1. Introduction
Herbivorous insects are natural catalysts for disturbance within forest ecosystems. However, the increasing frequency and heightened virulence of outbreaks, attributed to climate warming, have raised significant concerns in forest management [
1,
2]. Insect outbreaks have impacts on forest resilience and crucial forest ecosystem services, including carbon sequestration and timber production [
3,
4]. The effects of outbreaks on host trees span a spectrum, ranging from growth decline to eventual mortality. Moreover, they often interact with other pathogens and drought stress, leading to non-lineal responses and excessive mortality [
5]. In the Mediterranean region, one of the major defoliating insects affecting pines and cedars is the pine processionary moth (PPM,
Thaumetopoea pityocampa) [
6]. PPM defoliations are widespread in Spanish forests, annually affecting 500,000 ha [
7], reducing tree growth and subsequently its carbon storage capacity [
4].
The life cycle of PPM commences in summer when adult female moths, which have a brief lifespan of approximately one night, lay up to 300 eggs on pine needles after mating [
8]. These eggs then hatch into larvae, which proceed to feed on pine needles throughout the winter. Once the fifth and last instar of the larval development stage starts concluding, the larvae organize into processions and move from the canopy to the ground, undergo pupation in the soil, and eventually give rise to new adult moths in the following summer. Alternatively, some may enter a short or prolonged diapause [
9]. PPM has the potential to instigate large-scale outbreaks characterized by intense defoliation, although it rarely causes tree mortality, underscoring the high resilience of tree host species to this endemic pest. Nevertheless, it does curtail tree growth and reproductive capacity [
4,
10]. Additionally, the caterpillar hairs can induce severe allergic reactions for humans and animals, resulting in skin lesions or respiratory difficulties, posing a threat to public health [
11]. The population size of PPM is significantly influenced by environmental conditions, such as climatic factors, interactions with other species, and the composition and structure of the forests [
12,
13]. The performance of PPM is strongly constrained by temperature, with lethal thresholds for PPM larvae ranging from −12 to −16 °C in winter [
14,
15] and for PPM eggs between 36 and 42 °C in summer [
16,
17]. Warmer winters are expanding PPM outbreaks northward and to higher elevations [
15], while adverse effects have also been observed due to the increasing severity of heat events [
18,
19,
20]. Precipitation levels influence the lignin content of pine needles, leading, in cases of water stress, to lower nutritional properties that adversely affect PPM populations [
10,
21]. Furthermore, there is consensus on the high vulnerability of
Pinus nigra [
22] over other species within the same genus. Forest composition, generally measured by species richness, reduces PPM abundance particularly when non-host tree species are present [
23,
24,
25]. PPM performance also depends on forest structure, increasing its incidence in structurally simple pine forests such as monospecific plantations [
13]. Less consensus has been found on the effects of forest age [
12]. Tree location at the landscape scale matters, causing forest edges and isolated trees to be more vulnerable to PPM defoliation [
26]. Consequently, given the anticipated rise in temperatures and the frequency of extreme events, which may result in unpredictable PPM development, it is paramount to develop cost-effective tools for monitoring the spatiotemporal effects on conifer forests that serve as base information for forest composition and structure management.
Practically, the monitoring of PPM traditionally relies on visual assessments by forestry technicians or photo interpreters gauging the degree of infestation in the field [
8]. In Spain, for instance, about 70% of regions employ PPM monitoring programs, predominantly based on in situ surveys. These programs are supplemented by additional assessments on specific plots by the International Co-operative Programme on Assessment and Monitoring of Air Pollution Effects on Forests (ICP Forest). While in situ surveys are valuable, their associated costs, resource demands and, at times, temporal intervals may constrain effective forest management actions. The integration of remote sensing, leveraging the rapid advancements in platforms and sensors, provides quantitative insights that not only complements in situ surveys but also enhances the spatiotemporal frequency of observations.
