
Monitoring Plateau Pika and Revealing the Associated Influencing Mechanisms in the Alpine Grasslands Using Unmanned Aerial Vehicles


Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Data Acquisition and Processing
2.2.1. Field Investigation and Pika Density Extracting
2.2.2. Pika Hazard Categories Base on EPH
2.2.3. Environmental Variables
2.3. Data Analyses
2.3.1. Relationships Between Pika Density and Environmental Variables
2.3.2. The Key Controlling Factors of Pika Density
2.3.3. Statistical Analyses
3. Results
3.1. Spatiotemporal Variation in Environmental Variables
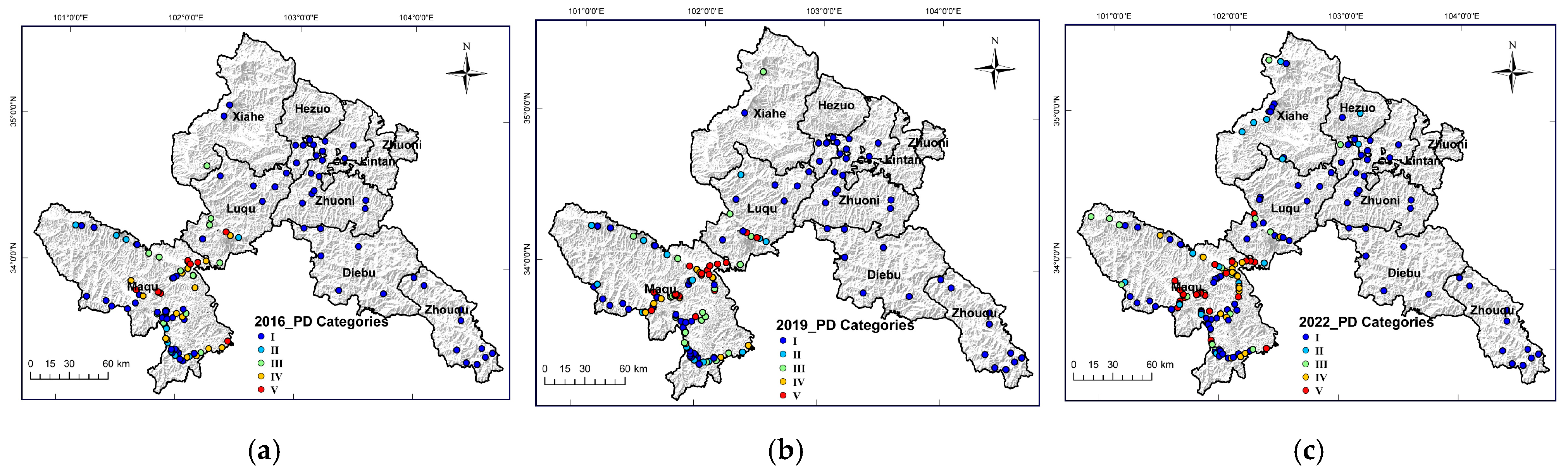
3.2. Spatiotemporal Distribution of Pika Density Spatiotemporal Differentiation Characteristics of the Distribution of Pika Density
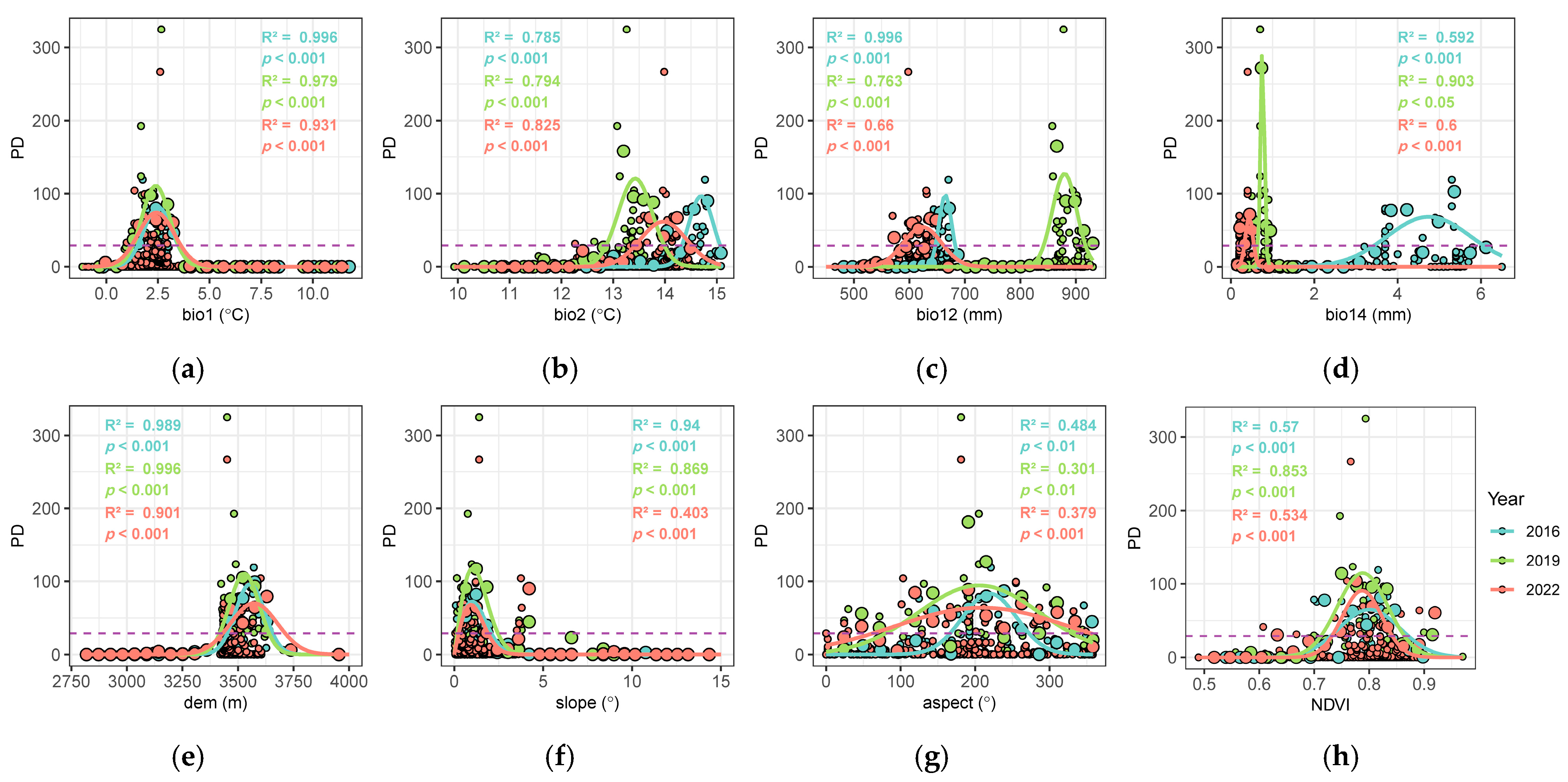
3.3. Response of Pika Density to Environmental Variables
4. Discussion
4.1. Spatiotemporal Distribution of Pika Density
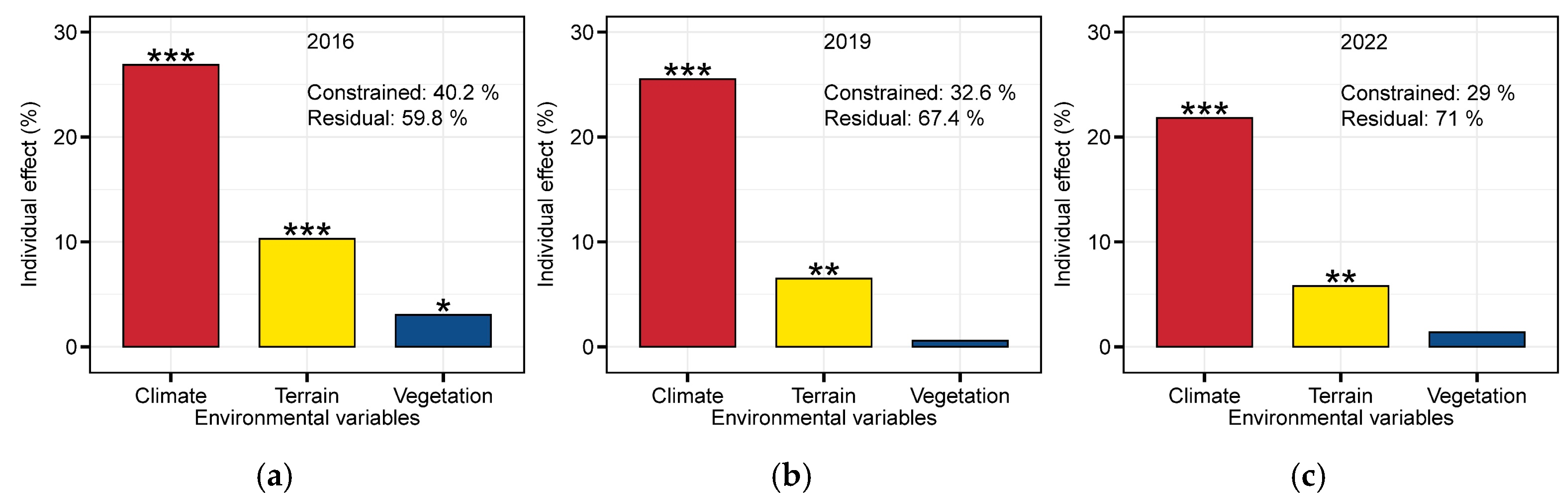
4.2. Key Factors Influencing Pika Distribution
4.3. Study Limitations and Future Research
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Correction Statement
References
- Yao, T.; Bolch, T.; Chen, D.; Gao, J.; Immerzeel, W.; Piao, S.; Su, F.; Thompson, L.; Wada, Y.; Wang, L. The imbalance of the Asian water tower. Nat. Rev. Earth Environ. 2022, 3, 618–632. [Google Scholar] [CrossRef]
- Miehe, G.; Schleuss, P.-M.; Seeber, E.; Babel, W.; Biermann, T.; Braendle, M.; Chen, F.; Coners, H.; Foken, T.; Gerken, T. The Kobresia pygmaea ecosystem of the Tibetan highlands–Origin, functioning and degradation of the world’s largest pastoral alpine ecosystem: Kobresia pastures of Tibet. Sci. Total Environ. 2019, 648, 754–771. [Google Scholar] [CrossRef] [PubMed]
- Pan, Q.; Sun, J.; Yang, Y.; Liu, W.; Li, A.; Peng, Y.; Xiu, J.; Xia, H.; Huang, J. Issues and Solutions on Grassland Restoration and Conservation in China. Bull. Chin. Acad. Sci. 2021, 36, 666–674. [Google Scholar]
- Fu, B.; Ouyang, Z.; Shi, P.; Fan, J.; Wang, X.; Zheng, H.; Zhao, W.; Wu, F. Current Condition and Protection Strategies of Qinghai-Tibet Plateau Ecological Security Barrier. Bull. Chin. Acad. Sci. 2021, 36, 1298–1306. [Google Scholar]
- Wang, Z.; Zhang, Y.; Yang, Y.; Zhou, W.; Gang, C.; Zhang, Y.; Li, J.; An, R.; Wang, K.; Odeh, I. Quantitative assess the driving forces on the grassland degradation in the Qinghai–Tibet Plateau, in China. Ecol. Inform. 2016, 33, 32–44. [Google Scholar] [CrossRef]
- Wang, G.; Li, Y.; Wang, Y.; Wu, Q. Effects of permafrost thawing on vegetation and soil carbon pool losses on the Qinghai–Tibet Plateau, China. Geoderma 2008, 143, 143–152. [Google Scholar]
- Chen, B.; Zhang, X.; Tao, J.; Wu, J.; Wang, J.; Shi, P.; Zhang, Y.; Yu, C. The impact of climate change and anthropogenic activities on alpine grassland over the Qinghai-Tibet Plateau. Agric. For. Meteorol. 2014, 189, 11–18. [Google Scholar] [CrossRef]
