

Structural Traits, Production, Biomass Allocation, and Changes in Leaf Investment in Megathyrsus maximus cv. MG12 Paredão Under Nitrogen Fertilization and Cutting Intervals



Abstract
1. Introduction
2. Materials and Methods
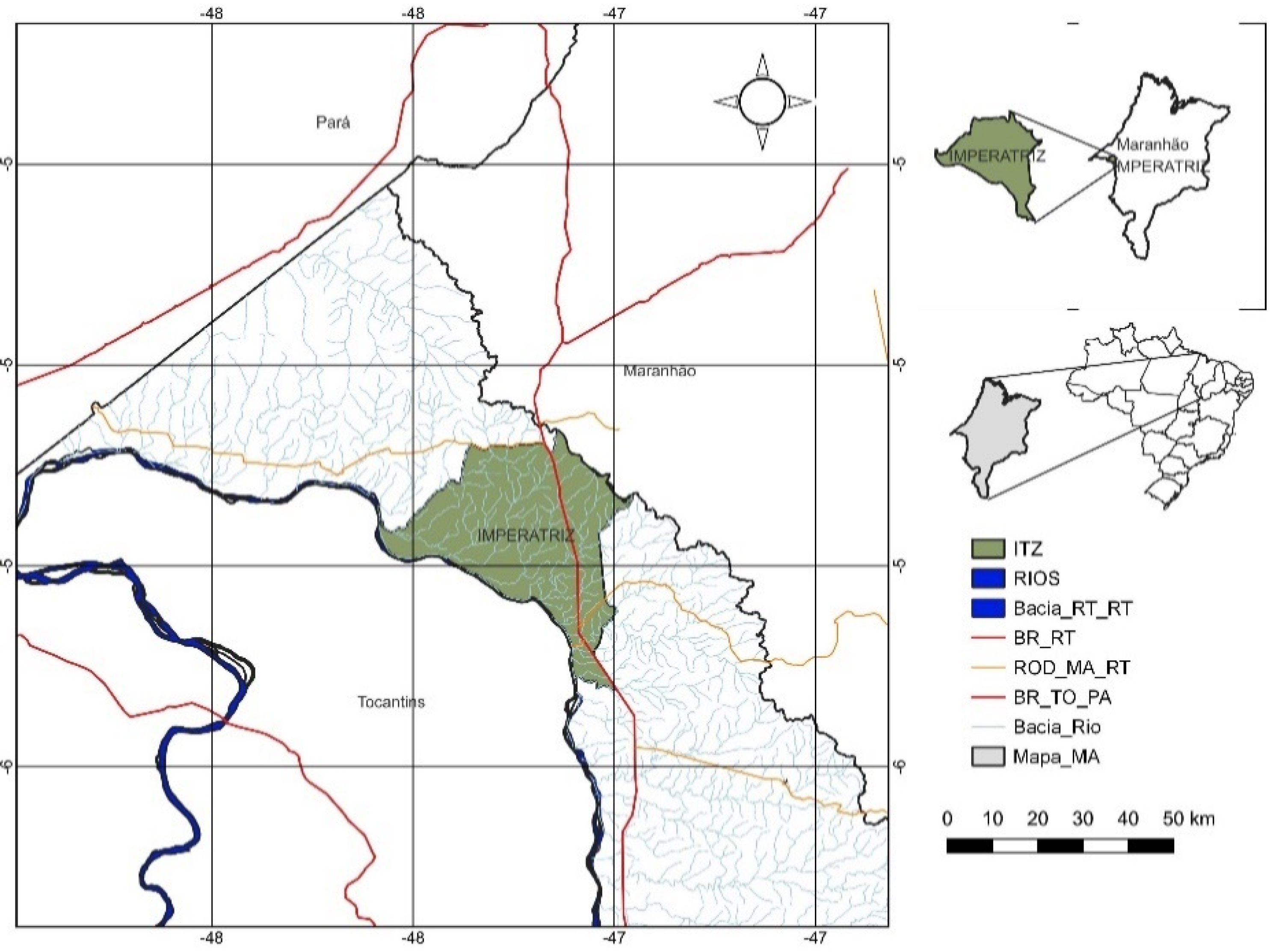
2.1. Experimental Area
2.2. Experimental Design and Soil Preparation
2.3. Fertilizer Application, Sample Collection, and Biomass Allocation
2.4. Soil Moisture Content
2.5. Specific Leaf Area and Leaf Green Color Index (SPAD Index)
2.6. Statistical Analysis
3. Results
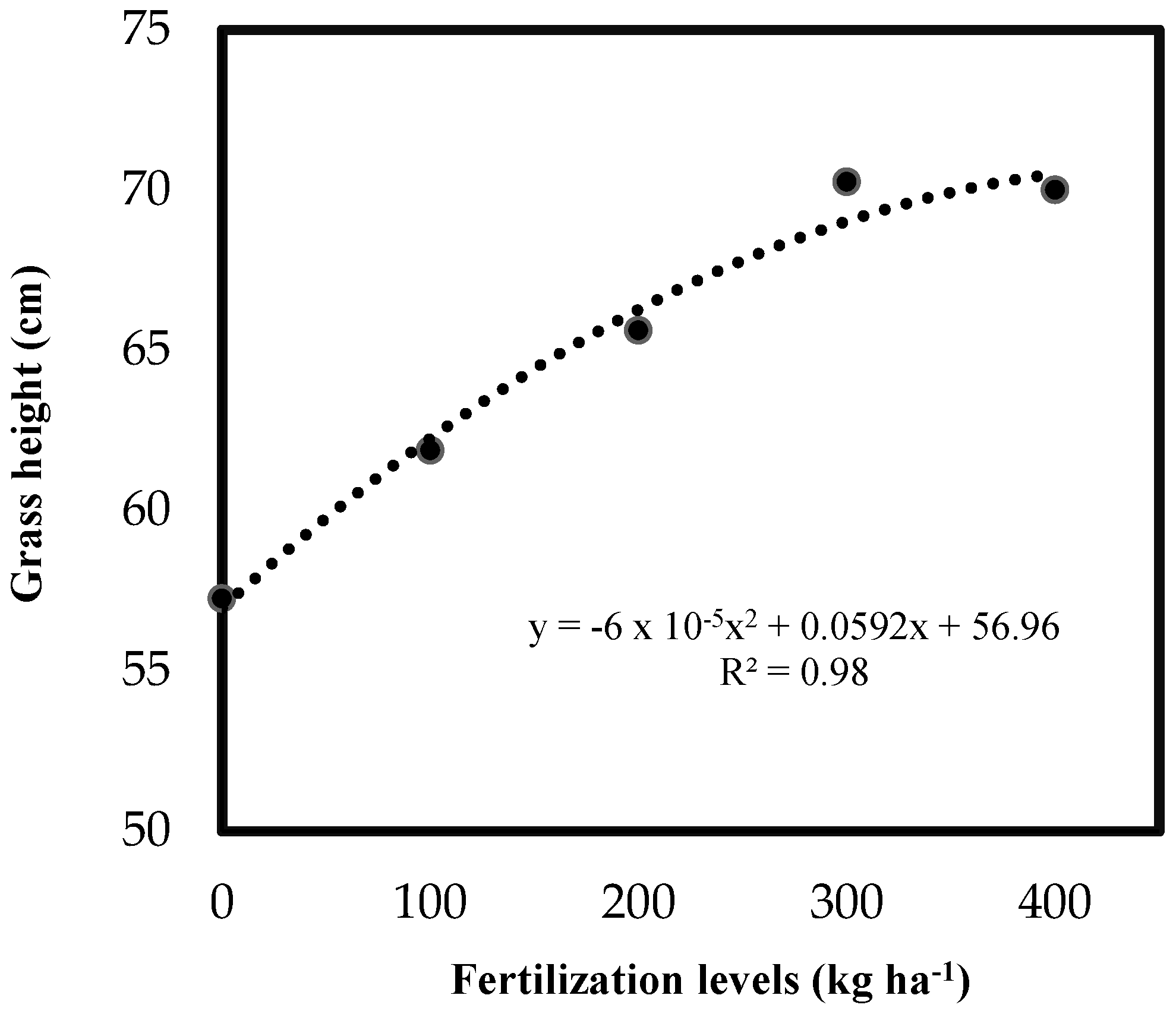
3.1. Structural and Productive Traits
3.2. Soil Moisture, Biomass Allocation, and Changes in Leaf Investment
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CV | Coefficient of variation |
| DML | Dry mass of leaves |
| DMP | Dry matter production |
| DMR | Dry mass of roots |
| DMS | Dry mass of stems |
| IBGE | Brazilian Institute of Geography and Statistics |
| LDM | Leaf dry mass |
| M | Aluminum saturation |
| N | Nitrogen |
| OM | Organic matter of the soil |
| PVC | Polyvinyl chloride |
| RDM | Root dry mass |
| SDM | Stem dry mass |
| SB | Sum of bases |
| SLA | Specific leaf area |
| SPAD-502 | Soil–Plant Analysis Development |
| TFM | Total fresh mass |
| UEMASUL | University of the Maranhão Tocantina Region |
| V | Base saturation |