Remote sensing has gained significant attention for analyzing insect disturbances and monitoring their dynamics across three key species groups: bark beetles and broadleaved and coniferous defoliators [
27]. Notably, time series data acquired by the Moderate Resolution Imaging Spectroradiometer (MODIS) have been employed to monitor PPM defoliations in homogeneous managed stands of
Pinus pinaster at southwest France [
28] with correlations up to 0.9 when using the enhanced vegetation index. Likewise, Landsat data have been utilized to significantly differentiate up to three defoliation levels in northeastern [
29] and southeastern [
30] Spain, with the moisture stress index (MSI), normalized difference infrared index (NDII), and normalized burn ratio (NBR) being the most suitable spectral indices for this purpose. The increased availability of drone derived data has introduced novel spatial scales for analyzing PPM defoliation at the individual tree level. RGB orthomosaics showed a 79% accuracy in discriminating between defoliated and non-defoliated trees in two study sites dominated by
Pinus sylvestris,
Pinus halepensis, and
Pinus nigra at northeast Spain (Solsona) [
8]. Multispectral drone information was used to determine three defoliation classes with 82% accuracy in a mixed forest dominated by
Quercus ilex and
Pinus sylvestris at northeast Spain (Solsona) [
31,
32]. Additionally, Otsu et al. [
33] calibrated Landsat 8 with drone multispectral imagery to detect PPM defoliations. While much of the literature concentrates on PPM detection and the quantification of its infestation levels, García et al. [