- Wang, G.; Wang, Y.; Li, Y.; Cheng, H. Influences of alpine ecosystem responses to climatic change on soil properties on the Qinghai–Tibet Plateau, China. Catena 2007, 70, 506–514. [Google Scholar] [CrossRef]
- Cheng, G.; Wu, T. Responses of permafrost to climate change and their environmental significance, Qinghai-Tibet Plateau. J. Geophys. Res. Earth Surf. 2007, 112. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, L.; Liu, W.; Qi, Y.; Wo, X. Livestock-carrying capacity and overgrazing status of alpine grassland in the Three-River Headwaters region, China. J. Geogr. Sci. 2014, 24, 303–312. [Google Scholar] [CrossRef]
- Arthur, A.D.; Pech, R.P.; Jiebu, Z.Y.; Lin, H. Grassland degradation on the Tibetan Plateau: The role of small mammals and methods of control. Aust. Cent. Int. Agric. Res. Tech. Rep. 2007, 67, 1–35. [Google Scholar]
- Shao, Z.; Qin, Y. Advance in Studying the Influence of Plateau Pikas Disturbance on Carbon Cycle of Alpine Grasslands. Acta Agrestia Sin. 2022, 30, 1086–1094. [Google Scholar]
- Li, W.; Zhang, Y. Impacts of plateau pikas on soil organic matter and moisture content in alpine meadow. Acta Theriol. Sin. 2006, 26, 331. [Google Scholar]
- Smith, A.T.; Foggin, J.M. The plateau pika (Ochotona curzoniae) is a keystone species for biodiversity on the Tibetan plateau. Anim. Conserv. Forum 1999, 2, 235–240. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, Q.; Zhang, J.; Pang, X.; Xu, H.; Wang, J.; Guo, Z. Clipping by plateau pikas and impacts to plant community. Rangel. Ecol. Manag. 2020, 73, 368–374. [Google Scholar] [CrossRef]
- Pech, R.P.; Jiebu Arthur, A.D.; Yanming, Z.; Hui, L. Population dynamics and responses to management of plateau pikas Ochotona curzoniae. J. Appl. Ecol. 2007, 44, 615–624. [Google Scholar] [CrossRef]
- Song, Z.; Li, X.; Su, X.; Li, C. Analyzing the recovery mechanisms of patchy degradation and its response to mowing and plateau pika disturbances in alpine meadow. Ecol. Indic. 2023, 154, 110565. [Google Scholar] [CrossRef]
- Dong, Q.M.; Zhao, X.Q.; Wu, G.L.; Shi, J.J.; Ren, G.H. A review of formation mechanism and restoration measures of “black-soil-type” degraded grassland in the Qinghai-Tibetan Plateau. Environ. Earth Sci. 2013, 70, 2359–2370. [Google Scholar] [CrossRef]
- Sun, F.; Chen, W.; Liu, L.; Liu, W.; Cai, Y.; Smith, P. Effects of plateau pika activities on seasonal plant biomass and soil properties in the alpine meadow ecosystems of the Tibetan Plateau. Grassl. Sci. 2015, 61, 195–203. [Google Scholar] [CrossRef]
- Wei, W.; He, J.; Zheng, Q.; He, Y.; Oosthuizen, M.K. A comparison of density estimation methods in plateau pika populations in an alpine meadow ecosystem. J. Mammal. 2020, 101, 1091–1096. [Google Scholar] [CrossRef]
- Wei, W.; Yao, X.; Zhang, Y.; Zhen, Q.; Qin, M.; Tang, Z.; Oosthuizen, M.K.; Zhang, W. Vegetation restoration measures: Increasing plant height suppresses population densities of plateau pikas. Land Degrad. Dev. 2023, 34, 2201–2213. [Google Scholar] [CrossRef]
