
Drone-Based Identification and Monitoring of Two Invasive Alien Plant Species in Open Sand Grasslands by Six RGB Vegetation Indices

 ,
,  , ,
, ,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Studied Species
2.1.1. Common Milkweed—Asclepias syriaca
2.1.2. Indian Blanket Flower—Gaillardia pulchella
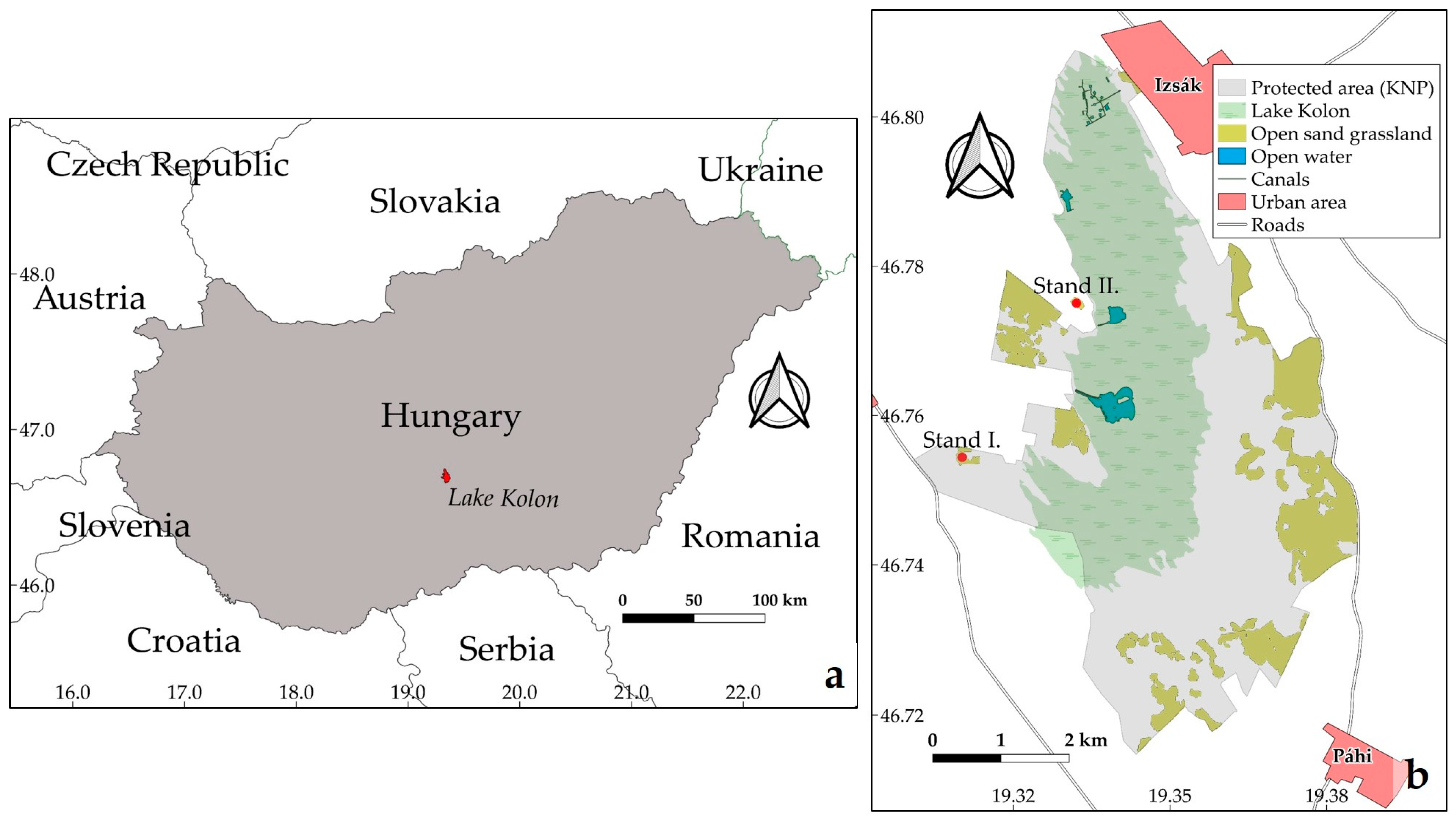
2.2. Study Area
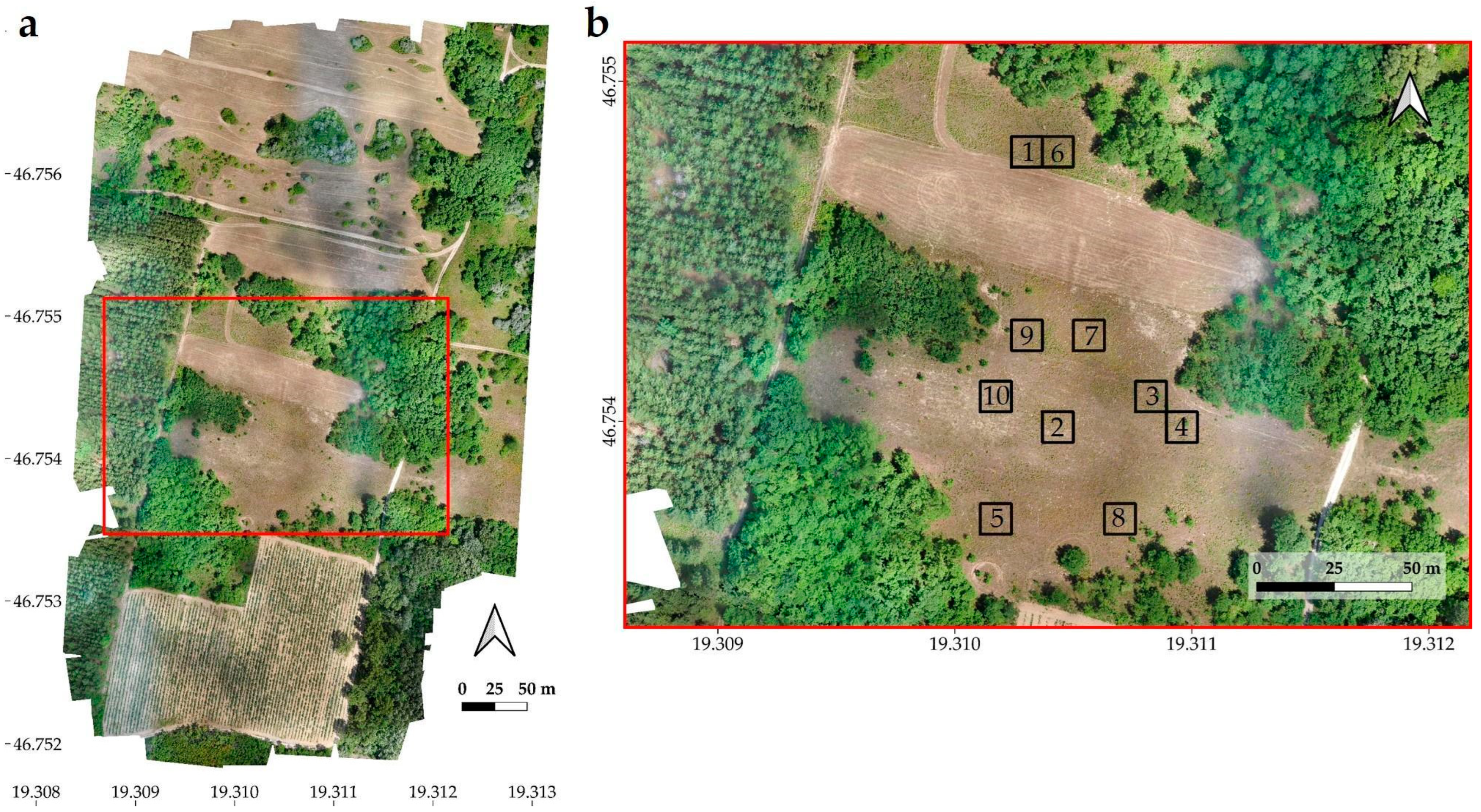
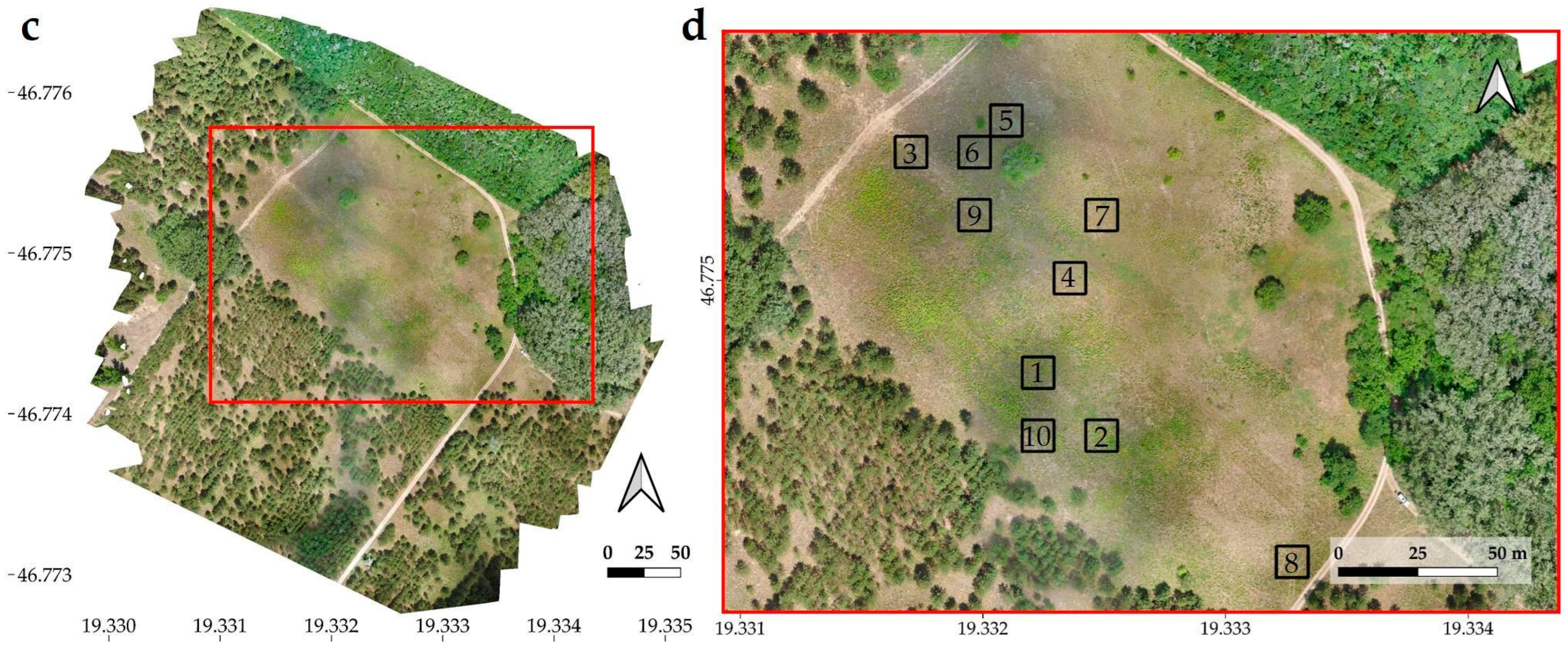
2.3. Documentation Methods
2.4. Data Processing Methods
2.4.1. The Used RGB Indices
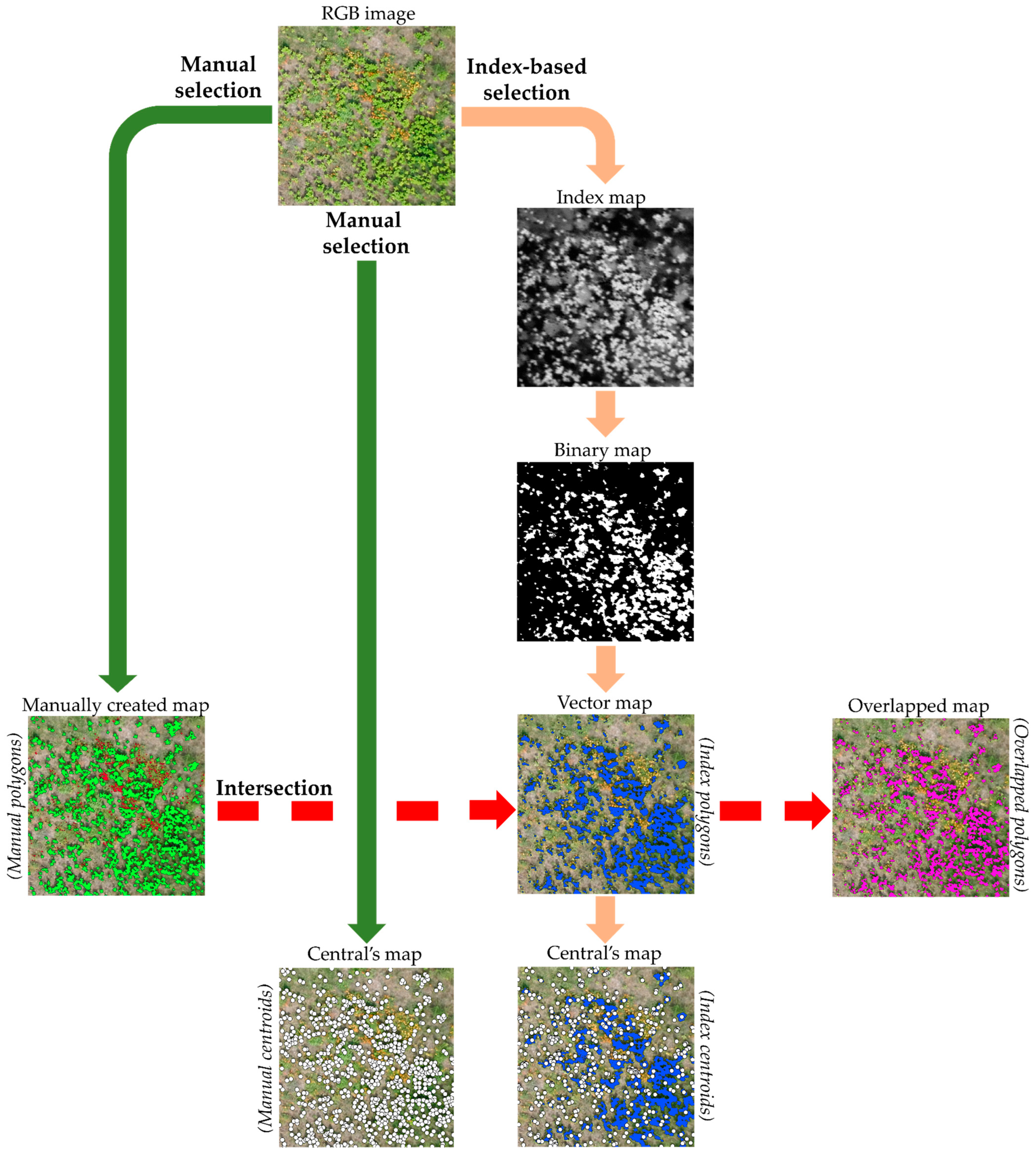
2.4.2. Data Validation Processes
2.5. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Richardson, D.M.; Pyšek, P.; Rejmánek, M.; Barbour, M.G.; Panetta, F.D.; West, C.J. Naturalization and Invasion of Alien Plants: Concepts and Definitions. Divers. Distrib. 2000, 6, 93–107. [Google Scholar] [CrossRef]
- Kettunen, M.; Genovesi, P.; Gollasch, S.; Pagad, S.; Starfinger, U.; ten Brink, P.; Shine, C. Technical Support to EU Strategy on Invasive Alien SPECIES (IAS); Institute for European Environmental Policy (IEEP): Brussels, Belgium, 2009; pp. 3–28. [Google Scholar]
- European Commission. Regulation (EU) No 1143/2014 of the European Parliament and of the Council 22 October 2014 on the Prevention and Management of the Introduction and Spread of Invasive Alien Species. Off. J. Eur. Union. 2014, L174, 511. Available online: https://www.eea.europa.eu/policy-documents/ec-2014-regulation-eu-no (accessed on 13 March 2023).