34] explored the use of drone-derived RGB data in conjunction with artificial intelligence models to detect PPM nests. Despite previous studies having predominantly focused on the analysis of PPM defoliations and their effects based on two-dimensional data, there has been limited attention given to the three-dimensional changes in forest cover due to PPM disturbances. This is crucial to better understand the links between coverage change and tree growth reduction. This scarcity may be attributed to the lack of high-density 3D multitemporal flights, such as those derived from LiDAR sensors carried out by drones. This information presents innovative perspectives for addressing questions such as what are the effects of insect defoliation on crown morphology and forest structure? What are the volume changes (e.g., crown volume) caused by an insect outbreak? These inquiries contribute to the monitoring of forest ecosystems, providing pertinent information for effective forest management actions.
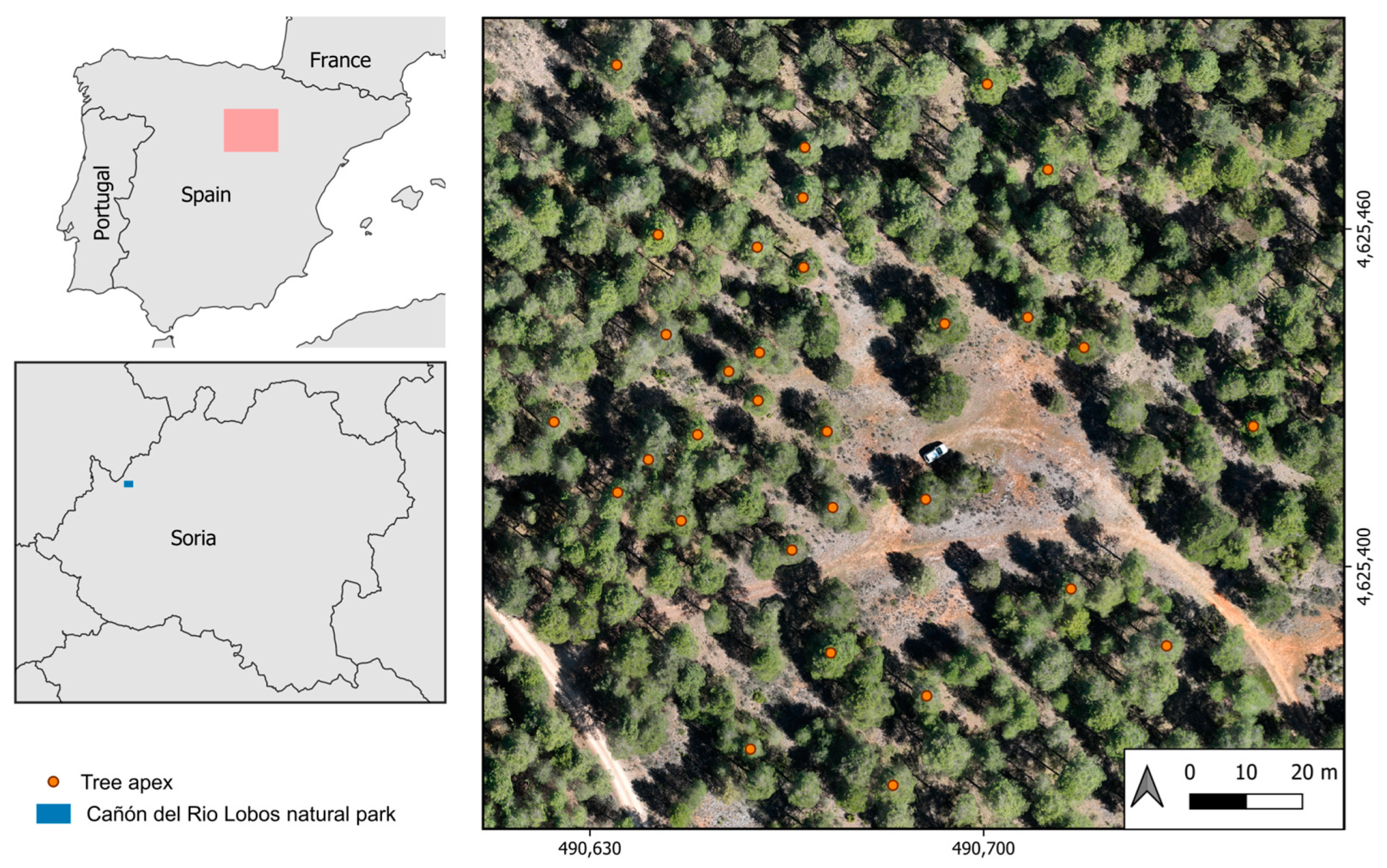
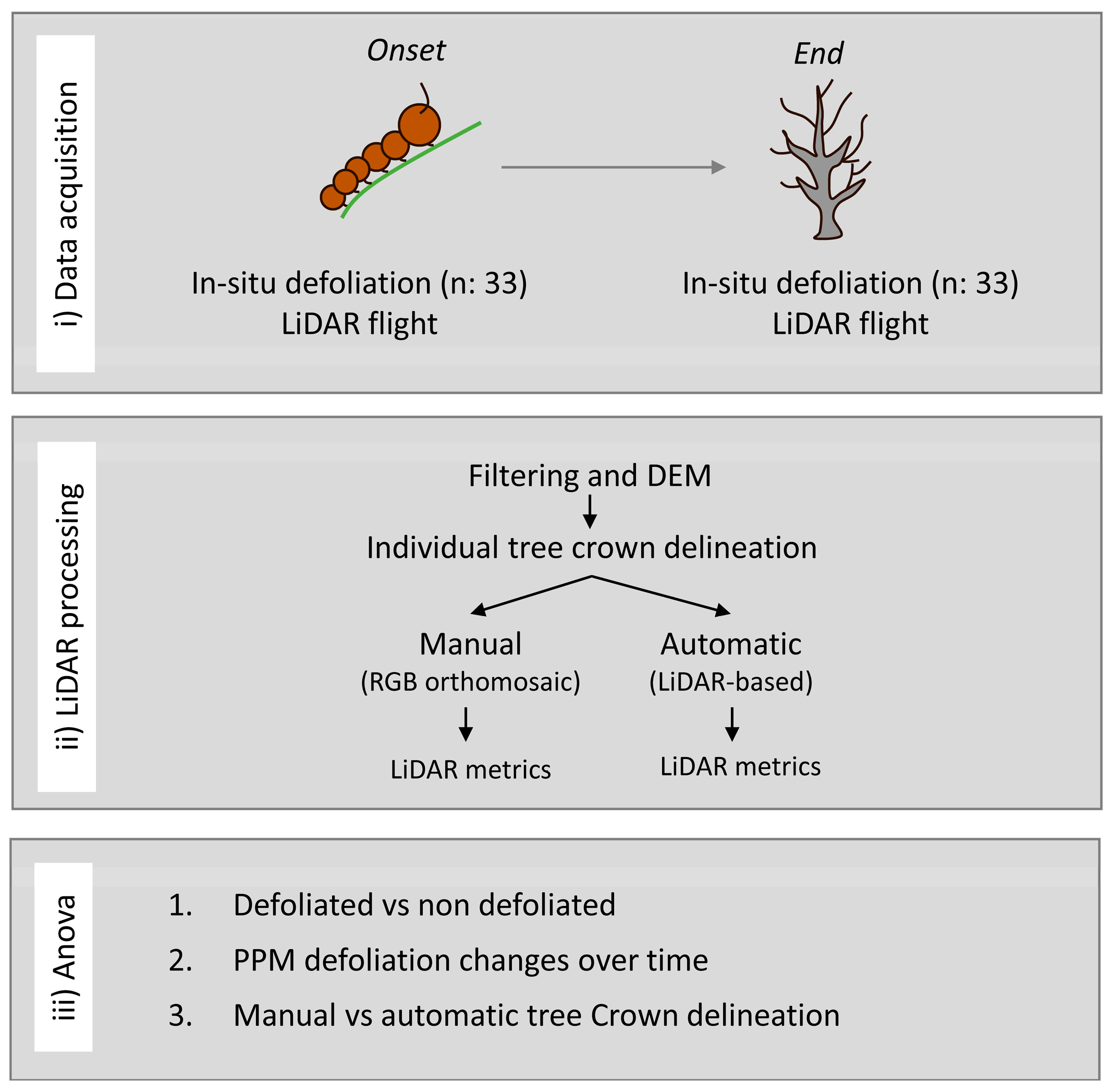
In this study, our primary objective is to assess the efficacy of high-density drone LiDAR-derived point clouds in quantifying the impact of PPM on canopy structure and its development over time at individual tree level scale. In particular, this study attempts to (i) investigate the existence of structural differences between non-defoliated and PPM-defoliated trees, (ii) evaluate the potential of drone-derived LiDAR data to discern structural changes attributable to PPM defoliation over time (i.e., along a feeding winter period), and (iii) analyze the disparities between manual tree crown detection and automatized LiDAR-derived crowns concerning structural metrics associated with PPM defoliations. To achieve these objectives, our focus is on a black pine (Pinus nigra subsp. salzmannii) forest, as it represents a highly susceptible species to PPM defoliations in the Mediterranean environment.