- Wang, X.; Li, J.; Li, J.; Guo, Z.; Pang, X. Plateau pika (Ochotona curzoniae) bioturbation triggers soil organic carbon loss by altering plant inputs and soil properties and their control on microbial composition. Catena 2025, 250, 108737. [Google Scholar] [CrossRef]
- Pang, X.P.; Wang, Q.; Guo, Z.G. The impact of the plateau pika on the relationship between plant aboveground biomass and plant species richness. Land Degrad. Dev. 2021, 32, 1205–1212. [Google Scholar] [CrossRef]
- Qi, Y.; Pu, X.; Li, Y.; Li, D.; Huang, M.; Zheng, X.; Guo, J.; Chen, Z. Prediction of Suitable Distribution Area of Plateau pika (Ochotona curzoniae) in the Qinghai–Tibet Plateau under Shared Socioeconomic Pathways (SSPs). Sustainability 2022, 14, 12114. [Google Scholar] [CrossRef]
- Hua, R.; Hua, L.; Tang, Z.; Dong, R.; Bao, D.; Ye, G.; La, M.; Sun, W.; Zhang, Z.; Wang, L. Maxent Modeling for Predicting Habitat Suitability and Potential Distribution of Plateau Pika (Ochotona curzoniae) on the Qinghai-Tibet Plateau, China. Rangel. Ecol. Manag. 2023, 87, 34–43. [Google Scholar] [CrossRef]
- Wang, Z.; Deng, Y.; Kang, Y.; Wang, Y.; Bao, D.; Tan, Y.; An, K.; Su, J. Impacts of climate change and human activities on three Glires pests of the Qinghai–Tibet Plateau. Pest Manag. Sci. 2024, 80, 5233–5545. [Google Scholar] [CrossRef]
- Wu, Y.; Ma, Y.; Liu, W.; Zhang, W. Modeling the spatial distribution of plateau pika (Ochotona curzoniae) in the Qinghai Lake Basin, China. Animals 2019, 9, 843. [Google Scholar] [CrossRef]
- Yi, S.; Chen, J.; Qin, Y. Preliminary application of aerial photos using UAV on the ecological studies of alpine grassland on Qinghai-Tibetan Plateau. J. Southwest Univ. Natl. 2016, 42, 1–7. [Google Scholar]
- Qin, Y.; Yi, S.; Ding, Y.; Qin, Y.; Zhang, W.; Sun, Y.; Hou, X.; Yu, H.; Meng, B.; Zhang, H. Effects of plateau pikas’ foraging and burrowing activities on vegetation biomass and soil organic carbon of alpine grasslands. Plant Soil 2021, 458, 201–216. [Google Scholar] [CrossRef]
- Du, J.; Yi, S.; Qin, Y.; Yu, H.; Ma, J.; Sun, Y.; Qin, Y. Distribution and cause of plateau pika (Ochotona curzoniae) burrows in Henan Mongolian Autonomous county, Qinghai Province. J. Anhui Agric. Univ. 2019, 46, 415–419. [Google Scholar]
- Guo, X.; Yi, S.; Qin, Y.; Chen, J. Habitat environment affects the distribution of plateau pikas: A study based on an unmanned aerial vehicle. Pratacultural Sci. 2017, 34, 1306–1313. [Google Scholar]
- Du, J.; Sun, Y.; Xiang, B.; Chen, J.; Qin, Y.; Hou, X.; Yu, H.; Yi, S. Potential distribution of plateau pika and its influence factors in the source region of the Yellow River Basin using BIOMOD. Pratacultural Sci. 2019, 36, 1074–1083+1917. [Google Scholar]
