3.2.1. Canis etruscus Forsyth Major, 1877
Synonyms. 1997 Xenocyon sp. Koufos and Kostopoulos, p. 48. (partim)
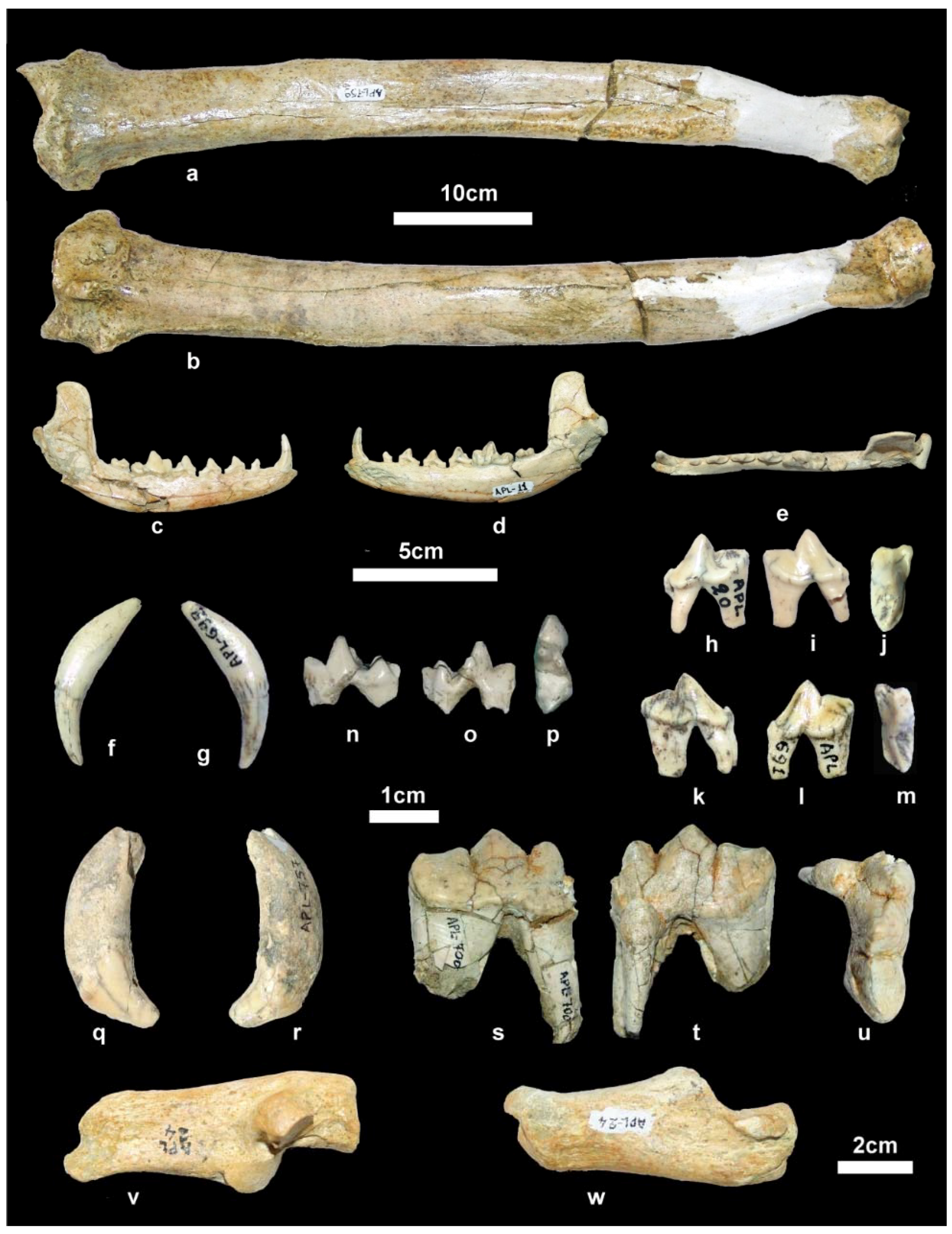
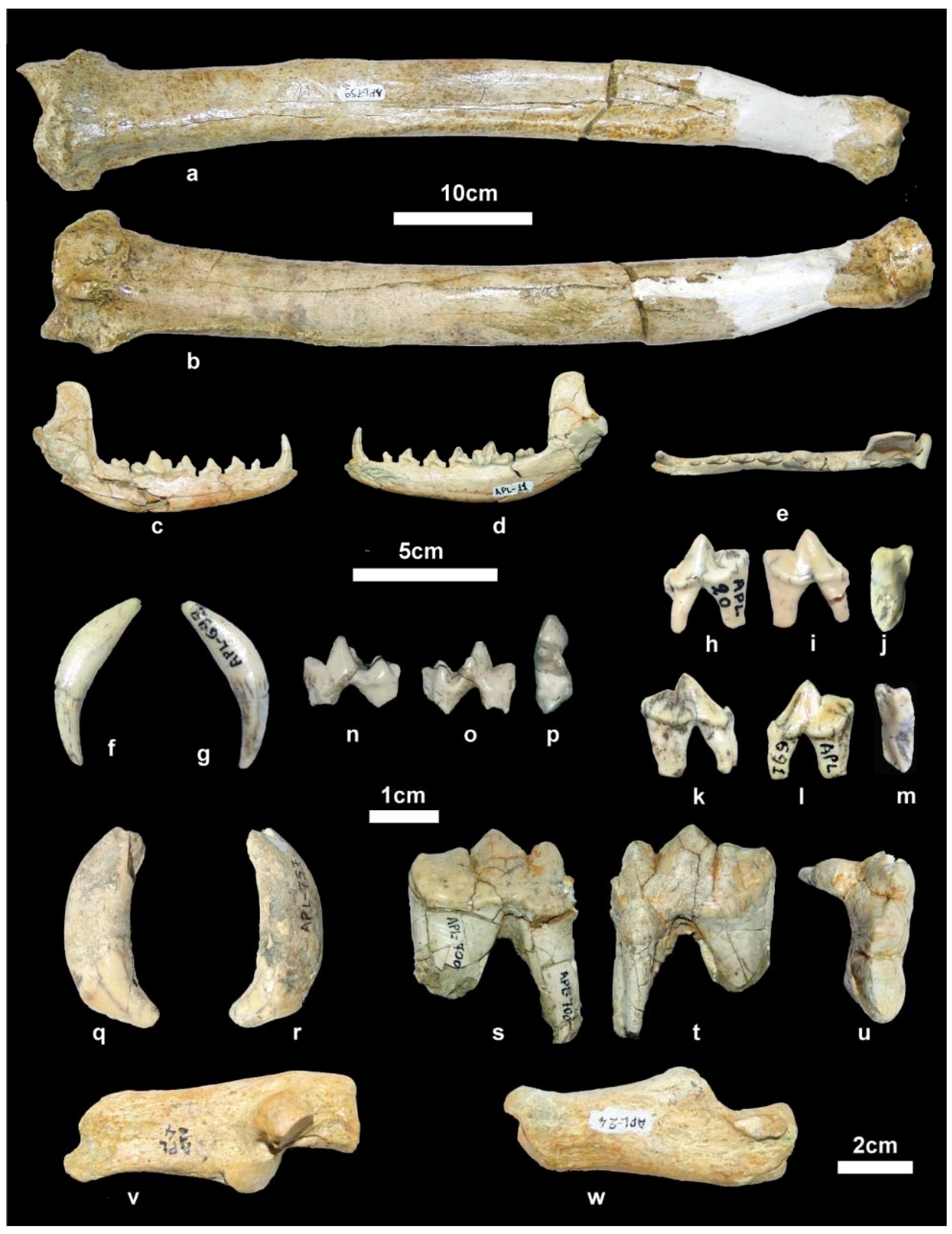
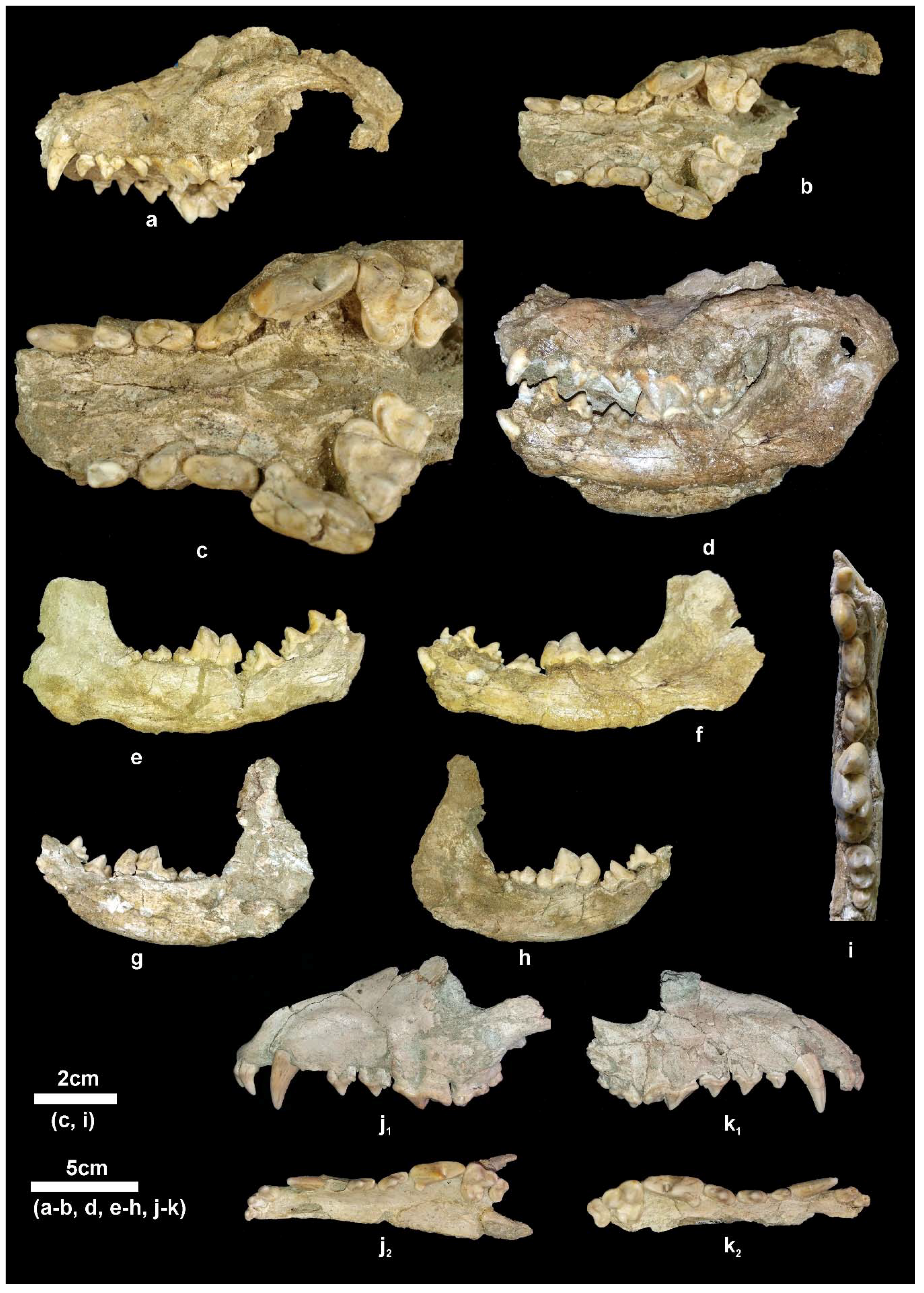
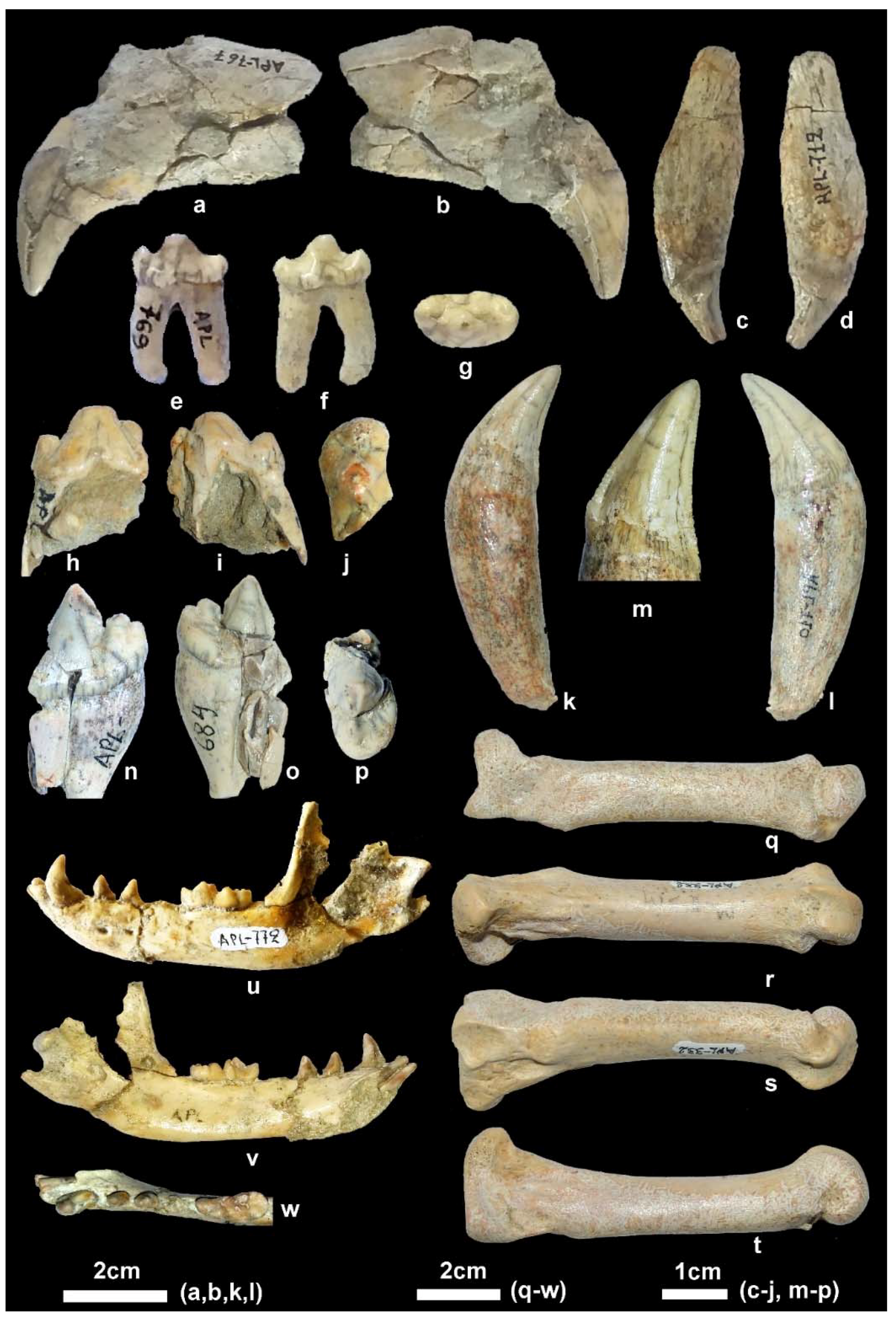
Material. New collection: Right mandibular fragment with p2-m2, APL-689.