4. Discussion
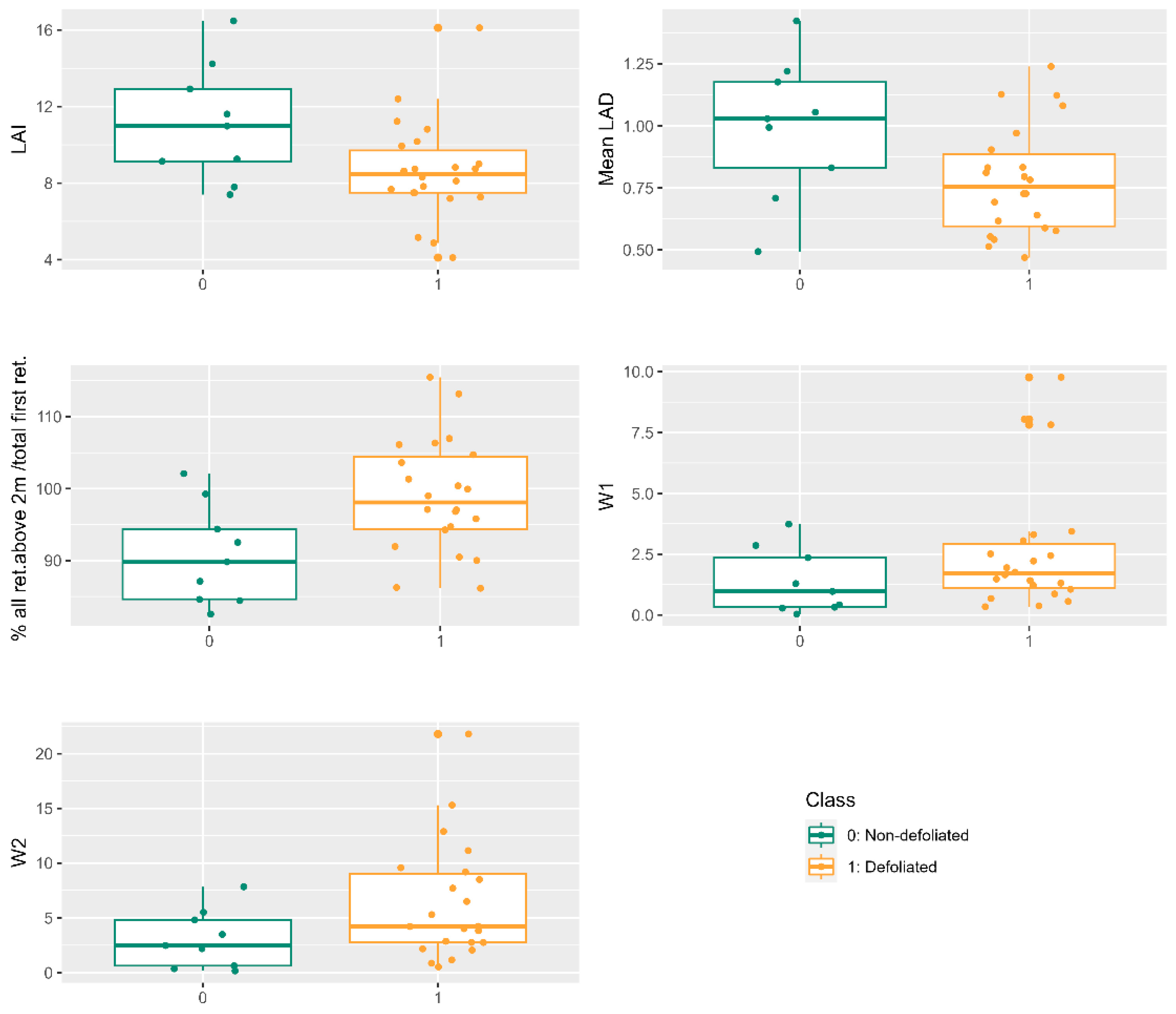
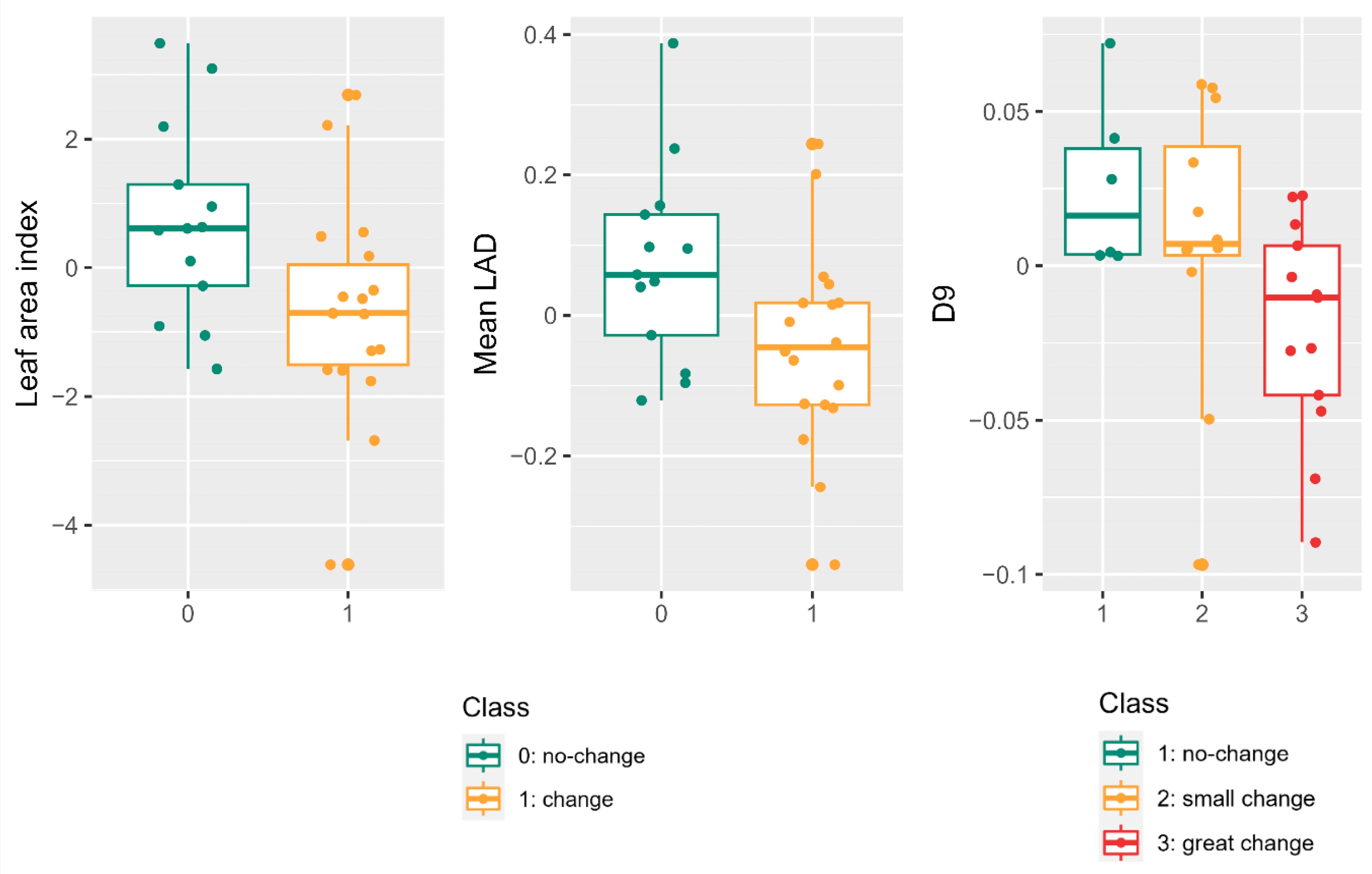
The impact of PPM in pine forests manifests in noteworthy alterations to tree crown density. The reduction in pine needles alters plant dynamics and nutrient cycling, resulting in diminished radial tree growth and volume [
52,
53]. PPM-induced defoliations lead to substantial reductions in leaf area index and mean leaf area density, reaching up to 23% when comparing non-defoliated to defoliated trees. The concurrent decrease in upper canopy density enhances LiDAR pulse penetration, yielding a notably higher percentage of returns in lower tree canopy branches. Moreover, our study unveils evident morphological transformations throughout the winter feeding period, showing statistically significant differences in two to three classes of defoliation change using drone LiDAR-derived canopy density metrics. These results hold for LiDAR-based crown delineation and provide comparable information to manual delineations. Such findings offer essential insights into the development of dynamic replicas or digital twins, facilitating enhanced forest ecosystem monitoring and informed decision-making by forest managers.