- Hua, R.; Zhou, R.; Baodaerhan Zhang, F.; Zhou, F.; Tang, Z.; Hua, L. Prediction of geographical distribution for plateau pikas and its potential response to climate change in Maqu. Grassl. Turf 2020, 40, 1–8. [Google Scholar]
- Zhao, J.; Jiang, C.; Ding, Y.; Li, G.; Li, Q. Potential distribution and disturbance intensity analysis of plateau pika in the source region of the Yellow River via BIOMOD2 integrated model. Chin. J. Ecol. 2024, 43, 1192–1201. [Google Scholar]
- Qu, J.; Russell, J.C.; Ji, W.; Yang, M.; Chen, Q.; Li, W.; Zhang, Y. Five-year population dynamics of plateau pikas (Ochotona curzoniae) on the east of Tibetan Plateau. Eur. J. Wildl. Res. 2017, 63, 51. [Google Scholar] [CrossRef]
- Smith, A.T. Conservation status of American pikas (Ochotona princeps). J. Mammal. 2020, 101, 1466–1488. [Google Scholar] [CrossRef]
- Peng, S.; Ding, Y.; Liu, W.; Li, Z. 1-km monthly precipitation dataset for China (1901–2023). Earth Syst. Sci. Data 2024, 11, 1931–1946. [Google Scholar] [CrossRef]
- Liu, X.; Chen, Q.; Wang, Y. The effect of grassland degeneration for ecological security and economie development in Gannan region of Gansu province. Pratacultural Sci. 2006, 23, 39–42. [Google Scholar]
- Yu, H.; Feng, Q.; Chen, S.; Han, L.; Huang, X.; Liang, T. Grassland biomass dynamic monitoring in Gannan prefecture based on the microwave vegetation indices. J. Lanzhou Univ. 2011, 47, 69–74. [Google Scholar]
- Liu, M.; Zhang, Y.; Li, Q.; Li, B.; Sun, R.; Song, J. Species abundance distribution characteristics of alpine meadow plant community in Gannan. China Environ. Sci. 2021, 41, 1405–1414. [Google Scholar] [CrossRef]
- Yi, S. FragMAP: A tool for long-term and cooperative monitoring and analysis of small-scale habitat fragmentation using an unmanned aerial vehicle. Int. J. Remote Sens. 2017, 38, 2686–2697. [Google Scholar] [CrossRef]
- Qu, J.; Li, W.; Yang, M.; Ji, W.; Zhang, Y. Life history of the plateau pika (Ochotona curzoniae) in alpine meadows of the Tibetan Plateau. Mamm. Biol. 2013, 78, 68–72. [Google Scholar] [CrossRef]
- Zhang, W.; Jin, S.; Yu, C.; Pang, X.; Wang, J.; Guo, Z. Influence of the density of burrow entrances of plateau pika on the concentration of soil nutrients in a Kobresia pygmaea meadow. Pratacultural Sci. 2018, 35, 1593–1601. [Google Scholar]
- Wei, W. The Relationship between Population Density, Vegetation Community Structure and Predation Risk of Plateau Pika. Acta Agrestia Sin. 2019, 27, 350–355. [Google Scholar]
- Hua, R.; Bao d Dong, R.; Tang, Z.; Chu, B.; Hao, Y.; Hua, L. Monitoring of rodent damage areas in grassland using unmanned aerial vehicle remote sensing technology. Acta Prataculturae Sin. 2023, 32, 71–82. [Google Scholar]
- Liu, J.; Si, W. Correlation between population density of plateau pika and community structure of grassland. J. South. Agric. 2012, 43, 2083–2086. [Google Scholar]
- Chen, J.; Wang, Z.; Wang, Y.; Li, B.; Zhaxi, X.Z.; Luosangdawa, Z.M. Methods for investigating the density of the plateau pika in Northern Tibetan Plateau. Plant Prot. 2008, 34, 114–117. [Google Scholar]
