Exploration of Predicted Nitrogen-Containing Natural Products from Northern Canadian Bark Beetle-Associated Bacteria

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection, Bacterial Isolation, and Cultivation
2.1.1. Sample Collection and Isolation
2.1.2. Cryogenic Storage of Bacterial Isolates
2.1.3. Assays for Strain Prioritization and Subsequent Crude Extract Preparation
2.2. Genomic Sequencing and BGC Analysis
2.3. NMR and Spectroscopic Profiling
2.4. General Experimental Procedures
2.5. List of Sample Name Abbreviations Used in the Text
- NBB—Northern bark beetle
- MPB—Mountain pine beetle (D. ponderosae)
- SBB—Spruce bark beetle (D. rufipennis)
- DFB—Douglas fir beetle (D. pseudotsugae)
- IPS—Ips perturbatus (a bark beetle species)
2.6. Use of Generative AI in Publication Preparation
3. Results
3.1. Bacterial Isolation and Cultivation
3.2. Secondary Metabolite Biosynthetic Gene Cluster Analysis
3.2.1. Overview of Genomic Features
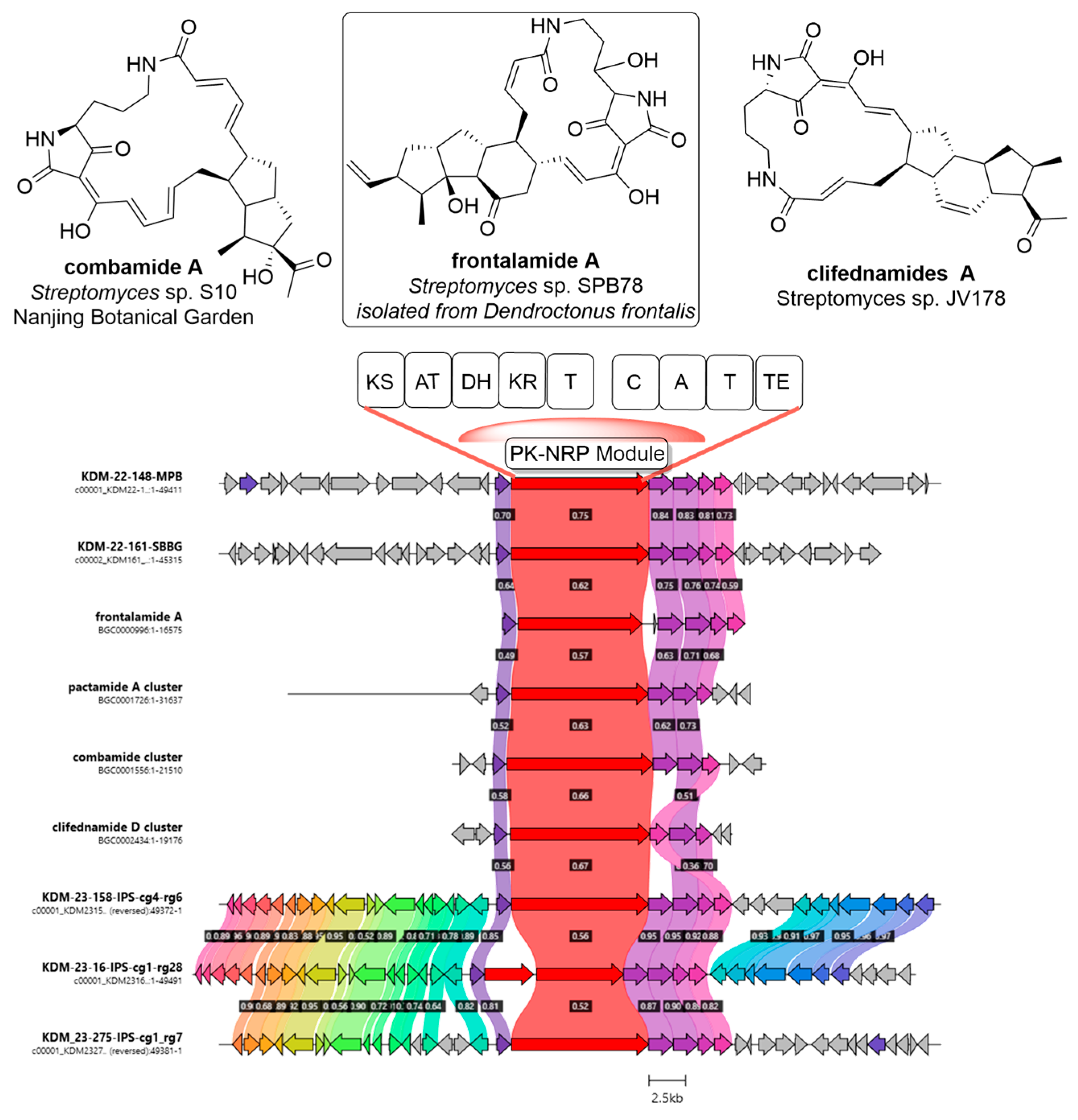
3.2.2. BGC Identification and Analysis
3.2.3. Gene Cluster Families, BGC Diversity, and Nitrogen-Containing BGCs
3.3. Nitrogen-15 NMR and Spectroscopic Profiling
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs over the Nearly Four Decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef] [PubMed]
- Bernardini, S.; Tiezzi, A.; Laghezza Masci, V.; Ovidi, E. Natural Products for Human Health: An Historical Overview of the Drug Discovery Approaches. Nat. Prod. Res. 2018, 32, 1926–1950. [Google Scholar] [CrossRef] [PubMed]
- Genilloud, O. Natural Products Discovery and Potential for New Antibiotics. Curr. Opin. Microbiol. 2019, 51, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Gerwick, B.C.; Sparks, T.C. Natural Products for Pest Control: An Analysis of Their Role, Value and Future. Pest Manag. Sci. 2014, 70, 1169–1185. [Google Scholar] [CrossRef] [PubMed]
- Choudoir, M.J.; Pepe-Ranney, C.; Buckley, D.H. Diversification of Secondary Metabolite Biosynthetic Gene Clusters Coincides with Lineage Divergence in Streptomyces. Antibiotics 2018, 7, 12. [Google Scholar] [CrossRef]
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; Supuran, C.T. Natural Products in Drug Discovery: Advances and Opportunities. Nat. Rev. Drug Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef]