- Haubrock, P.J.; Turbelin, A.J.; Cuthbert, R.N.; Novoa, A.; Taylor, N.G.; Angulo, E.; Ballesteros-Mejia, L.; Bodey, T.W.; Capinha, C.; Diagne, C.; et al. Economic Costs of Invasive Alien Species Across Europe. NeoBiota 2021, 67, 153–190. [Google Scholar] [CrossRef]
- Schiffleithner, V.; Essl, F. Is it Worth the Effort? Spread and Management Success of Invasive Alien Plant Species in a Central European National Park. NeoBiota 2016, 31, 43–61. [Google Scholar] [CrossRef]
- Csiszár, Á.; Korda, M. Practical Experiences in Invasive Alien Plant Control; Duna-Ipoly Nemzeti Park Igazgatóság: Budapest, Hungary, 2015; pp. 17–25. [Google Scholar]
- Bakó, G. Az Özönnövények Feltérképezése a Beavatkozás Megtervezéséhez és Precíziós Kivitelezéséhez. In Practical Experiences in Invasive Alien Plant Control; Csiszár, Á., Korda, M., Eds.; Duna-Ipoly Nemzeti Park Igazgatóság: Budapest, Hungary, 2015; pp. 17–25. [Google Scholar]
- Bolch, E.A.; Santos, M.J.; Ade, C.; Khanna, S.; Basinger, N.T.; Reader, M.O.; Hestir, E.L. Remote Detection of Invasive Alien Species. In Remote Sensing of Plant Biodiversity; Cavender-Bares, J., Gamon, J.A., Townsend, P.A., Eds.; Springer Nature: Cham, Switzerland, 2020; pp. 267–307. [Google Scholar]
- Bakacsy, L. Invasion Impact is Conditioned by Initial Vegetation States. Commun. Ecol. 2019, 20, 11–19. [Google Scholar] [CrossRef]
- Cruzan, M.B.; Weinstein, B.G.; Grasty, M.R.; Kohrn, B.F.; Hendrickson, E.C.; Arredondo, T.M.; Thompson, P.G. Small Unmanned Aerial Vehicles (Micro-UAVs, Drones) in Plant Ecology. App. Plant Sci. 2016, 4, 1600041. [Google Scholar] [CrossRef] [PubMed]
- Dvořák, P.; Müllerová, J.; Bartaloš, T.; Brůna, J. 2015 Unmanned Aerial Vehicles for Alien Plant Species Detection and Monitoring. Int. Arch. Photogramm. Remote Sens. Spat. Inf. Sci. 2015, XL-1/W4, 83–90. [Google Scholar] [CrossRef]
- Michez, A.; Piégay, H.; Jonathan, L.; Claessens, H.; Lejeune, P. Mapping of Riparian Invasive Species with Supervised Classification of Unmanned Aerial System (UAS) Imagery. Int. J. Appl. Earth Obs. Geoinf. 2016, 44, 88–94. [Google Scholar] [CrossRef]
- Hill, D.J.; Tarasoff, C.; Whitworth, G.E.; Baron, J.; Bradshaw, J.L.; Church, J.S. Utility of Unmanned Aerial Vehicles for Mapping Invasive Plant Species: A Case Study on Yellow Flag Iris (Iris pseudacorus L.). Int. J. Remote Sens. 2017, 38, 2083–2105. [Google Scholar] [CrossRef]
- Müllerová, J.; Bartaloš, T.; Brůna, J.; Dvořák, P.; Vítková, M. Unmanned Aircraft in Nature Conservation: An Example from Plant Invasions. Int. J. Remote Sens. 2017, 38, 2177–2198. [Google Scholar] [CrossRef]
- Lehmann, J.R.; Prinz, T.; Ziller, S.R.; Thiele, J.; Heringer, G.; Meira-Neto, J.A.; Buttschardt, T.K. Open-Source Processing and Analysis of Aerial Imagery Acquired with a Low-Cost Unmanned Aerial System to Support Invasive Plant Management. Front. Environm. Sci. 2017, 5, 44. [Google Scholar] [CrossRef]
- Wijesingha, J.; Astor, T.; Schulze-Brüninghoff, D.; Wachendorf, M. Mapping Invasive Lupinus polyphyllus Lindl. in Semi-Natural Grasslands Using Object-Based Image Analysis of UAV-Borne Images. PFG J. Photogramm. Remote Sens. Geoinf. Sci. 2020, 88, 391–406. [Google Scholar] [CrossRef]
- Niphadkar, M.; Nagendra, H. Remote Sensing of Invasive Plants: Incorporating Functional Traits into the Picture. Int. J. Remote Sens. 2016, 37, 3074–3085. [Google Scholar] [CrossRef]
- de Sá, N.C.; Castro, P.; Carvalho, S.; Marchante, E.; López-Núñez, F.A.; Marchante, H. Mapping the Flowering of an Invasive Plant Using Unmanned Aerial Vehicles: Is There Potential for Biocontrol Monitoring? Front. Plant Sci. 2018, 9, 293. [Google Scholar] [PubMed]
- Botta-Dukát, Z. Invasion of Alien Species to Hungarian (Semi-) Natural Habitats. Acta Bot. Hung. 2008, 50, 219–227. [Google Scholar] [CrossRef]
- Molnár, Z.; Bölöni, J.; Horváth, F. Threatening Factors Encountered: Actual Endangerment of the Hungarian (Semi-) Natural Habitats. Acta Bot. Hung. 2008, 50, 199–217. [Google Scholar] [CrossRef]
- Martin, F.M.; Müllerová, J.; Borgniet, L.; Dommanget, F.; Breton, V.; Evette, A. Using Single- and Multi-Date UAV and Satellite Imagery to Accurately Monitor Invasive Knotweed Species. Remote Sens. 2018, 10, 1662. [Google Scholar] [CrossRef]
- Müllerová, J.; Brůna, J.; Bartaloš, T.; Dvořák, P.; Vítková, M.; Pyšek, P. Timing Is Important: Unmanned Aircraft vs. Satellite Imagery in Plant Invasion Monitoring. Front. Plant Sci. 2017, 8, 887. [Google Scholar] [CrossRef]
- Horning, N.; Fleishman, E.; Ersts, P.J.; Fogarty, F.A.; Wohlfeil Zillig, M. Mapping of Land Cover with Open-Source Software and Ultra-High-Resolution Imagery Acquired with Unmanned Aerial Vehicles. Remote Sens. Ecol. Conserv. 2020, 6, 487–497. [Google Scholar] [CrossRef]
- Sankey, T.T.; McVay, J.; Swetnam, T.L.; McClaran, M.P.; Heilman, P.; Nichols, M. UAV Hyperspectral and Lidar Data and Their Fusion for Arid and Semi-arid Land Vegetation Monitoring. Remote Sens. Ecol. Conserv. 2018, 4, 20–33. [Google Scholar] [CrossRef]
- Elkind, K.; Sankey, T.T.; Munson, S.M.; Aslan, C.E. Invasive Buffelgrass Detection Using High-resolution Satellite and UAV Imagery on Google Earth Engine. Remote Sens. Ecol. Conserv. 2019, 5, 318–331. [Google Scholar] [CrossRef]
- Narumalani, S.; Mishra, D.R.; Wilson, R.; Reece, P.; Kohler, A. Detecting and Mapping four Invasive Species Along the Floodplain of North Platte River, Nebraska. Weed Technol. 2009, 23, 99–107. [Google Scholar] [CrossRef]
- Lass, L.W.; Prather, T.S.; Glenn, N.F.; Weber, K.T.; Mundt, J.T.; Pettingill, J. A Review of Remote Sensing of Invasive Weeds and Example of the Early Detection of Spotted Knapweed (Centaurea maculosa) and Babysbreath (Gypsophila paniculata) with a Hyperspectral Sensor. Weed Sci. 2005, 53, 242–251. [Google Scholar] [CrossRef]
- Wan, H.; Wang, C.; Li, Y.; Wang, Q.; Li, J.; Liu, X. Monitoring an invasive plant using hyperspectral remote sensing data. Trans. Chin. Soc. Agric. Eng. 2010, 26, 59–63. [Google Scholar] [CrossRef]