- Yu, H. Investigation on The Field Route of Rodent Damage in The Grassland of Huangnan State in Qinghai Provine. Qinghai Prataculture 2018, 27, 36–40. [Google Scholar]
- Sun, F.; Long, R.; Lu, C. Effects of plateau pikas (Ochotona curzoniae) burrow densities on plant community composition and population diversity in alpine meadow. J. Arid. Land Resour. Environ. 2010, 24, 181–186. [Google Scholar]
- Li, B.; Jiebu Li, S.; Tao, Y. The survey report of main meadow type mouse of north plateau in Tibet. Tibet. Sci. Technol. 2007, 3, 29–30. [Google Scholar]
- Gong, A.; Zhang, S.; Li, Q. Discussion on population types of redents and damage in alpine meadow grassland in QingHai. Qinghai Prataculture 2003, 12, 19–23. [Google Scholar]
- Liu, W.; Wang, X.; Zhou, L.; Zhou, H. Studies on Destruction, Prevention and Control of Plateau Pikas in Kobresia pygmaea Meadow. Acta Theriol. Sin. 2003, 23, 214–219. [Google Scholar]
- Yi, S.; Chen, J.; Qin, Y.; Xu, G. The burying and grazing effects of plateau pika on alpine grassland are small: A pilot study in a semiarid basin on the Qinghai-Tibet Plateau. Biogeosciences 2016, 13, 6273–6284. [Google Scholar] [CrossRef]
- Villén Peréz, S.; Heikkinen, J.; Salemaa, M.; Mäkipää, R. Global warming will affect the maximum potential abundance of boreal plant species. Ecography 2020, 43, 801–811. [Google Scholar] [CrossRef]
- Cade, B.S.; Noon, B.R.; Flather, C.H. Quantile regression reveals hidden bias and uncertainty in habitat models. Ecology 2005, 86, 786–800. [Google Scholar] [CrossRef]
- Legendre, P.; Gallagher, E.D. Ecologically meaningful transformations for ordination of species data. Oecologia 2001, 129, 271–280. [Google Scholar] [CrossRef]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Hijmans, R.J.; Phillips, S.; Leathwick, J.; Elith, J. dismo: Species Distribution Modeling, R Package Version 1.3-14. 2023. Available online: https://CRAN.R-project.org/package=dismo (accessed on 26 September 2023).
- Huang, H.; Zhou, L.; Chen, J.; Wei, T. ggcor: Extended Tools for Correlation Analysis and Visualization, R Package Version 0.9.7. 2020.
- Lai, J.; Zou, Y.; Zhang, J.; Peres-Neto, P.R. Generalizing hierarchical and variation partitioning in multiple regression and canonical analyses using the rdacca. hp R package. Methods Ecol. Evol. 2022, 13, 782–788. [Google Scholar] [CrossRef]
- Qin, Y.; Huang, B.; Zhang, W.; Yu, Y.; Yi, S.; Sun, Y. Pikas burrowing activity promotes vegetation species diversity in alpine grasslands on the Qinghai-Tibetan Plateau. Glob. Ecol. Conserv. 2021, 31, e01806. [Google Scholar] [CrossRef]
- Sumbh, O.; Hof, A.R. Can pikas hold the umbrella? Understanding the current and future umbrella potential of keystone species Pika (Ochotona spp.). Glob. Ecol. Conserv. 2022, 38, e02247. [Google Scholar] [CrossRef]