- Park, C.J.; Caimi, N.A.; Buecher, D.C.; Valdez, E.W.; Northup, D.E.; Andam, C.P. Unexpected Genomic, Biosynthetic and Species Diversity of Streptomyces Bacteria from Bats in Arizona and New Mexico, USA. BMC Genom. 2021, 22, 247. [Google Scholar] [CrossRef]
- Beemelmanns, C.; Guo, H.; Rischer, M.; Poulsen, M. Natural Products from Microbes Associated with Insects. Beilstein J. Org. Chem. 2016, 12, 314–327. [Google Scholar] [CrossRef]
- Chanson, A.; Moreau, C.S.; Duplais, C. Assessing Biosynthetic Gene Cluster Diversity of Specialized Metabolites in the Conserved Gut Symbionts of Herbivorous Turtle Ants. Front. Microbiol. 2021, 12, 1640. [Google Scholar] [CrossRef]
- Arnam, E.B.V.; Currie, C.R.; Clardy, J. Defense Contracts: Molecular Protection in Insect-Microbe Symbioses. Chem. Soc. Rev. 2018, 47, 1638–1651. [Google Scholar] [CrossRef]
- Oh, D.-C.; Poulsen, M.; Currie, C.R.; Clardy, J. Dentigerumycin: A Bacterial Mediator of an Ant-Fungus Symbiosis. Nat. Chem. Biol. 2009, 5, 391–393. [Google Scholar] [CrossRef]
- Chevrette, M.G.; Carlson, C.M.; Ortega, H.E.; Thomas, C.; Ananiev, G.E.; Barns, K.J.; Book, A.J.; Cagnazzo, J.; Carlos, C.; Flanigan, W.; et al. The Antimicrobial Potential of Streptomyces from Insect Microbiomes. Nat. Commun. 2019, 10, 516. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-H.; Ko, H.; Bang, H.-S.; Park, S.-H.; Kim, D.-G.; Kwon, H.C.; Kim, S.Y.; Shin, J.; Oh, D.-C. Coprismycins A and B, Neuroprotective Phenylpyridines from the Dung Beetle-Associated Bacterium, Streptomyces sp. Bioorganic Med. Chem. Lett. 2011, 21, 5715–5718. [Google Scholar] [CrossRef] [PubMed]
- Dalisay, D.S.; Williams, D.E.; Wang, X.L.; Centko, R.; Chen, J.; Andersen, R.J. Marine Sediment-Derived Streptomyces Bacteria from British Columbia, Canada Are a Promising Microbiota Resource for the Discovery of Antimicrobial Natural Products. PLoS ONE 2013, 8, e77078. [Google Scholar] [CrossRef] [PubMed]
- Six, D.L. The Bark Beetle Holobiont: Why Microbes Matter. J. Chem. Ecol. 2013, 39, 989–1002. [Google Scholar] [CrossRef]
- Sickle, A.V.; Fiddick, R.L.; Wood, C.S. History of Forest Insect Investigations in British Columbia II. The Forest Insect and Disease Survey in the Pacific Region. J. Entomol. Soc. Br. Columbia 2001, 98, 169–176. [Google Scholar]
- 2021 Yukon Forest Health Report 2022. Available online: https://yukon.ca/en/2022-yukon-forest-health-report (accessed on 1 June 2023).
