
N Losses from an Andisol via Gaseous N2O and N2 Emissions Increase with Increasing Ruminant Urinary–N Deposition Rate

 ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Soil Sampling and Preconditioning
2.2. Urine Collection and Preparation
2.3. Experimental Setup
2.4. Gas Analyses and Data Management
2.5. Soil Analysis
2.6. Statistical Analysis
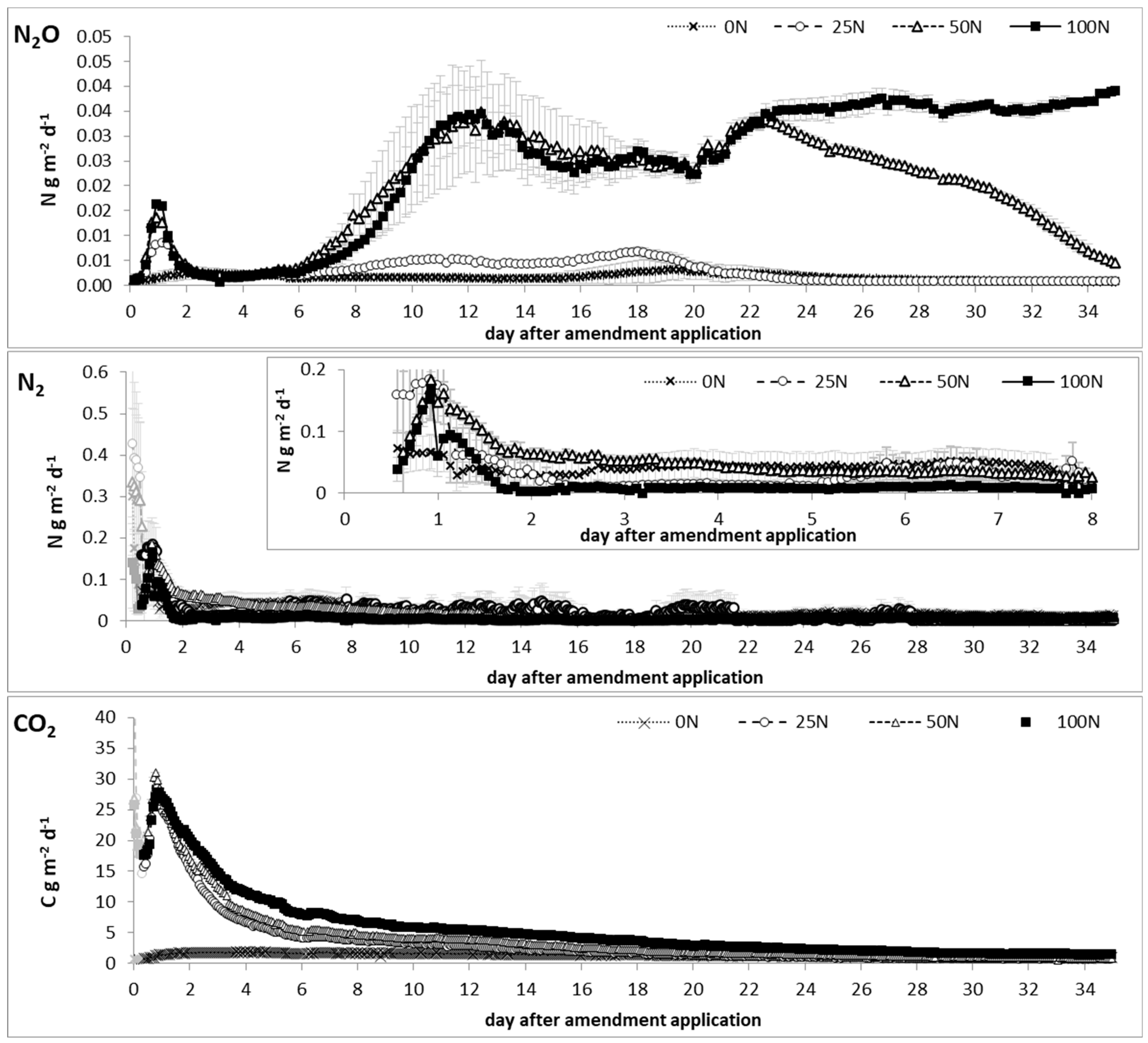
3. Results
3.1. Gaseous Emissions
3.2. Soil Chemistry
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Haynes, R.J.; Williams, P.H. Nutrient Cycling and Soil Fertility in the Grazed Pasture Ecosystem. In Advances in Agronomy; Sparks, D.L., Ed.; Academic Press: Cambridge, MA, USA, 1993; pp. 119–199. [Google Scholar]
- Ledgard, S.F.; Sprosen, M.S.; Penno, J.W.; Rajendram, G.S. Nitrogen fixation by white clover in pastures grazed by dairy cows: Temporal variation and effects of nitrogen fertilization. Plant Soil 2001, 229, 177–187. [Google Scholar] [CrossRef]
- Bolan, N.S.; Saggar, S.; Luo, J.; Bhandral, R.; Singh, J. Gaseous Emissions of Nitrogen from Grazed Pastures: Processes, Measurements and Modelling, Environmental Implications, and Mitigation. In Advances in Agronomy; Academic Press: Cambridge, MA, USA, 2004; pp. 37–120. [Google Scholar]
- Saggar, S.; Luo, J.; Kim, D.-G.; Jha, N. Intensification in Pastoral Farming: Impacts on Soil Attributes and Gaseous Emissions. In Soil Health and Climate Change; Singh, B.P., Cowie, A.L., Chan, K.Y., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 207–236. [Google Scholar]
- Somers, C.; Girkin, N.T.; Rippey, B.; Lanigan, G.J.; Richards, K.G. The effects of urine nitrogen application rate on nitrogen transformations in grassland soils. J. Agric. Sci. 2019, 157, 515–522. [Google Scholar] [CrossRef]
- Moreau, D.; Bardgett, R.D.; Finlay, R.D.; Jones, D.L.; Philippot, L. A plant perspective on nitrogen cycling in the rhizosphere. Funct. Ecol. 2019, 33, 540–552. [Google Scholar] [CrossRef]
- Kuypers, M.M.M.; Marchant, H.K.; Kartal, B. The microbial nitrogen-cycling network. Nat. Rev. Microbiol. 2018, 16, 263–276. [Google Scholar] [CrossRef]
- Heil, J.; Vereecken, H.; Brüggemann, N. A review of chemical reactions of nitrification intermediates and their role in nitrogen cycling and nitrogen trace gas formation in soil. Eur. J. Soil Sci. 2016, 67, 23–39. [Google Scholar] [CrossRef]
- Cameron, K.C.; Di, H.J.; Moir, J.L. Nitrogen losses from the soil/plant system: A review. Ann. Appl. Biol. 2013, 162, 145–173. [Google Scholar] [CrossRef]