Old collection: Skull, APL-522; Right and left mandibular fragments with c-m2, APL-526; left mandibular fragment with p2-m1, APL-569.
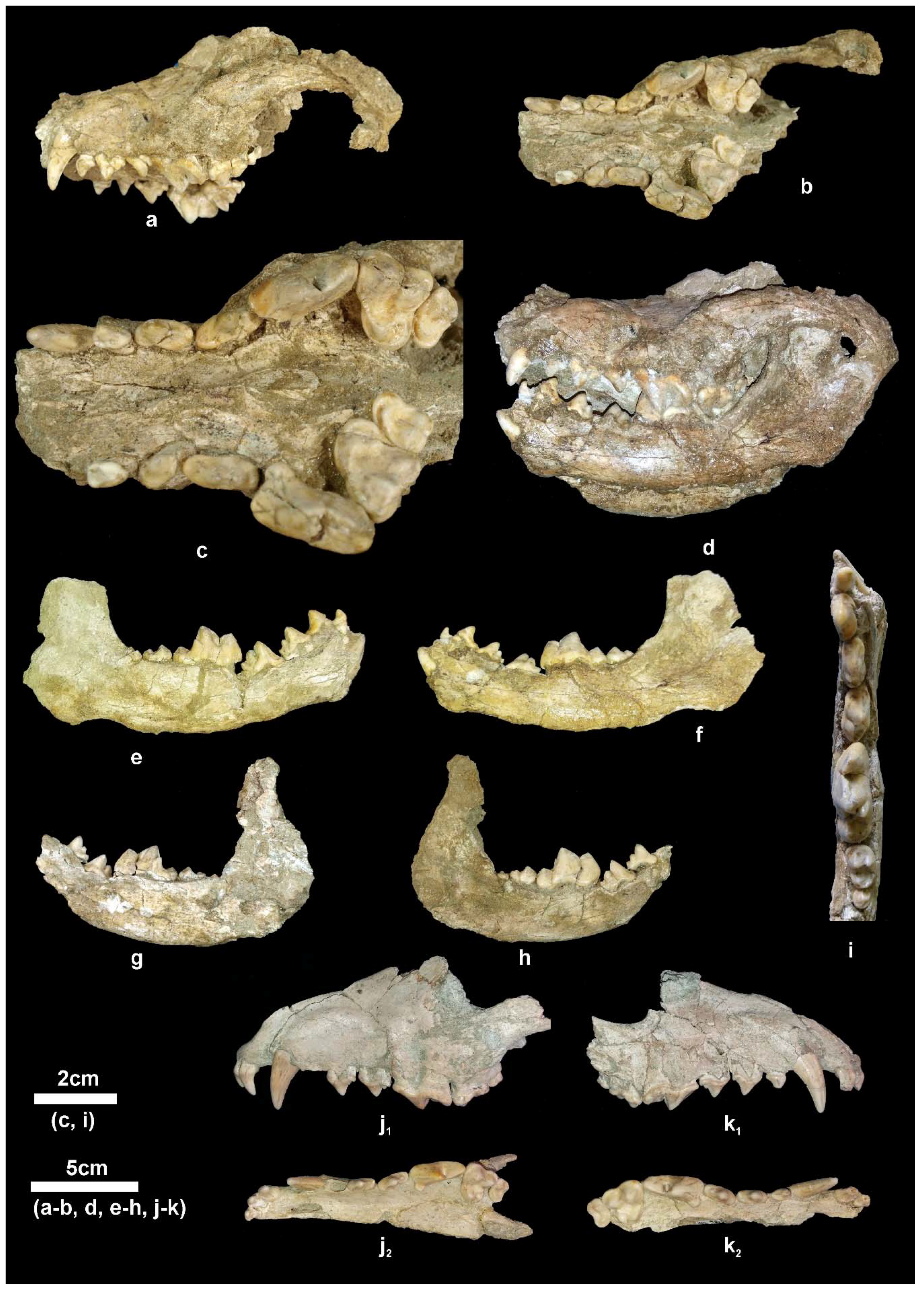
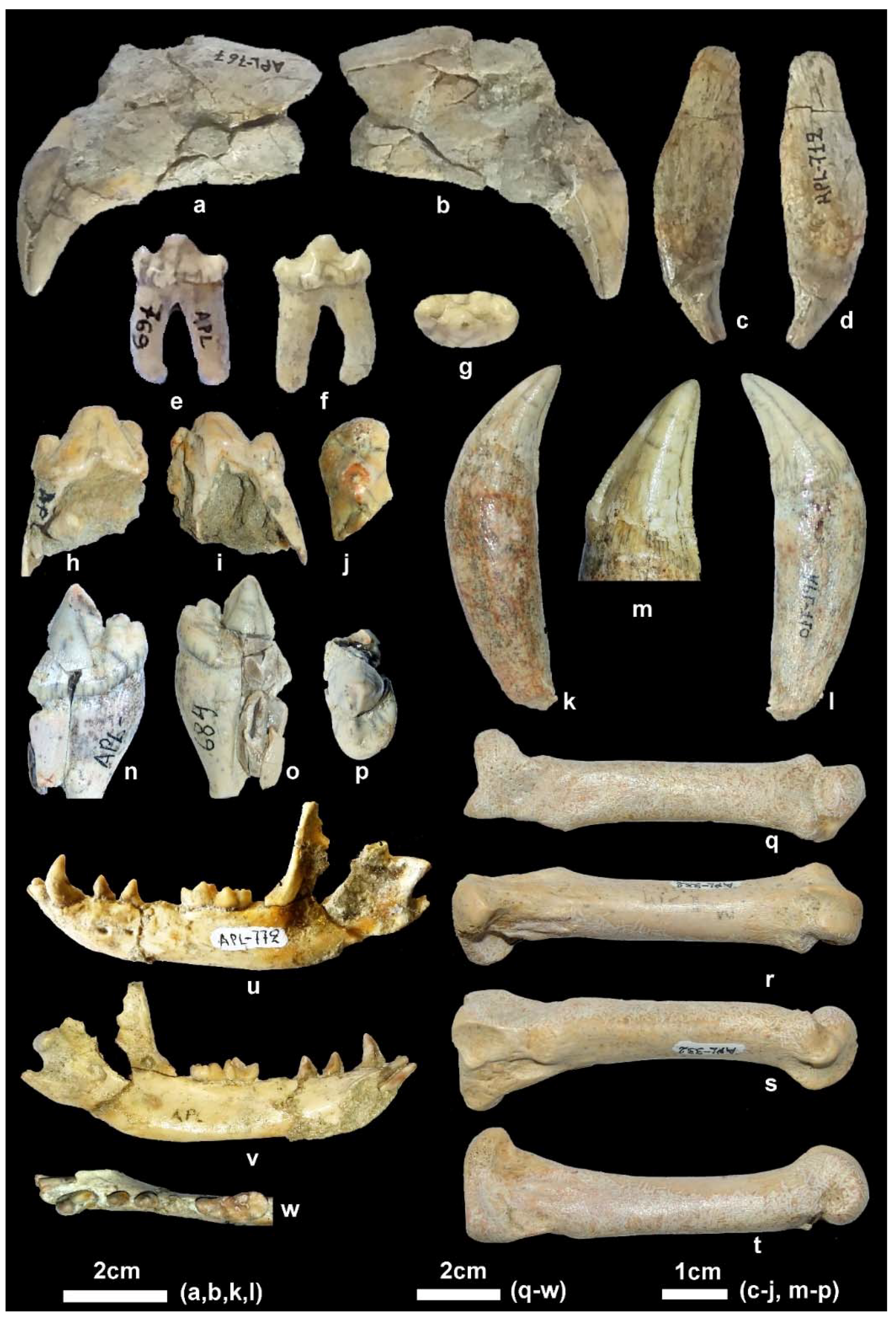
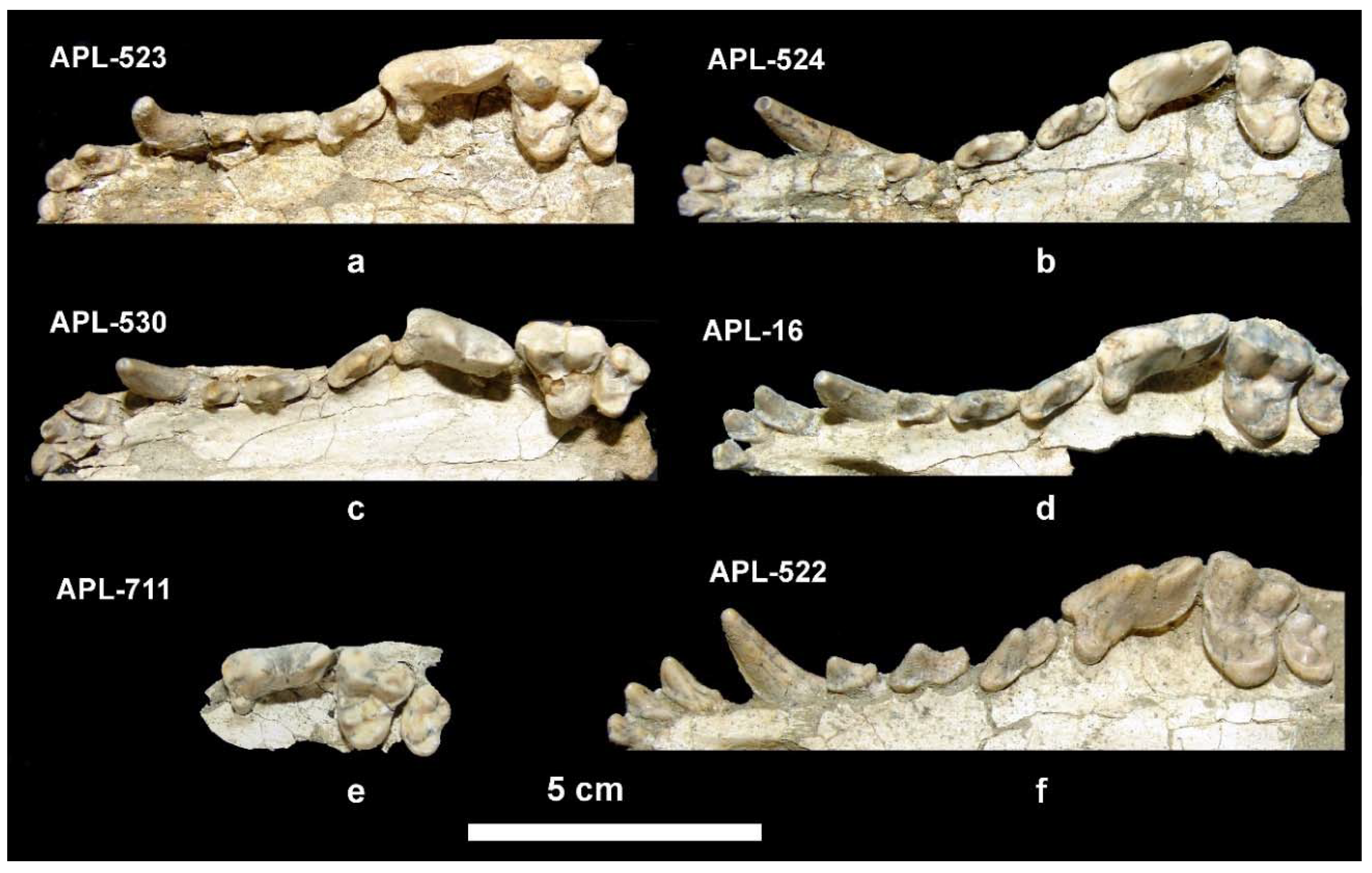
Description. The partial cranium APL-522 (
Figure 4f) has been already described [
4]. The available mandibular remains are badly preserved or fragmentary (
Figure 5i–k) providing limited morphological information. The mandibular corpus is higher than that of
C. apolloniensis and its ventral margin is slightly convex below the m1 and m2. It bears a large mental foramen below the mesial part of the p2 and a small one below the middle of the p3 (
Figure 5j). The masseteric fossa is partly preserved in APL-526 and its mesial margin (
Figure 5i
1) is situated below the m3. The dentition of APL-569 and APL-689 is worn but that of APL-526 is less worn. The canine is small relative to the size of the mandible and strongly curved distally; in the right one, a lingual crest starts from its apex and, after the middle of the tooth’s height, is curved distally and connected with the pronounced distal cingulum. The p1 is small, monocuspid and single-rooted as in the other APL canids. The p2 lacks a.a.c. but it bears a p.a.c. on a large distal cingular projection. The p3 lacks a.a.c. but bears a small cingular mesio-lingual prominence; the p.a.c. is larger than that of the p2 and there is an elevation of the distal cingulum. The p4 is like the p3 but it bears a small secondary p.a.c., which originates from the strong elevation of the distal cingulum. The carnassial is relatively