- Gayathri Samarasekera, G.D.N.; Bartell, N.V.; Lindgren, B.S.; Cooke, J.E.K.; Davis, C.S.; James, P.M.A.; Coltman, D.W.; Mock, K.E.; Murray, B.W. Spatial Genetic Structure of the Mountain Pine Beetle (Dendroctonus ponderosae) Outbreak in Western Canada: Historical Patterns and Contemporary Dispersal. Mol. Ecol. 2012, 21, 2931–2948. [Google Scholar] [CrossRef] [PubMed]
- Oh, D.-C.; Scott, J.J.; Currie, C.R.; Clardy, J. Mycangimycin, a Polyene Peroxide from a Mutualist Streptomyces sp. Org. Lett. 2009, 11, 633–636. [Google Scholar] [CrossRef] [PubMed]
- Kautsar, S.A.; Blin, K.; Shaw, S.; Navarro-Muñoz, J.C.; Terlouw, B.R.; van der Hooft, J.J.J.; van Santen, J.A.; Tracanna, V.; Suarez Duran, H.G.; Pascal Andreu, V.; et al. MIBiG 2.0: A Repository for Biosynthetic Gene Clusters of Known Function. Nucleic Acids Res. 2020, 48, D454–D458. [Google Scholar] [CrossRef] [PubMed]
- Chevrette, M.G.; Gavrilidou, A.; Mantri, S.; Selem-Mojica, N.; Ziemert, N.; Barona-Gómez, F. The Confluence of Big Data and Evolutionary Genome Mining for the Discovery of Natural Products. Nat. Prod. Rep. 2021, 38, 2024–2040. [Google Scholar] [CrossRef] [PubMed]
- Zdouc, M.M.; Blin, K.; Louwen, N.L.L.; Navarro, J.; Loureiro, C.; Bader, C.D.; Bailey, C.B.; Barra, L.; Booth, T.J.; Bozhüyük, K.A.J.; et al. MIBiG 4.0: Advancing Biosynthetic Gene Cluster Curation through Global Collaboration. Nucleic Acids Res. 2025, 53, D678–D690. [Google Scholar] [CrossRef]
- Blin, K.; Shaw, S.; Augustijn, H.E.; Reitz, Z.L.; Biermann, F.; Alanjary, M.; Fetter, A.; Terlouw, B.R.; Metcalf, W.W.; Helfrich, E.J.N.; et al. antiSMASH 7.0: New and Improved Predictions for Detection, Regulation, Chemical Structures and Visualisation. Nucleic Acids Res. 2023, 51, W46–W50. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Muñoz, J.C.; Selem-Mojica, N.; Mullowney, M.W.; Kautsar, S.A.; Tryon, J.H.; Parkinson, E.I.; De Los Santos, E.L.C.; Yeong, M.; Cruz-Morales, P.; Abubucker, S.; et al. A Computational Framework to Explore Large-Scale Biosynthetic Diversity. Nat. Chem. Biol. 2020, 16, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Morgan, K.D.; Williams, D.E.; Patrick, B.O.; Remigy, M.; Banuelos, C.A.; Sadar, M.D.; Ryan, K.S.; Andersen, R.J. Incarnatapeptins A and B, Nonribosomal Peptides Discovered Using Genome Mining and 1H/15N HSQC-TOCSY. Org. Lett. 2020, 22, 4053–4057. [Google Scholar] [CrossRef] [PubMed]
- Morgan, K.D.; Williams, D.E.; Ryan, K.S.; Andersen, R.J. Dentigerumycin F and G: Dynamic Structures Retrieved through a Genome-Mining/Nitrogen-NMR Methodology. Tetrahedron Lett. 2022, 94, 153688. [Google Scholar] [CrossRef]
- Hagar, M.; Morgan, K.D.; Stumpf, S.D.; Tsingos, M.; Banuelos, C.A.; Sadar, M.D.; Blodgett, J.A.V.; Andersen, R.J.; Ryan, K.S. Piperazate-Guided Isolation of Caveamides A and B, Cyclohexenylalanine-Containing Nonribosomal Peptides from a Cave Actinomycete. Org. Lett. 2024, 26, 4127–4131. [Google Scholar] [CrossRef]
