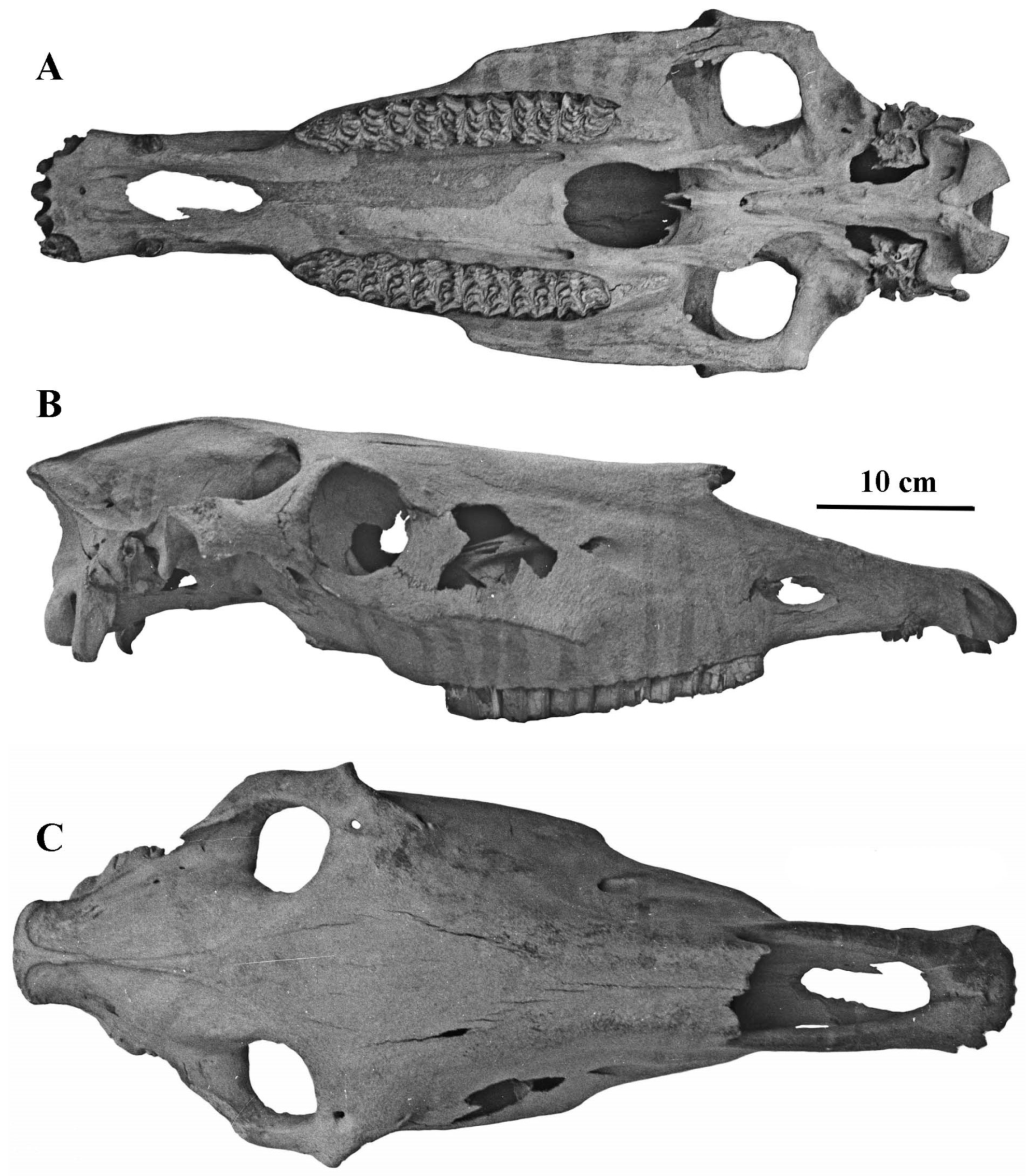
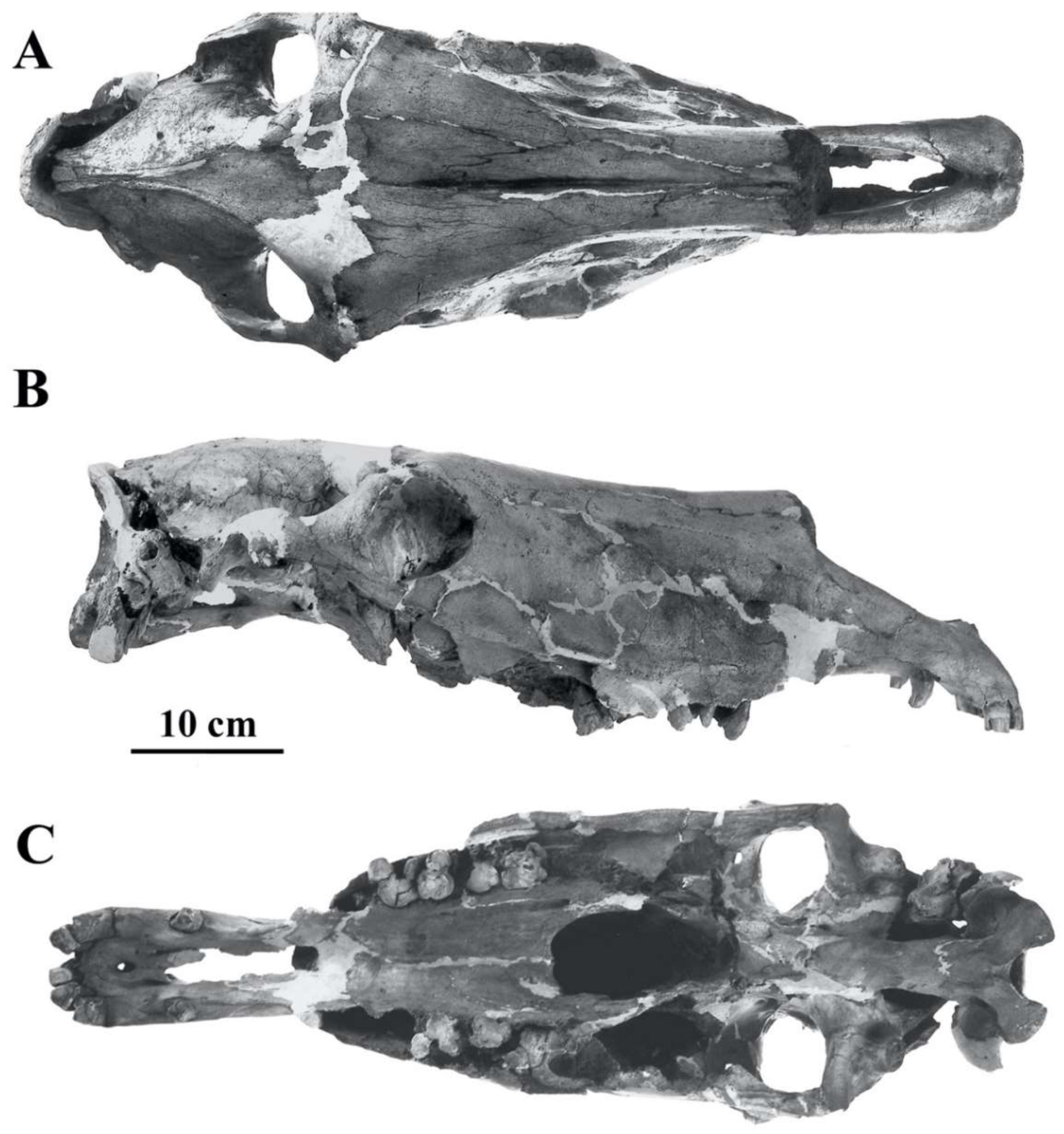
- Cranium characteristics: Franck’s and Palatal indices [89].
- 2.
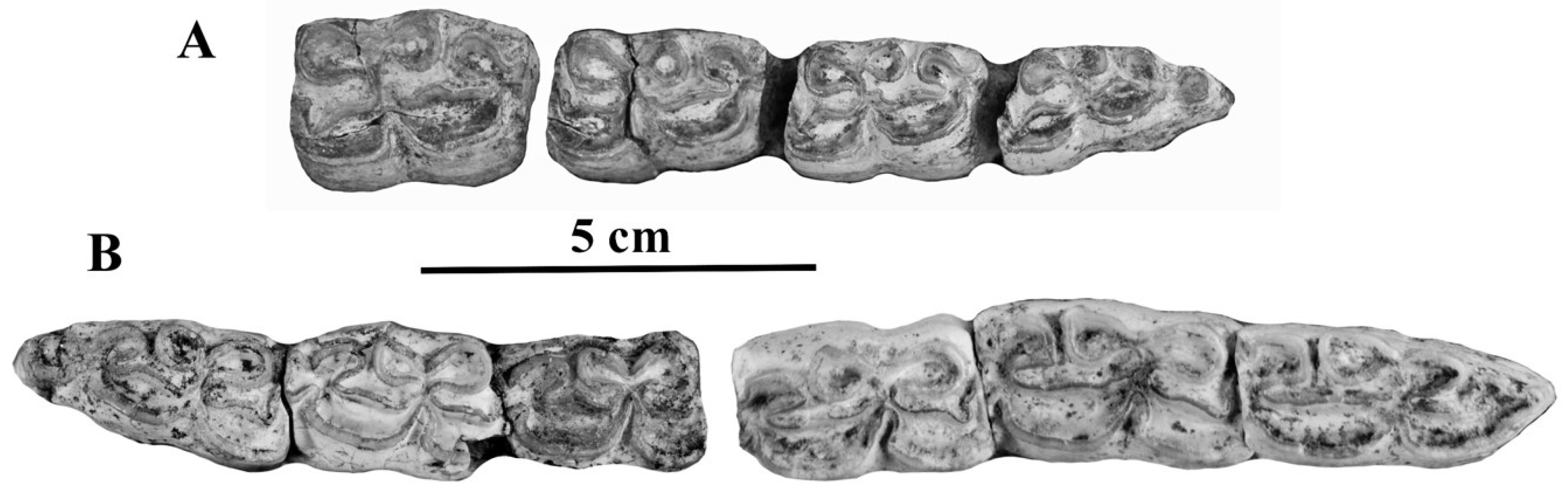
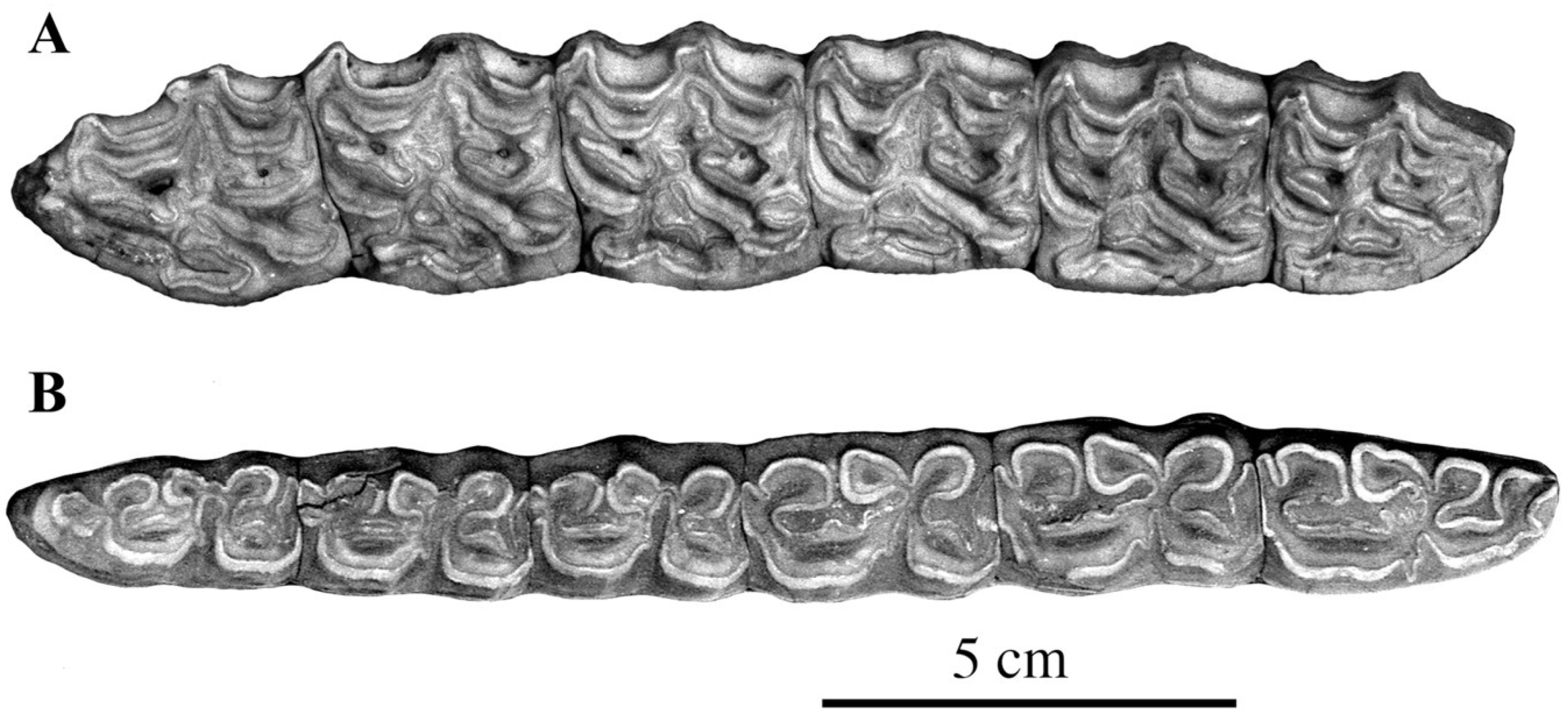
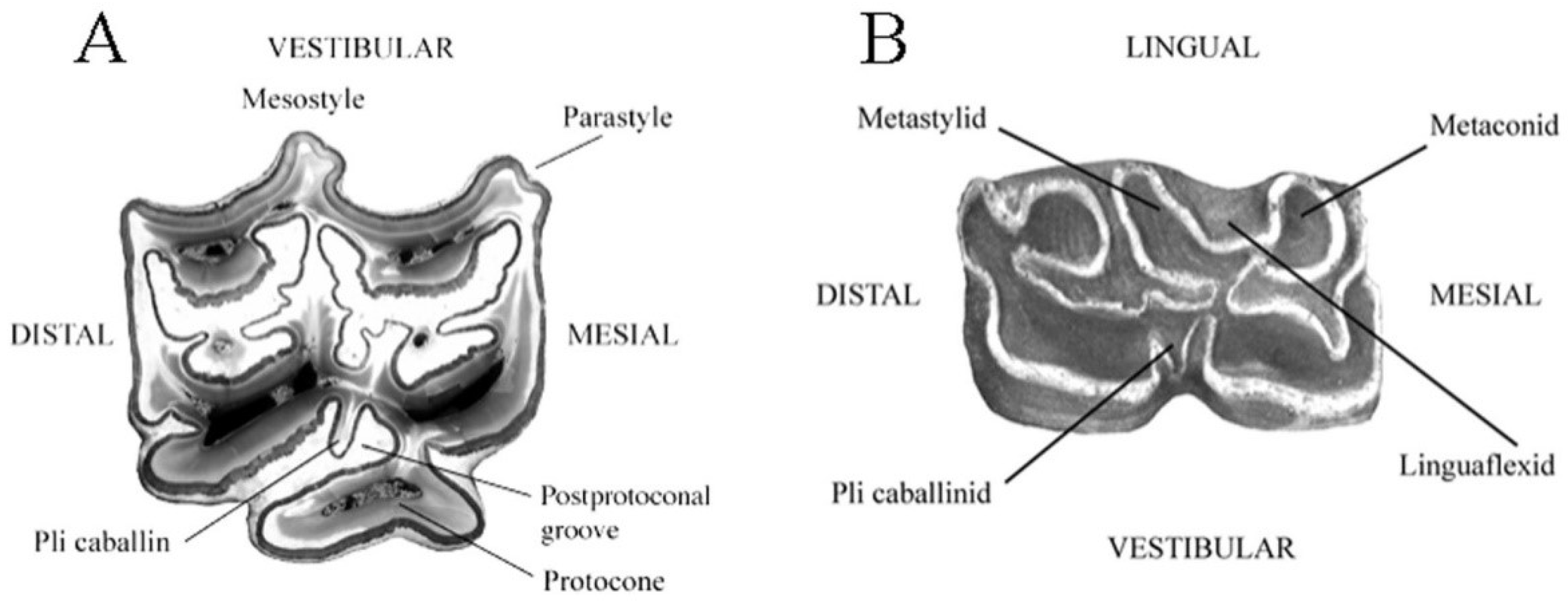
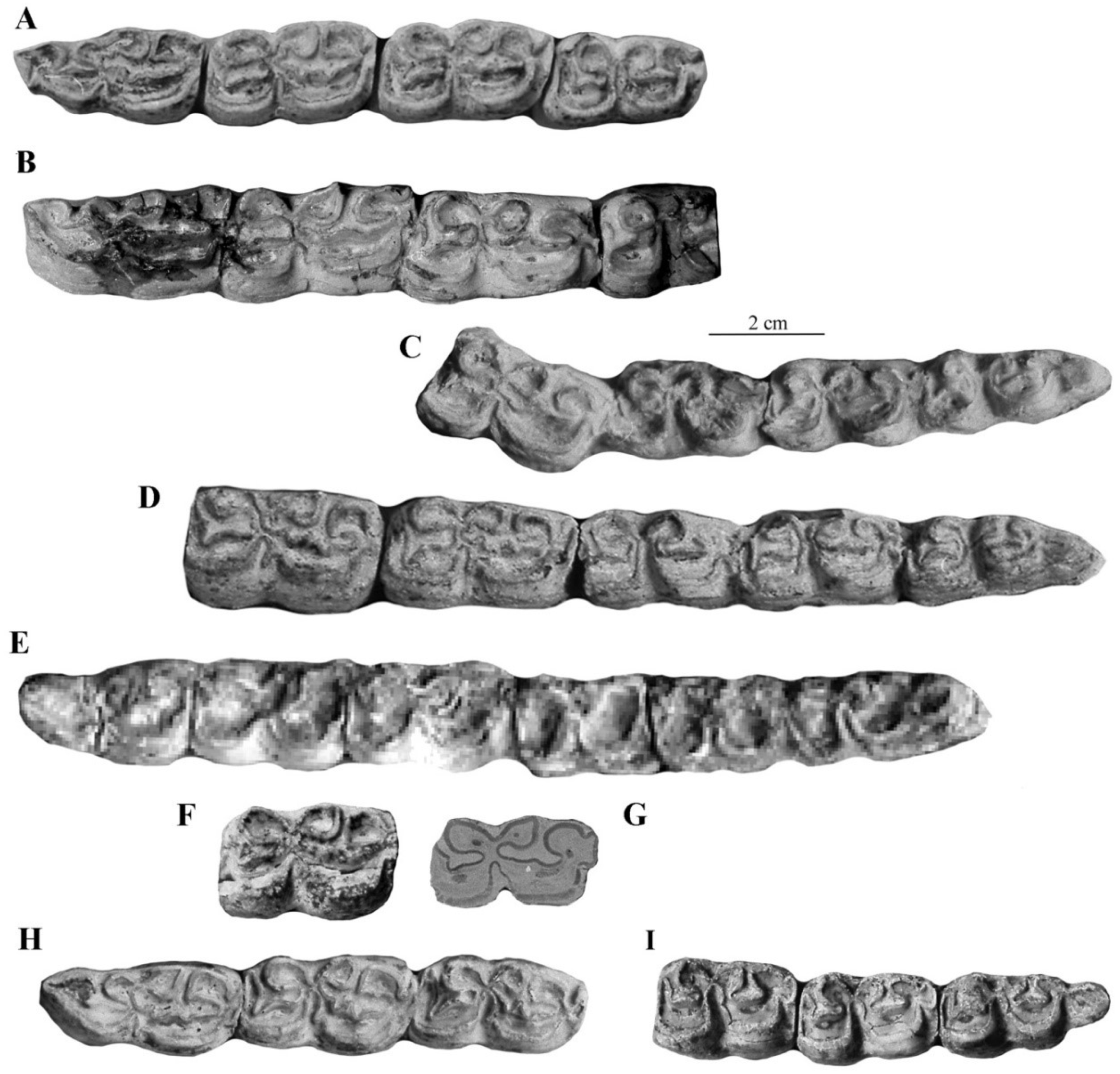
- Cheek teeth (Figure 55)
- -
- The parastyle and mesostyle are grooved at least on premolars.
- -
- The vestibular enamel in the front and back of the mesostyle is concave.
- -
- The pli caballin is present at least on premolars.
- -
- The post-protoconal groove is not very deep.
- -
- The protocone is long and asymmetric, being less developed medially than distally.
- -
- The linguaflexid is angular.
- -
- The double knot (metaconid + linguaflexid + metastylid) is asymmetric.
- -
- The metastylid is pointed.
- -
- The pli caballinid is present at least on premolars.
- 3.