short and robust with a small metaconid situated slightly distally to the protoconid; albeit the advanced wear of the talonid, the hypoconid and entoconid are well distinguished and a distal crest, well distinguished in APL-689, connects them. The m2 has an oval occlusal outline with well-developed metaconid and protoconid separated by a shallow furrow; the hypoconid is situated disto-bucally and is smaller than the other two cuspids from which it is separated by a deep valley; the shelf like mesio-buccal cingulum is pronounced.
Remarks. The cranium, APL-522, and a mandibular fragment with badly preserved p4 and m1 (APL-569) of
C. etruscus from Apollonia (
Figure 4e and
Figure 5i) have previously been described [
4]. The higher paracone than metacone in the M1, the reduced contact between the M1 and M2, the continuous cingulum in the M1 and M2, the relatively small paraconid and protoconid in the m1, the linked hypoconid and entoconid by a distal sinuous crest in the m1, and the relatively small paraconid and protoconid in comparison with
C. lupus of the APL sample agree with the diagnosis for
C. etruscus [
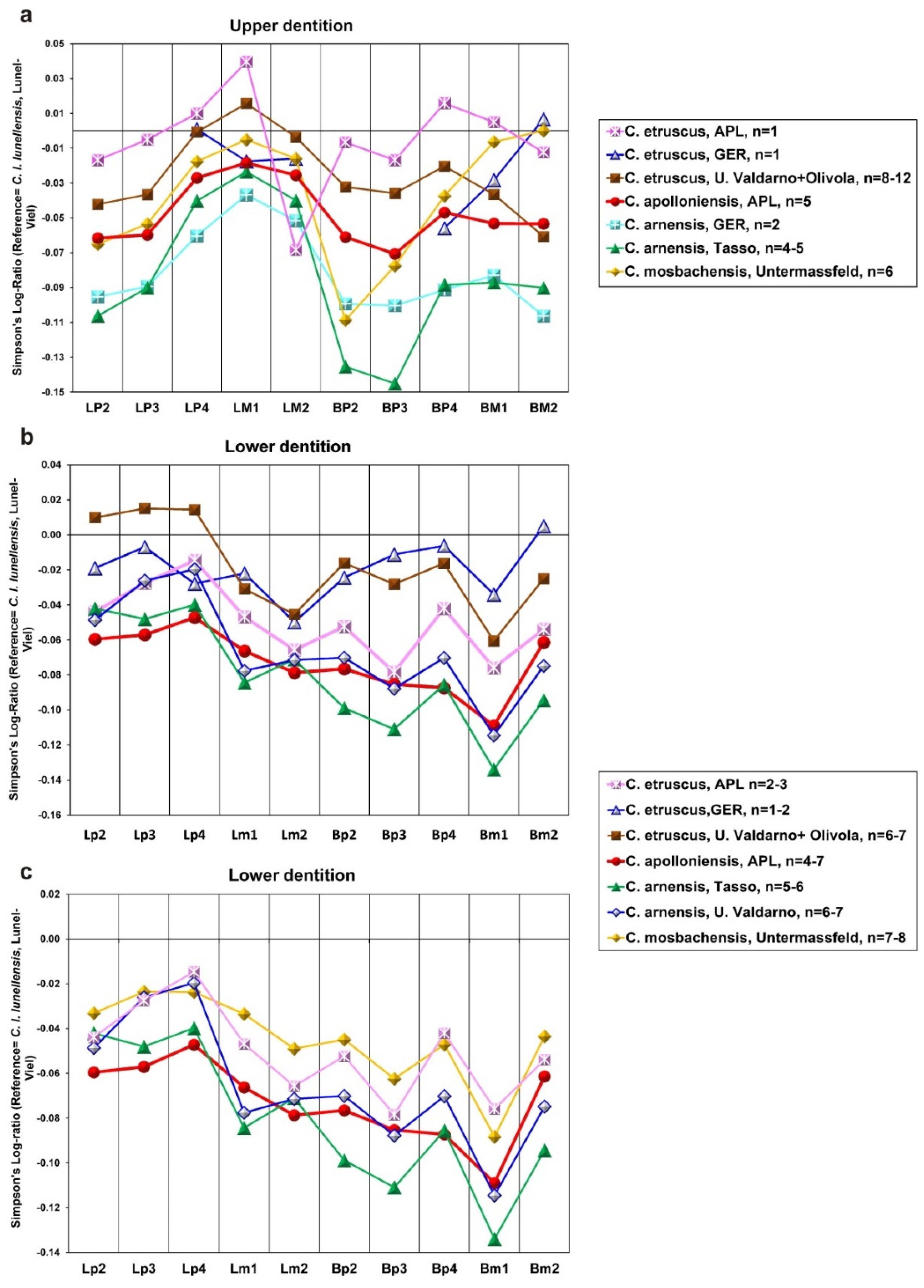
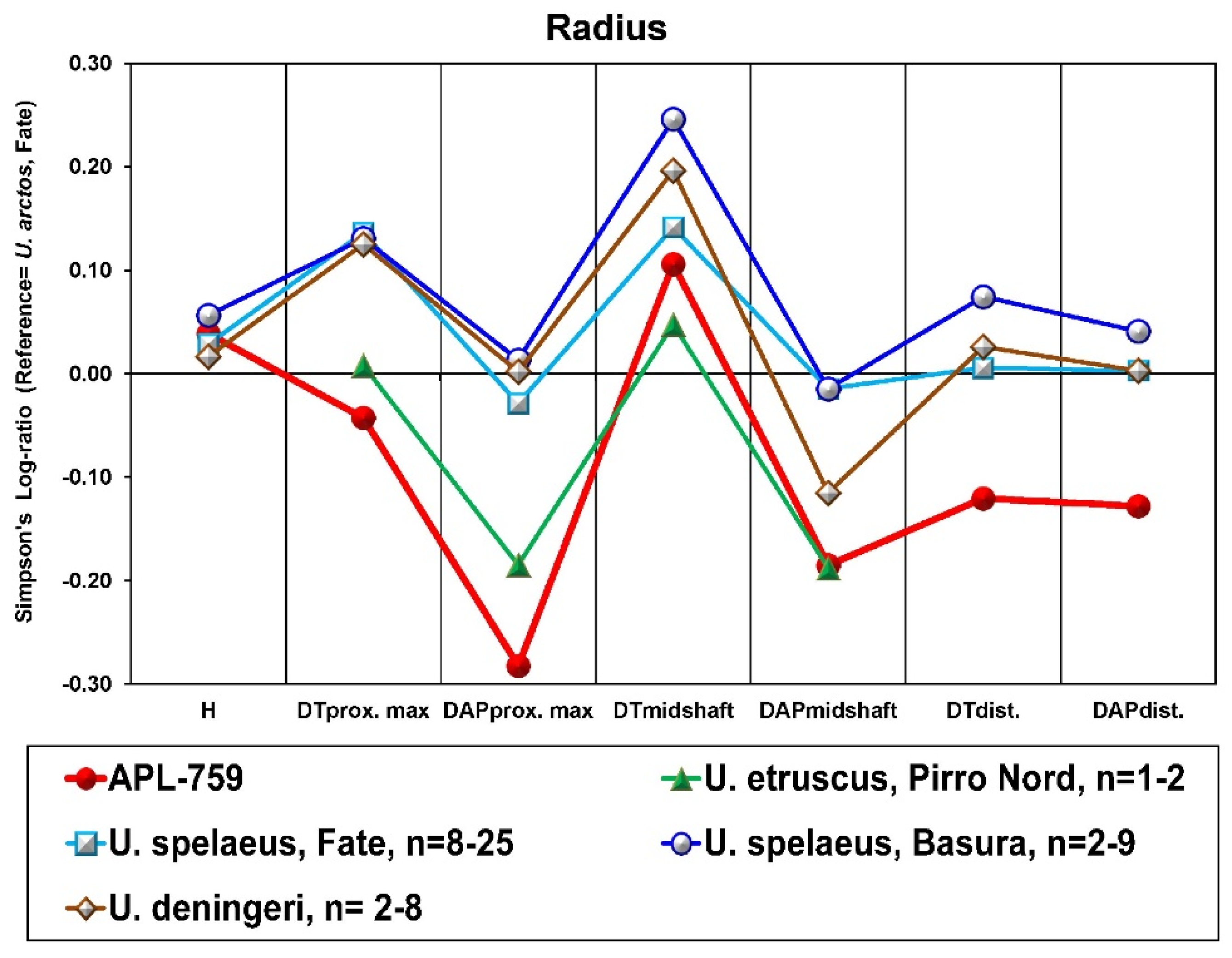
12]. The dental dimensions of the APL sample are also close to those of
C. etruscus (
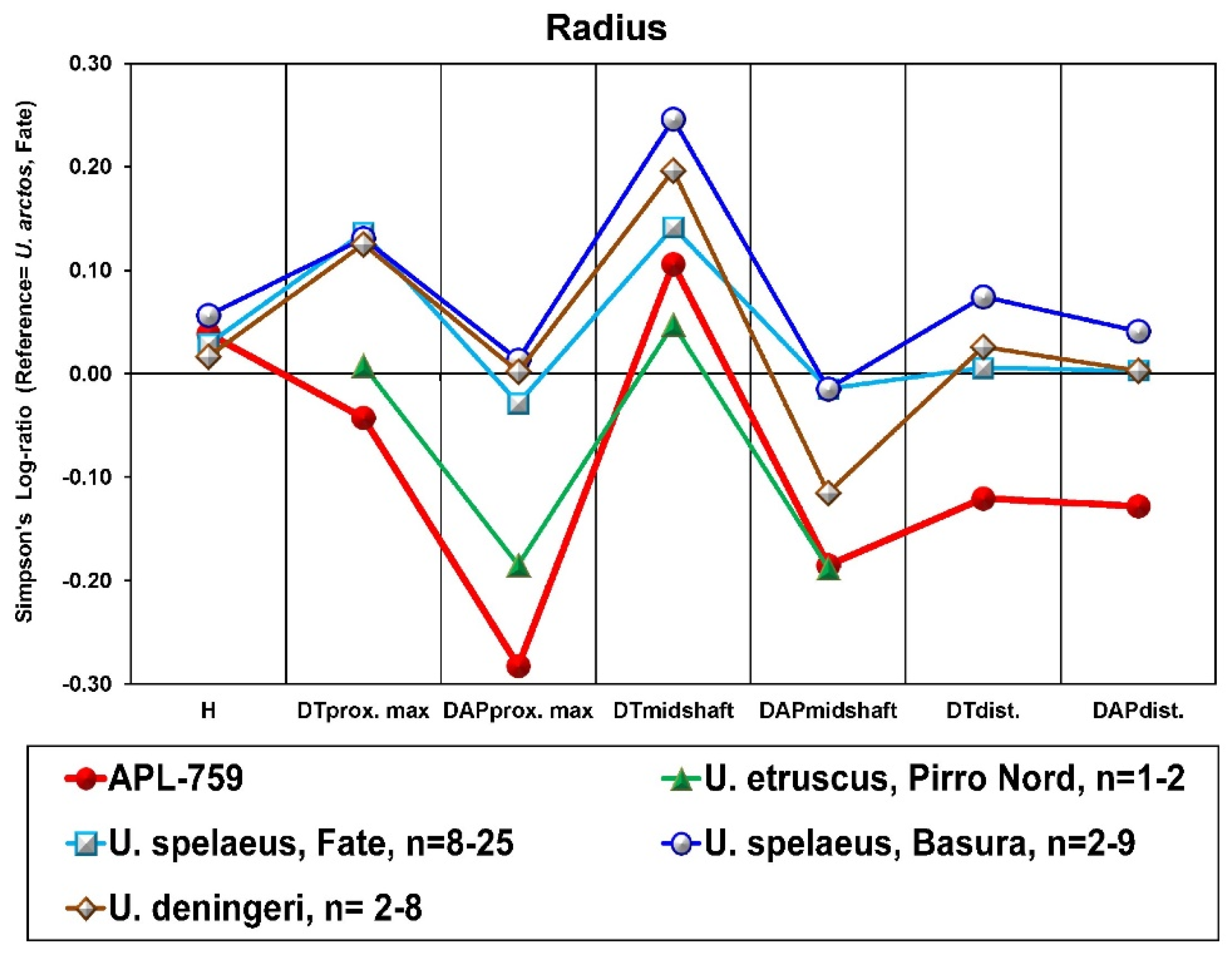
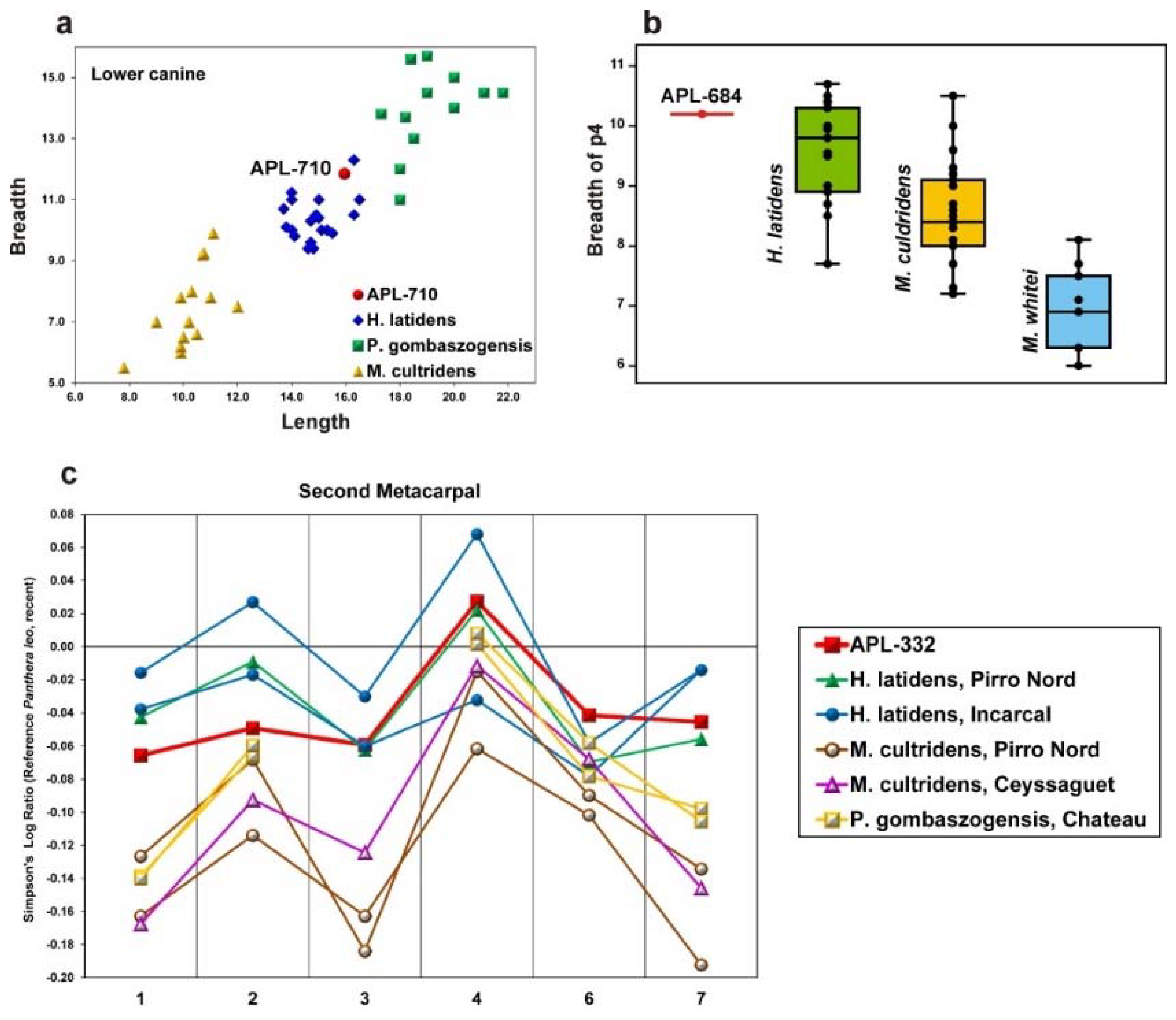
Figure 6). However, the APL upper teeth are larger than those from Upper Valdarno + Olivola (Italy) and Gerakarou (Mygdonia Basin, Greece) except for the M2, which is relatively short (
Figure 6a). This might be due to the very limited APL sample, which includes only one specimen. On the other hand, the APL lower teeth have similar proportions to