- Breton, R.C.; Reynolds, W.F. Using NMR to Identify and Characterize Natural Products. Nat. Prod. Rep. 2013, 30, 501. [Google Scholar] [CrossRef]
- Witanowski, M.; Webb, G.A. Nitrogen NMR Spectroscopy. In Annual Reports on NMR Spectroscopy; Mooney, E.F., Ed.; Academic Press: London, UK, 1972; Volume 5, pp. 395–464. ISBN 978-0-12-505305-1. [Google Scholar]
- Martin, G.E.; Williams, A.J. Utilizing Long-Range 1 H–15 N 2-D NMR Spectroscopy for Chemical Structure Elucidation and Confirmation. In eMagRes; Harris, R.K., Wasylishen, R.L., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2010; ISBN 978-0-470-03459-0. [Google Scholar]
- Marek, R.; Lycka, A. 15N NMR Spectroscopy in Structural Analysis. COC 2002, 6, 35–66. [Google Scholar] [CrossRef]
- Hu, J.-Q.; Wang, J.-J.; Li, Y.-L.; Zhuo, L.; Zhang, A.; Sui, H.-Y.; Li, X.-J.; Shen, T.; Yin, Y.; Wu, Z.-H.; et al. Combining NMR-Based Metabolic Profiling and Genome Mining for the Accelerated Discovery of Archangiumide, an Allenic Macrolide from the Myxobacterium Archangium violaceum SDU8. Org. Lett. 2021, 23, 2114–2119. [Google Scholar] [CrossRef]
- Shin, D.; Byun, W.S.; Kang, S.; Kang, I.; Bae, E.S.; An, J.S.; Im, J.H.; Park, J.; Kim, E.; Ko, K.; et al. Targeted and Logical Discovery of Piperazic Acid-Bearing Natural Products Based on Genomic and Spectroscopic Signatures. J. Am. Chem. Soc. 2023, 145, 19676–19690. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Reheman, A.; Wan, C. Discovery of Anti-Mycobacterium Tuberculosis Desertomycins from Streptomyces Flavofungini TRM90047 Based on Genome Mining and HSQC-TOCSY. Sci. Rep. 2024, 14, 17006. [Google Scholar] [CrossRef] [PubMed]
- Williamson, R.T.; Márquez, B.L.; Gerwick, W.H. Use of 1H-15N PEP-HSQC-TOCSY at Natural Abundance to Facilitate the Structure Elucidation of Naturally Occurring Peptides. Tetrahedron 1999, 55, 2881–2888. [Google Scholar] [CrossRef]
- Morgan, K.D. The Use of Nitrogen-15 in Microbial Natural Product Discovery and Biosynthetic Characterization. Front. Microbiol. 2023, 14, 1174591. [Google Scholar] [CrossRef]
- Bauman, K.D.; Butler, K.S.; Moore, B.S.; Jonathan, R. Chekan Genome Mining Methods to Discover Bioactive Natural Products. Nat. Prod. Rep. 2021. [Google Scholar] [CrossRef]
- Pan, R.; Bai, X.; Chen, J.; Zhang, H.; Wang, H. Exploring Structural Diversity of Microbe Secondary Metabolites Using OSMAC Strategy: A Literature Review. Front. Microbiol. 2019, 10, 294. [Google Scholar] [CrossRef] [PubMed]
- Shin, Y.-H.; Im, J.H.; Kang, I.; Kim, E.; Jang, S.C.; Cho, E.; Shin, D.; Hwang, S.; Du, Y.E.; Huynh, T.-H.; et al. Genomic and Spectroscopic Signature-Based Discovery of Natural Macrolactams. J. Am. Chem. Soc. 2023, 145, 1886–1896. [Google Scholar] [CrossRef]