References
- Moraes, D.A.; Moares, D.B.; Giacon, M.; Diogo, R. A Relação da Pecuária na Economia Brasileira; Repositório Institucional do Conhecimento do Centro Paula Souza (RIC-CPS): São Paulo, Brazil, 2021. [Google Scholar]
- Brasil. Ministério da Agricultura e Pecuária. Produção da Pecuária Nacional. 2023. Available online: https://www.gov.br/agricultura (accessed on 14 December 2024).
- Lavres, J., Jr.; Santos, J.D.G., Jr.; Monteiro, F.A. Nitrate reductase activity and spad readings in leaf tissues of guinea grass submitted to nitrogen and potassium rates. Rev. Bras. Ciênc. Solo 2010, 34, 801–809. [Google Scholar] [CrossRef]
- Witkowski, E.T.F.; Lamont, B.B. Leaf specific mass confounds leaf density and thickness. Oecologia 1991, 88, 486–493. [Google Scholar] [CrossRef] [PubMed]
- Garnier, E.; Cordonnier, P.; Giullerm, J.L.; Sonie, L. Specific leaf area and leaf nitrogen concentration in annual and perennial grass species growing in Mediterranean old-fields. Oecologia 1997, 111, 409–498. [Google Scholar] [CrossRef] [PubMed]
- Reich, P.B. Reconciling apparent discrepancies among studies relating life span, structure and function of leaves in contrasting plant life forms and climates: “the blind men and the elephant retold”. Funct. Ecol. 1993, 7, 721–725. [Google Scholar] [CrossRef]