- Metapodials
4.6.1. E. (Equus) ferus Boddaert, 1785
4.6.2. E. (Equus) przewalskii Polyakoff, 1881
4.6.3. E. (Equus) ferus cf. scotti Gidley, 1900
4.6.4. E. (Equus) ferus chosaricus Gromova, 1949
4.6.5. E. (Equus) ferus germanicus Nehring, 1884
4.6.6. E. (Equus) antunesi Cardoso and Eisenmann, 1989
4.7. Sussemionus Eisenmann, 2010 [96]
4.7.1. E. (Sussemionus) coliemensis, Lazarev, 1980
4.7.2. E. (Sussemionus) verae Sher, 1971
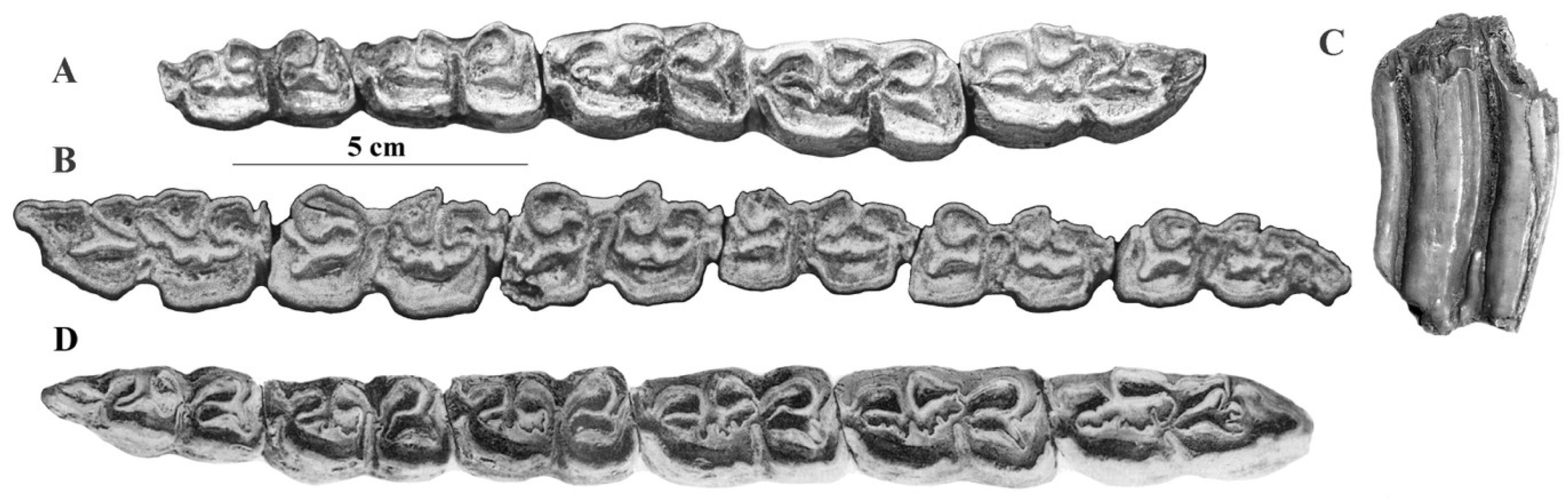
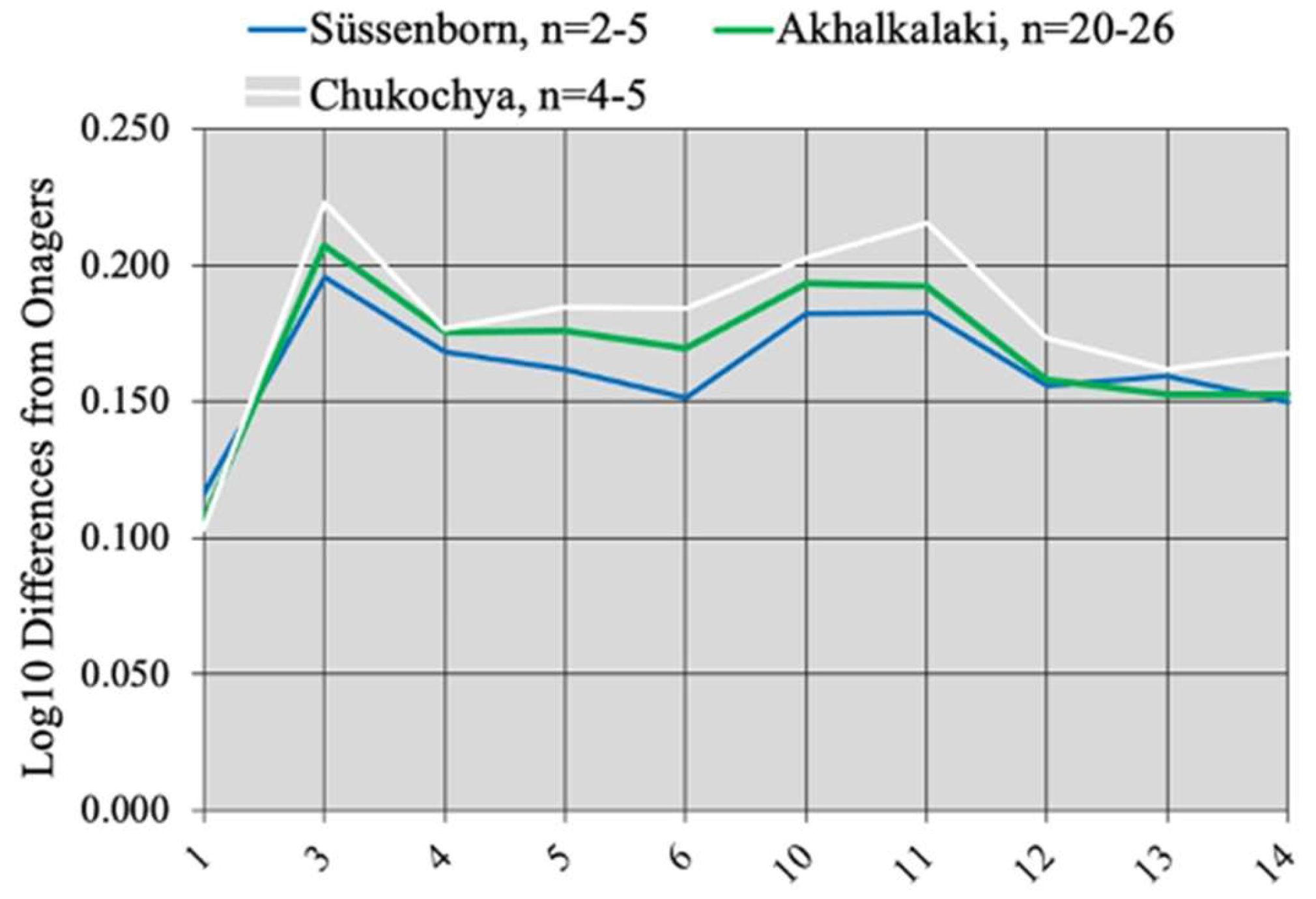
4.7.3. E. (Sussemionus) suessenbornensis Wüst, 1901
4.7.4. E. (Sussemionus) aff. sussenbornensis Vekua, 1962
4.7.5. Concluding Remarks
4.8. Pseudohydruntines and E. altidens
4.8.1. The Problem with ‘Hippotigris’ altidens Reichenau, 1915
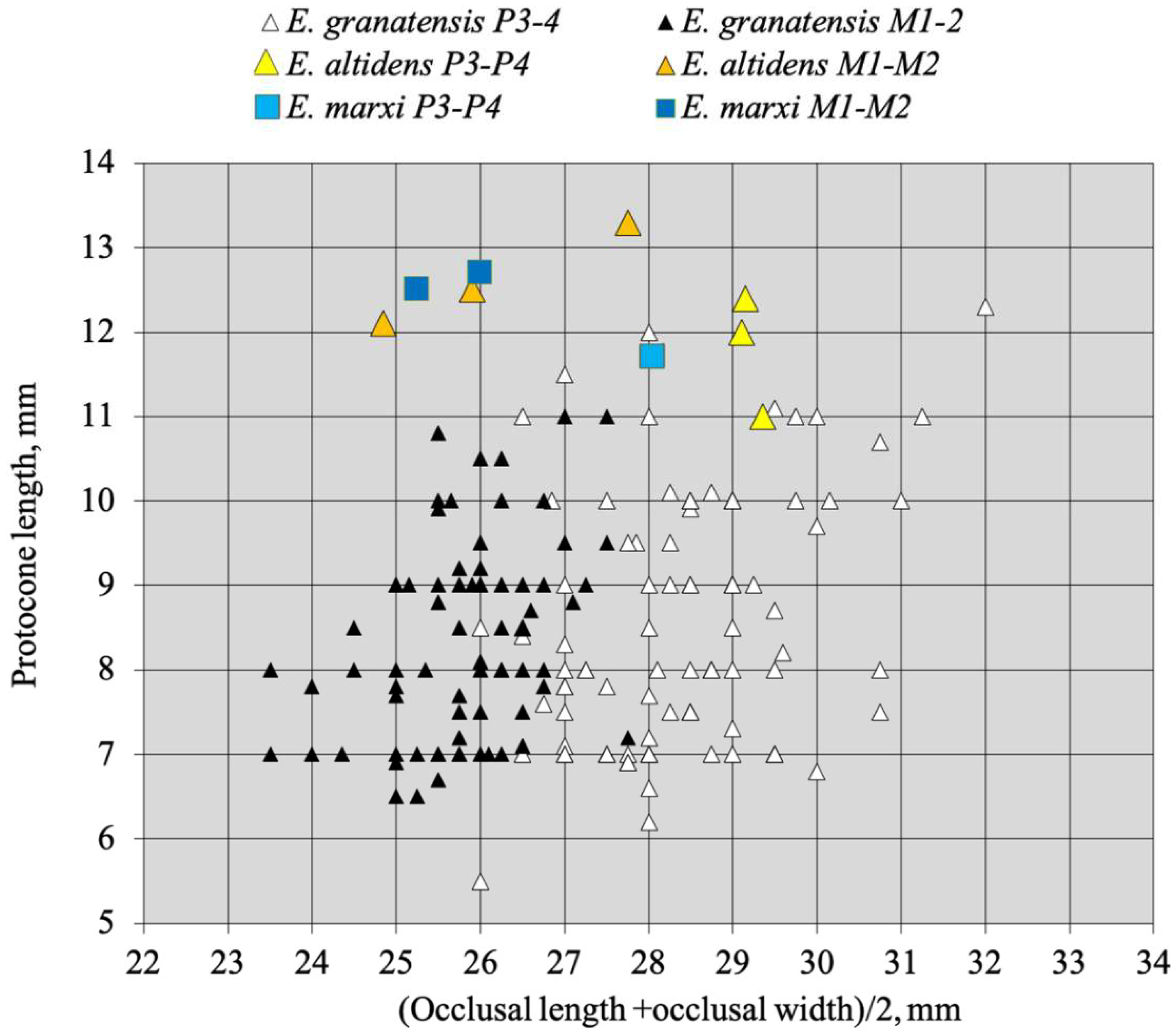
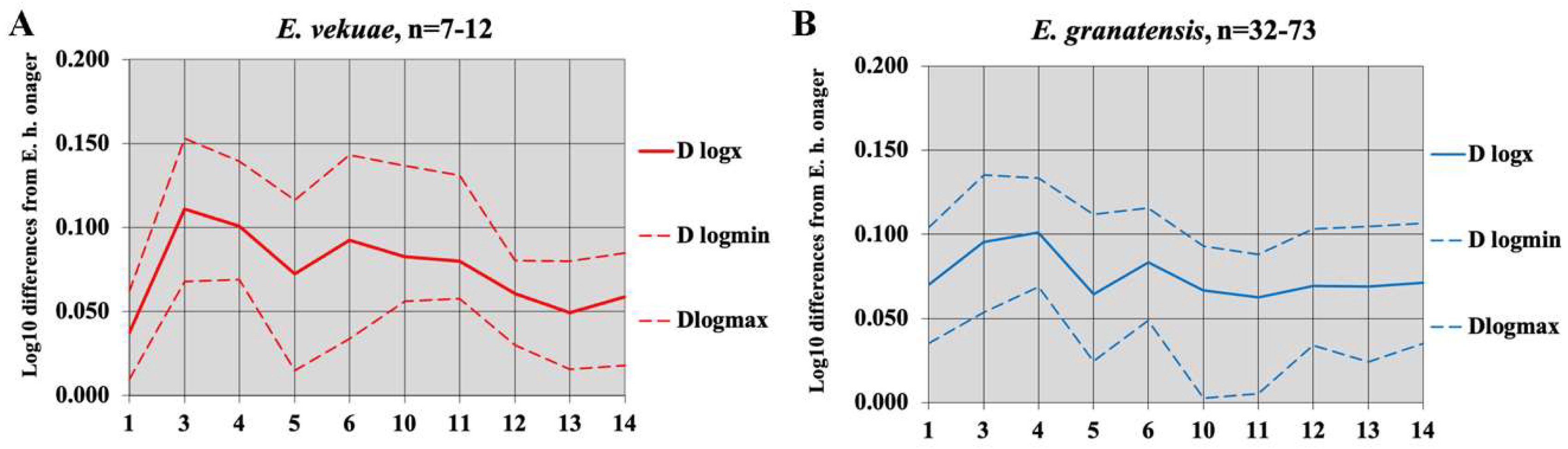
4.8.2. E. granatensis Alberdi and Bustos, 1985
4.8.3. E. aff. granatensis, Pirro, Italy