- Barras, S.J. Improved White’s Solution for Surface Sterilization of Pupae of Dendroctonus Frontalis1. J. Econ. Entomol. 1972, 65, 1504. [Google Scholar] [CrossRef] [PubMed]
- Blodgett, J.A.V.; Oh, D.-C.; Cao, S.; Currie, C.R.; Kolter, R.; Clardy, J. Common Biosynthetic Origins for Polycyclic Tetramate Macrolactams from Phylogenetically Diverse Bacteria. Proc. Natl. Acad. Sci. USA 2010, 107, 11692–11697. [Google Scholar] [CrossRef]
- Lauer, A.; Baal, J.D.; Mendes, S.D.; Casimiro, K.N.; Passaglia, A.K.; Valenzuela, A.H.; Guibert, G. Valley Fever on the Rise—Searching for Microbial Antagonists to the Fungal Pathogen Coccidioides Immitis. Microorganisms 2019, 7, 31. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Gilchrist, C.L.M.; Chooi, Y.-H. Clinker & Clustermap.Js: Automatic Generation of Gene Cluster Comparison Figures. Bioinformatics 2021, 37, 2473–2475. [Google Scholar] [CrossRef] [PubMed]
- Ha, S.-M.; Kim, C.K.; Roh, J.; Byun, J.-H.; Yang, S.-J.; Choi, S.-B.; Chun, J.; Yong, D. Application of the Whole Genome-Based Bacterial Identification System, TrueBac ID, Using Clinical Isolates That Were Not Identified With Three Matrix-Assisted Laser Desorption/Ionization Time-of-Flight Mass Spectrometry (MALDI-TOF MS) Systems. Ann. Lab. Med. 2019, 39, 530–536. [Google Scholar] [CrossRef] [PubMed]
- Mohite, O.S.; Jørgensen, T.S.; Booth, T.J.; Charusanti, P.; Phaneuf, P.V.; Weber, T.; Palsson, B.O. Pangenome Mining of the Streptomyces Genus Redefines Species’ Biosynthetic Potential. Genome Biol. 2025, 26, 9. [Google Scholar] [CrossRef] [PubMed]
- Van Den Belt, M.; Gilchrist, C.; Booth, T.J.; Chooi, Y.-H.; Medema, M.H.; Alanjary, M. CAGECAT: The CompArative GEne Cluster Analysis Toolbox for Rapid Search and Visualisation of Homologous Gene Clusters. BMC Bioinform. 2023, 24, 181. [Google Scholar] [CrossRef] [PubMed]
- Adams, A.S.; Boone, C.K.; Bohlmann, J.; Raffa, K.F. Responses of Bark Beetle-Associated Bacteria to Host Monoterpenes and Their Relationship to Insect Life Histories. J. Chem. Ecol. 2011, 37, 808–817. [Google Scholar] [CrossRef]
- Malit, J.J.L.; Wu, C.; Liu, L.-L.; Qian, P.-Y. Global Genome Mining Reveals the Distribution of Diverse Thioamidated RiPP Biosynthesis Gene Clusters. Front. Microbiol. 2021, 12, 635389. [Google Scholar] [CrossRef] [PubMed]
- Bracegirdle, J.; Hou, P.; Nowak, V.V.; Ackerley, D.F.; Keyzers, R.A.; Owen, J.G. Skyllamycins D and E, Non-Ribosomal Cyclic Depsipeptides from Lichen-Sourced Streptomyces Anulatus. J. Nat. Prod. 2021, 84, 2536–2543. Available online: https://pubs-acs-org.prxy.lib.unbc.ca/doi/10.1021/acs.jnatprod.1c00547 (accessed on 16 February 2025). [CrossRef]