C. etruscus from Gerakarou and U. Valdarno+Olivola, but the lower premolars are narrower than those from the previous localities (
Figure 6b). The dental dimensions of the APL
C. etruscus are more or less similar to those of
C. mosbachensis but larger than those of
C. apolloniensis and
C. arnensis (
Figure 6c). The similar dental morphology and dimensions of the APL sample with
C. etruscus from different European localities suggest that it can be attributed to this taxon. Besides APL,
C. etruscus is known from two other localities of the Mygdonia Basin, i.e., the early Late Villafranchian locality Gerakarou and the Late Villafranchian locality Tsiotra Vryssi [
2,
7]. Some dental remains of
C. etruscus are also known from the Late Villafranchian locality Alykes (Thessaly, Greece) [
13].
3.2.2. Canis apolloniensis Koufos and Kostopoulos, 1997
Synonyms. 1992 Canis arnensis Koufos, p. 7–8 (partim). 1997 Canis arnensis Koufos and Kostopoulos.
Material. New collection: Left maxillary fragment with P4-M3, APL-711; right mandibular corpus with i3-m2, APL-715; left hemimandible with c, p2-m2, APL-690; right hemimandible with p3-m2, APL-703.
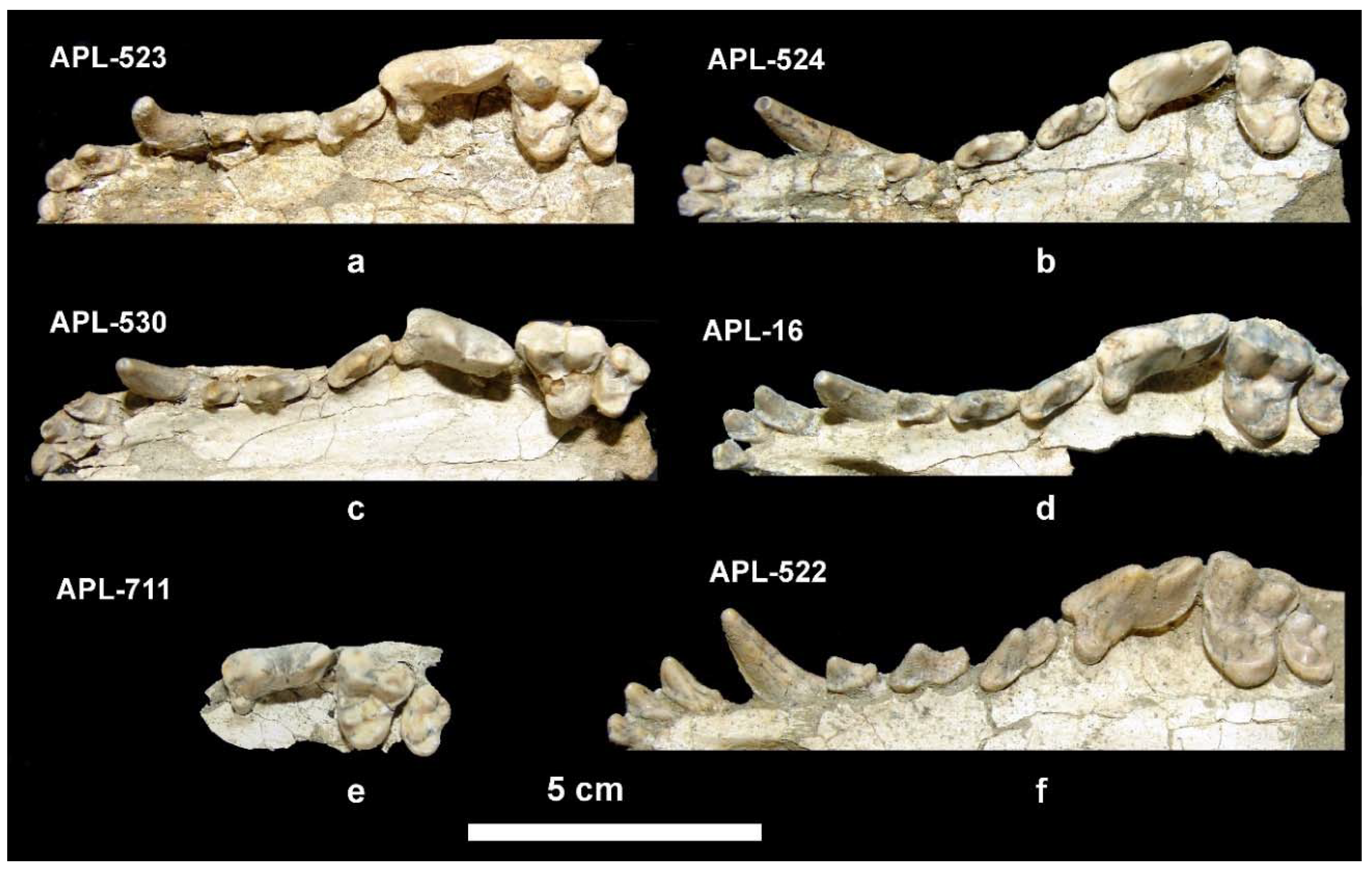
Old collection: Frontal cranial part and mandible, APL-530 (holotype); frontal cranial part, APL-523; maxilla with both I1-M2 tooth rows, APl-524; right maxilla with left I1-M2 and right I1-P2, APL-16; left maxillary fragment with i2-p3, APL-1; left mandibular fragment with c-m3, APL-17; right mandibular fragment with c-m1, APL-528; right mandibular fragment with p2-m2, APL-527.