The assessment of defoliation through drone LiDAR point clouds enables the discrimination of non-defoliated and defoliated trees at the individual level. Statistically significant differences were identified for defoliation levels below or above 15%, consistent with previous assessments that employed drone multispectral images and determined a 20% defoliation threshold as optimal for recognizing defoliated trees [
8]. Both LAI and mean leaf area density LiDAR metrics exhibited significant reductions in the upper canopy due to PPM infestation, aligning with findings from Solberg [
54]. Solberg utilized LAI derived from high-density airborne laser scanning (ALS) data to estimate defoliations caused by
Neodiprion sertifer and compared the results to forest cutting practices. PPM infestation prompted increased pulse penetration to lower canopy strata, resulting in significant modifications to canopy morphology resembling changes induced by low–medium–severity forest fires [
55]. Interestingly, two metrics related to the cumulative proportion of returns in lower percentages of maximum elevation (W1 and W2) proved more effective than fixed height strata thresholds (e.g., returns between 3 and 4 m) in capturing the defoliation-induced changes in tree structure. The utilization of LiDAR drone technology opens new avenues for analyzing defoliations at the individual tree scale, expanding the range of available drone tools [
31,
32]. This underscores LiDAR’s capacity to provide a three-dimensional perspective on canopy structure changes.
PPM defoliation instigates morphological transformations throughout the winter, distinctly discernible through multitemporal LiDAR drone-derived metrics. LAI and mean leaf area density exhibited significant differences, particularly in determining defoliation changes exceeding 10%, a common threshold applied in field measurements. The percentage of returns in the upper canopy layer (D9) proved valuable in detecting up to three levels of defoliation change, although caution is warranted due to potential ambiguities between no-change and small-change detections [