- Toki, S.; Agatsuma, T.; Ochiai, K.; Saitoh, Y.; Ando, K.; Nakanishi, S.; Lokker, N.A.; Giese, N.A.; Matsuda, Y. RP-1776, a Novel Cyclic Peptide Produced by Streptomyces sp., Inhibits the Binding of PDGF to the Extracellular Domain of Its Receptor. J. Antibiot. 2001, 54, 405–414. [Google Scholar] [CrossRef]
- Soldatou, S.; Eldjárn, G.H.; Ramsay, A.; van der Hooft, J.J.J.; Hughes, A.H.; Rogers, S.; Duncan, K.R. Comparative Metabologenomics Analysis of Polar Actinomycetes. Mar. Drugs 2021, 19, 103. [Google Scholar] [CrossRef]
- Baltz, R.H. Gifted Microbes for Genome Mining and Natural Product Discovery. J. Ind. Microbiol. Biotechnol. 2017, 44, 573–588. [Google Scholar] [CrossRef]
- Xu, S.; Wang, N.; Meng, Q.; Ma, W.; Li, H. Metabologenomics-Driven Discovery of Nocardimicins from a Psychrophilic Nocardia Sp. Strain. J. Nat. Prod. 2025, 88, 103–109. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | Bark Beetle Source | Bark Beetle or Gallery | Location |
|---|---|---|---|
| KDM-22-177-SBBG | Dendroctonus rufipennis | Gallery of Infected Tree | Whisker’s Point, BC, Canada |
| KDM-22-269-MPB | Dendroctonus ponderosae | Crushed beetle | Cranbrook, BC, Canada |
| KDM-22-148-DFB | Dendroctonus pseudotsugae | Crushed beetle | Prince George, BC, Canada |
| KDM-22-161-SBBG | Dendroctonus rufipennis | Gallery of Infected Tree | Whisker’s Point, BC, Canada |
| KDM-22-185-DFB | Dendroctonus pseudotsugae | Crushed beetle | Prince George, BC, Canada |
| KDM-23-16-IPS | Ips perturbatus | Crushed beetle | Haines Junction, YT, Canada |
| KDM-23-410-IPS | Ips perturbatus | Crushed beetle | Haines Junction, YT, Canada |
| KDM-23-158-IPS | Ips perturbatus | Crushed beetle | Haines Junction, YT, Canada |
| KDM-23-275-IPS | Ips perturbatus | Crushed beetle | Haines Junction, YT, Canada |
| Isolate Name | Bacteria Species Most Closely Related To | Sequence ID %/Coverage % | Method of Analysis |
|---|---|---|---|
| KDM-22-177-SBBG | Bacillus velezensis | 98.3/69.8 | UBCG |
| KDM-22-269-MPB | Bacillus altitudinis | 99.4/68.1 | UBCG |
| KDM-22-148-DFB | Streptomyces albidoflavus | 99.1/69.9 | UBCG |
| KDM-22-161-SBBG | Streptomyces brevispora | 91.0/42.6 | UBCG |
| KDM-22-185-DFB | Streptomyces cinereospinus | 99.4/94.8 | UBCG |
| KDM-23-16-IPS | Streptomyces sp. CNB091 | 95.1/51.3 | UBCG |
| KDM-23-410-IPS | Streptomyces albidoflavus | 99.1/68.9 | UBCG |
| KDM-23-158-IPS | Streptomyces setonii | 96.2/51.4 | UBCG |
| KDM-23-275-IPS | Streptomyces microflavus | 98.2/66.5 | UBCG |
| Isolate Name | Genome Size (Mb) | No. of Contigs | GC Content (%) | N50 (bp) |
|---|---|---|---|---|
| Dendroctonus rufipennis | ||||
| KDM-22-177-SBBG | 3.94 | 52 | 46.42 | 319,638 |
| KDM-22-161-SBBG | 8.49 | 94 | 70.87 | 6,636,282 |
| Dendroctonus pseudotsugae | ||||
| KDM-22-148-DFB | 7.02 | 84 | 73.46 | 151,321 |
| KDM-22-185-DFB | 7.20 | 13 | 72.10 | 3,731,363 |
| Dendroctonus ponderosae | ||||