4.8.4. E. wuesti Musil, 2001
4.8.5. E. hipparionoides Vekua, 1962
4.8.6. E. ovodovi Eisenmann and Vasiliev, 2011
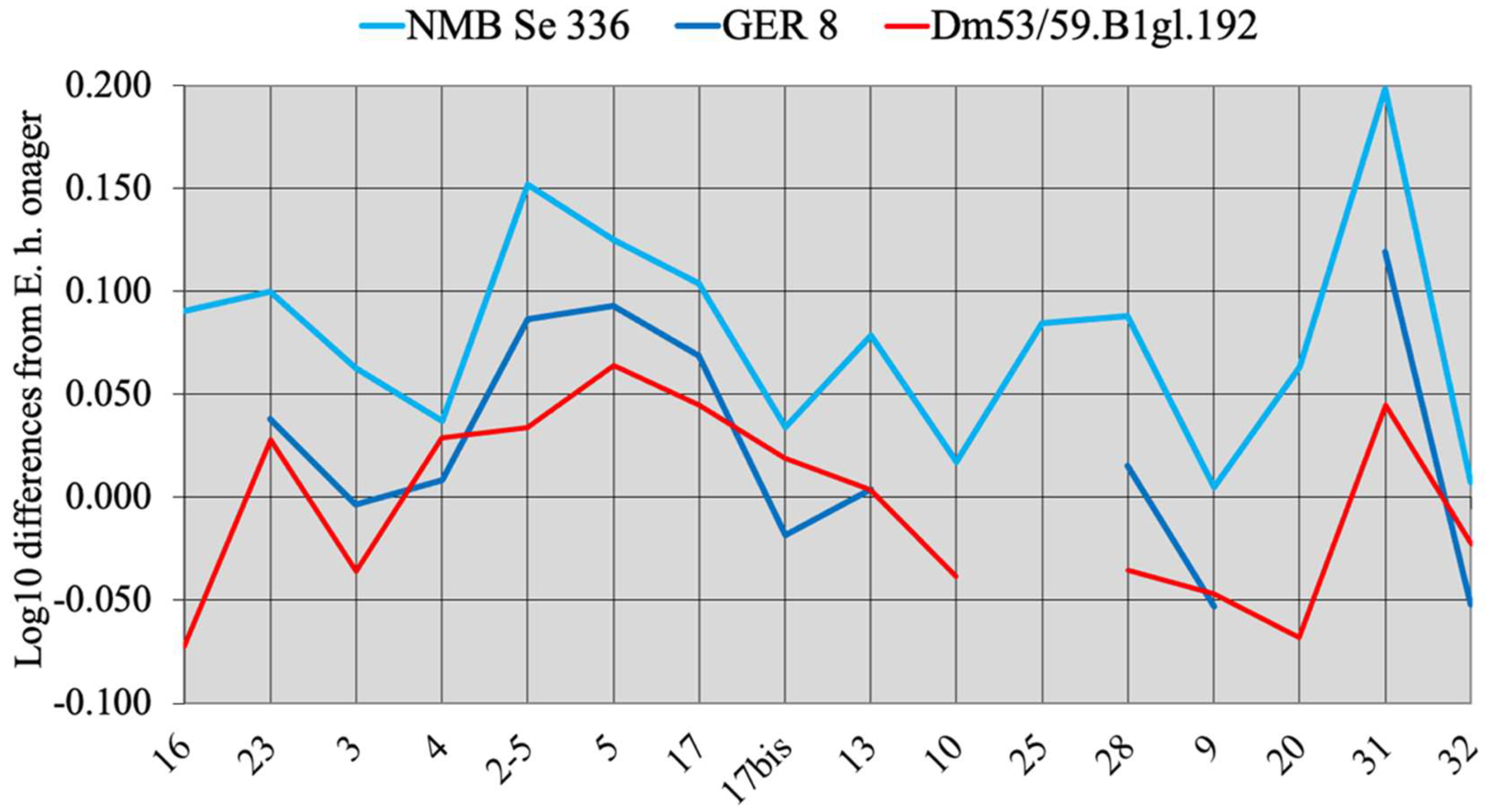
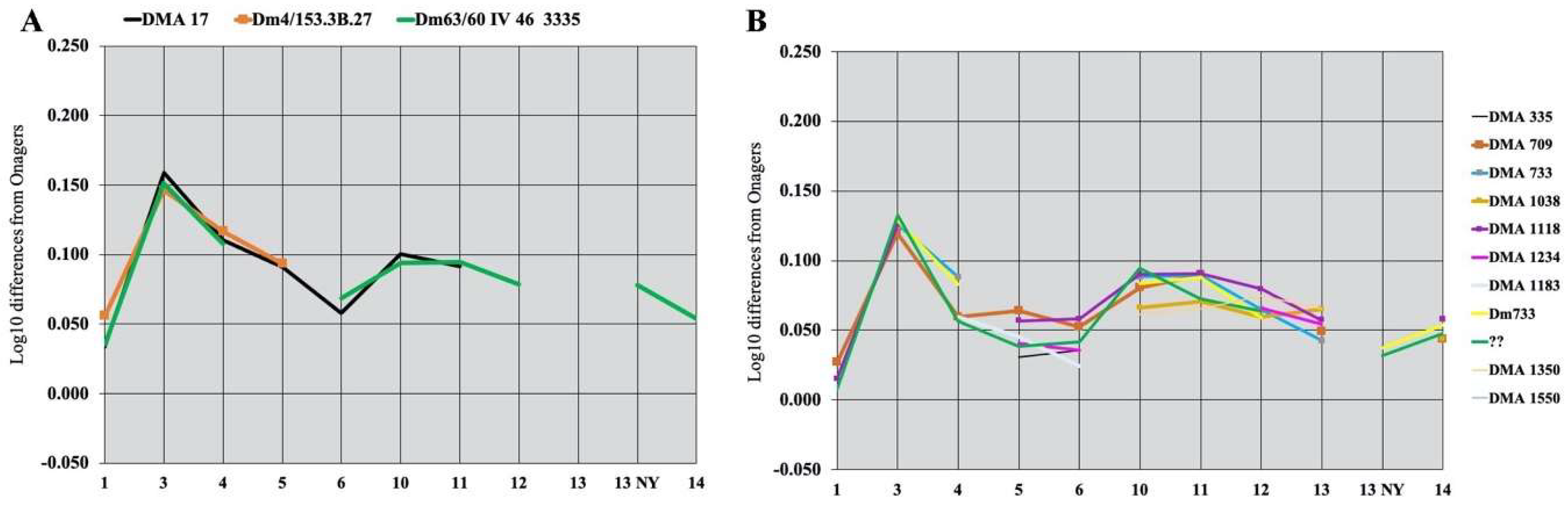
4.8.7. Dmanisi
Preliminary Observations
Equus vekuae n. sp.
- -
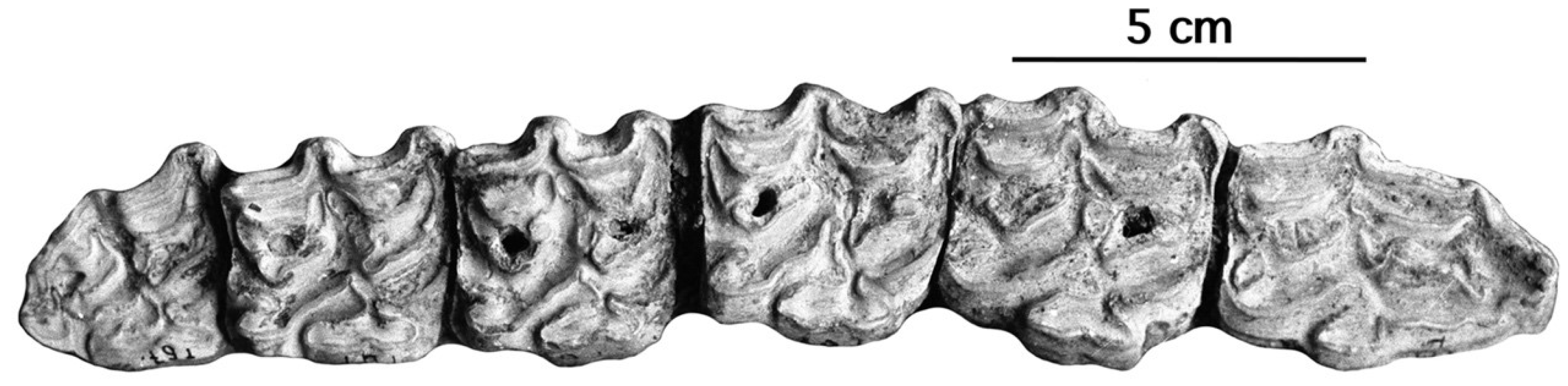
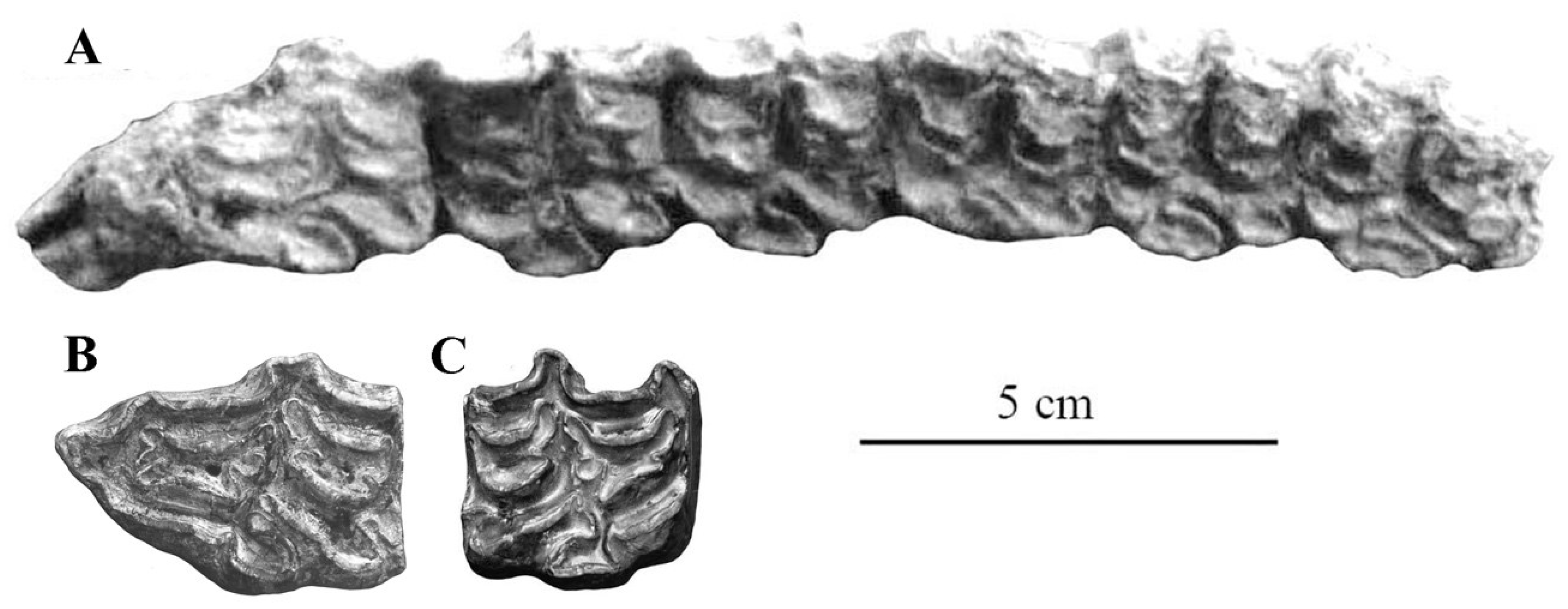
- From all Allohippus crania, the cranium of E. vekuae differs by the longer distance between the Basion and the Hormion (measurement 4) and a shorter naso-incisival notch (measurement 32). From Allohippus mygdoniensis and Allohippus senezensis in particular (Figure 91), it differs also by shorter choanae (measurement 9). From Allohippus upper cheek teeth, those of E. vekuae differ by deeper post-protoconal grooves. From Allohippus lower cheek teeth, those of E. vekuae differ by the elongated metaconids and the occurrence of a pli protostylid on the p2.