- Saggar, S.; Jha, N.; Deslippe, J.; Bolan, N.S.; Luo, J.; Giltrap, D.L.; Kim, D.G.; Zaman, M.; Tillman, R.W. Denitrification and N2O:N2 production in temperate grasslands: Processes, measurements, modelling and mitigating negative impacts. Sci. Total Environ. 2013, 465, 173–195. [Google Scholar] [CrossRef]
- Knowles, R. Denitrification. Microbiol. Rev. 1982, 46, 43–70. [Google Scholar] [CrossRef]
- van Spanning, R.J.M.; Richardson, D.J.; Ferguson, S.J. Chapter 1—Introduction to the Biochemistry and Molecular Biology of Denitrification. In Biology of the Nitrogen Cycle; Bothe, H., Ferguson, S.J., Newton, W.E., Eds.; Elsevier: Amsterdam, The Netherlands, 2007; pp. 3–20. [Google Scholar]
- IPCC. Climate change. In Synthesis Report of the Fourth Assessment Report of IPCC, Chapter 3; IPCC: Geneva, Switzerland, 2007; 49p. [Google Scholar]
- Kroeze, C.; Mosier, A.; Bouwman, L. Closing the global N2O budget: A retrospective analysis 1500–1994. Glob. Biogeochem. Cycles 1999, 13, 1–8. [Google Scholar] [CrossRef]
- Mosier, A.R.; Duxbury, J.M.; Freney, J.R.; Heinemeyer, O.; Minami, K. Assessing and Mitigating N2O Emissions from Agricultural Soils. Clim. Chang. 1998, 40, 7–38. [Google Scholar] [CrossRef]
- Skopp, J.; Jawson, M.D.; Doran, J.W. Steady-State Aerobic Microbial Activity as a Function of Soil Water Content. Soil Sci. Soc. Am. J. 1990, 54, 1619–1625. [Google Scholar] [CrossRef]
- Saggar, S.; Bolan, N.S.; Bhandral, R.; Hedley, C.B.; Luo, J. A review of emissions of methane, ammonia, and nitrous oxide from animal excreta deposition and farm effluent application in grazed pastures. N. Z. J. Agric. Res. 2004, 47, 513–544. [Google Scholar] [CrossRef]
- Van Vuuren, A.M.; Meijs, J.A.C. Effects of herbage composition and supplement feeding on the excretion of nitrogen in dung and urine by grazing dairy cows. In Animal Manure on Grassland and Fodder Crops. Fertilizer or Waste? Proceedings of an International Symposium of the European Grassland Federation, Wageningen, The Netherlands, 31 August–3 September 1987; Van Der Meer, H.G., Unwin, R.J., Van Dijk, T.A., Ennik, G.C., Eds.; Springer: Dordrecht, The Netherlands, 1987; pp. 17–25. [Google Scholar]
- Oenema, O.; Velthof, G.L.; Yamulki, S.; Jarvis, S.C. Nitrous oxide emissions from grazed grassland. Soil Use Manag. 1997, 13, 288–295. [Google Scholar] [CrossRef]
- Misselbrook, T.; Fleming, H.; Camp, V.; Umstatter, C.; Duthie, C.A.; Nicoll, L.; Waterhouse, T. Automated monitoring of urination events from grazing cattle. Agric. Ecosyst. Environ. 2016, 230, 191–198. [Google Scholar] [CrossRef]
- Clough, T.J.; Sherlock, R.R.; Cameron, K.C.; Ledgard, S.F. Fate of urine nitrogen on mineral and peat soils in New Zealand. Plant Soil 1996, 178, 141–152. [Google Scholar] [CrossRef]