- Wellstein, C.; Poschlod, P.; Gohlke, A.; Chelli, S.; Campetella, G.; Rosbakh, S.; Beierkuhnlein, C. Effects of extreme drought on specific leaf area of grassland species: A meta-analysis of experimental studies in temperate and sub-Mediterranean systems. Glob. Change Biol. 2017, 23, 2473–2481. [Google Scholar] [CrossRef] [PubMed]
- Freschet, G.T.; Valverde-Barrantes, O.J.; Tucker, C.M.; Craine, J.M.; Mccormack, M.L.; Violle, C.; Fort, F.; Blackwood, C.B.; Urban-Mead, K.R.; Iversen, C.M.; et al. Climate, soil and plant functional types as drivers of global fine-root trait variation. J. Ecol. 2017, 105, 1182–1196. [Google Scholar] [CrossRef]
- Costa, N.L. Formação, Manejo e Recuperação de Pastagens em Rondônia; Embrapa Rondônia: Porto Velho, Brazil, 2004; 212p. [Google Scholar]
- Lima, U.F.; Castro, L.M.O.; Pimentel, A.J.B.; Silva, J.L. Emergence of Megathyrsus maximus seedlings grown under salt stress. Rev. Agric. Neotrop. 2024, 11, e8266. [Google Scholar] [CrossRef]
- Silva, A.L.; Sayd, R.M. Levantamento e Descrição de Cultivares Forrageiras Utilizadas no Brasil; REAL—Repositório Institucional; ICESP: Brasilia, Brazil, 2023; Volume 2. [Google Scholar]
- Matsuda. Sementes MG 12 Paredão. Sementes Matsuda. 2022. Available online: https://www.matsuda.com.br/sementes-forrageiras/produto/mg12-paredao (accessed on 4 November 2022).