- -
- From E. (suessemionus) granatensis—the best documented of all Equids usually referred to as ‘E. altidens’, E. vekuae differs by the lack of isolated stylids and MC robustness (Figure 99).
4.9. Incertae Sedis
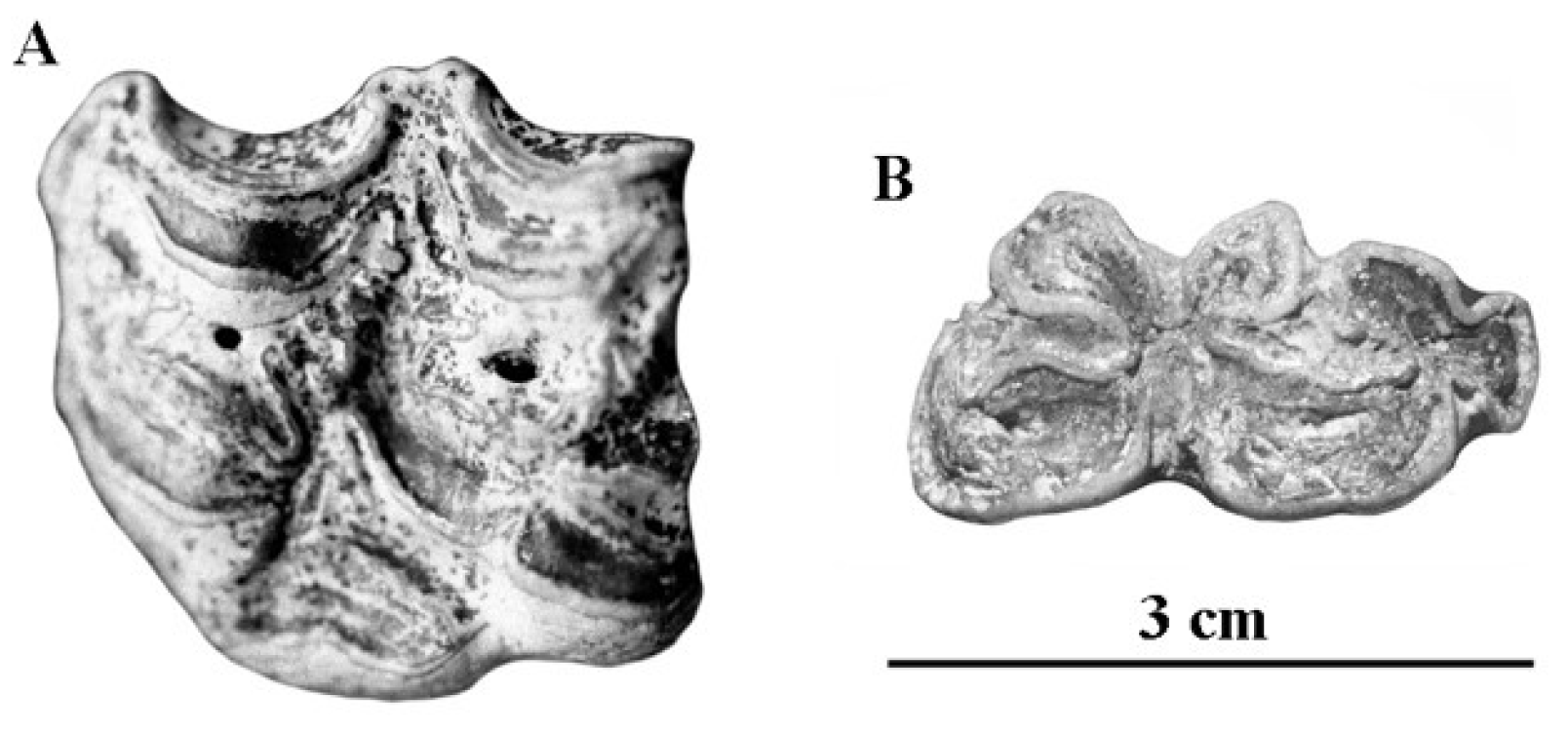
4.9.1. E. marxi Reichenau, 1915
4.9.2. E. tabeti Arambourg, 1970
4.9.3. E. nalaikhaensis Kuznetsova and Zhegallo, 1996 [128]
4.9.4. Equus sp. from Konso, Ethiopia
5. Remarks on the Recurrence of Some Morphological Patterns
6. Conclusions
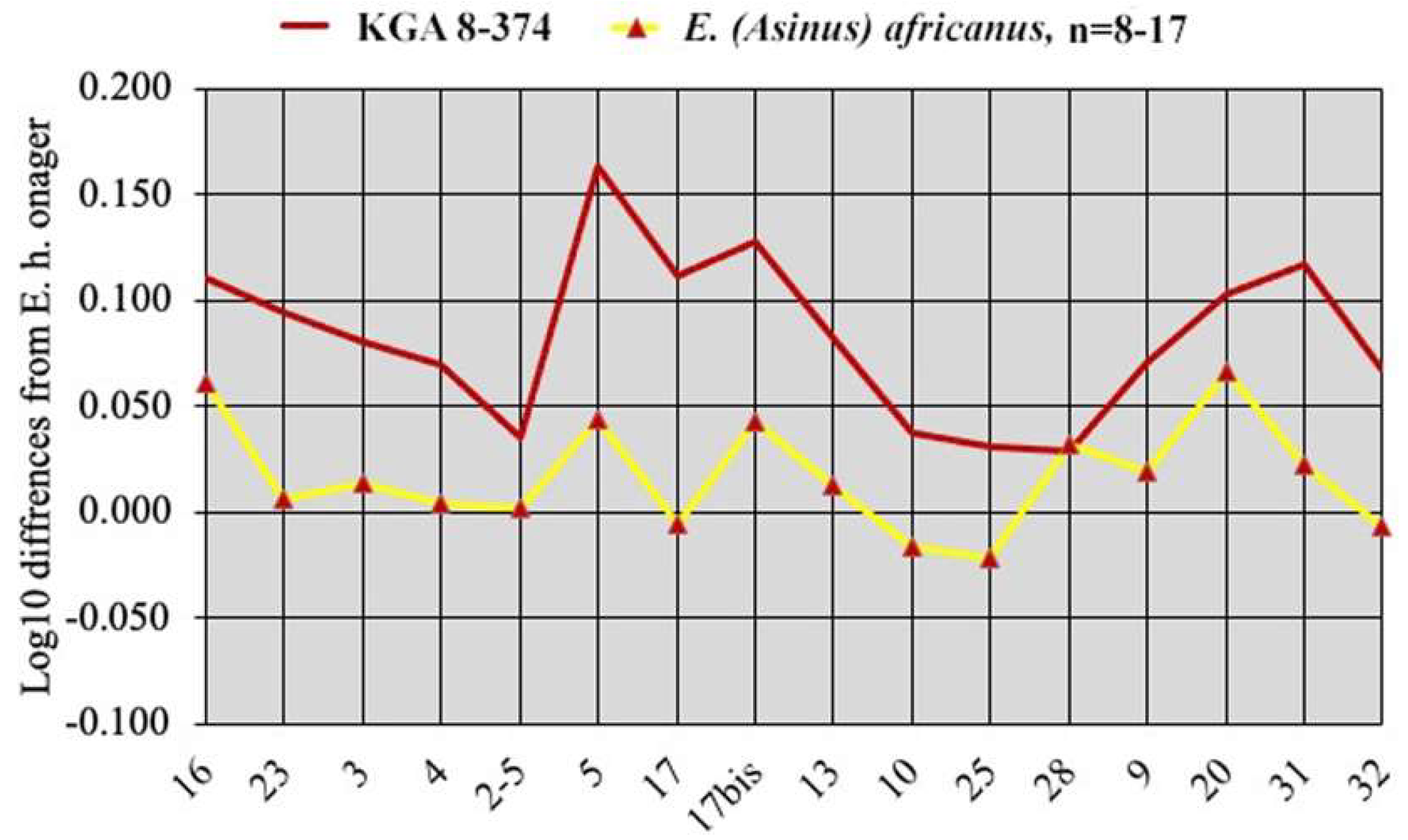
- Age of modern Equus subgeneraThe oldest fossils reliably related to extant African species are not older than 1 Ma:
- -
- The age of the fossil cranium of E. (Dolichohippus) grevyi from the Kapthurin Formation falls between 547 and 396 ka [13]; after recording and discussing the “proposed fossil occurrences of E. grevyi”, it is conclusively argued that most of them are not characteristic enough to be referred to that taxon, and certainly not those older than ca. 500 ka.
- -
- -
- -
- -
- 2.
- Age of genus Equus
- 3.
- In between 1 Ma–2 Ma Equid species
Supplementary Materials
Funding
Acknowledgments
Conflicts of Interest
References
- Churcher, C.S.; Richardson, M.L. Equidae. In Evolution of African Mammals; Maglio, V.J., Basil, H., Cooke, S., Eds.; Harvard University Press: Cambridge, MA, USA, 1978. [Google Scholar]
- MacFadden, B.J. Fossil Horses. In Systematics, Paleobiology, and Evolution of the Family Equidae; Cambridge University Press: Cambridge, UK, 1994; p. 369. [Google Scholar]
- MacFadden, B.J. Fossil Horses. In Systematics, Paleobiology, and Evolution of the Family Equidae, 2nd ed.; Cambridge University Press: Cambridge, UK, 1999; p. 369. [Google Scholar]
- Barron-Ortiz, C.I.; Avilla, L.S.; Jass, C.N.; Bravo-Cuevas, V.M.; Machado, H.; Mothé, D. What is Equus? Reconciling Taxonomy and Phylogenetic Analyses. Front. Ecol. Evol. 2019, 7, 343. [Google Scholar] [CrossRef]
- Cirilli, O.; Pandolfi, L.; Rook, L.; Bernor, R.L. Evolution of OLd World Equus and origin of the zebra-ass clade. Sci. Rep. 2021, 11, 10156. [Google Scholar] [CrossRef] [PubMed]
- Cirilli, O.; Saarinen, J.; Pandolfi, L.; Rook, L.; Bernor, R.L. An updated review on Equus stenonis (Mammalia, Perissodactyla): New implications for European early Pleistocene Equus taxonomy and paleoecology, and remarks on the Old World Equus evolution. Quat. Sci. Rev. 2021, 269, 107155. [Google Scholar] [CrossRef]
- Eisenmann, V.; Baylac, M. Extant and fossil Equus (Mammalia, Perissodactyla) skulls: A morphometric definition of the genus Equus. Zologica Scr. 2000, 29, 89–100. [Google Scholar] [CrossRef]
- Samson, P. Les Equidés fossiles de Roumanie. Geol. Romana 1975, 14, 165–352. [Google Scholar]
- Forsten, A.; Eisenmann, V. Equus (Plesippus) simplicidens (Cope), not Dolichohippus. Mammalia 1995, 59, 85–89. [Google Scholar] [CrossRef]
- Eisenmann, V.; Deng, T. Equus qingyangensis (Equidae, Perissodactyla) of the Lower Pleistocene of Bajiazui, China: Evidence for the North American origin of an Old World lineage distinct from E. stenonis. Quaternaire 2005, 2, 113–122. [Google Scholar]
- Eisenmann, V.; Delson, E. The Senèze Equids. In Senèze: Life in Central France Two Million Years Ago. Paleontology, Geochronology, Stratigraphy and Taphonomy; Delson, E., Faure, M., Guérin, C., Eds.; Springer: Cham, Switzerland.