| KDM-22-269-MPB | 3.70 | 12 | 41.24 | 463,929 |
| Ips perterbatus | ||||
| KDM-23-16-IPS | 8.60 | 2 | 71.40 | 8,456,531 |
| KDM-23-410-IPS | 7.10 | 2 | 73.40 | 7,051,502 |
| KDM-23-158-IPS | 8.90 | 6 | 71.40 | 3,306,723 |
| KDM-23-275-IPS | 8.10 | 3 | 71.40 | 7,914,817 |
| Isolate Name | Number of BGCs * | # of PKS | # of NRPS | # of PKS/NRPS Hybrids | # of Terpenes | # of RIPPS |
|---|---|---|---|---|---|---|
| Dendroctonus rufipennis | ||||||
| KDM-22-177-SBBG | 10 # | 5 | 1 | 1 | 2 | 0 |
| KDM-22-161-SBBG | 30 # | 7 | 7 | 2 | 5 | 5 |
| Dendroctonus pseudotsugae | ||||||
| KDM-22-148-DFB | 21 # | 7 | 3 | 1 | 3 | 6 |
| KDM-22-185-DFB | 36 | 7 | 3 | 6 | 4 | 4 |
| Dendroctonus ponderosae | ||||||
| KDM-22-269-MPB | 9 | |||||
| Ips perterbatus | ||||||
| KDM-23-16-IPS | 35 | 4 | 2 | 3 | 4 | 10 |
| KDM-23-410-IPS | 21 | 1 | 2 | 2 | 3 | 6 |
| KDM-23-158-IPS | 31 | 4 | 2 | 4 | 5 | 6 |
| KDM-23-275-IPS | 33 | 5 | 3 | 2 | 5 | 6 |
| Total Numbers | 226 | 40 | 29 | 23 | 31 | 43 |
| Isolate Name | Crude Extract Media | 1H-15N HSQC Signals | Range 100–150 ppm | Range <100 ppm | Range > 150 ppm |
|---|---|---|---|---|---|
| Dendroctonus rufipennis | |||||
| KDM-22-177-SBBG | RBMII | Yes | 0 | 1 | 0 |
| KDM-22-161-SBBG | YMS/YEME | Yes | 8 | 1 | 1 |
| Dendroctonus pseudotsugae | |||||
| KDM-22-148-DFB | YMS | Yes | 4 | 0 | 0 |
| Dendroctonus ponderosae | |||||
| KDM-22-269-MPB | RBMII | Yes | 0 | 1 | 0 |
| KDM-22-185-DFB | YMS | Yes | 5 | 1 | 0 |
| Ips perterbatus | |||||
| KDM-23-16-IPS | YMS | Yes | 2 | 1 | 0 |
| KDM-23-410-IPS | YMS | Yes | 1 | 0 | 0 |
| KDM-23-158-IPS | YMS | Yes | 3 | 0 | 0 |
| KDM-23-275-IPS | YMS | Yes | 0 | 1 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Atapattu, N.; Justus, N.; Selvarajan, H.; Baylosis, M.; Schieven, M.; Morgan, K.D. Exploration of Predicted Nitrogen-Containing Natural Products from Northern Canadian Bark Beetle-Associated Bacteria. Nitrogen 2025, 6, 13. https://doi.org/10.3390/nitrogen6010013
Atapattu N, Justus N, Selvarajan H, Baylosis M, Schieven M, Morgan KD. Exploration of Predicted Nitrogen-Containing Natural Products from Northern Canadian Bark Beetle-Associated Bacteria. Nitrogen. 2025; 6(1):13. https://doi.org/10.3390/nitrogen6010013
Chicago/Turabian StyleAtapattu, Nirasha, Nicolas Justus, Hariniha Selvarajan, Mitzchilouise Baylosis, Marc Schieven, and Kalindi D. Morgan. 2025. "Exploration of Predicted Nitrogen-Containing Natural Products from Northern Canadian Bark Beetle-Associated Bacteria" Nitrogen 6, no. 1: 13. https://doi.org/10.3390/nitrogen6010013
APA StyleAtapattu, N., Justus, N., Selvarajan, H., Baylosis, M., Schieven, M., & Morgan, K. D. (2025). Exploration of Predicted Nitrogen-Containing Natural Products from Northern Canadian Bark Beetle-Associated Bacteria. Nitrogen, 6(1), 13. https://doi.org/10.3390/nitrogen6010013