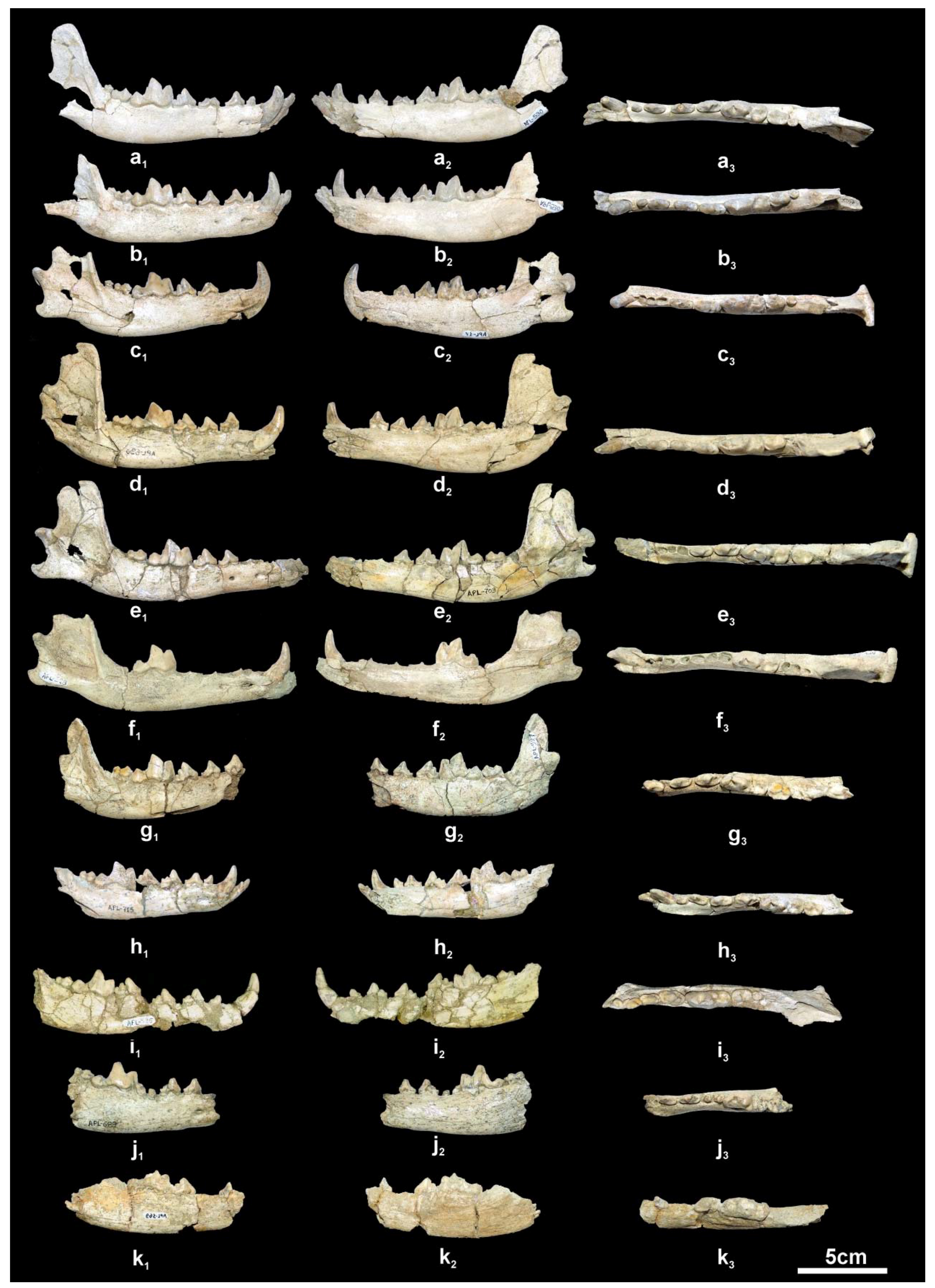
Description. The available cranial remains (APL-523, APL-530) preserve the frontal part of the cranium; they are compressed and the cranial bones are crushed. The nasal bones are better preserved in the holotype APL-530; they are narrow and elongated extending distally behind the anterior border of the orbit. The nasal cavity is small, oval-shaped and slightly inclined distally. The muzzle is relatively short. The well-preserved orbit in APL-523 is rounded with its anterior margin above the distal end of the carnassial. The palate is elongated and narrow. The tooth rows are well preserved in all specimens (
Figure 4).
The upper incisors increase in size from I1 to I3 (
Figure 4a–f); the last incisor is canine-like with strong lingual and distal cingula and bears a crest across its distal margin. The I1 and I2 have a triangular lingual outline and bear a distal cingular projection; two strong cingula start from this projection and are directed respectively towards the anteromesial and anterodistal extremities of the tooth forming two cusplets. The upper canine is strong, curved distally and strongly flattened bucco-lingually; it bears a weak mesio-lingual and distal crest, running from its apex to the base. The P1 is small, monocuspid, single-rooted with strong buccal and distal cingula; the P1 of APL-523 and APL-530 is slightly smaller than that of the other specimens. The P2 is long and narrow, bearing a p.a.c. situated in the middle of an elongated distal cingular projection; the p.a.c. is large in APL-523 and APL-524 but it is very small in APL-16 and APL-530; the distal margin of the distal cingualar projection is elevated and looks like a small secondary p.a.c.; there is a well-developed lingual cingulum. The P3 is morphologically like the P2, but is larger with clear and strong p.a.c. and with more expressed elevation of the distal cingulum; it bears a small mesio-lingual prominence of the cingulum from which starts a faint mesial crest to the apex of the main cusp. The carnassial is relatively short and wide; it lacks a parastyle and the protocone is small, low, and well separated from the paracone; its mesial margin is aligned with that of the paracone; there is a crest across the mesial border of the paracone from its apex to the base of the crown. The paracone is high and separated from the metacone by a deep valley; the metacone is blade-like and its distal part slightly curves buccally; a well-developed cingulum is observed in the disto-lingual half of the tooth. The M1 is triangular-shaped with slightly convex mesial margin and concave distal one; the paracone is larger and higher than the metacone; the protocone is small and low; the hypocone is crest-like; there is a small protoconule clearly distinguished in the less worn teeth; the entocone is also well distinguished in all specimens and is connected with the protocone by a crest; the main basin is not very deep and that between the protocone and hypocone is small and shallow; the mesial and disto-buccal cingula are well developed but the buccal cingulum is weak. The M2 has elliptical occlusal outline and similar morphology to the M1, but is smaller with faint or absent entocone and stronger buccal cingulum.
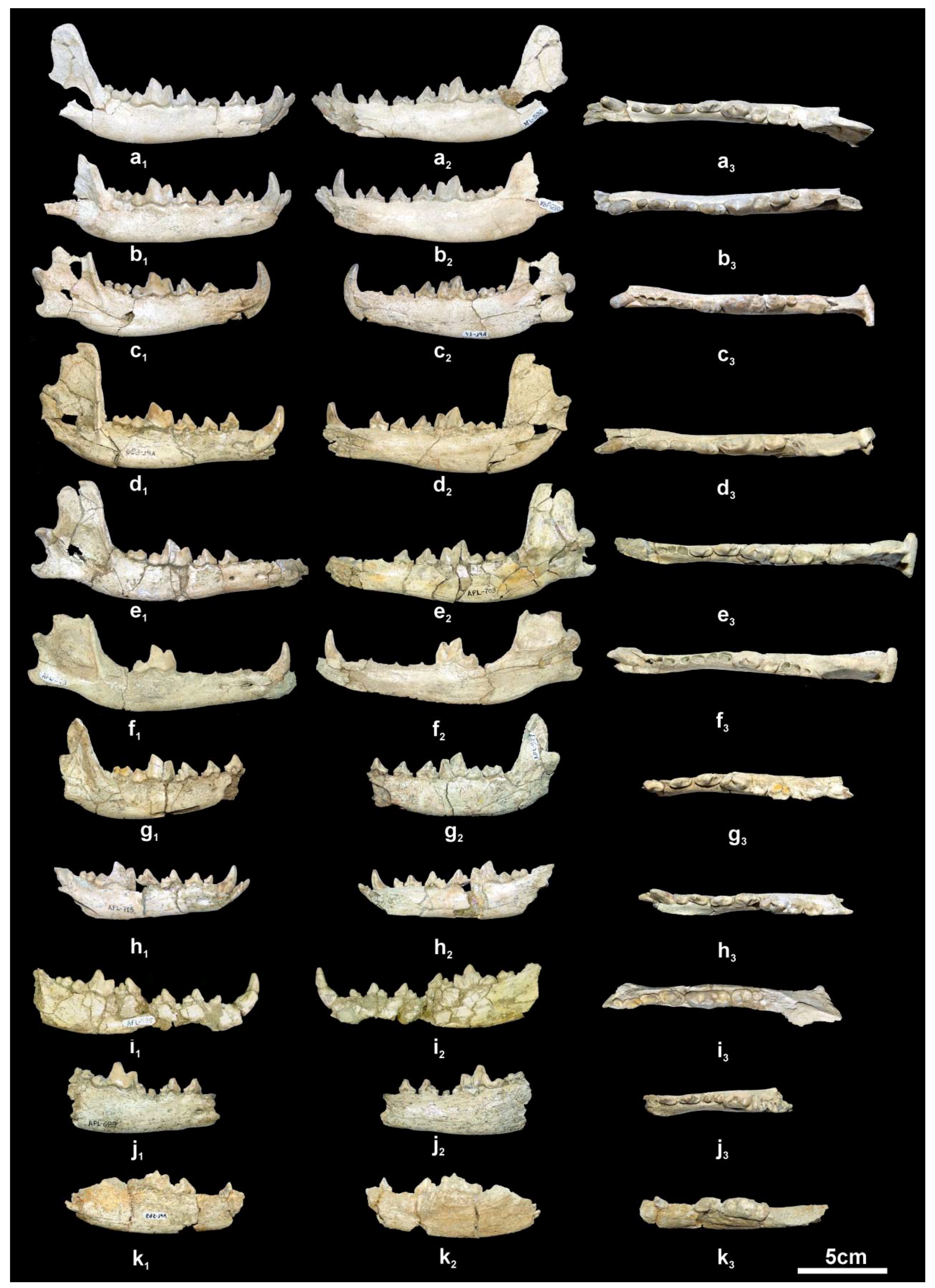
The mandibular corpus (
Figure 5a–h) is elongated and relatively shallow; the ventral margin of the mandibular corpus bears a faint concavity from below the p2 to the p4 and then it curves gradually till the angular process, where it is strongly concave; the angular process is short, sharp and does not exceed the posterior margin of the condyle. The coronoid process is relatively low and separated from the condyle by a shallow mandibular incision. There are two mental foramina, one below the middle of the p2 and another below the middle of the p3. The masseteric fossa is oval and deep with its anterior margin below the m3.