- Barros, L.F.; Araújo, L.C.; Andrade, K.S.P.; Mota, C.R.D.M.; Silva, A.M.; Ribeiro, A.L.; Rodrigues, W.P.; Rocha, T.C. Efeito dos dias de corte nas características morfológicas do Panicum maximum cv. Paredão com níveis de adubação nitrogenada no período da seca. Rev. Aracê 2024, 6, 13281–13294. [Google Scholar] [CrossRef]
- Zhang, K.; Zhai, C.; Li, Y.; Li, Y.; Qu, H.; Shen, Y. Effect of nitrogen application and cutting frequency on the yield and forage quality of alfalfa in seasonal cultivation. Agriculture 2023, 13, 1063. [Google Scholar] [CrossRef]
- Silva, F.C.; Eira, P.A.; Barreto, W.O.; Pérez, D.V.; Silva, C.A. Manual of Chemical Analysis Methods for Soil Fertility Evaluation; Embrapa CNPS: Rio de Janeiro, Brazil, 1998; Volume 56, p. 3. [Google Scholar]
- Gomide, C.A.M.; Paciullo, D.S.C.; Leite, J.L.B.; Resende, H. Panicum maximum cvs. Tanzânia e Mombaça para uso em Pastejo: Produção e Custo; Circular Técnica; Embrapa Gado de Corte: Juiz de Fora, Brazil, 2016; p. 7. [Google Scholar]
- Caputo, H.P.; Caputo, A.N. Mecânica dos solos e Suas Aplicações, 7th ed.; LTC: Rio de Janeiro, Brazil, 2017. [Google Scholar]
- Reis, M.B.; Araújo, L.C.; Santos, L.C.; Silva, A.M.; Ribeiro, A.L.; Silva, J.V.S.; Andrade, K.S.P.; Rocha, T.C. Influência dos dias de corte e níveis de adubação nitrogenada nas características morfológicas no Panicum maximum cv. Mombaça e cv. Paredão. Cad. Pedagóg. 2024, 21, e9719. [Google Scholar] [CrossRef]
- Motta, M.M.; Mota, L.G.; Melo, K.K.; Silva, P.R.; Santos, A.R.M.; Motta, L.J.M.; Cabral, C.H.A.; Cabral, C.E.A. Interval between defoliation and nitrogen fertilization of Panicum maximum cultivars. Bol. Ind. Anim. 2021, 78, 1–12. [Google Scholar] [CrossRef]
- Werner, J.C. Adubação de pastagens. Bol. Téc. 1986, 18. [Google Scholar]
- Araújo, L.M.B.; Andrade, A.L.; Rodrigues, B.H.N.; Santos, F.J.S.; Magalhães, J.A.; Rodrigues, R.C.; Oliveira, I.V.L. Produtividade do capim-Mombaça sob diferentes idades de rebrotação no Norte do Piauí. Nucleus 2019, 16, 233–244. [Google Scholar] [CrossRef]
- Luo, L.; Zhang, Y.; Xu, G. How does nitrogen shape plant architecture? J. Exp. Bot. 2020, 71, 4415–4427. [Google Scholar] [CrossRef] [PubMed]
- Santos, M.E.R.; Fonseca, D.M.; Euclides, V.P.B.; Ribeiro Júnior, J.I.R.; Balbino, E.M.; Casagrande, D.R. Valor nutritivo da forragem e de seus componentes morfológicos em pastagens de Brachiaria decumbens diferida. Bol. De Indústria Anim. 2008, 65, 303–311. [Google Scholar]
- Fontaneli, R.S.; Fontaneli, R.S. Qualidade e valor nutritivo de forragem. In Forrageiras ILPF—Integração Lavoura-Pecuária-Floresta, 1st ed.; Embrapa Trigo: Passos Fundo, Brazil, 2009; 28p. [Google Scholar]