- Song, Y.; Chong, H.; Wang, D.; Xiao, H. Simulation and response mechanism of potential geographical distribution of plateau pika Ochotona curzoniae under climate change. Glob. Ecol. Conserv. 2025, 57, e03409. [Google Scholar] [CrossRef]
- Tessarolo, G.; Lobo, J.M.; Rangel, T.F.; Hortal, J. High uncertainty in the effects of data characteristics on the performance of species distribution models. Ecol. Indic. 2021, 121, 107147. [Google Scholar] [CrossRef]
- Guisan, A.; Thuiller, W. Predicting species distribution: Offering more than simple habitat models. Ecol. Lett. 2005, 8, 993–1009. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.; Li, C.; Lin, K.; Fu, H.; Zhang, Z. Current situation, research progress and management strategies of plant diseases, insect pests, and rodent pests in grasslands of China. Bull. Natl. Nat. Sci. Found. China 2023, 37, 580–586. [Google Scholar]
- Li, J.; Li, X.; Gao, J.; Ma, G.; Qi, X. Micro-scale fragmentation of the alpine meadow landscape on the Qinghai-Tibet Plateau under external disturbances. Catena 2021, 201, 105220. [Google Scholar] [CrossRef]
- Pan, X.; Migmar, W.-D. Plateau pika ecology: A review. Chin. J. Ecol. 2016, 35, 7. [Google Scholar]
- Wang, X.; Smith, A.T. On the natural winter mortality of the plateau pika (Ochotona curzoniae). Acta Theriol. Sin. 1988, 8, 152–156. [Google Scholar]
- Nie, H. Study on the Evolutionary Ecology of Small Herbivorous Ammals: Life History Strategy of Plateau Pika (Ochotona curzoniae). Ph.D. Thesis, Zhejiang University, Hangzhou, China, 2005. [Google Scholar]
- Wang, J.; Zhang, Y.; Wang, D. Seasonal thermogenesis and body mass regulation in plateau pikas (Ochotona curzoniae). Oecologia 2006, 149, 373–382. [Google Scholar] [CrossRef]
- Wang, D.; Sun, R.; Wang, Z.; Liu, J. Effects of temperature and photoperiod on thermogenesis in plateau pikas (Ochotona curzoniae) and root voles (Microtus oeconomus). J. Comp. Physiol. B 1999, 169, 77–83. [Google Scholar] [CrossRef]
- Yi, S.; Cao, W.; Zhang, J.; Qin, Y.; Sun, Y.; Qin, Y. Research Progress on Plateau Pika Dynamic Distribution in the Context of Climate Change and Human Activities. J. Nantong Univ. 2020, 19, 16–30. [Google Scholar]
- Zong, H.; Xia, W.; Sun, D. The influence of a heavy snow on the population density of small mammals. Acta Biol. Plateau Sin. 1986, 5, 85–90. [Google Scholar]
- Speakman, J.R.; Chi, Q.; Ołdakowski, Ł.; Fu, H.; Fletcher, Q.E.; Hambly, C.; Togo, J.; Liu, X.; Piertney, S.B.; Wang, X. Surviving winter on the Qinghai-Tibetan Plateau: Pikas suppress energy demands and exploit yak feces to survive winter. Proc. Natl. Acad. Sci. USA 2021, 118, e2100707118. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Yang, Q.; Bai, Z.; Zhou, W.; Semenza, G.L.; Ge, R. Chronic cold exposure results in subcutaneous adipose tissue browning and altered global metabolism in Qinghai-Tibetan plateau pika (Ochotona curzoniae). Biochem. Biophys. Res. Commun. 2018, 500, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Bhandari, J.; Zhang, Y. Effect of altitude and soil properties on biomass and plant richness in the grasslands of Tibet, China, and Manang District, Nepal. Ecosphere 2019, 10, e02915. [Google Scholar] [CrossRef]