The lower incisors are well preserved in APL-530 (
Figure 5a); they have a triangular shape in lingual view with strong distal projection, like a small cuspid, which is quite large in the i3; the latter is canine-like and stronger than the other two incisors. The lower canine is relatively small, situated bucco-lingually, strongly curved distally and flattened bucco-lingually; it bears a faint mesio-lingual and distal crest. The p1 is small, monocuspid and single-rooted with a weak distal cingular projection. The p2 is elongated and narrow with a strong distal cingular projection, the distal margin of which is elevated and looks like a p.a.c.; a clear crest runs across the mesial and distal margins of the main cuspid. The p3 has similar morphology to the p2 but is larger and has stronger mesial cingulum and distal cingular projection; it lacks a p.a.c. but there is a small one in APL-715 and a vestigial one in APL-527 and APL-703; it bears a weak lingual cingulum. The p4 is more robust than the other premolars, with strong mesial cingulum; the distal cingular projection is large and bears a clear and large p.a.c. situated buccally; in APL-527, APL-530, APL-690 and APL-715 there is a small to faint secondary p.a.c.; the lingual cingulum is well developed. The lower carnassial is relatively short and wide, with a talonid about 1/3 of the tooth’s length. The protoconid is high and separated from the paraconid by a deep valley; the metaconid is small and situated disto-lingually to the protoconid; the talonid bears a large hypoconid and a small entoconid connected distally by a crest. The m2 has an oval occlusal outline with a large protoconid and a relatively small metaconid and hypoconid; it lacks an entoconid, but in some specimens (APL-530, APL-715) there is a crest in its position connected with the hypoconid; the mesial cingulum is strong. The m3 is present in APL-530; it is small, rounded, and single-rooted, with two cuspids; the large protoconid dominates and the paraconid is very small, situated lingually; a strongly developed cingulum is visible around the entire tooth.
Remarks.
Canis apolloniensis was originally described from APL based on some maxillary and mandibular remains together with
C. etruscus. Some authors consider
C. apolloniensis as belonging to the
C. etruscus-C. mosbachensis line, or as a southern European form of
C. mosbachensis [
17,
18,
19,
20]. However, other researchers synonymized it with
C. arnensis [
14,
21]. Although an extensive definition of
C. apolloniensis is given in the original description of the taxon [
4], the new findings and data published during the last twenty years, allow the knowledge of the taxon.
The inclusion of
C. apolloniensis in the evolutionary line of
C. etruscus–C. mosbachensis is based on the following distinctive characteristics separating
C. etruscus and
C. arnensis [
17].
“In
C. arnensis the talonid of […] M
1 […] presents two clear different and separated peaks [hypoconid and entoconid] and in
C. etruscus both cusplets are linked by a sinuous crest”; the last character is also mentioned in the emended diagnosis of
C. etruscus [
12].
The m1 talonid of
C. apolloniensis consists of two separated cuspids and bears a small sinuous cristid in its distal part, less pronounced than that of
C. etruscus; in APL-690 and APL-703 it is very small, like a vestigial cusplet. In
C. mosbachensis the m1 talonid is bicuspid but the two cuspids are connected through a strong transverse crest [
15].
The m2 in C. etruscus “... shows a very well pronounced cingular antero-external border but it is not present in C. arnensis”.
The pronounced mesial cingulum is clear in the m2 of
C. etruscus from APL and Gerakarou (Mygdonia Basin, Greece) [
2]; it looks like a shelf in the mesial and mesio-buccal margin of the tooth.
Canis apolloniensis bears a less pronounced mesial cingulum than that of
C. etruscus; in some specimens it is faint especially in the lingual half of the tooth. On the contrary, the mesial and mesio-buccal cingula of
C. mosbachensis are well developed, like in
C. etruscus.
“In […] M1 of C. arnensis the protocone is linked to the parastyle and metastyle by a well-defined crest, and the paraconule and metaconule are well marked”.
In
C. apolloniensis we cannot say that there is a crest linking the protocone with the parastyle and metastyle. Additionally, the paraconule of
