Validation of an In-Vitro Parkinson’s Disease Model for the Study of Neuroprotection †


{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Transient Transfection
2.2. XTT Assay
2.3. qPCR
2.4. Western Blotting
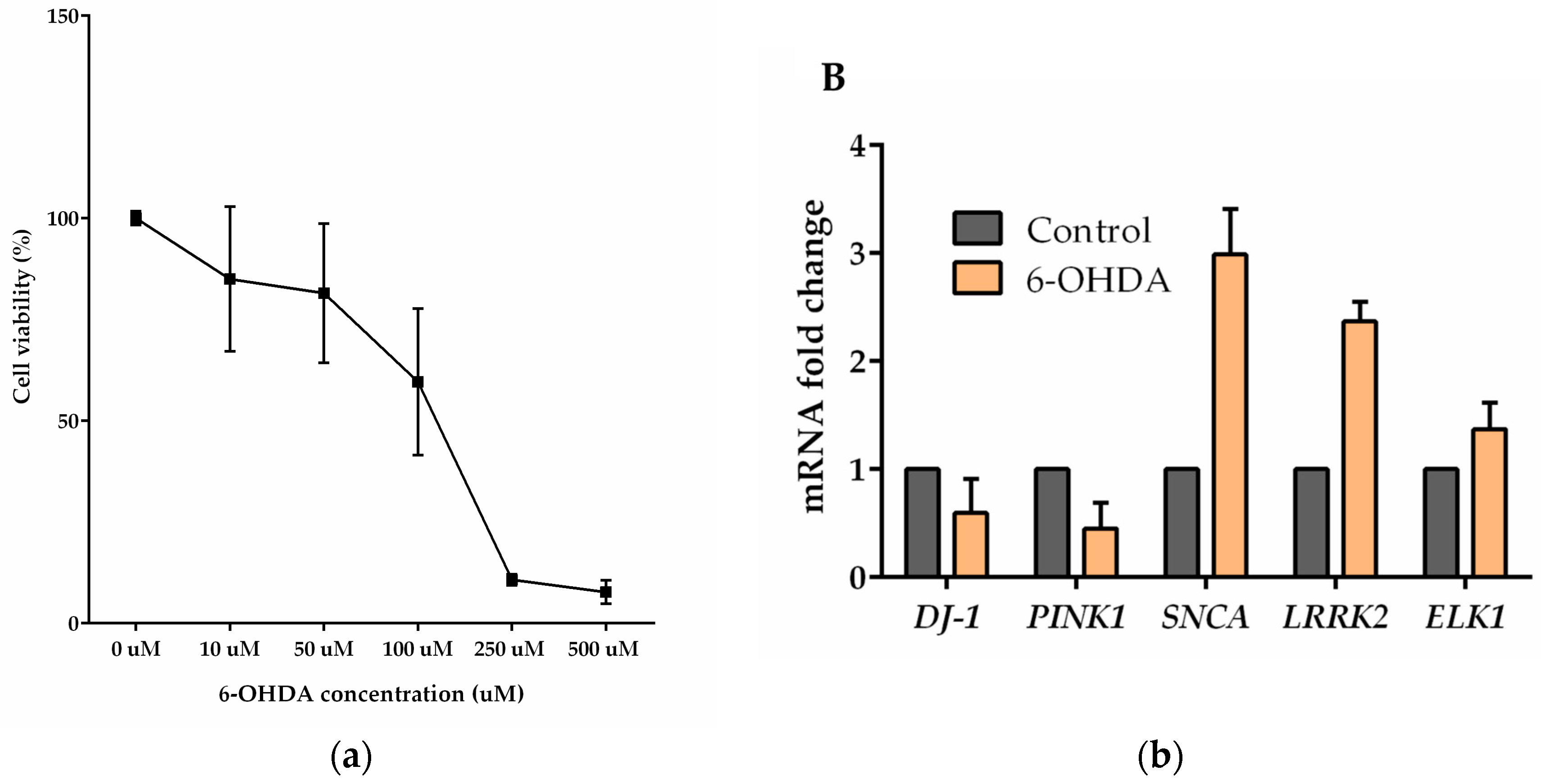
3. Results
4. Discussion
References
- Klein, C.; Westenberger, A. Genetics of Parkinson’s Disease. Cold Spring Harb. Perspect. Med. 2012, 2, a008888. [Google Scholar] [CrossRef] [PubMed]
- Singleton, A.B.; Farrer, M.; Johnson, J.; Singleton, A.; Hague, S.; Kachergus, J.; Hulihan, M.; Peuralinna, T.; Dutra, A.; Nussbaum, R.; et al. alpha-Synuclein locus triplication causes Parkinson’s disease. Science 2003, 302, 841. [Google Scholar] [CrossRef] [PubMed]
- Bové, J.; Prou, D.; Perier, C.; Przedborski, S. Toxin-induced models of Parkinson’s disease. NeuroRx 2005, 2, 484–494. [Google Scholar] [CrossRef] [PubMed]
- Junn, E.; Mouradian, M.M. Human alpha-synuclein over-expression increases intracellular reactive oxygen species levels and susceptibility to dopamine. Neurosci. Lett. 2002, 320, 146–150. [Google Scholar] [CrossRef]
- Koukouraki, P.; Doxakis, E. Constitutive translation of human alpha-synuclein is mediated by the 5′-untranslated region. Open Biol. 2016, 6, 160022. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.E.; Mouradian, M.M. Cytoprotective mechanisms of DJ-1 against oxidative stress through modulating ERK1/2 and ASK1 signal transduction. Redox Biol. 2018, 14, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Iwata, A.; Miura, S.; Kanazawa, I.; Sawada, M.; Nukina, N. α-Synuclein forms a complex with transcription factor Elk-1. J. Neurochem. 2001, 77, 239–252. [Google Scholar] [CrossRef] [PubMed]
- Demir, O.; Aysit, N.; Onder, Z.; Turkel, N.; Ozturk, G.; Sharrocks, A.D.; Kurnaz, I.A. ETS-domain Transcription Factor Elk-1 Mediates Neuronal Survival: SMN as A Potential Target. Biochim. Biophys. Acta BBA Mol. Basis Dis. 2011, 1812, 652–662. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yiğit, E.N.; Sönmez, E.; Söğüt, M.S.; Çakır, T.; Kurnaz, I.A. Validation of an In-Vitro Parkinson’s Disease Model for the Study of Neuroprotection. Proceedings 2018, 2, 1559. https://doi.org/10.3390/proceedings2251559
Yiğit EN, Sönmez E, Söğüt MS, Çakır T, Kurnaz IA. Validation of an In-Vitro Parkinson’s Disease Model for the Study of Neuroprotection. Proceedings. 2018; 2(25):1559. https://doi.org/10.3390/proceedings2251559
Chicago/Turabian StyleYiğit, Esra Nur, Ekin Sönmez, Melis Savaşan Söğüt, Tunahan Çakır, and Işıl Aksan Kurnaz. 2018. "Validation of an In-Vitro Parkinson’s Disease Model for the Study of Neuroprotection" Proceedings 2, no. 25: 1559. https://doi.org/10.3390/proceedings2251559
APA StyleYiğit, E. N., Sönmez, E., Söğüt, M. S., Çakır, T., & Kurnaz, I. A. (2018). Validation of an In-Vitro Parkinson’s Disease Model for the Study of Neuroprotection. Proceedings, 2(25), 1559. https://doi.org/10.3390/proceedings2251559