- Sun, J.; Zhang, Z.; Dong, S. Adaptive management of alpine grassland ecosystems over Tibetan Plateau. Pratacultural Sci. 2019, 36, 933–938. [Google Scholar]
- Smith, A.T.; Badingqiuying Wilson, M.C.; Hogan, B.W. Functional-trait ecology of the plateau pika Ochotona curzoniae in the Qinghai–Tibetan Plateau ecosystem. Integr. Zool. 2019, 14, 87–103. [Google Scholar] [CrossRef]
- Wei, W.; Zhang, L.; Yang, G.; Xu, J.; Fan, X.; Zhang, W. A study on the burrow features and functions of plateau pika. Acta Prataculturae Sin. 2013, 22, 198–204. [Google Scholar]
- Ren, Y.; Liu, J.; Liu, S.; Wang, Z.; Liu, T.; Shalamzari, M.J. Effects of Climate Change on Vegetation Growth in the Yellow River Basin from 2000 to2019. Remote Sens. 2022, 14, 687. [Google Scholar] [CrossRef]
- Hu, R.; Chen, X.; Chen, J.; Zhang, S.; Kuang, Y.; Yu, H.; Ji, H.; Zhao, X.; Yi, S.; Meng, B.; et al. MODIS NDVI saturation assessment of alpine meadow grassland biomass estimation using remote sensing: A case study in the eastern edge of the Qinghai-Tibet Plateau. Acta Ecol. Sin. 2024, 44, 6357–6372. [Google Scholar]
- Zhang, H.; Tang, Z.; Wang, B.; Meng, B.; Qin, Y.; Sun, Y.; Lv, Y.; Zhang, J.; Yi, S. A non-destructive method for rapid acquisition of grassland aboveground biomass for satellite ground verification using UAV RGB images. Glob. Ecol. Conserv. 2022, 33, e01999. [Google Scholar] [CrossRef]
- Kan, H.; Zhang, H.; Meng, B.; Shuhua, Y. LiDAR flight parameter influence on low vegetation height inversion: A case study of DJI Zenmuse L1. Pratacultural Sci. 2023, 40, 2513–2527. [Google Scholar]
- Tang, Z.; Zhang, Y.; Zheng, Z.; Cong, N.; Zhu, Y.; Chen, Y.; Gao, J.; Zhu, W. Grazing affects ecosystem traits by regulating plateau pika activities at the landscape scale. Sci. Total Environ. 2024, 946, 174356. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Yi, S.; Hou, F.; Luo, D.; Hu, J.; Zhou, Z. Quantifying the dynamics of livestock distribution by unmanned aerial vehicles (UAVs): A case study of yak grazing at the household scale. Rangel. Ecol. Manag. 2020, 73, 642–648. [Google Scholar] [CrossRef]
- Zhang, H.J.; Xie, J.X.; Li, X.L. The distribution of the plateau pika and correlation with soil properties of Henan County in Qinghai. Qinghai Univ. (Nat. Sci. Ed.) 2010, 34, 14–21. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Area | Ceph | Cpd | References |
|---|---|---|---|
| Gannan Tibetan Autonomous Prefecture | 0.38 ± 0.14 | - | This study |
| Maqu county | 0.44 | 0.46 | [43,44,45] |
| Yushu prefecture | 0.74 | 0.37 | [46] |
| Naqu county | 0.48 | 0.11 | [47] |
| Huangnan prefecture | 0.27 | - | [48] |