C. apolloniensis is weak and just distinguished, while the metaconule is well defined but relatively smaller than in
C. arnensis. In
C. mosbachensis both paraconule and metaconule of the m1 are prominent and the buccal cingulum complete and strong [
15], distinguishing it from
C. apolloniensis.
“In […] [M2] of C. arnensis there are 4 cusplets very well isolated [...] C. etruscus has only three peaks, never presents entoconid...”.
Nevertheless, the presence of accessory cuspids in the disto-lingual portion and sometimes the presence of an entoconid are mentioned for the m2 of
C. arnensis [
20]. In all available specimens of
C. apolloniensis the m2 is tricuspid and in this feature, it is closer to
C. etruscus.
“C. arnensis has diastemes between all the premolars and C. etruscus has no separation between the premolars”.
This feature was also mentioned earlier [
22,
23], while a small diastema between the premolars is mentioned in the original description of
C. apolloniensis [
4]. A more detailed study of the available material indicates that this character varies in the APL material. In APL-524, there are diastemata between the premolars but the specimen is crushed and probably this affected the position of the teeth. In APL-523 there are diastemata, except between P3 and P4 but the specimen is deformed. In APL-530 there is a small diastema between P1 and P2 and a larger one between P2 and P3. In
C. mosbachensis there are specimens with diastemata between the premolars (IQW 1982/18052), but the premolars set closely in others (IQW 1984/20177) [
15]. Based on the great variation observed in this character, it seems that it cannot be used as diagnostic and rightly was not included in the emended diagnosis of
C. arnensis [
20].
Based on the abovementioned characteristics,
C. apolloniensis has more similarities with
C. etruscus than with
C. arnensis and for this reason it is considered closer to the
C. etruscus-
C. mosbachensis evolutionary line [
17].
Canis etruscus from the Villafranchian of Italy was recently revised [
12]; based on the proposed diagnosis,
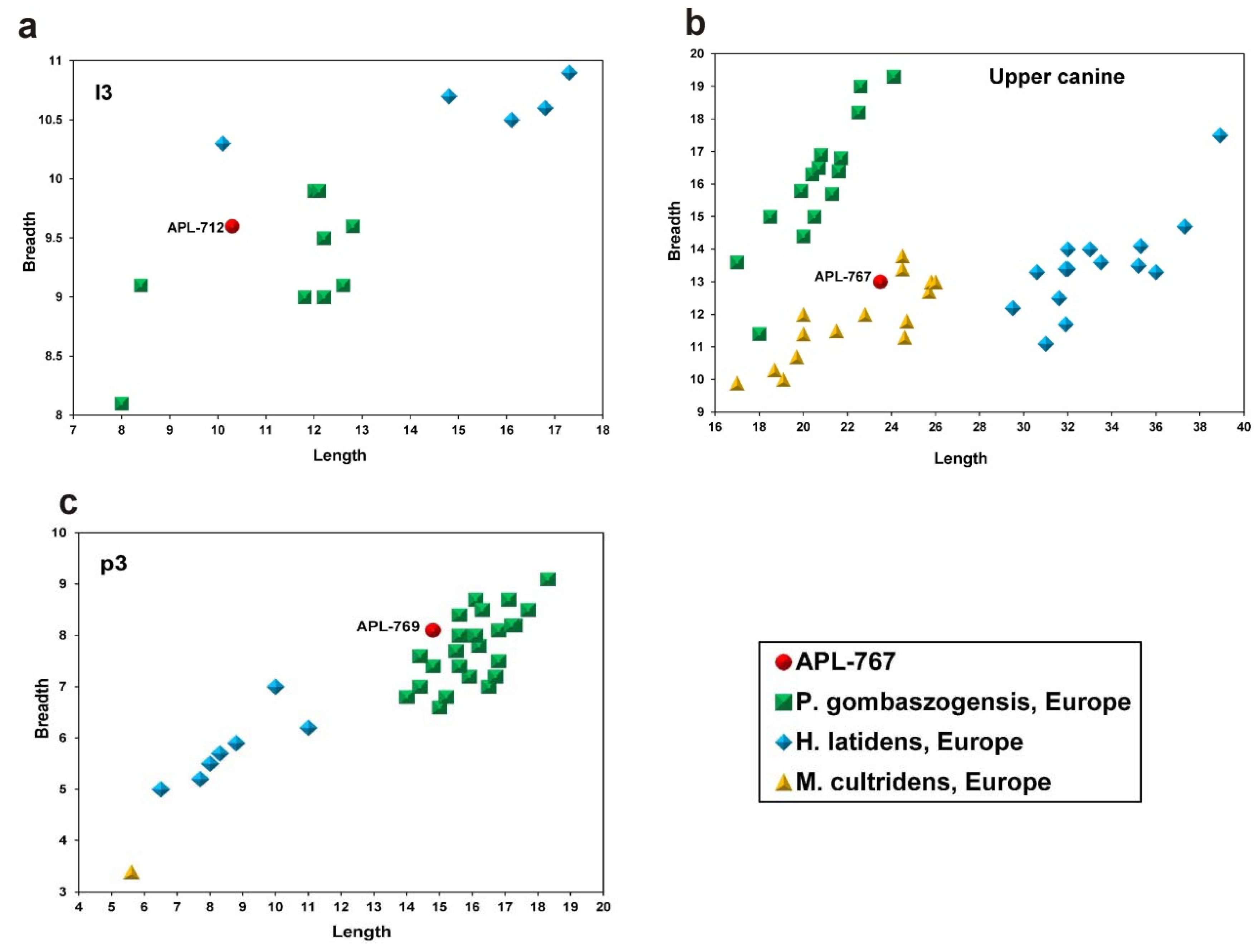
C. apolloniensis differs from this taxon displaying absence of lingual cingulum in the P1, a vestigial secondary p.a.c. (modified distal cingulum) in the P3 (it is strong in
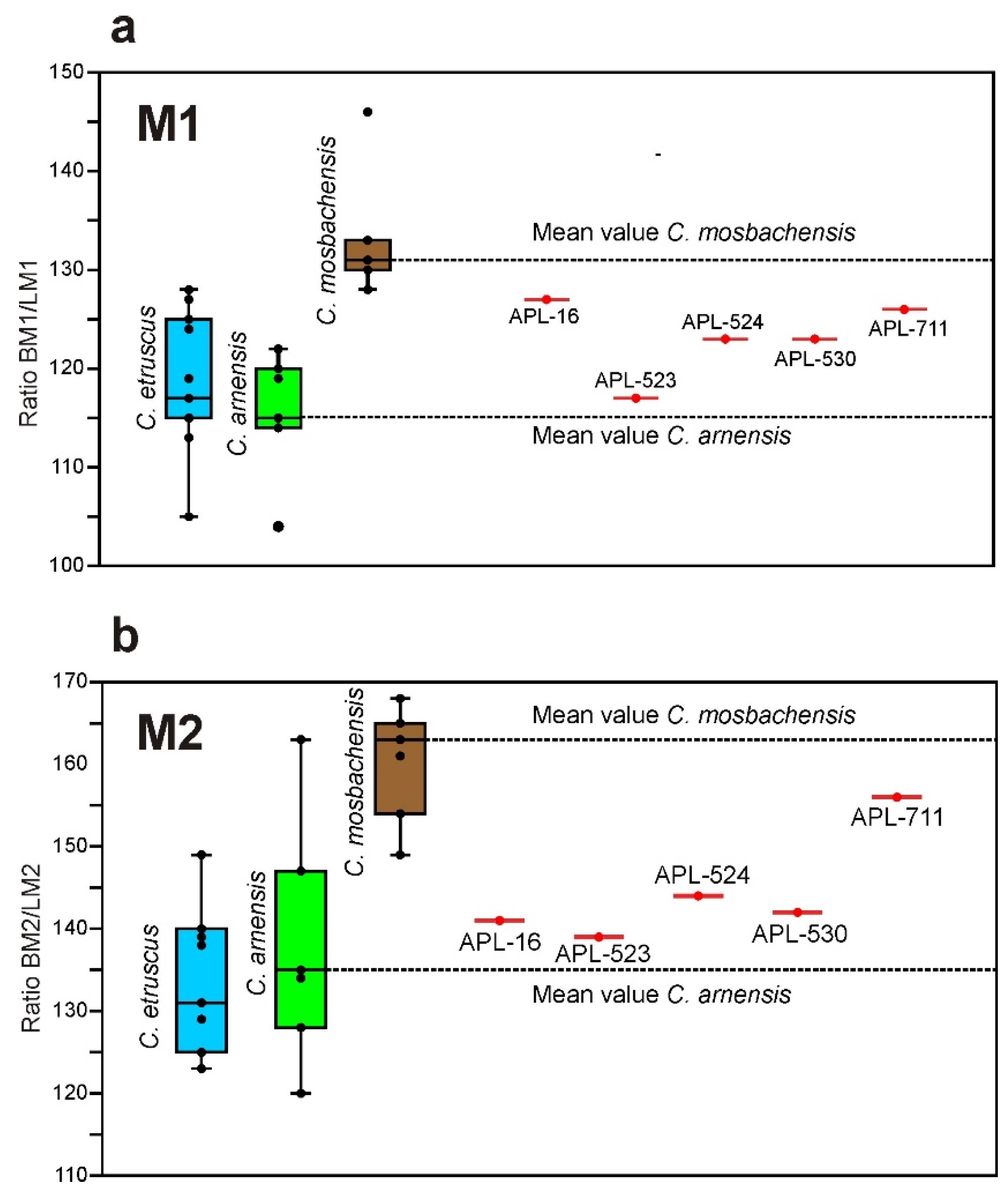
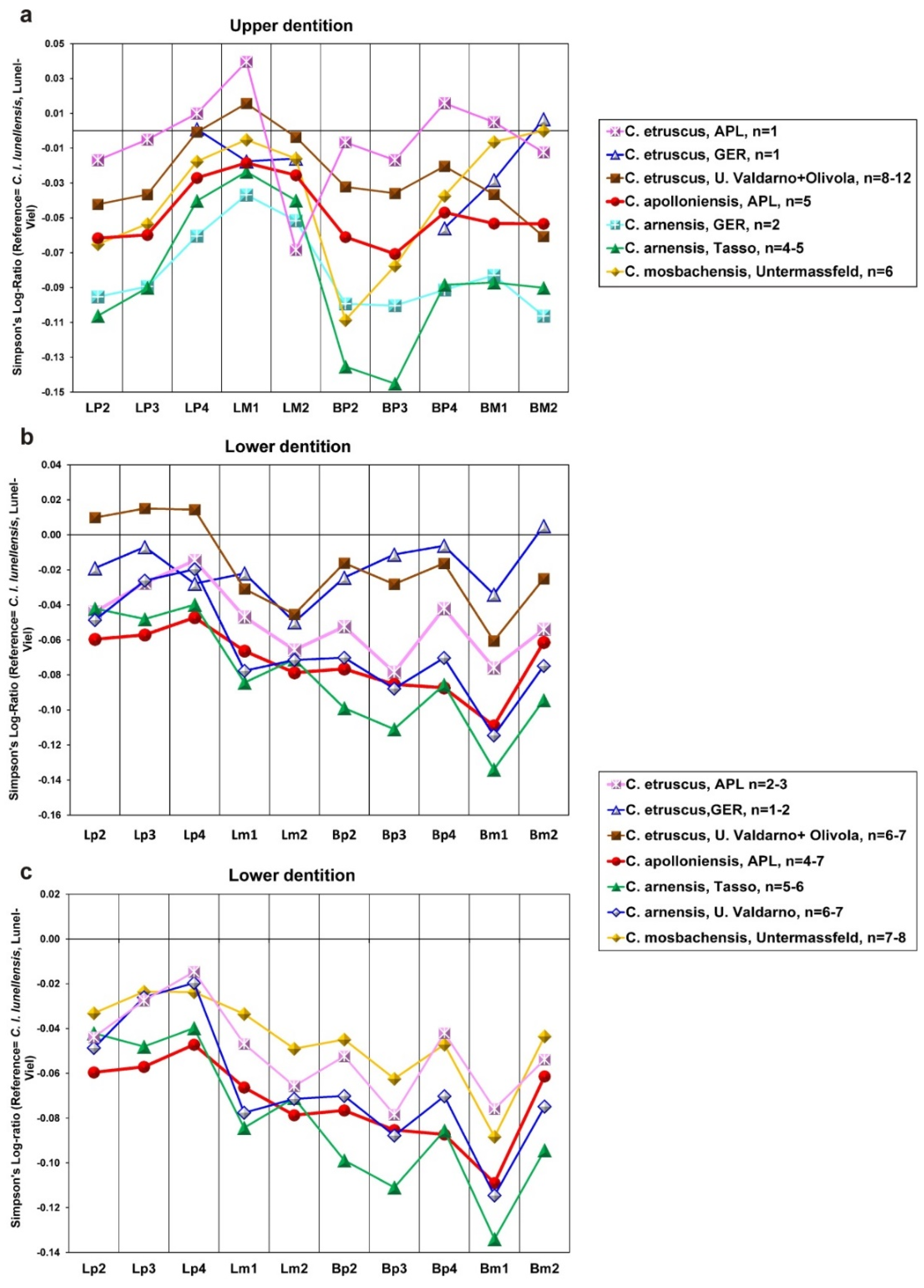
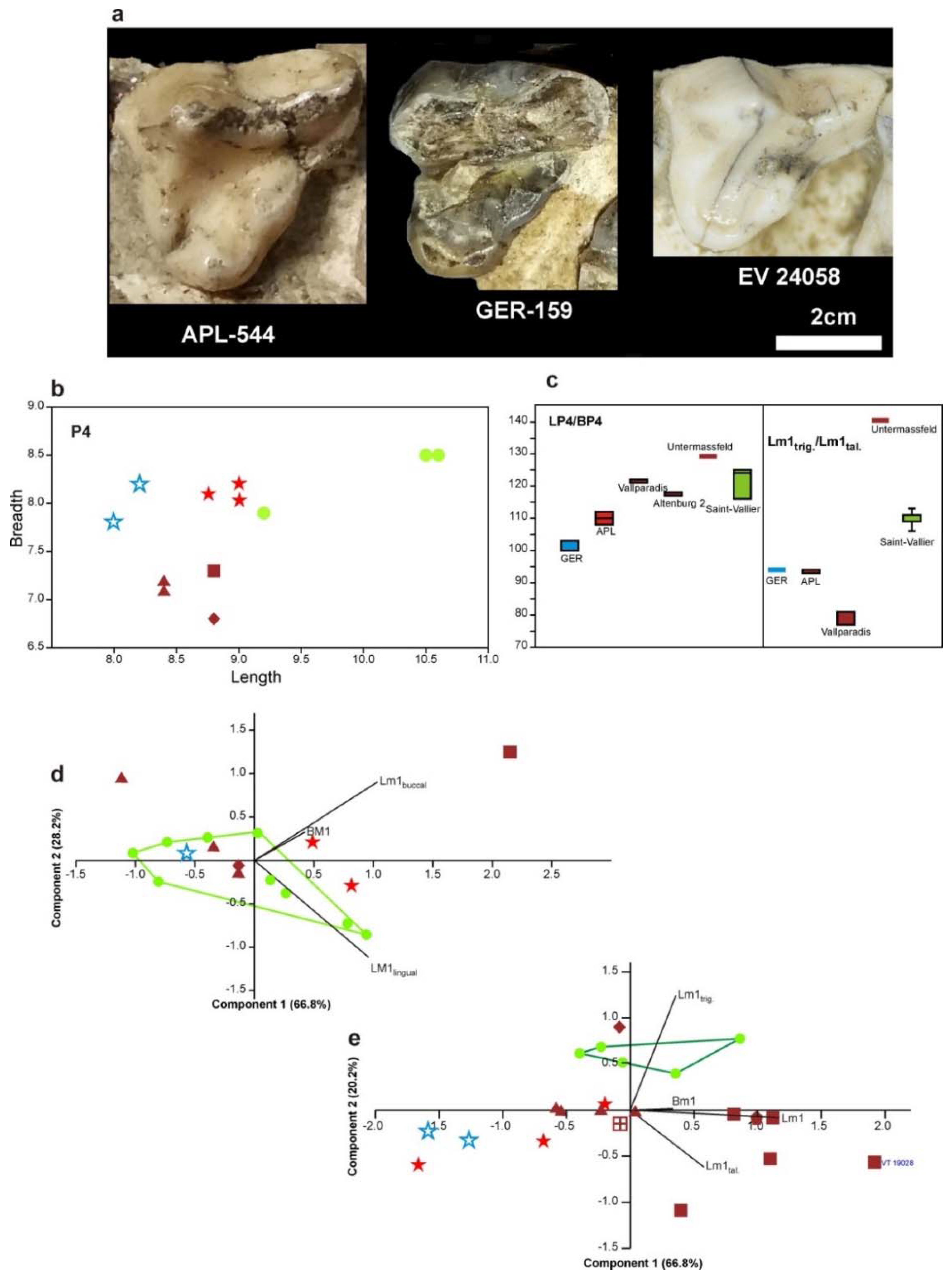
C. etruscus), a weak paraconule in the M1, more constricted medially M1, on average more slender M1 and M2 (
Figure 7), absence of a secondary p.a.c. in the p3 (it is strong in
C. etruscus, weak mesial cingulum in the m2, and smaller size (
Figure 6a–c). Based on the abovementioned comparisons, the APL canid is separated from
C. etruscus.
The revision of the Italian sample of
C. arnensis and the description of some new material from Poggio Rosso (Italy) provided an emended diagnosis for the taxon [
20]. On that ground,
C. apolloniensis differs from
C. arnensis in having a small p.a.c. in the P2 (it is absent in
C. arnensis), larger paracone than metacone in the M1 (they are equal-sized in
C. arnensis), deeper protocone than hypocone basin in the M1 (they have the same depth in
C. arnensis), remarkably larger hypoconid than entoconid in the m1, absence of p.a.c. in the p3 (there is a variably developed one in
C. arnensis, APL-715 bears a small p.a.c. and APL-527 a vestigial one), usually absence of accessory cuspids between the entoconid and metaconid of the m1 (only two specimens present a vestigial cuspid), absence of a well-developed hypoconulid shelf on the distal margin of the m1 (there is a small and variably developed hypoconulid in the m1 of
C. arnensis), absence or very weak disto-buccal cingulum in the m1, and larger protoconid than metaconid in the m2 (equal-sized in
C. arnensis). In addition to these differences, the comparison with the known Greek material suggests that the two species have more or less the same size, though the upper teeth of
C. apolloniensis are larger (
Figure 6). Moreover, the M1 and M2 of
C. apolloniensis are on average more robust than those of
C. arnensis (
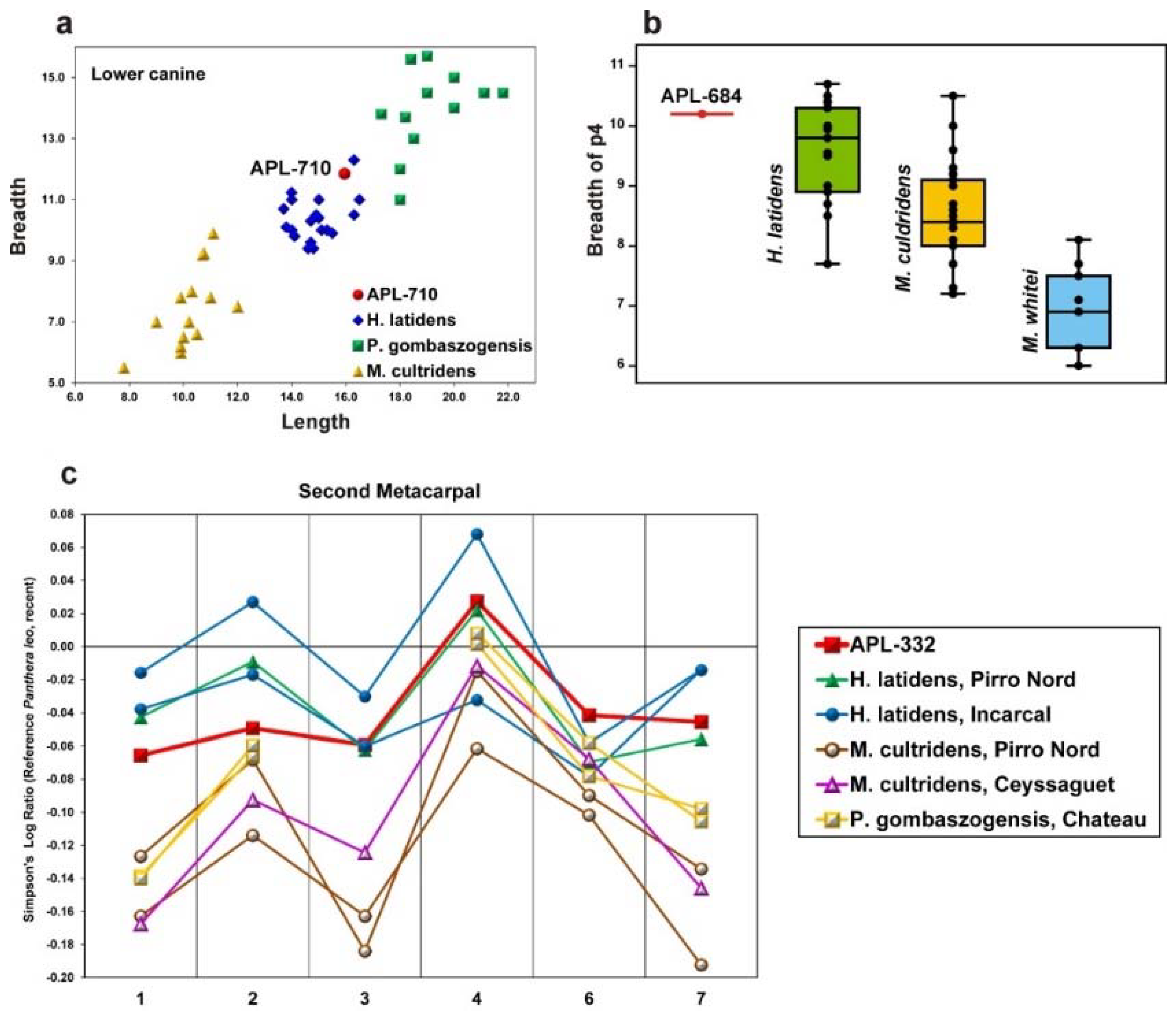
Figure 7) and the trigonid of the m1 is longer in relation to the talonid (the % ratio L