- Skowronek, S.; Asner, G.P.; Feilhauer, H. Performance of One-class Classifiers for Invasive Species Mapping Using Airborne Imaging Spectroscopy. Ecol. Inform. 2017, 37, 66–76. [Google Scholar] [CrossRef]
- Paz-Kagan, T.; Silver, M.; Panov, N.; Karnieli, A. Multispectral Approach for Identifying Invasive Plant Species Based on Flowering Phenology Characteristics. Remote Sens. 2019, 11, 953. [Google Scholar] [CrossRef]
- Kattenborn, T.; Lopatin, J.; Förster, M.; Braun, A.C.; Fassnacht, F.E. UAV Data as Alternative to Field Sampling to Map Woody Invasive Species Based on Combined Sentinel-1 and Sentinel-2 Data. Remote Sens. Environ. 2019, 227, 61–73. [Google Scholar] [CrossRef]
- Lopatin, J.; Dolos, K.; Kattenborn, T.; Fassnacht, F.E. How Canopy Shadow Affects Invasive Plant Species Classification in High Spatial Resolution Remote Sensing. Remote Sens. Ecol. Conserv. 2019, 5, 302–317. [Google Scholar] [CrossRef]
- Kawashima, S.; Nakatani, M. An Algorithm for Estimating Chlorophyll Content in Leaves Using a Video Camera. Ann. Bot. 1998, 81, 49–54. [Google Scholar] [CrossRef]
- Hunt, E.R., Jr.; Doraiswamy, P.C.; McMurtrey, J.E.; Daughtry, C.S.; Perry, E.M.; Akhmedov, B. A Visible Band Index for Remote Sensing Leaf Chlorophyll Content at the Canopy Scale. Int. J. Appl. Earth Obs. Geoinf. 2013, 21, 103–112. [Google Scholar] [CrossRef]
- Escadafal, R.; Belghith, A.; Ben-Moussa, H. Indices Spectraux pour la Dégradation des Milieux Maturels en Tunisie Aride. In Proceedings of the 6ème Symp. Int. “Mesures Physiques et Signatures en Télédétection”, Val d’Isere, France, 17–21 January 1994; pp. 253–259. [Google Scholar]
- Chen, Y.; Su, W.; Li, J.; Sun, Z. Hierarchical Object Oriented Classification Using Very High Resolution Imagery and LIDAR Data Over Urban Areas. Adv. Sp. Res. 2009, 43, 1101–1110. [Google Scholar] [CrossRef]
- Smith, R.G.; Maxwell, B.D.; Menalled, F.D.; Rew, L.J. Lessons from Agriculture may Improve the Management of Invasive Plants in Wildland Systems. Front. Ecol. Environ. 2006, 4, 428–434. [Google Scholar] [CrossRef]
- Bagi, I. Common Milkweed (Asclepias syriaca L.). In The Most Important Invasive Plants in Hungary; Botta-Dukát, Z., Balogh, L., Eds.; Institute of Ecology and Botany, Hungarian Academy of Sciences: Vácrátót, Hungary, 2008; pp. 151–159. [Google Scholar]
- Follak, S.; Bakacsy, L.; Essl, F.; Hochfellner, L.; Lapin, K.; Schwarz, M.; Tokarska-Guzik, B.; Wołkowycki, D. Monograph of Invasive Plants in Europe N° 6: Asclepias syriaca L. Bot. Lett. 2021, 168, 422–451. [Google Scholar] [CrossRef]
- Commonwealth Agricultural Bureau International (CABI). Asclepias Syriaca (Common Milkweed). Available online: http://www.cabi.org/isc/datasheet/7249 (accessed on 5 April 2015).
- European Commission. List of Invasive Alien Species of Union Concern. 2017. Available online: http://ec.europa.eu/environment/nature/invasivealien/list/index_en.htm (accessed on 13 March 2023).
- The European and Mediterranean Plant Protection Organization (EPPO). EPPO Global Database. 2019. Available online: https://gd.eppo.int/taxon/ASCSY (accessed on 13 March 2023).
- Szatmári, J.; Tobak, Z.; Novák, Z. Environmental Monitoring Supported by Aerial Photography—A Case Study of the Burnt Down Bugac Juniper Forest, Hungary. J. Environ. Geogr. 2016, 9, 31–38. [Google Scholar] [CrossRef]
- Bagi, I.; Bakacsy, L. Közönséges selyemkóró (Asclepias syriaca). In Inváziós növényfajok Magyarországon; Csiszár, Á., Eds.; Nyugat-magyarországi Egyetem Kiadó: Sopron, Hungary, 2012; pp. 183–187.syriaca). In Inváziós növényfajok Magyarországon; Csiszár, Á., Ed.; Nyugat-magyarországi Egyetem Kiadó: Sopron, Hungary, 2012; Nyugat-magyarországi Egyetem Kiadó: Sopron, Hungary, 2012; pp. 183–187. [Google Scholar]
- Szilassi, P.; Szatmári, G.; Pásztor, L.; Árvai, M.; Szatmári, J.; Szitár, K.; Papp, L. Understanding the Environmental Background of an Invasive Plant Species (Asclepias syriaca) for the Future: An Application of LUCAS Field Photographs and Machine Learning Algorithm Methods. Plants 2019, 8, 593. [Google Scholar] [CrossRef]
- Szilassi, P.; Soóky, A.; Bátori, Z.; Hábenczyus, A.A.; Frei, K.; Tölgyesi, C.; Van Leeuwen, B.; Tobak, Z.; Csikós, N. Natura 2000 Areas, Road, Railway, Water, and Ecological Networks May Provide Pathways for Biological Invasion: A Country Scale Analysis. Plants 2021, 10, 2670. [Google Scholar] [CrossRef]
- Stoutamire, W. Chromosome Races of Gaillardia pulchella (Asteraceae). Brittonia 1977, 29, 297–309. [Google Scholar] [CrossRef]
- Simon, T. A Magyarországi Edényes Flóra Határozója (Vascular Flora of Hungary); Nemzeti Tankönyvkiadó: Budapest, Hungary, 2000; p. 976. [Google Scholar]
- Molnár, C.; Csathó, A.I.; Molnár, Á.P.; Pifkó, D. Amendments to the Alien Flora of the Republic of Moldova. Studia Bot. Hung. 2019, 50, 225–240. [Google Scholar] [CrossRef]
- Daehler, C.C. The Taxonomic Distribution of Invasive Angiosperm Plants: Ecological Insights and Comparison to Agricultural Weeds. Biol. Conserv. 1998, 84, 167–180. [Google Scholar] [CrossRef]
- Pyšek, P. Is There a Taxonomic Pattern to Plant Invasions? Oikos 1998, 82, 282–294. [Google Scholar] [CrossRef]
- Tóth, K. 20 éves a Kiskunsági Nemzeti Park 1975-1995; Kiskunság Nemzeti Park Igazgatósága: Kecskemét, Hungary, 1996; p. 234. [Google Scholar]
- Sipos, G.; Tóth, O.; Pécsi, E.; Bíró, C. Bracketing the Age of Freshwater Carbonate Formation by OSL Dating Near Lake Kolon, Hungary. J. Environ. Geogr. 2014, 7, 53–59. [Google Scholar] [CrossRef]
- Bölöni, J.; Molnár, Z.; Kun, A. Magyarország élőhelyei: Vegetációtipusok Leirása és Határozója: ÁNÉR 2011; MTA Ökológiai és Botanikai Kutatóintézete: Vácrátót, Hungary, 2011; p. 439. [Google Scholar]
- QGIS Development Team. QGIS Geographic Information System. QGIS Association. 2021. Available online: https://www.qgis.org (accessed on 17 June 2021).