| Maqin county | - | 0.175 | [49] |
| Naqu county | 0.26 | 0.25 | [50] |
| Qinghai | - | 0.27 | [51] |
| Dari county | - | 0.22 | [52] |
| Tianjun county | 0.29 | 0.44 | [53] |
| Average | 0.41 | 0.29 | - |
| Categories | Hazard | EPH (ha−1) | Pika Density (ha−1) |
|---|---|---|---|
| I | Without pika | 0~3 | 0 |
| II | No hazard | 4~8 | 1.16~2.32 |
| III | Slight hazard | 9~30 | 2.61~8.7 |
| IV | Moderate hazard | 31~100 | 8.99~29 |
| V | Severe hazard | ≥101 | ≥29.29 |
| Symbol (Unit) | Description |
|---|---|
| Bio1 (°C) | Annual Mean Temperature |
| Bio2 (°C) | Mean Diurnal Range (Mean of monthly (max temp − min temp)) |
| Bio3 (-) | Isothermality (BIO2/BIO7) (×100) |
| Bio4 (°C) | Temperature Seasonality (standard deviation × 100) |
| Bio5 (°C) | Max Temperature of Warmest Month |
| Bio6 (°C) | Min Temperature of Coldest Month |
| Bio7 (°C) | Temperature Annual Range (BIO5-BIO6) |
| Bio8 (°C) | Mean Temperature of Wettest Quarter |
| Bio9 (°C) | Mean Temperature of Driest Quarter |
| Bio10 (°C) | Mean Temperature of Warmest Quarter |
| Bio11 (°C) | Mean Temperature of Coldest Quarter |
| Bio12 (mm) | Annual Precipitation |
| Bio13 (mm) | Precipitation of Wettest Month |
| Bio14 (mm) | Precipitation of Driest Month |
| Bio15 (-) | Precipitation Seasonality (Coefficient of Variation) |
| Bio16 (mm) | Precipitation of Wettest Quarter |
| Bio17 (mm) | Precipitation of Driest Quarter |
| Bio18 (mm) | Precipitation of Warmest Quarter |
| Bio19 (mm) | Precipitation of Coldest Quarter |
| Pika Density | 2016 | 2019 | 2022 |
|---|---|---|---|
| I | 0 a | 0 a | 0 a |
| II | 1.42 ± 0.48 a | 1.33 ± 0.45 a | 1.36 ± 0.39 a |
| III | 5.12 ± 1.83 a | 5.33 ± 2.21 a | 5.08 ± 1.80 a |
| IV | 17.03 ± 5.68 a | 19.01 ± 6.24 a | 15.88 ± 5.22 a |
| V | 65.36 ± 23.45 a | 79.04 ± 67.11 a | 61.37 ± 47.89 a |
| Mean | 9.87 ± 21.03 a | 14.43 ± 37.79 a | 11.20 ± 26.72 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, X.; Qin, Y.; Sun, Y.; Yi, S. Monitoring Plateau Pika and Revealing the Associated Influencing Mechanisms in the Alpine Grasslands Using Unmanned Aerial Vehicles. Drones 2025, 9, 298. https://doi.org/10.3390/drones9040298
Liu X, Qin Y, Sun Y, Yi S. Monitoring Plateau Pika and Revealing the Associated Influencing Mechanisms in the Alpine Grasslands Using Unmanned Aerial Vehicles. Drones. 2025; 9(4):298. https://doi.org/10.3390/drones9040298
Chicago/Turabian StyleLiu, Xinyu, Yu Qin, Yi Sun, and Shuhua Yi. 2025. "Monitoring Plateau Pika and Revealing the Associated Influencing Mechanisms in the Alpine Grasslands Using Unmanned Aerial Vehicles" Drones 9, no. 4: 298. https://doi.org/10.3390/drones9040298
APA StyleLiu, X., Qin, Y., Sun, Y., & Yi, S. (2025). Monitoring Plateau Pika and Revealing the Associated Influencing Mechanisms in the Alpine Grasslands Using Unmanned Aerial Vehicles. Drones, 9(4), 298. https://doi.org/10.3390/drones9040298