- Bernor, R.L.; Cirilli, O.; Jukar, A.M.; Potts, R.; Buskianidze, M.; Rook, L. Evolution of Early Equus in Italy, Georgia, the Indian Subcontinent, East Africa, and the Origins of African Zebras. Front. Ecol. Evol. 2019, 7, 65. [Google Scholar] [CrossRef]
- O’Brien, K.; Tryon, C.A.; Blegen, N.; Kimeu, B.; Rowan, J.; Faith, T.J. First appearance of Grévy’s zebra (Equus grevyi), from the Middle Pleistocene Kapthurin Formation, Kenya, sheds light on the evolution and ecology of large zebras. Quat. Sci. Rev. 2021, 256, 106835. [Google Scholar] [CrossRef]
- Gkeme, A.G.; Koufos, G.D.; Kostopoulos, D.S. Reconsidering the Equids from the Early Pleistocene fauna of Apollonia 1 (Mygdonian Basin, Greece). Quaternary 2021, 4, 12. [Google Scholar] [CrossRef]
- Wang, S.-Q.; Deng, T. Some evolutionary trends of Equus eisenmannae (Mammalia, Perissodactyla) in the stratigraphic sequence of Longdan, China, in comparison to modern Equus. J. Vertebr. Paleontol. 2011, 31, 1356–1365. [Google Scholar] [CrossRef]
- Eisenmann, V.; Mashkour, M. Data base for Teeth and Limb Bones of Modern Hemiones. In Fiches d’Ostéologie Animale Pour l’Archéologie; Série B: Mammifères; Centre de Recherches Archéologiques du CNRS, APDCA: Paris, France, 2000; Volume 9, p. 46. [Google Scholar]
- Eisenmann, V. Comparative osteology of modern and fossil Horses, Halfasses and Asses. In Equids in the Ancient World; Measow, R.H., Uerpmann, H.P., Eds.; Beihefte zum Tübinger Atlas des Vorderen Orients; Reihe A: Wiesbaden, Germany, 1986; pp. 67–116. [Google Scholar]
- Simpson, G.G. Large pleistocene felines of North America. Am. Mus. Novit. 1941, 1136, 1–27. [Google Scholar]
- Bourdelle, E. Chevaux, Anes et Zèbres. Bull. Société Natl. D’acclimatation Fr. 1944, 91, 49–71. [Google Scholar]
- Bourdelle, E. Notes anatomiques et considérations zoologiques sur les Zèbres. Actes Congrès Natl. Sociétés Savantes 1934, 67, 99–112. [Google Scholar]
- Bourdelle, E. Quelques caractères anatomiques du Zèbre de Hartmann (E. zebra hartmannae Matschie). Bull. Mus. Natn. Hist. Nat. 1936, 8, 473–477. [Google Scholar]
- Orlando, L.; Metcalf, J.L.; Alberdi, M.T.; Telles-Antunes, M.; Bonjean, D.; Otte, M.; Martin, F.; Eisenmann, V.; Mashkour, M.; Morello, F.; et al. Revising the recent evolutionary history of equids using ancient DNA. Proc. Natl. Acad. Sci. USA 2009, 106, 21754–21759. [Google Scholar] [CrossRef]
- Eisenmann, V.; Turlot, J.C. Sur la taxinomie du genre Equus (Equidés). Les Cahiers de l’Analyse des Données 1978, 3, 179–201. [Google Scholar]
- Groves, C.P.; Grubb, P. Ungulate Taxonomy; The Johns Hopkins University Press: Baltimore, MD, USA, 2011; p. 309. [Google Scholar]
- Jónsson, H.; Schubert, M.; Seguin-Orlando, A.; Ginolhac, A.; Petersen, L.; Fumagalli, M.; Albrechtsen, A.; Petersen, B.; Korneliussen, T.S.; Vilstrup, J.T.; et al. Speciation with gene flow in equids despite extensive chromosomal plasticity. Proc. Natl. Acad. Sci. USA 2014, 111, 18655–18660. [Google Scholar] [CrossRef]
- Bennett, E.A.; Champlot, S.; Peter, J.; Arbuckle, B.; Guimaraes, S.; Pruvost, M.; Bar-David, S.; Davis, S.; Gautier, M.; Kaczensky, P.; et al. Taming the Late Quaternary phylogeography of the Eurasiatic wild ass through ancient and modern DNA. Proc. Natl. Acad. Sci. USA 2009, 106, 21754–21759, Erratum in PLoS ONE 2017, 12. [Google Scholar] [CrossRef]
- Groves, C.P.; Willoughby, D.P. Studies on the taxonomy and phylogeny of the genus Equus. Mammalia 1981, 45, 321–354. [Google Scholar] [CrossRef]
- Eisenmann, V. Pliocene and Pleistocene Equids: Paleontology versus Molecular Biology. In Late Neogene and Quaternary Biodiversity and Evolution, Regional Developments and Interregional Correlations. Proceedings Volume of the 18th International Senckenberg Conference (VI International Palaeontological Colloquium in Weimar), Weimar, Germany, 20–25 April 2004; Kahlke, R.-D., Maul, L.-C., Mazza, P., Eds.; Courier Forschungsinstitut Senckenberg: Frankfurt, Germany, 2006; p. 256. [Google Scholar]
- Eisenmann, V.; Howe, J.; Pichardo, M. Old World Hemiones and New World slender species (Mammalia, Equidae). Palaeovertebrata 2008, 36, 159–233. [Google Scholar] [CrossRef]
- Vilstrup, J.T.; Seguin-Orlando, A.; Stiller, M.; Ginolhac, A.; Raghavan, M.; Nielsen, S.C.A.; Weintstock, J.; Froese, D.; Vasiliev, S.K.; Ovodov, N.D.; et al. Mitochondrial phylogenomics of modern and ancient. PLoS ONE 2013, 8, e55950. [Google Scholar] [CrossRef]
- Eisenmann, V. Etude des cornets des dents incisives inférieures des Equus actuels et fossiles. Palaeontogr. Ital. 1979, 71, 55–75. [Google Scholar]
- Orlando, L.; Mashkour, M.; Burke, A.; Douady, C.J.; Eisenmann, V. Geographic distribution of an extinct equid (E. hydruntinus, Mammalia, Equidae) revealed by morphological and genetical analyses of fossils. Mol. Ecol. 2006, 15, 2083–2093. [Google Scholar] [CrossRef]
- Burke, A.; Eisenmann, V.; Ambler, G. The systematic position of Equus hydruntinus, an extinct species of Pleistocene equid. Quat. Res. 2003, 59, 459–469. [Google Scholar] [CrossRef]
- Eisenmann, V.; Mashkour, M. The small equids of Binagady (Azerbaidjan) and Qazvin (Iran): E. hemionus binagadensis nov. subsp., and E. hydruntinus. Géobios 1999, 32, 105–122. [Google Scholar] [CrossRef]
- Bonifay, M.-F. Equus hydruntinus Regalia minor n.ssp. from the caves of Lunel-Viel (Hérault, France). In Equids In the Ancient World; Meadow, R.H., Uerpmann, H.-P., Eds.; Beihefte zum Tübinger Atlas des Vorderen Orients, Reihe A (Naturwissenschaften), Nr. 19/2; Dr Ludwig Reichert Verlag: Wiesbaden, Germany, 1991; Volume 2. [Google Scholar]
- van Asperen, E.N.; Stefaniak, K.; Proskurnyak, L.; Ridush, B. Equids from Emine-Bair-Khosar cave (Crimea, Ukraine): Co-occurrence of the stenonid Equus hydruntinus and the caballoid E. ferus latipes based on skull and postcranial remains. Palaeontol. Electron. 2012, 15, 28. [Google Scholar] [CrossRef]
- Boulbes, N.; van Asperen, E.N. Biostratigraphy and Palaeoecology of European Equus. Front. Ecol. Evol. 2019, 7, 301. [Google Scholar] [CrossRef]
- Alberdi, M.T.; Palombo, M.R. The Early Pleistocene Equidae from Pirro Nord (Apricena, Southern Italy). Paleontographica, Abt. A: Paleozoolgy Stratigr. 2013, 298, 147–167. [Google Scholar] [CrossRef]
- Tsoukala, E. Contribution to the study of the Pleistocene fauna of large mammals (Carnivora, Perissodactyla, Artiodactyla) from Petralona Cave (Chalkidiki, N. Greece). Preliminary report. C. R. Acad. Sci. 1991, 312, 331–336. [Google Scholar]
- Stehlin, H.G.; Graziosi, P. Ricerche sugli Asinidi fossili d’Europa. Mémoires Société Paléontologique Suisse 1935, 56, 1–73. [Google Scholar]
- Antunes, M.T. The Zebro (Equidae) and its extinction in Portugal, with an Appendix on the noun zebro and the modern “zebra”. 9th ICAZ Conference, Durham 2002. In Equids in Time and Space; Mashkour, M., Ed.; Oxbow Books: Oxford, UK, 2006; Volume 17, pp. 211–236. [Google Scholar]
- Blanc, G.A. Sulla esistenza di “Equus (Asinus) hydruntinus” Regalia nel Pleistocene del Nord Africa. Bolletino Soc. Geol. Ital. 1956, 75, 3–14. [Google Scholar]
- Bibikova, V.I.; Vereshchagin, N.K.; Garutt, V.E.; Iourv, K.B. Novye materialy po chetvertichnoj faune Zabaijkalia. Mater. Arkheologiii SSSR 1953, 39, 463–475. [Google Scholar]
- Vogt, T.; Erbajeva, H. Premières preuves de conditions périglaciaires au Pléistocène inférieur en Transbaïkalie (Sibérie, Russie). C. R. Acad. Sci. Paris 1995, 320, 861–866. [Google Scholar]
- Foronova, I. Late quaternary equids (genus Equus) of South-western and South-central Siberia. 9th ICAZ Conference, Durham 2002. In Equids in Time and Space; Mashkour, M., Ed.; Oxbow Books: Oxford, UK, 2006; Volume 2, pp. 20–30. [Google Scholar]
- Boule, M.; de Chardin, T.P. Paléontologie, Equus hemionus. In Le paléolithique de la Chine; Archives de l’institut de Paléontologie Humaine; Masson: Paris, France, 1928; Volume 4, pp. 36–46. [Google Scholar]
- Zhou, X.; Sun, Y.; Wang, Z.; Wang, H. Dalian Gulongshan Yizhi Yanjiu (Gulongshan Cave Site, A Upper Paleolithic site at Dalian City); Beijing Kexue Jishu Chubanshe (Beijing Scientific and Technical Publishing House): Beijing, China, 1990; p. 94. [Google Scholar]
- Sam, Y. African origins of modern asses as seen from paleontology and DNA: What about the Atlas wild ass? Geobios 2020, 58, 73–84. [Google Scholar] [CrossRef]
- Ducos, P. The Equid from Tell Muraibit, Syria. In Equids in the Ancient World; Meadow, R.H., Uerpmann, H.P., Eds.; Beihefte zum Tübinger Atlas des Vorderen Orients; Reihe A.: Wiesbaden, Germany, 1986; pp. 237–245. [Google Scholar]
- Uerpmann, H.-P. Equus africanus in Arabia. In Equids in the Ancient World, Vol. II; Meadow, R.H., Uerpmann, H.-P., Eds.; Beihefte zum Tübinger Atlas des Vorderen Orients; Reihe A (Naturwissenschaften); Dr. Ludwig Reichert Verlag: Wiesbaden, Germany, 1991; pp. 12–33. [Google Scholar]
- Astre, G. Petit Asinien Pléistocène du Lauraguais. Bull. Société D’histoire Nat. Toulouse 1948, 83, 118–128. [Google Scholar]