- eBee X mapping drone—Drones. Available online: https://ageagle.com/drones/ebee-x/ (accessed on 15 December 2022).
- S.O.D.A.—Sensors. Available online: https://ageagle.com/dronessensors/soda/ (accessed on 15 December 2022).
- Agisoft, L.L.C. Agisoft Metashape User Manual Professional Edition, Version 1.5.4.; Agisoft LLC.: St. Petersburg, Russia, 2020. [Google Scholar]
- Jacquemoud, S.; Verhoef, W.; Baret, F.; Bacour, C.; Zarco-Tejada, P.J.; Asner, G.P.; François, C.; Ustin, S.L. PROSPECT + SAIL Models: A Review of Use for Vegetation Characterization. Remote Sens. Environ. 2009, 113, S56–S66. [Google Scholar] [CrossRef]
- Feret, J.B.; François, C.; Asner, G.P.; Gitelson, A.A.; Martin, R.E.; Bidel, L.P.R.; Ustin, S.L.; le Maire, G.; Jacquemoud, S. PROSPECT-4 and 5: Advances in the Leaf Optical Properties Model Separating Photosynthetic Pigments. Remote Sens. Environ. 2008, 112, 3030–3043. [Google Scholar] [CrossRef]
- Bannari, A.; El-Harti, A.; Haboudane, D.; Bachaoui, E.M.; El-Ghmari, A. Intégration des Variables Spectrales et Géomorphométriques dans un SIG pour la Cartographie des Aires Exposées á L’Érosion (Integration of Spectral and Geomorphometric Variables in a GIS for Erosion Risk Mapping). Revue Télédétection. 2007, 7, 327–342. [Google Scholar]
- Congalton, R.G. A Review of Assessing the Accuracy of Classifications of Remotely Sensed Data. Remote Sens. Environ. 1991, 37, 35–46. [Google Scholar] [CrossRef]
- Papp, L.; Van Leeuwen, B.; Szilassi, P.; Tobak, Z.; Szatmári, J.; Árvai, M.; Mészáros, J.; Pásztor, L. Monitoring Invasive Plant Species Using Hyperspectral Remote Sensing Data. Land 2021, 10, 29. [Google Scholar] [CrossRef]
- Ozcan, K.; Sharma, A.; Bradbury, S.P.; Schweitzer, D.; Blader, T.; Blodgett, S. Milkweed (Asclepias syriaca) Plant Detection Using Mobile Cameras. Ecosphere 2020, 11, e02992. [Google Scholar] [CrossRef]
- Bakó, G.; (Interspect Ltd., II. Rákóczi Ferenc út 42, H-2314 Halásztelek, Hungary). Personal communication, 2021.
- Kunah, O.M.; Papka, O.S. Ecogeographical Determinants of the Ecological Niche of the Common Milkweed (Asclepias syriaca) on the Basis of indices of Remote Sensing of Land Images. Biosyst. Divers. 2016, 24, 78–86. [Google Scholar] [CrossRef]
- Csecserits, A.; Halassy, M.; Rédei, T.; Szitár, K.; Botta-Dukát, Z. Changes in Abundance of Common Milkweed (Asclepias syriaca L.) on Sandy Old-Fields During Succession and Due to Conservation Management. Természetvédelmi Közlemények. 2020, 26, 1–15. [Google Scholar] [CrossRef]
- Csecserits, A.; Berki, B.; Botta-Dukát, Z.; Csákvári, E.; Halassy, M.; Mártonffy, A.; Rédei, T.; Szitár, K. Has the Vegetation and Severity of Invasion Changed in Sandy Grasslands and Old-Fields of the Kiskunság in the Last Decade?—Results of a Repeated Survey. Természetvédelmi Közlemények. 2022, 28, 13–28. [Google Scholar] [CrossRef]
- Gröschler, K.C.; Oppelt, N. Using Drones to Monitor Broad-Leaved Orchids (Dactylorhiza majalis) in High-Nature-Value Grassland. Drones 2022, 6, 174. [Google Scholar] [CrossRef]
- Carl, C.; Landgraf, D.; van der Maaten-Theunissen, M.; Biber, P.; Pretzsch, H. Robinia pseudoacacia L. Flowers Analyzed by Using an Unmanned Aerial Vehicle (UAV). Remote Sensing. 2017, 9, 1091. [Google Scholar] [CrossRef]
- Bakó, G. Vegetációtérképezés Nagyfelbontású Valósszínes-és Multispektrális Légifelvételek Alapján. [Vegetation Mapping Based on High-Resolution True Color and Multispectral Aerial Images]. Kitaibelia 2013, 18, 152–160, (in Hungarian with English abstract). [Google Scholar]
- Schulte, A.J.; Mail, M.; Hahn, L.A.; Barthlott, W. Ultraviolet Patterns of Flowers Revealed in Polymer Replica–Caused by Surface Architecture. Beilstein J. Nanotechnol. 2019, 10, 459–466. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Applicability to Species | Milkweed | Blanket Flower | Blanket Flower | Milkweed | Milkweed | Blanket Flower | Milkweed | |
|---|---|---|---|---|---|---|---|---|