8]. Prior research has often relied on multitemporal datasets from middle-resolution (e.g., Landsat) and low spatial resolution (e.g., MODIS) multispectral sensors for monitoring PPM outbreaks. Spectral changes observed throughout the PPM cycle enable the detection and characterization of infestation levels [
28,
29,
30], typically categorizing them into two to three levels, mirroring those determined with drone LiDAR. While three-dimensional datasets are progressively becoming available, those derived from ALS remain relatively scarce [
41]. Drone LiDAR has opened new opportunities for tailored multitemporal flights, augmenting spatiotemporal monitoring frequencies and facilitating fine change detections of PPM defoliations, akin to field measurements (i.e., 10% change).
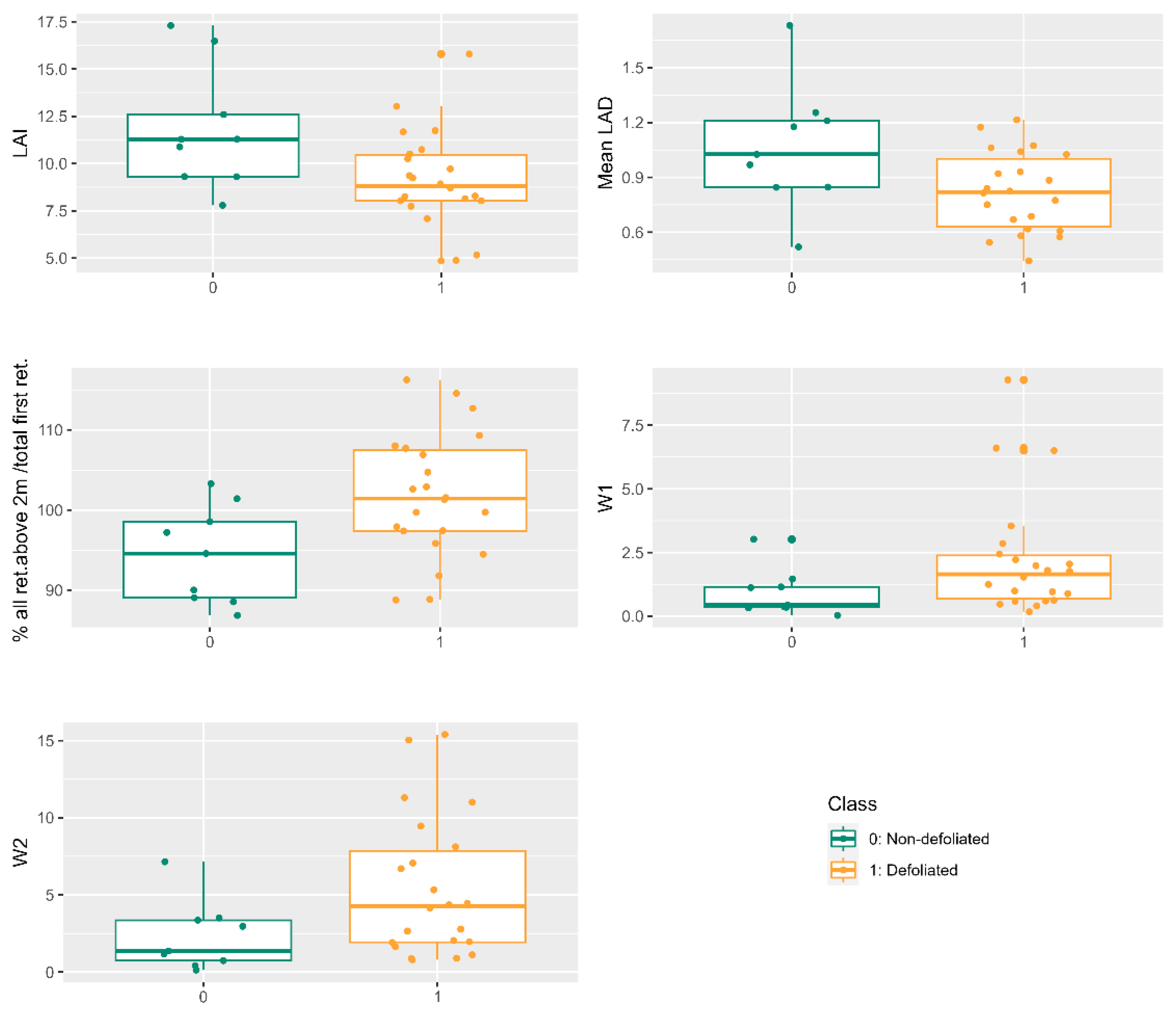
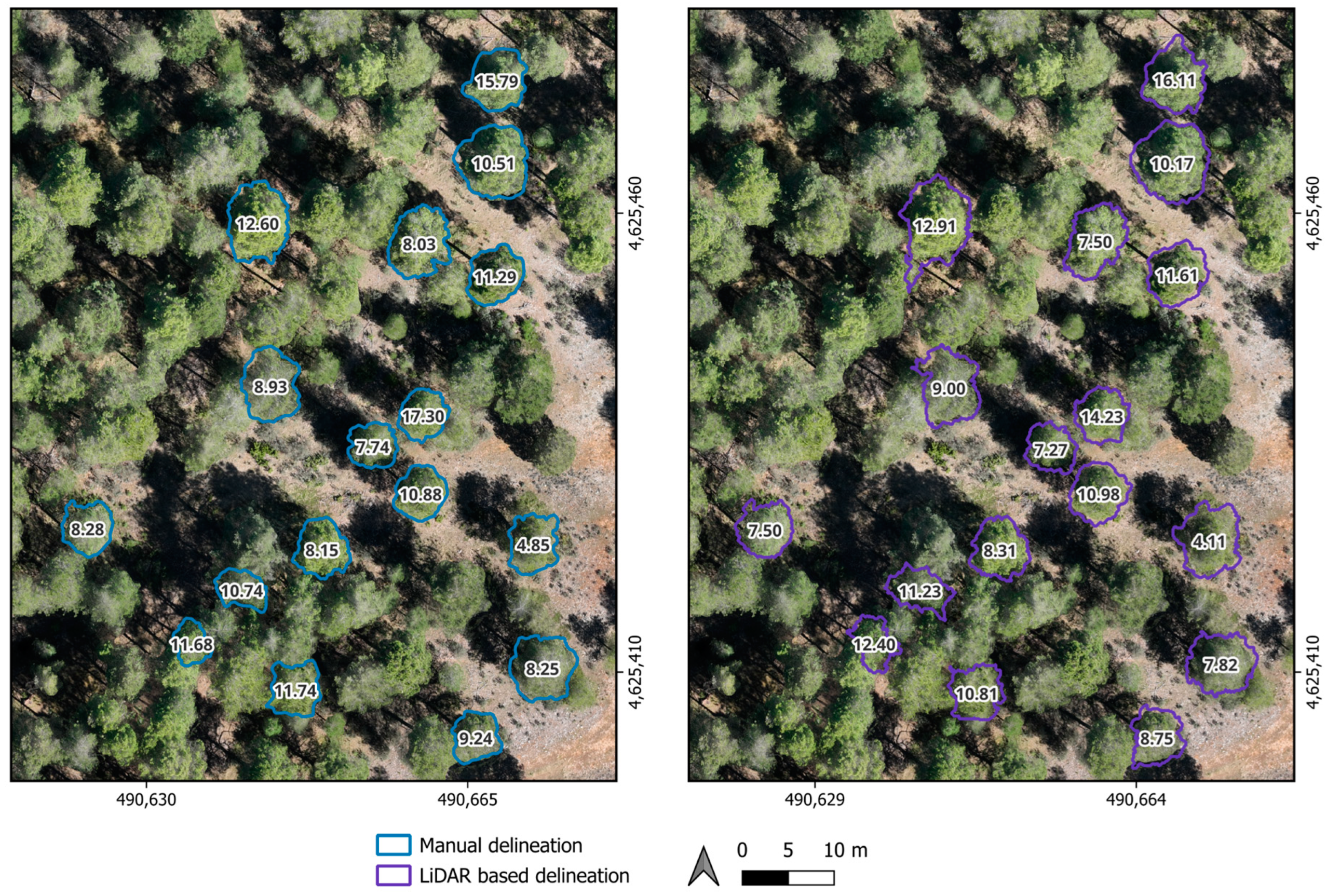
Automated tree-crown delineation from 3D drone LiDAR-derived point clouds yields comparable information to manual delineation in monitoring PPM defoliation. Though our focus does not center on identifying the optimal automatic crown delineation algorithm, we have observed that commonly employed methods such as tree top [
39] and tree crown watershed segmentation [
40], deliver accurate LiDAR derived metrics. LAI, the mean leaf area density, the percentage of all returns above 2 m concerning the total first returns, and metrics W1 and W2 exhibit no significant differences compared to manual delineations. However, there are notable distinctions in the upper canopy layer density (D9) metric between manual and automatic delineations, necessitating further investigation across more representative spatial scales. Despite additional examinations using alternative automatic delineation methodologies and encompassing a broader range of forest stand densities are warranted, automatic LiDAR delineation appears promising for monitoring PPM defoliation outbreaks over larger areas.