- Eisenmann, V. Les Chevaux (Equus Sensu Lato) Fossiles et Actuels: Crânes et Dents Jugales Supérieures. Cahiers de Paléontologie; Editions du CNRS: Paris, France, 1980; 186p. [Google Scholar]
- Eisenmann, V. Etude des dents jugales inférieures des Equus actuels et fossiles. Palaeovertebrata 1981, 10, 3–4. [Google Scholar]
- Groves, C.P. Taxonomy. In Sull’asino Selvatico Africano; Groves, C.P., Ziccardi, F., Toschi, A., Eds.; Supplemento alle Ricerche di Zoologia Applicata alla Caccia; Universita di Bologna: Bologna, Italy, 1966; Volume 5, pp. 2–11. [Google Scholar]
- Thomas, P. Recherches stratigraphiques et paléontologiques sur quelques formations d’eau douce de l’Algérie. Mémoires Société Géologique Fr. 1884, 3, 1–51. [Google Scholar]
- Boule, M. Observations sur quelques équidés fossiles. Bull. Société Géologique Fr. 1900, 27, 531–542. [Google Scholar]
- Bagtache, B.; Hadjouis, D.; Eisenmann, V. Présence d’un Equus caballin (E. algericus n. sp.) et d’une autre espèce nouvelle d’Equus (E. melkiensis n. sp.) dans l’Atérien des Allobroges, Algérie. C. R. Ac. Sc. Paris 1984, 298, 609–612. [Google Scholar]
- Ginsburg, L.; Hilly, J.; Taquet, P. Une faune würmienne dans un remplissage de fente du massif du Filfila (littoral nord-constantinois, Algérie). C. R. Somm. Séances Soc. Géol. Fr. 1968, 5, 157. [Google Scholar]
- Souville, G. Atlas Préhistorique de l’Algérie, Feuille N°5P; Jourdan, A., Ed.; Gouvernement Général de l’Algérie: Alger, Algérie, 1958. [Google Scholar]
- Ennouchi, E. La faune des limons rouges de Bou-Knadel (Maroc). In Actes du IVème Congrès International du Quaternaire: 3–4; Août—Septembre: Rome, Italy, 1953. [Google Scholar]
- Arambourg, C. Mammifères fossiles du Maroc. Mémoires Soc. Sci. Nat. Maroc 1938, 46, 1–74. [Google Scholar]
- Tchernov, E. An attempt to synchronize the faunal changes with the radiometricdates and the cultural chronology in Southwest Asia. In Archaeozoology of the Near East III; ARC Publicaties: Groningen, The Netherlands, 1998; Volume 18, pp. 7–44. [Google Scholar]
- Marom, N.; Lazagabaster, I.A.; Horwitz, L.K. The late middle pleistocene mammalian fauna of Oumm Qatafa cave, Judean desert: Taphonomy and paleoenvironment. J. Quat. Sci. 2022, 37, 612–638. [Google Scholar] [CrossRef]
- Dodonov, A.; Zhegallo Penkov, V.A.; Maria Sotnikova, M. Stratigrafia mestonakhozhdenij pozdnepliotsenovykh pozvonochnykh Iuzhnogo Tadjikistana. Izvestia Akademii Nauk SSSR Serija Geologicheskaja 1991, 5, 12–20. (In Russian) [Google Scholar]
- Sotnikova, M.V.; Baigusheva, V.S.; Titov, V.V. Carnivores of the Khapry faunal assemblage and their stratigraphic implications. Stratigr. Geol. Correl. 2002, 10, 375–390. [Google Scholar]
- Gromova, V. Istorija Loshadej (roda Equus) v Starom Svete. Chast’ 1. Obzor i Opisanie Form; Trudy Paleontological Institute Akademi Nauk SSSR: Moscow, Russia, 1949; Volume 17, p. 373. (In Russian) [Google Scholar]
- Hadjouis, D. La faune mammaliennne d’Algérie, un renouvellement constant. Préhistoire Algérienne. Dossiers Archéol. 2003, 282, 42–53. [Google Scholar]
- Aouadi, N.; Bkhairi, A.; Belhouchhet, L.; Khedaer-El-Asmi, R. Les sites préhistoriques de l’Aïn Metherchem (Foussana, Kasserine) revisités. Bull. Mus. Anthropol. préhist. Monaco 2014, 54, 137–151. [Google Scholar]
- Van Couvering, J.A.; Delson, E. African land mammal ages. J. Vertebr. Paleontol. 2020, 40, e1803340. [Google Scholar] [CrossRef]
- Azzaroli, A. On a Late Pleistocene Ass from Tuscany; with notes on the History of Asses. Palaeontographia Italica 1979, 71, 27–47. [Google Scholar]
- Azzaroli, A. Pleistocene and living horses of the Old World. Palaeontogr. Ital. 1966, 61, 1–15. [Google Scholar]
- Koufos, G.D.; Kostopoulos, D.S.; Sylvestrou, I.A. Equus apolloniensis n. sp. (Mammalia, Equidae) from the latest Villafranchian locality of Apollonia, Macedonia, Greece. Paleontol. Evol. 1997, 30–31, 49–76. [Google Scholar]
- Kahlke, R.D.; García, N.; Kostopoulos, D.S.; Lacombat, F.; Lister, A.M.; Mazza, P.P.A.; Spassov, N.; Titov, V.V. Western Palaearctic palaeoenvironmental conditions during the Early and early Middle Pleistocene inferred from large mammal communities, and implications for hominin dispersal in Europe. Quat. Sci. Rev. 2011, 30, 1368–1395. [Google Scholar] [CrossRef]
- Eisenmann, V. Le protostylide: Valeur systématique et signification phylétique chez les espèces actuelles et fossiles du genre Equus (Perissodactyla, Mammalia). Z. Säugetierkunde 1976, 41, 349–365. [Google Scholar]
- Eisenmann, V. Equus capensis (Mammalia, Perissodactyla) from Elandsfontein. Palaeontol. Afr. 2000, 36, 91–96. [Google Scholar]
- Geraads, D.; Hublin, J.J.; Jaeger, J.J.; Tong, T.; Sen, S.; Toureau, P. The Pleistocene hominid site of Ternifine, Algeria: New results on the environment, age, and human industries. Quat. Res. 1986, 25, 380–386. [Google Scholar] [CrossRef]
- Potts, R.; Behrensmeyer, A.K.; Tyler Faith, J.; Tryon, C.A.; Brooks, A.S.; Yellen, J.E. Environmental dynamics during the onset of the Middle Stone Age in eastern Africa. Science 2018, 360, 86–90. [Google Scholar] [CrossRef]
- Churcher, C.S.; Hooijer, D.A. The Olduvai zebra (Equus oldowayensis) from the Later Omo Beds, Ethiopia. Zool. Meded. Rijksmus. Nat. Hist. Leiden 1980, 55, 265–280. [Google Scholar]
- Eisenmann, V.; De Giuli, C. Caractères distinctifs entre vrais Zèbres (Equus zebra) et Zèbres de Chapman (Equus burchelli antiquorum) d’après l’étude de 60 têtes osseuses. Mammalia 1974, 38, 509–543. [Google Scholar] [CrossRef]
- Lundholm, B. Equus zebra greadheadi n. subsp., a new South African fossil zebra. Ann. Transvaal Mus. 1952, 22, 25–27. [Google Scholar]
- Lazarev, P.A. Antropogenovye Loshadi Iakutii; Nauka: Moscow, Russia, 1980; p. 190. (In Russian) [Google Scholar]
- Kuzmina, I.E. Late Pleistocene horse of the Ussurian region. Trudy Zool. Inst. Akad. Nauk SSSR. 1990, 212, 49–59. (In Russian) [Google Scholar]
- Cardoso, J.L. Les grands Mammifères du Pléistocène supérieur du Portugal. Essai de synthèse. Géobios 1996, 29, 235–250. [Google Scholar] [CrossRef]
- Eisenmann, V.; Crégut-Bonnoure, E.; Moigne, A.-M. Equus mosbachensis et les grands Chevaux de la Caune de l’Arago et de Lunel-Viel: Crâniologie comparée. Bull. Mus. Nat. Hist. Nat. 1985, 7, 157–173. [Google Scholar]
- Eisenmann, V. Proportions squelettiques de Chevaux quaternaires et actuels. Géobios 1991, 13, 25–32. [Google Scholar] [CrossRef]
- Madriaga de la Campa, B. Estudio zootecnico de las pinturas rupestres en la region cantabrica. Zephyrus 1963, XIV, 29–45. [Google Scholar]
- Eisenmann, V.; David, F. Evolution de la taille des Chevaux d’Arcy-sur-Cure et de quelques autres Chevaux quaternaires. In L’Aurignacien de la Grotte du Renne; XXXIVème Supplément à Gallia Préhistoire; CNRS: Paris, France, 2002; pp. 97–102. [Google Scholar]
- Turbanti, L. Equus Caballus nel Pleistocene delle Puglie; Università degli Studi di Firenze: Firenze, Italy, 1982; Volume 78. [Google Scholar]
- Eisenmann, V. Discriminating Equus skulls: The Franck’s Index and the new Palatal Index. 9th ICAZ Conference, Durham 2002. In Equids in Time and Space; Mashkour, M., Ed.; Oxbow Books: Oxford, UK, 2006; pp. 172–182. [Google Scholar]
- Eisenmann, V. Origins, dispersals, and migrations of Equus (Mammalia, Perissodactyla). Cour. Forsch. Senckenberg 1992, 153, 161–170. [Google Scholar]
- Vangenheim, E.A.; Zazhigin, V.S. Obzor faunisticheskikh kompleksov i faun territorii SSSR. In Chetvertichnaya Sistema, 1; Shantser, E.V., Ed.; Nedra: Moscow, Russia, 1982; pp. 267–279. (In Russian) [Google Scholar]
- Forsten, A.; Ziegler, R. The horses (Mammalia, Equidae) from the early Wuermian of Villa Seckendorff, Stuttgart-Bad Cannstadt, Germany. Stuttg. Beiträge Nat. Ser. B 1995, 224, 1–22. [Google Scholar]
- Nehring, A. Fossile Pferde aus Deutschen Diluvial-Ablagerungen und ihre Beziehungen zu den lebenden Pferden: Ein Beitrag zur Geschichte des Hauspferdes. Landwirtsch. Jahrb. 1884, 13, 81–160. [Google Scholar]
- Philippe, M. La faune würmienne du gisement paléontologique de Siréjol à Gignac (Lot). Bull. Société Sci. Hist. Archéologique Corrèze 1975, 97, 1–9. [Google Scholar] [CrossRef]
- Cardoso, J.L.; Eisenmann, V. Equus caballus antunesi, nouvelle sous-espèce quaternaire du Portugal. Palaeovertebrata 1989, 19, 47–72. [Google Scholar]
- Eisenmann, V. Sussemionus, a new subgenus of Equus (Perissodactyla, Mammalia). C. R. Biol. 2010, 333, 235–240. [Google Scholar] [CrossRef]
- Saarinen, J.; Cirilli, O.; Strani, F.; Meshida, K.; Bernor, R.L. Testing Equid Body Mass Estimate Equations on Modern Zebras—With Implications to Understanding the Relationship of Body Size, Diet, and Habitats of Equus in the Pleistocene of Europe. Front. Ecol. Evol. 2021, 9, 622412. [Google Scholar] [CrossRef]
- Sher, A.V. Mlekopitaiushchie i Stratigrafia Pleistotsena Krainego Severo-Vostoka SSSR i Severnoi Ameriki; Nauka: Moscow, Russia, 1971; p. 310. (In Russsian) [Google Scholar]
- Sher, A.V. Olyorian land mammal age of Northeastern Siberia. Palaeontogr. Ital. 1987, 74, 97–112. [Google Scholar]