| Applied Thresholds | R-G | R-G | R-B | G-B | TGI | IF | SSI | |
| STAND I. | 1.study area | <−1 | >38 | >99 | >71 | >4050 | >130 | >67 |
| 2.study area | <5 | >37 | >100 | >64 | >3500 | >119 | >55 | |
| 3.study area | <−3 | >40 | >102 | >71 | >4250 | >135 | >72 | |
| 4.study area | <−5 | >35 | >87 | >68 | >4250 | >113 | >74 | |
| 5.study area | <−5 | >42 | >98 | >72 | >4150 | >135 | >70 | |
| 6.study area | <−1 | >38 | >108 | >70 | >4050 | >137 | >67 | |
| 7.study area | <−3 | >37 | >96 | >79 | >4250 | >129 | >69 | |
| 8.study area | <−3 | >38 | >92 | >71 | >4200 | >119 | >71 | |
| 9.study area | <−1 | >38 | >98 | >71 | >4050 | >128 | >68 | |
| 10.study area | <2 | >34 | >89 | >64 | >3550 | >111 | >58 | |
| STAND II. | 1.study area | <−17 | >27 | >97 | >68 | >4500 | >116 | >82 |
| 2.study area | <−22 | >25 | >98 | >78 | >5300 | >116 | >97 | |
| 3.study area | <−5 | >29 | >93 | >57 | >3500 | >110 | >60 | |
| 4.study area | <−7 | >30 | >94 | >72 | >4350 | >116 | >75 | |
| 5.study area | <−23 | >21 | >72 | >53 | >3500 | >88 | >65 | |
| 6.study area | <−17 | >23 | >78 | >52 | >3500 | >92 | >65 | |
| 7.study area | <−10 | >33 | >89 | >73 | >4550 | >116 | >79 | |
| 8.study area | <−11 | >38 | >101 | >73 | >4600 | >132 | >81 | |
| 9.study area | <−12 | >29 | >89 | >60 | >3850 | >110 | >68 | |
| 10.study area | <−12 | >24 | >86 | >57 | >3800 | >92 | >68 | |
| Unpaired t Test | Stand I. vs. Stand II. | ||||||
|---|---|---|---|---|---|---|---|
| R-G (Milkweed) | R-G (Blanket Flower) | R-B (Blanket Flower) | G-B (Milkweed) | TGI (Milkweed) | IF (Blanket Flower) | SSI (Milkweed) | |
| p value | <0.0001 | <0.0001 | 0.0537 | 0.096 | 0.5934 | 0.0054 | 0.1002 |
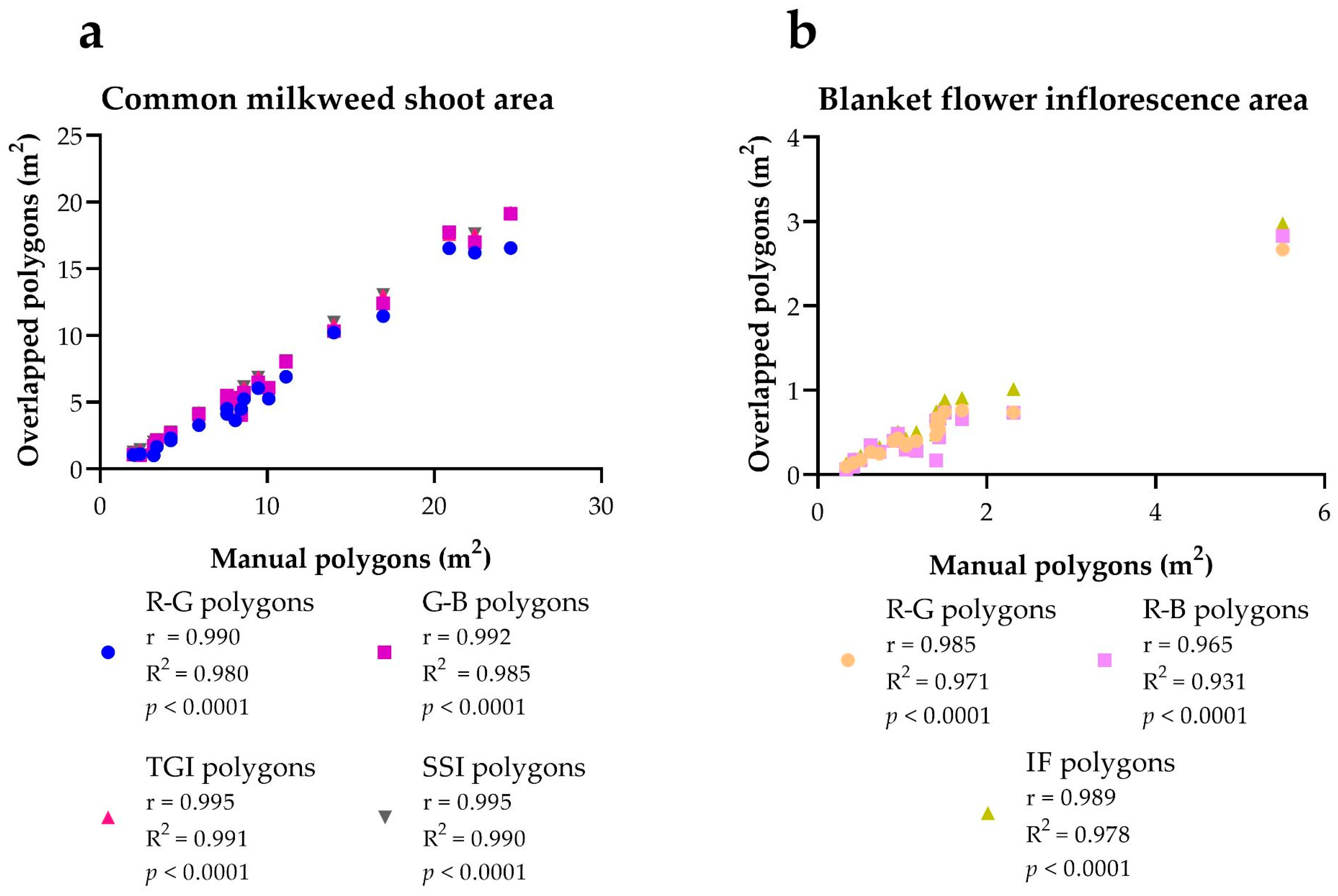
| Milkweed SHOOT AREA | Manual Polygons | R-G Overlapped Polygons | G-B Overlapped Polygons | TGI Overlapped Polygons | SSI Overlapped Polygons | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| m2 | m2 | % | m2 | % | m2 | % | m2 | % | ||
| STAND I. | 1.study area | 8.601 | 5.235 | 60.86 | 5.719 | 66.49 | 6.077 | 70.65 | 6.139 | 71.37 |
| 2.study area | 7.594 | 4.109 | 54.11 | 5.495 | 72.35 | 5.530 | 72.82 | 5.424 | 71.42 | |
| 3.study area | 8.440 | 4.496 | 53.23 | 4.026 | 47.66 | 5.362 | 63.48 | 5.341 | 63.24 | |
| 4.study area | 10.098 | 5.259 | 52.08 | 6.078 | 60.19 | 6.108 | 60.48 | 5.986 | 59.28 | |
| 5.study area | 1.984 | 1.049 | 52.87 | 1.082 | 54.54 | 1.232 | 62.11 | 1.247 | 62.86 | |
| 6.study area | 16.944 | 11.450 | 67.57 | 12.415 | 73.26 | 12.991 | 76.66 | 13.028 | 76.88 | |
| 7.study area | 2.393 | 1.113 | 46.52 | 1.022 | 42.72 | 1.361 | 56.86 | 1.423 | 59.44 | |
| 8.study area | 7.596 | 4.517 | 59.47 | 4.909 | 64.63 | 5.126 | 67.49 | 5.138 | 67.64 | |
| 9.study area | 9.456 | 6.046 | 63.93 | 6.482 | 68.55 | 6.894 | 72.90 | 6.849 | 72.42 | |
| 10.study area | 4.232 | 2.305 | 54.48 | 2.672 | 63.16 | 2.774 | 65.56 | 2.762 | 65.27 | |
| STAND II. | 1.study area | 22.422 | 16.198 | 72.24 | 16.991 | 75.77 | 17.572 | 78.37 | 17.593 | 78.46 |
| 2.study area | 24.573 | 16.550 | 67.35 | 19.116 | 77.79 | 19.210 | 78.17 | 19.101 | 77.72 | |
| 3.study area | 11.129 | 6.916 | 62.14 | 8.034 | 72.19 | 8.110 | 72.87 | 8.102 | 72.80 | |
| 4.study area | 4.222 | 2.140 | 50.68 | 2.409 | 57.06 | 2.584 | 61.20 | 2.580 | 61.11 | |
| 5.study area | 3.200 | 1.002 | 31.33 | 1.717 | 53.65 | 1.980 | 61.88 | 1.994 | 62.30 | |
| 6.study area | 5.904 | 3.279 | 55.55 | 4.087 | 69.23 | 4.215 | 71.40 | 4.163 | 70.51 | |
| 7.study area | 3.389 | 1.649 | 48.66 | 2.138 | 63.09 | 2.196 | 64.79 | 2.184 | 64.44 | |
| 8.study area | 8.088 | 3.631 | 44.89 | 5.331 | 65.91 | 5.147 | 63.64 | 4.960 | 61.32 | |
| 9.study area | 13.992 | 10.224 | 73.07 | 10.324 | 73.78 | 10.779 | 77.03 | 10.961 | 78.33 | |
| 10.study area | 20.889 | 16.531 | 79.13 | 17.730 | 84.88 | 17.588 | 84.20 | 17.582 | 84.17 | |
| Average % | 57.51 | 65.34 | 69.13 | 69.05 | ||||||
| SD % | 11.20 | 10.47 | 7.42 | 7.41 | ||||||