- Soratto, R.P.; Carvalho, M.A.C.; ARF, O. Teor de clorofila e produtividade do feijoeiro em razão da adubação nitrogenada. Pesqui. Agropecu. 2004, 39, 895–901. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sorption Complex | Saturation of the Sorption Complex | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| pH | OM | P | K | Ca | Mg | AI | H + AI | SB | CTC | V | m | Ca | Mg | K |
| CaCI2 | g/kg | mg/dm3 | cmol/dm3 | % | ||||||||||
| 4.6 | 13.8 | 3.8 | 0.08 | 1.94 | 0.59 | 0.00 | 3.24 | 2.61 | 5.85 | 44.6 | 0.0 | 33.2 | 10.1 | 1.3 |
| Cutting Days | Fertilization Levels (kg ha−1) | CV (%) | |||||
|---|---|---|---|---|---|---|---|
| Item | 0 | 100 | 200 | 300 | 400 | ||
| Grass height (m) | 28 | 0.325 Bb | 0.475 Bab | 0.550 Ba | 0.500 Ba | 0.500 Ba | 7.5 |
| 56 | 0.800 A | 0.950 A | 0.825 A | 0.875 A | 0.900 A | ||
| Number of tillers | 28 | 108.0 | 62.0 | 62.5 | 80.0 | 91.0 | 35.8 |
| 56 | 94.5 | 96.0 | 93.5 | 101.0 | 59.5 | ||
| Leaves (%) | 28 | 100.0 | 100.0 | 100.0 | 100.0 | 92.8 | 5.5 |
| 56 | 100.0 | 100.0 | 100.0 | 90.4 | 100.0 | ||
| DMP (g) | 28 | 8000.0 | 4700.0 | 7400.0 | 6000.0 | 10600.0 A | 33.6 |
| 56 | 6700.0 | 8100.0 | 9100.0 | 11100.0 | 4300.0 B | ||
| Cutting Days | Fertilization Levels (kg N ha−1) | CV (%) | |||||
|---|---|---|---|---|---|---|---|
| Item | 0 | 100 | 200 | 300 | 400 | ||
| Gass height (m) | 28 | 0.400 Bb | 0.625 Bab | 0.750 Aa | 0.650 Ba | 0.650 Ba | 9.1 |
| 56 | 0.825 A | 0.975 A | 0.825 A | 0.925 A | 0.925 A | ||
| Number of tillers | 28 | 32.5 b | 50.0 Aab | 51.5 Aab | 58.0 Aa | 56.0 Aa | 17.5 |
| 56 | 26.0 | 24.5 B | 23.0 B | 37.5 B | 35.0 B | ||
| Leaves% | 28 | 100.0 | 81.6 | 100.0 | 83.7 | 100.0 | 11.4 |
| 56 | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 | ||
| DMP (g) | 28 | 2120.0 | 1440.0 | 1780.0 | 2160.0 B | 2380.0 | 24.0 |
| 56 | 1940.0 | 2300.0 | 2540.0 | 3540 A | 2560.0 | ||
| Cutting Days | Fertilization Levels (kg N ha−1) | CV (%) | |||||
|---|---|---|---|---|---|---|---|
| Item | 0 | 100 | 200 | 300 | 400 | ||
| Grass height (m) | 28 | 0.400 Bb | 0.375 Bb | 0.425 Bab | 0.525 Ba | 0.425 Bab | 5.7 |
| 56 | 0.800 Ab | 0.875 Aab | 0.850 Aab | 0.875 Aab | 0.925 Aa | ||
| Number of tillers | 28 | 90.0 | 75.0 | 84.5 | 98.5 | 103.5 | 21.8 |
| 56 | 94.0 | 116.0 | 96.5 | 90.0 | 96.5 | ||
| Leaves (%) | 28 | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 | 0.0 |
| 56 | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 | ||
| DMP (g) | 28 | 1400.0 B | 1400.0 B | 1000.0 B | 1200.0 B | 1100.0 B | 24.0 |
| 56 | 5800.0 Aab | 4300.0 Ab | 5220.0 Ab | 5760.0 Aab | 8100.0 Aa | ||