- Wagner, G.A.; Maul, L.C.; Löscher, M.; Schreiber, H.D. Mauer—The type site of Homo heidelbergensis: Palaeoenvironment and age. Quat. Sci. Rev. 2011, 30, 1464–1473. [Google Scholar] [CrossRef]
- Maul, L.C.; Parfitt, S.A. Micromammals from the 1995 Mammoth Excavation at West Runton, Norfolk, UK: Morphometric data, biostratigraphy and taxonomic reappraisal. Quat. Int. 2010, 228, 91–115. [Google Scholar] [CrossRef][Green Version]
- Musil, R. Die Equidenreste aus dem Pleistozän von Süssenborn bei Weimar. Paläontol. Abhandl. A 1969, 3–4, 617–666. [Google Scholar]
- Wüst, E. Untersuchungen über das Pliozän und das älteste Pleistozän Thüringens Nördlich vom Thüringer Walde und Westlich von der Saale. Abhandl. Natur. Gesellschaft Halle. 1901, XXIII, 566. [Google Scholar]
- Vekua, A.K. The Lower Pleistocene Mammalian Fauna of Akhalkalaki (Southern Georgia, USSR). Palaeontogr. Ital. 1986, 74, 63–96. [Google Scholar]
- Von Reichenau, W. Beiträge zur näheren Kenntnis fossiler Pferde aus deutschen Pleistozän, insbesondere über die Entwicklung und die Abkaustadien des Gebisses vom Hochterrassenpferd (Equus mosbachensis v.R.). Abh. Grossherz.-Hess. Geol. Landesanst. Darmstadt 1915, 7, 155. [Google Scholar]
- Alberdi, M.T.; Palombo, M.R. The late Early to early Middle Pleistocene stenonoid horses from Italy. Quat. Int. 2013, 288, 25–44. [Google Scholar] [CrossRef]
- Arzarello, M.; Peretto, C.; Moncel, M.-H. The Pirro Nord site (Apricena, Fg, Southern Italy) in the context of the first European peopling: Convergences and divergences. Quat. Int. 2015, 389, 255–263. [Google Scholar] [CrossRef]
- Maul, L.; Masini, F.; Parfitt, S.A.; Rekovetz, L.; Savorelli, A. Evolutionary trends in arvicolids and the endemic murid Mikrotia—New data and a critical overview. Quat. Sci. Rev. 2014, 96, 240–258. [Google Scholar] [CrossRef]
- Ortiz, J.E.; Torres, T.; Ramon, J.; Delgado, A.; Llamas, J.; Soler, V.; Delgado, J. Numerical dating algorithm of amino acid racemization ratios from continental ostracodes. Application to the Guadix-Baza Basin (southern Spain). Qyaternary Sci. Rev. 2004, 23, 717–730. [Google Scholar] [CrossRef]
- Alberdi, M.T.; Ruiz Bustos, A. Descripcion y significado biostratigrafico y climatico del Equus e Hippopotamus, en el yacimiento de Venta Micena (Granada). Estud. Geol. 1985, 41, 251–261. [Google Scholar] [CrossRef]
- Duval, M.; Falguères, C.; Jean-Jacques Bahain, J.-J. Age of the oldest hominin sttelements in Spain: Contribution of the combined U-series/ESR dating method applied tyo fossil teeth. Quat. Geochronol. 2012, 10, 412–417. [Google Scholar] [CrossRef]
- De Giuli, C.; Masini, F.; Torre, D. The latest villafranchian faunas in Italy: The Pirro Nord fauna (Apricena, Gargano). Palaeontogr. Ital. 1987, 74, 51–62. [Google Scholar]
- De Giuli, C. Late villafranchian faunas of Italy: The Selvella local fauna in the southern Chiana valley—Umbria. Palaeontogr. Ital. 1987, 74, 11–50. [Google Scholar]
- Musil, R. Die Equiden-Reste aus dem Unterpleistozän von Untermaßfeld. In Das Pleistozän von Untermassfeld bei Meiningen (Thüringen); Kahlke, R.-D., Ed.; Teil 2; Monographien des Römisch-Germanischen Zentralmuseums: Mainz, Germany, 2001; Volume 40, pp. 557–587. [Google Scholar]
- Kahlke, R.D.; Linnermann, U.; Gärtner, A. New results on the origin and geological history of the Early Pleistocene site of Untermassfeld. In The Pleistocene of Untermassfeld Near Meiningen (Thüringen, Germany); Kahlke, R.-D., Ed.; Part 4; Monographien des Römisch-Germanischen Zentralmuseums: Mainz, Germany, 2020; Volume 40, pp. 1079–1104. [Google Scholar]
- Eisenmann, V.; Boulbes, N. New results on equids from the Early Pleistocene site of Untermassfeld. In The Pleistocene of Untermassfeld Near Meiningen (Thüringen, Germany); Kahlke, R.-D., Ed.; Part 4; Monographien des Römisch-Germanischen Zentralmuseums: Mainz, Germany, 2020; Volume 40, pp. 1295–1322. [Google Scholar]
- Eisenmann, V.; Vasiliev, S. Unexpected finding of a new Equus species (Mammalia, Perissodactyla) belonging to a supposedly extinct subgenus in late Pleistocene deposits of Khakassia (southwestern Siberia). Geodiversitas 2011, 33, 519–530. [Google Scholar] [CrossRef]
- Lordkipanidze, D.; Jashashvili, T.; Vekua, A.; Ponce de Leon, M.S.; Zollikofer, C.P.E.; Rightmire, G.P.; Pontzer, H.; Ferring, R.; Oms, O.; Tappen, M.; et al. Postcranial evidence from early Homo from Dmanisi, Georgia. Nature 2007, 449, 305–310. [Google Scholar] [CrossRef]
- Ferring, R.; Oms, O.; Agusti, J.; Berna, F.; Nioradze, M.; Shelia, T.; Tappen, M.; Vekua, A.; Zhvania, D.; Lordkipanidze, D. Earliest human occupations at Dmanisi (Georgian Caucasus) dated to 1.85–1.78 Ma. Proc. Natl. Acad. Sci. USA 2011, 108, 10432–10436. [Google Scholar] [CrossRef]
- Bernor, R.L.; Cirilli, O.; Buskianidze, M.; Lordkipanidze, D. The Dmanisi Equus: Systematics, biogeography, and paleoecology. J. Hum. Evol. 2021, 158, 1–14. [Google Scholar] [CrossRef]
- Vekua, A.K. Die Wirbeltierfauna des Villafranchium von Dmanisi und ihre biostratigraphische Bedeutung. Jahrb. Des Römisch-Ger. Zent. Mainz 1995, 42, 180. [Google Scholar]
- Downs, T.; Miller, G.J. Late Cenozoic Equids from Anza-Borrego Desert of Clifornia. Contrib. Sci. Nat. Hist. Mus. Los Angeles Cty. 1994, 440, 90. [Google Scholar]
- Arambourg, C. Les Vertébrés du Pléistocène de l’Afrique du Nord. Arch. Mus. Nat. Hist. Nat. 1970, 10, 1–127. [Google Scholar]
- Geraads, D.; Raynal, J.-P.; Eisenmann, V. The earliest human occupation of North Africa: A reply to Sahnouni et al. (2002). J. Hum. Evol. 2004, 46, 751–761. [Google Scholar] [CrossRef]
- Duval, M.; Sahnoini, M.; Parés, J.M.; van der Made, J.; Abdessadok, S.; Harichane, Z.; Chebeb, R.C.; Boulaghraif, K.; Pérez-Gonzales, A. The Plio-Pleistocene sequence of Oued Boucherit (Algeria): A unique chronologically—Constrained archaeological and palaeontological record in North Africa. Quat. Sci. Rev. 2021, 271, 1–23. [Google Scholar] [CrossRef]
- Eisenmann, V. Les métapodes d’Equus sensu lato (Mammalia, Perissodactyla). Géobios 1979, 12, 863–886. [Google Scholar] [CrossRef]
- Aouadi, N. Les Chevaux Fossiles Nord-Africains. Nouvelle Analyse, Nouvelle Phylogénie; Ministère de la Culture, de la Jeuness et des Loisirs: République Tunisienne, Africa, 2004; pp. 5–16. [Google Scholar]
- Kuznetsova, T.V.; Zhegallo, V.I. Taksonomicheskoie Raznoobrazie Ekvid Mestonakhojdenia Nalaikha (Mongolia); Trudy Mejdunarodnogo soveshchania ’Sostoianie Teriofauny v Rossii i Blijnem Zarubejie’ 1-3 fevralia 1995; Nauka: Moscow, Russia, 1996; pp. 48–53. (In Rusian) [Google Scholar]
- Eisenmann, V.; Kuznetsova, T. Early Pleistocene equids (Mammalia, Perissodactyla) of Nalaikha, Mongolia, and the emergence of modern Equus Linnaeus 1758. Geodiversitas 2004, 26, 535–561. [Google Scholar]
- Nakaya, H.; Suwa, G.; Ode, S.; Asfaw, B.; Beyene, Y. A near-complete skull of Equus (Equidae, Perissodactyla, Mammalia) from the Early Pleistocene Konso Formation, Southern Ethiopia. In Proceedings of the Program and Abstracts 73rd Annual Meeting, Society of Vertebrate Paleontology, Los Angeles, CA, USA, 2 November 2013. [Google Scholar]
- Suwa, G.; Nakaya, H.; Asfaw, B.; Saegusa, H.; Amzaye, A.; Kono, R.T.; Yonas Beyene, Y.; Katoh, S. Plio-Pleistocene terrestrial mammal assemblage from Konso, Southern Ethiopia. J. Vertebr. Paleontol. 2003, 23, 901–916. [Google Scholar] [CrossRef]
- Suwa, G.; Asfaw, B.; Katoh, S.; Nakaya, H.; Beyene, Y. Early Pleistocene fauna and paleoenvironments at Konso, Ethiopia. In African Paleoecology and Human Evolution; Reynolds, S., Bobe, R., Eds.; Cambridge University Press: Cambridge, MA, USA, 2022; pp. 269–277. [Google Scholar]
- Bar-Yosef, O.; Belmaker, M. Archeology of Human Evolution. In Quaternary of the Levant, Environments, Climate Change, and Humans; Cambridge Uniersity Press: Cambridge, MA, USA, 2017; pp. 179–186. [Google Scholar]
- Kahlke, R.D. Typencatalog zu den Sammlungen des Instituts für Quartaärpaläontologie Weimar. Abh. Ber. Mus. Not. Gotha. 1984, 12, 57–63. [Google Scholar]
- Schwartz, E. Über diluviale Pferde der Equus caballus-Gruppe. Jahrbuch Preussischen Geologischen Landesanstalt 1928, 48, 429–476. [Google Scholar]
































































































Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eisenmann, V. Old World Fossil Equus (Perissodactyla, Mammalia), Extant Wild Relatives and Incertae Sedis Forms. Quaternary 2022, 5, 38. https://doi.org/10.3390/quat5030038
Eisenmann V. Old World Fossil Equus (Perissodactyla, Mammalia), Extant Wild Relatives and Incertae Sedis Forms. Quaternary. 2022; 5(3):38. https://doi.org/10.3390/quat5030038
Chicago/Turabian StyleEisenmann, Vera. 2022. "Old World Fossil Equus (Perissodactyla, Mammalia), Extant Wild Relatives and Incertae Sedis Forms" Quaternary 5, no. 3: 38. https://doi.org/10.3390/quat5030038
APA StyleEisenmann, V. (2022). Old World Fossil Equus (Perissodactyla, Mammalia), Extant Wild Relatives and Incertae Sedis Forms. Quaternary, 5(3), 38. https://doi.org/10.3390/quat5030038