| Blanket Flower Inflorescence Area | Manual Polygons | R-G overlapped Polygons | R-B Overlapped Polygons | IF Overlapped Polygons | ||||
|---|---|---|---|---|---|---|---|---|
| m2 | m2 | % | m2 | % | m2 | % | ||
| STAND I. | 1.study area | 1.406 | 0.580 | 41.25 | 0.641 | 45.58 | 0.753 | 53.54 |
| 2.study area | 1.402 | 0.464 | 33.11 | 0.168 | 12.01 | 0.479 | 34.22 | |
| 3.study area | 0.899 | 0.397 | 44.16 | 0.402 | 44.77 | 0.476 | 52.91 | |
| 4.study area | 1.439 | 0.533 | 37.03 | 0.442 | 30.74 | 0.677 | 47.08 | |
| 5.study area | 0.949 | 0.427 | 45.06 | 0.485 | 51.12 | 0.512 | 53.92 | |
| 6.study area | 1.445 | 0.64 | 44.27 | 0.664 | 45.92 | 0.774 | 53.56 | |
| 7.study area | 5.505 | 2.667 | 48.44 | 2.827 | 51.37 | 2.974 | 54.02 | |
| 8.study area | 2.316 | 0.737 | 31.83 | 0.735 | 31.75 | 1.015 | 43.81 | |
| 9.study area | 1.706 | 0.761 | 44.59 | 0.657 | 38.52 | 0.908 | 53.22 | |
| 10.study area | 1.169 | 0.398 | 34.09 | 0.280 | 23.95 | 0.504 | 43.12 | |
| STAND II. | 1.study area | 1.506 | 0.746 | 49.55 | 0.734 | 48.78 | 0.881 | 58.54 |
| 2.study area | 1.413 | 0.67 | 47.43 | 0.639 | 45.28 | 0.747 | 52.85 | |
| 3.study area | 0.421 | 0.137 | 32.66 | 0.093 | 22.16 | 0.156 | 37.09 | |
| 4.study area | 1.043 | 0.365 | 35.05 | 0.339 | 32.52 | 0.431 | 41.33 | |
| 5.study area | 0.629 | 0.268 | 42.58 | 0.347 | 55.15 | 0.347 | 55.11 | |
| 6.study area | 0.509 | 0.172 | 33.75 | 0.172 | 33.86 | 0.221 | 43.34 | |
| 7.study area | 1.046 | 0.34 | 32.50 | 0.295 | 28.22 | 0.368 | 35.20 | |
| 8.study area | 0.436 | 0.129 | 29.62 | 0.174 | 39.94 | 0.187 | 42.96 | |
| 9.study area | 0.731 | 0.248 | 33.94 | 0.270 | 36.94 | 0.327 | 44.78 | |
| 10.study area | 0.336 | 0.096 | 28.53 | 0.060 | 18.05 | 0.132 | 39.18 | |
| Average % | 38.47 | 36.83 | 46.99 | |||||
| SD % | 6.76 | 11.95 | 7.41 | |||||
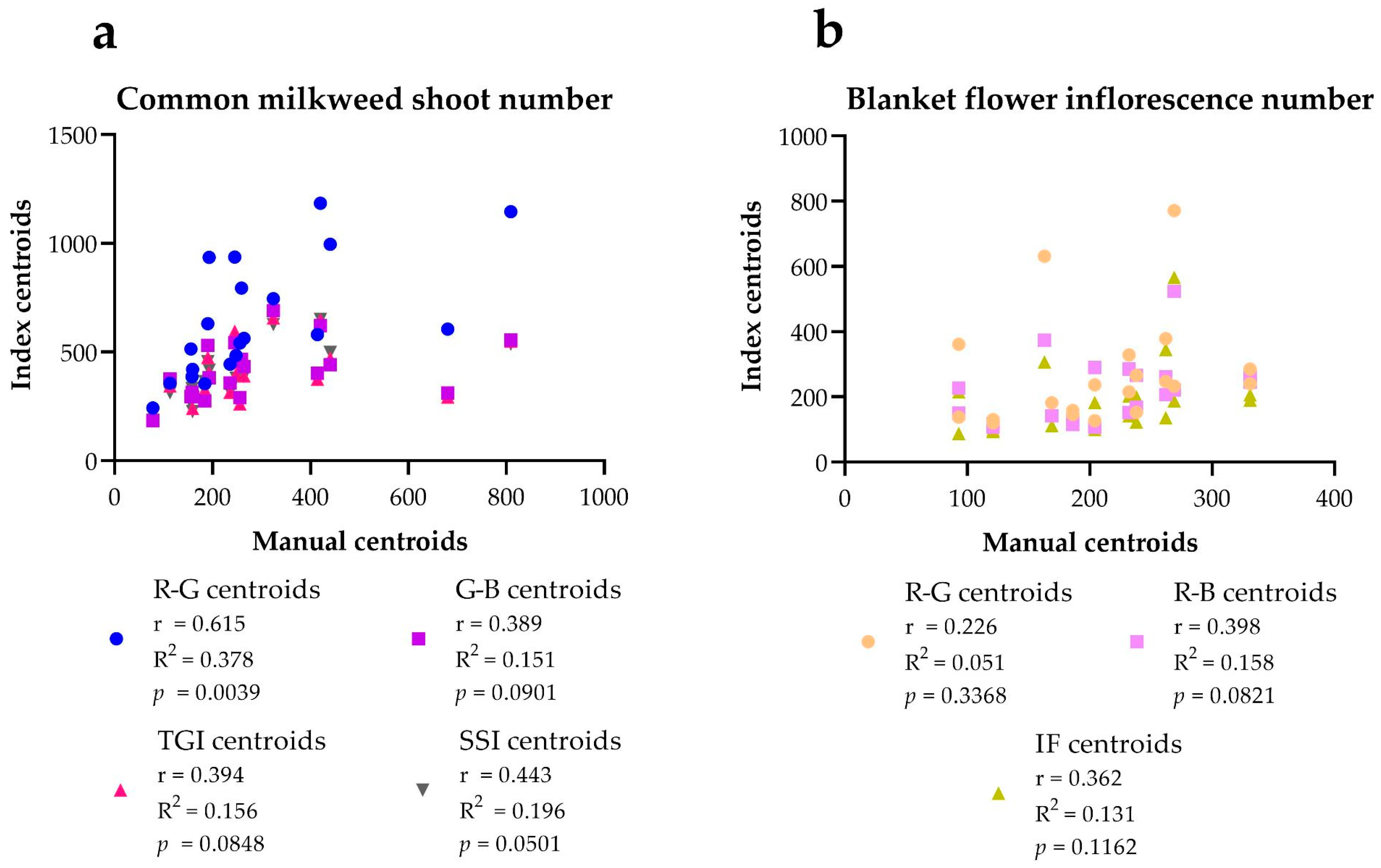
| Milkweed Shoot Number | Manual Centroids | R-G Centroids | G-B Centroids | TGI Centroids | SSI Centroids | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Number | Number | % | Number | % | Number | % | Number | % | ||
| STAND I. | 1.study area | 259 | 794 | 306.56 | 467 | 180.3 | 437 | 168.72 | 414 | 159.84 |
| 2.study area | 193 | 936 | 484.97 | 382 | 197.92 | 404 | 209.32 | 417 | 216.06 | |
| 3.study area | 236 | 444 | 188.13 | 358 | 151.69 | 315 | 133.47 | 315 | 133.47 | |
| 4.study area | 248 | 484 | 195.16 | 458 | 184.67 | 399 | 160.88 | 380 | 153.22 | |
| 5.study area | 78 | 243 | 311.53 | 184 | 235.89 | 204 | 261.53 | 188 | 241.02 | |
| 6.study area | 420 | 1184 | 281.9 | 622 | 148.09 | 646 | 153.8 | 650 | 154.76 | |
| 7.study area | 113 | 357 | 315.92 | 377 | 333.62 | 344 | 304.42 | 313 | 276.99 | |
| 8.study area | 190 | 630 | 331.57 | 531 | 279.47 | 475 | 250 | 455 | 239.47 | |
| 9.study area | 264 | 563 | 213.25 | 433 | 164.01 | 390 | 147.72 | 388 | 146.96 | |
| 10.study area | 156 | 514 | 329.48 | 296 | 189.74 | 298 | 191.02 | 264 | 169.23 | |
| STAND II. | 1.study area | 245 | 937 | 382.44 | 543 | 221.63 | 596 | 243.26 | 532 | 217.14 |
| 2.study area | 809 | 1145 | 141.53 | 555 | 68.6 | 551 | 68.1 | 537 | 66.37 | |
| 3.study area | 440 | 995 | 226.13 | 442 | 100.45 | 470 | 106.81 | 498 | 113.18 | |
| 4.study area | 159 | 420 | 264.15 | 294 | 184.9 | 240 | 150.94 | 227 | 142.76 | |
| 5.study area | 184 | 354 | 192.39 | 276 | 150 | 328 | 178.26 | 364 | 197.82 | |
| 6.study area | 256 | 542 | 211.71 | 290 | 113.28 | 262 | 102.34 | 281 | 109.76 | |
| 7.study area | 158 | 387 | 244.93 | 316 | 200 | 310 | 196.2 | 332 | 210.12 | |
| 8.study area | 324 | 746 | 230.24 | 690 | 212.96 | 657 | 202.77 | 627 | 193.51 | |
| 9.study area | 414 | 580 | 140.09 | 403 | 97.34 | 375 | 90.57 | 373 | 90.09 | |
| 10.study area | 680 | 606 | 89.11 | 312 | 45.88 | 293 | 43.08 | 304 | 44.7 | |
| Average % | 254.06 | 173.02 | 168.16 | 163.82 | ||||||
| SD % | 92.23 | 68.94 | 67.22 | 61.01 | ||||||
| Blanket Flower Inflorescence Number | Manual Centroids | R-G Centroids | R-B Centroids | IF Centroids | ||||