m1trig./L
m1tal. is on average 268 for
C. apolloniensis versus 245 for the Italian
C. arnensis). Based on the above comparative data,
C. apolloniensis, though it has a similar size to
C. arnensis, can be distinguished from it through several morphological and metrical characteristics.
The other opinion concerning the taxonomy of
C. apolloniensis is that it represents a southern European form of
C. mosbachensis [
20]. The latter species was originally described from the Middle Pleistocene locality Mosbach (Germany) [
24], while a rich sample was described from the locality Untermassfeld (Germany) [
15]. Based on the description and illustrations of
C. mosbachensis,
C. apolloniensis shows clear differences in displaying a laterally situated P3 in relation to the P4 (their main axes coincide in
C. mosbachensis), a vestigial secondary p.a.c. in the P3 (this is an elevation of the distal cingulum which is more pronounced in
C. mosbachensis), a small protocone aligned to the paracone, absence of p.a.c. in the p2 (sometimes it exists in
C. mosbachensis), a very small secondary p.a.c. in the p4 (it is markedly larger in
C. mosbachensis), and a small talonid relatively to the length of the m1 (the % ratio L
m1trig./L
m1tal. is 268 in
C. apolloniensis versus 224 in
C. mosbachensis). In addition to these differences our comparisons based on the description and illustrations of
C. mosbachensis from Untermassfeld [
15] indicate that:
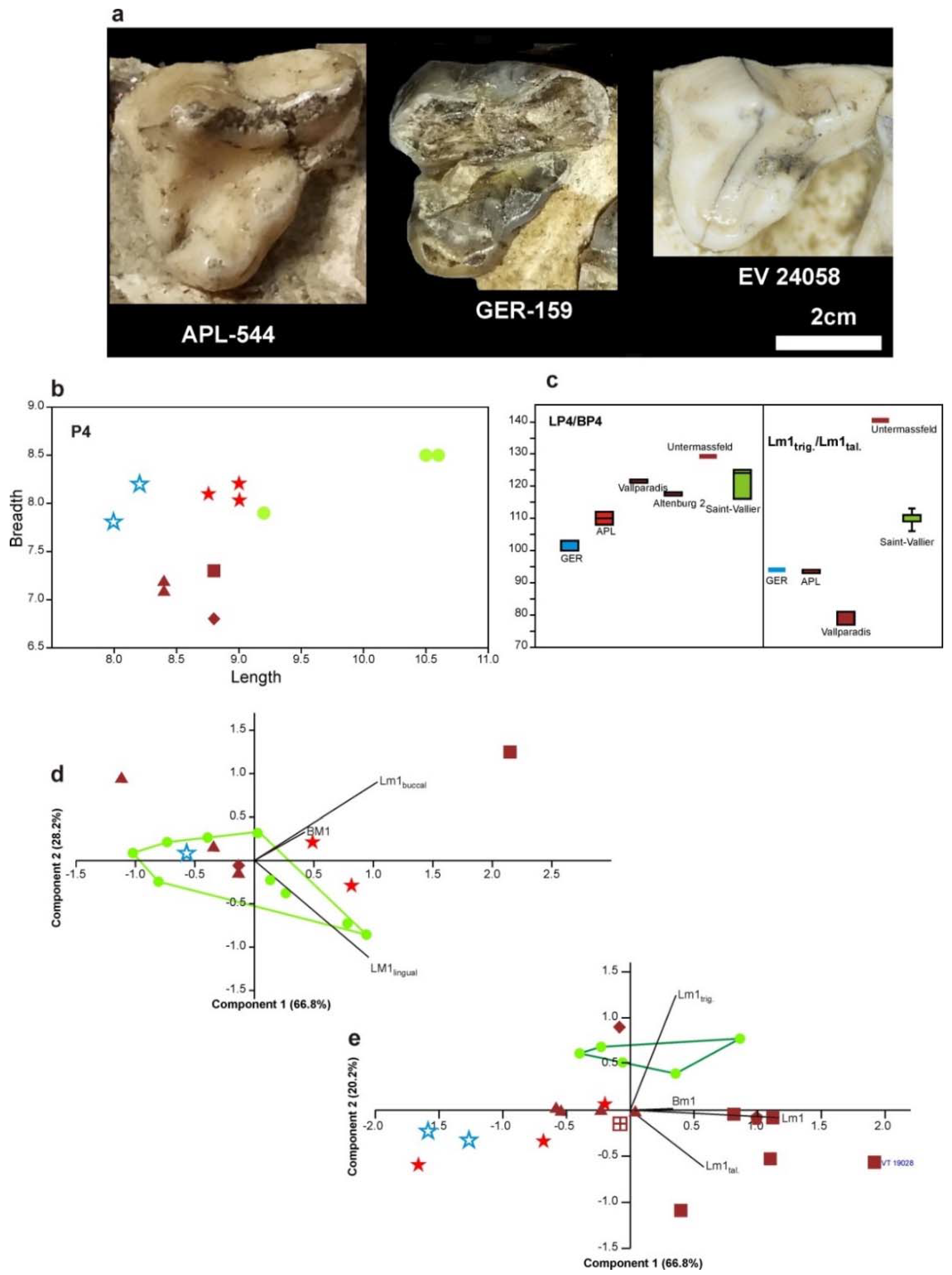
The occlusal shape of the M1 differs in the two species: it is sub-triangular and wide in
C. mosbachensis, while it is triangular with narrow lingual part in
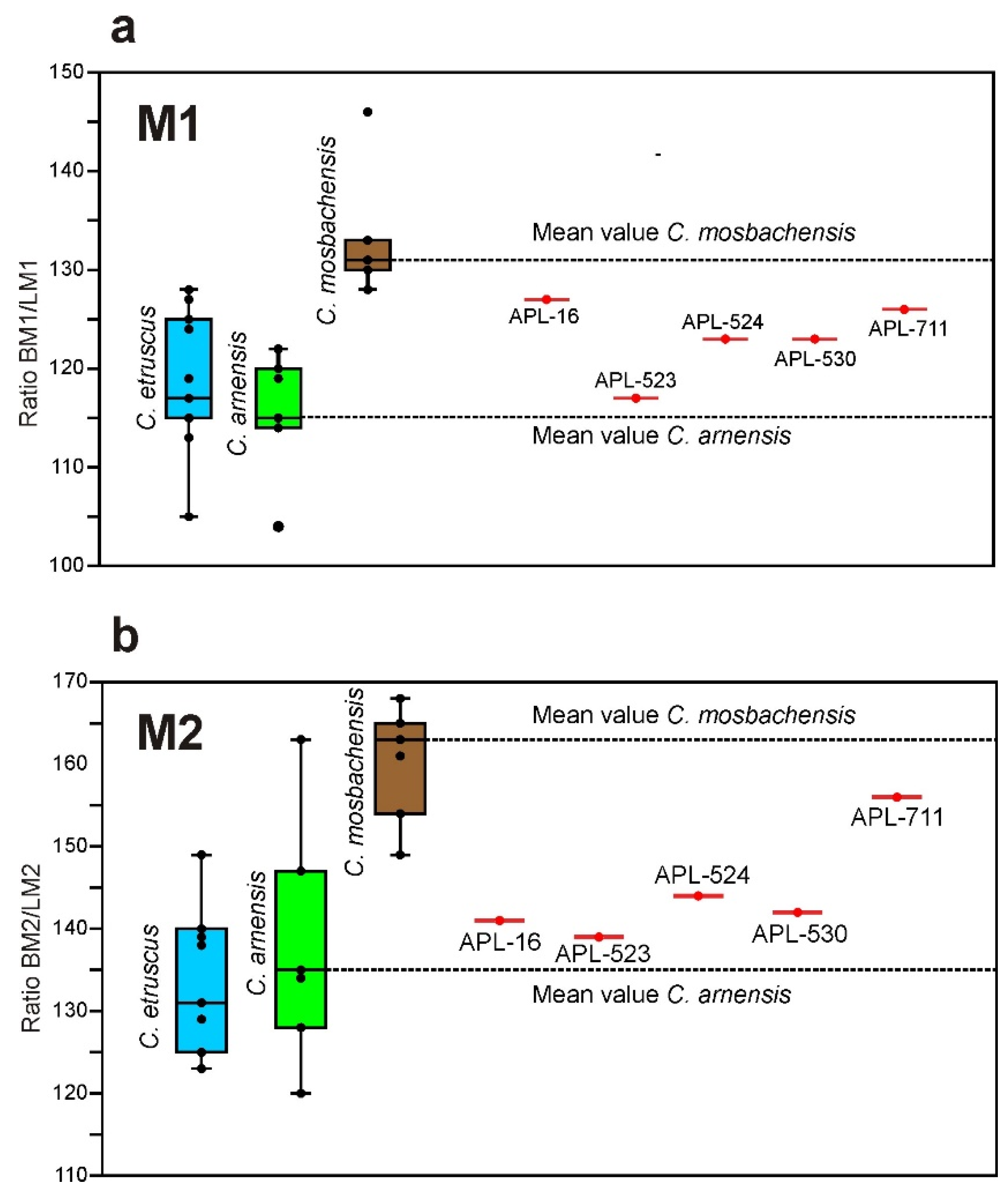
C. apolloniensis. The mean % ratio BM1/LM1 of
C. mosbachensis is remarkably larger than that of
C. apolloniensis; the mean value of the ratio for
C. apolloniensis is between the mean values for
C. arnensis (slender M1) and
C. mosbachensis (robust M1) (
Figure 7a).
The M2 of
C. mosbachensis is more robust than that of
C. apolloniensis; the mean % ratio BM2/LM2 of
C. apolloniensis is between those of
C. arnensis (slender M2) and
C. mosbachensis (robust M2), (
Figure 7b).
All the abovementioned comparative data suggest that C. apolloniensis can be separated from the known taxa C. etruscus, C. arnensis and C. mosbachensis; its morphology seems to be closer to C. etruscus and C. mosbachensis and thus could be a potential ancestor of the latter taxon.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}