| Cutting Days | Fertilization Levels (kg N ha−1) | Average | CV (%) | |||||
|---|---|---|---|---|---|---|---|---|
| Item | 0 | 100 | 200 | 300 | 400 | |||
| TFM (g) | 28 | 290.00 ns | 286.25 ns | 327.50 ns | 697.57 a | 306.25 ns | 381.50 ns | 43.83 |
| 56 | 287.50 ns | 375.00 ns | 379.50 ns | 423.75 b | 316.25 ns | 356.40 ns | ||
| SPAD-502 | 28 | 30.23 ns | 27.85 ns | 32.60 ns | 30.45 ns | 26.90 b | 29.6 ns | 19.83 |
| 56 | 24.95 ns | 25.33 ns | 27.08 ns | 22.88 ns | 37.48 a | 27.54 ns | ||
| DML % | 28 | 22.00 ns | 16.25 b | 18.75 ns | 20.50 ns | 25.75 ns | 20.65 ns | 26.39 |
| 56 | 14.50 ns | 27.25 a | 25.25 ns | 26.75 ns | 25.00 ns | 23.75 ns | ||
| DML % | 28 | 52.00 ns | 51.75 ns | 53.75 ns | 51.25 ns | 48.25 ns | 51.40 ns | 27.19 |
| 56 | 55.25 ns | 41.75 ns | 42.50 ns | 46.75 ns | 43.00 ns | 45.85 ns | ||
| DMS % | 28 | 26.00 ns | 32.00 ns | 27.00 ns | 28.50 ns | 26.00 ns | 27.90 ns | 37.39 |
| 56 | 30.25 ns | 30.50 ns | 31.75 ns | 26.50 ns | 31.75 ns | 30.15 ns | ||
| SLA (cm2 g−1) | 28 | 82.19 ns | 80.63 ns | 83.95 ns | 69.86 ns | 85.89 ns | 80.50 ns | 69.76 |
| 56 | 67.76 ns | 62.73 ns | 60.96 ns | 63.41ns | 64.50 ns | 63.87 ns | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pinheiro, V.L.; Mendonça, A.S.; Cruz, D.H.G.; Araújo, L.C.; Andrade, K.S.P.; Rocha, T.C.; Rodrigues, W.P. Structural Traits, Production, Biomass Allocation, and Changes in Leaf Investment in Megathyrsus maximus cv. MG12 Paredão Under Nitrogen Fertilization and Cutting Intervals. Nitrogen 2025, 6, 12. https://doi.org/10.3390/nitrogen6010012
Pinheiro VL, Mendonça AS, Cruz DHG, Araújo LC, Andrade KSP, Rocha TC, Rodrigues WP. Structural Traits, Production, Biomass Allocation, and Changes in Leaf Investment in Megathyrsus maximus cv. MG12 Paredão Under Nitrogen Fertilization and Cutting Intervals. Nitrogen. 2025; 6(1):12. https://doi.org/10.3390/nitrogen6010012
Chicago/Turabian StylePinheiro, Vinícus L., Andressa S. Mendonça, Danny Hellen G. Cruz, Laylles C. Araújo, Kele S. P. Andrade, Tiago C. Rocha, and Weverton P. Rodrigues. 2025. "Structural Traits, Production, Biomass Allocation, and Changes in Leaf Investment in Megathyrsus maximus cv. MG12 Paredão Under Nitrogen Fertilization and Cutting Intervals" Nitrogen 6, no. 1: 12. https://doi.org/10.3390/nitrogen6010012
APA StylePinheiro, V. L., Mendonça, A. S., Cruz, D. H. G., Araújo, L. C., Andrade, K. S. P., Rocha, T. C., & Rodrigues, W. P. (2025). Structural Traits, Production, Biomass Allocation, and Changes in Leaf Investment in Megathyrsus maximus cv. MG12 Paredão Under Nitrogen Fertilization and Cutting Intervals. Nitrogen, 6(1), 12. https://doi.org/10.3390/nitrogen6010012