|---|---|---|---|---|---|---|---|---|
| Number | Number | % | Number | % | Number | % | ||
| STAND I. | 1.study area | 331 | 285 | 86.10 | 245 | 74.01 | 190 | 57.40 |
| 2.study area | 262 | 379 | 144.65 | 262 | 100.00 | 345 | 131.67 | |
| 3.study area | 121 | 130 | 107.43 | 107 | 88.42 | 94 | 77.68 | |
| 4.study area | 232 | 329 | 141.81 | 285 | 122.84 | 203 | 87.50 | |
| 5.study area | 186 | 159 | 85.48 | 131 | 70.43 | 125 | 67.20 | |
| 6.study area | 204 | 237 | 116.17 | 290 | 142.15 | 183 | 89.70 | |
| 7.study area | 269 | 771 | 286.61 | 524 | 194.79 | 566 | 210.40 | |
| 8.study area | 163 | 631 | 387.11 | 374 | 229.44 | 307 | 188.34 | |
| 9.study area | 238 | 266 | 111.76 | 266 | 111.76 | 200 | 84.03 | |
| 10.study area | 93 | 361 | 388.17 | 227 | 244.08 | 215 | 231.18 | |
| STAND II. | 1.study area | 331 | 240 | 72.50 | 262 | 79.15 | 206 | 62.23 |
| 2.study area | 262 | 247 | 94.27 | 207 | 79.00 | 136 | 51.90 | |
| 3.study area | 121 | 120 | 99.17 | 118 | 97.52 | 98 | 80.99 | |
| 4.study area | 232 | 215 | 92.67 | 152 | 65.51 | 142 | 61.20 | |
| 5.study area | 186 | 146 | 78.49 | 115 | 61.82 | 120 | 64.51 | |
| 6.study area | 204 | 127 | 62.25 | 107 | 52.45 | 100 | 49.01 | |
| 7.study area | 269 | 233 | 86.61 | 221 | 82.15 | 188 | 69.88 | |
| 8.study area | 169 | 182 | 107.69 | 142 | 84.02 | 112 | 66.27 | |
| 9.study area | 238 | 153 | 64.28 | 169 | 71.00 | 123 | 51.68 | |
| 10.study area | 93 | 138 | 148.38 | 150 | 161.29 | 88 | 94.62 | |
| Average % | 138.08 | 110.59 | 93.87 | |||||
| SD % | 98.00 | 55.88 | 53.91 | |||||
| R-G | G-B | TGI | SSI | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Producer’s acc. % | User’s acc. % | Kappa co. | Producer’s acc. % | User’s acc. % | Kappa co. | Producer’s acc. % | User’s acc. % | Kappa co. | Producer’s acc. % | User’s acc. % | Kappa co. | |
| STAND I. | 51.51 | 63.53 | 0.57 | 57.45 | 71.66 | 0.63 | 59.62 | 74.31 | 0.66 | 59.44 | 74.27 | 0.66 |
| STAND II. | 65.67 | 66.67 | 0.66 | 73.79 | 74.67 | 0.74 | 75.14 | 76.52 | 0.76 | 74.96 | 75.97 | 0.75 |
| Average | 58.59 | 65.10 | 0.62 | 65.62 | 73.17 | 0.69 | 67.38 | 75.42 | 0.71 | 67.20 | 75.12 | 0.71 |
| SD | 10.01 | 2.22 | 0.06 | 11.56 | 2.13 | 0.08 | 10.98 | 1.56 | 0.07 | 10.97 | 1.20 | 0.06 |
| R-G | R-B | IF | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Producer’s acc. % | User’s acc. % | Kappa co. | Producer’s acc. % | User’s acc. % | Kappa co. | Producer’s acc. % | User’s acc. % | Kappa co. | |
| STAND I. | 34.54 | 47.96 | 0.40 | 45.90 | 45.90 | 0.39 | 41.02 | 56.02 | 0.47 |
| STAND II. | 39.06 | 39.44 | 0.39 | 39.35 | 39.35 | 0.39 | 46.46 | 46.77 | 0.47 |
| Average | 36.80 | 43.70 | 0.40 | 42.62 | 42.62 | 0.39 | 43.74 | 51.40 | 0.47 |
| SD | 3.20 | 6.02 | 0.01 | 4.63 | 4.63 | 0.00 | 3.85 | 6.54 | 0.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bakacsy, L.; Tobak, Z.; van Leeuwen, B.; Szilassi, P.; Biró, C.; Szatmári, J. Drone-Based Identification and Monitoring of Two Invasive Alien Plant Species in Open Sand Grasslands by Six RGB Vegetation Indices. Drones 2023, 7, 207. https://doi.org/10.3390/drones7030207
Bakacsy L, Tobak Z, van Leeuwen B, Szilassi P, Biró C, Szatmári J. Drone-Based Identification and Monitoring of Two Invasive Alien Plant Species in Open Sand Grasslands by Six RGB Vegetation Indices. Drones. 2023; 7(3):207. https://doi.org/10.3390/drones7030207
Chicago/Turabian StyleBakacsy, László, Zalán Tobak, Boudewijn van Leeuwen, Péter Szilassi, Csaba Biró, and József Szatmári. 2023. "Drone-Based Identification and Monitoring of Two Invasive Alien Plant Species in Open Sand Grasslands by Six RGB Vegetation Indices" Drones 7, no. 3: 207. https://doi.org/10.3390/drones7030207
APA StyleBakacsy, L., Tobak, Z., van Leeuwen, B., Szilassi, P., Biró, C., & Szatmári, J. (2023). Drone-Based Identification and Monitoring of Two Invasive Alien Plant Species in Open Sand Grasslands by Six RGB Vegetation Indices. Drones, 7(3), 207. https://doi.org/10.3390/drones7030207