The present study underscores the utility of LiDAR drone data in delineating morphological changes in tree crowns resulting from PPM defoliations within a Mediterranean forest environment dominated by Pinus nigra, a highly PPM palatable species. Periodic in situ defoliation sampling at tree level and the capturing of multitemporal LiDAR flights are costly and labor intensive. This study shows that LiDAR metrics are sensitive to changes in crown morphology caused by PPM defoliations. The sample size of our study may have potentially overlooked further LiDAR metrics sensitive to PPM-induced changes in crown morphology. Despite this sample size, those LiDAR metrics that showed statistical differences in ANOVA analyses and fulfilled the normality and equal variance hypothesis, also showed significant differences in non-parametric tests (no shown), which indicates that these variables are in fact sensitive to PPM defoliations. Future research should also focus on the monitoring of wider areas (in situ and with remote sensing data) to develop statistical models to further scale the results to larger areas. Such analysis cannot be undertaken with the present dataset due to limitations in our sample size. However, our results provide useful insights for modeling efforts in larger areas, presenting a set of LiDAR metrics to prioritize when developing predictive models. As high-density 3D LiDAR drone point clouds continue to represent a novel source of information, future research endeavors should prioritize their integration with multispectral drone-derived datasets, high-resolution satellite imagery or intensity-derived LiDAR metrics to characterize PPM defoliations at broader scales. Our study quasi covers a major feeding period in winter, but some specific trees presented defoliation levels during the end of December, and further research should focus on the complete PPM cycle period. Considering the effectiveness of specific LiDAR metrics for PPM monitoring, the utilization of photogrammetric point clouds may prove advantageous due to the increased affordability of RGB or multispectral sensors. Overall, future endeavors should focus on multi-scale and multi-sensor approaches, from in situ measurements up to satellite datasets, to enable near-real time monitoring and early warming detection for better forest management.


 ,
, 



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}