- Cahalan, E.; Minet, E.; Ernfors, M.; Müller, C.; Devaney, D.; Forrestal, P.J.; Richards, K.G. The effect of precipitation and application rate on dicyandiamide persistence and efficiency in two Irish grassland soils. Soil Use Manag. 2015, 31, 367–374. [Google Scholar] [CrossRef]
- McGeough, K.L.; Watson, C.J.; Müller, C.; Laughlin, R.J.; Chadwick, D.R. Evidence that the efficacy of the nitrification inhibitor dicyandiamide (DCD) is affected by soil properties in UK soils. Soil Biol. Biochem. 2016, 94, 222–232. [Google Scholar] [CrossRef]
- Padmanabhan, E.; Reich, P.F. World soil map based on soil taxonomy. In Encyclopedia of Soils in the Environment, 2nd ed.; Goss, M.J., Oliver, M., Eds.; Academic Press: Oxford, UK, 2023; pp. 218–231. [Google Scholar]
- Delmelle, P.; Opfergelt, S.; Cornelis, J.-T.; Ping, C.-L. Chapter 72—Volcanic Soils. In The Encyclopedia of Volcanoes, 2nd ed.; Sigurdsson, H., Ed.; Academic Press: Amsterdam, The Netherlands, 2015; pp. 1253–1264. [Google Scholar]
- Dörner, J.; Dec, D.; Peng, X.; Horn, R. Effect of land use change on the dynamic behaviour of structural properties of an Andisol in southern Chile under saturated and unsaturated hydraulic conditions. Geoderma 2010, 159, 189–197. [Google Scholar] [CrossRef]
- Salazar, F.; Martínez-Lagos, J.; Alfaro, M.; Misselbrook, T. Ammonia emissions from urea application to permanent pasture on a volcanic soil. Atmos. Environ. 2012, 61, 395–399. [Google Scholar] [CrossRef]
- Martínez-Lagos, J.; Salazar, F.; Alfaro, M.; Misselbrook, T. Ammonia volatilization following dairy slurry application to a permanent grassland on a volcanic soil. Atmos. Environ. 2013, 80, 226–231. [Google Scholar] [CrossRef]
- Vistoso, E.; Alfaro, M.; Saggar, S.; Salazar, F. Effect of Nitrogen Inhibitors on Nitrous Oxide Emissions and Pasture Growth After an Autumn Application in Volcanic Soil. Chil. J. Agric. Res. 2012, 72, 133–139. [Google Scholar] [CrossRef][Green Version]
- Hube, S.; Alfaro, M.A.; Scheer, C.; Brunk, C.; Ramírez, L.; Rowlings, D.; Grace, P. Effect of nitrification and urease inhibitors on nitrous oxide and methane emissions from an oat crop in a volcanic ash soil. Agric. Ecosyst. Environ. 2017, 238, 46–54. [Google Scholar] [CrossRef]
- Toro-Mujica, P.; Vera, R.; Vargas-Bello-Pérez, E.; Pinedo, P.; Bas, F. Trends and Drivers of Change of Pastoral Beef Production Systems in a Mediterranean-Temperate Climate Zone of Chile. Animals 2019, 9, 1135. [Google Scholar] [CrossRef] [PubMed]
- Kirchmann, H.; Esala, M.; Morken, J.; Ferm, M.; Bussink, W.; Gustavsson, J.; Jakobsson, C. Ammonia emissions from agriculture. Nutr. Cycl. Agroecosystems 1998, 51, 1–3. [Google Scholar] [CrossRef]
- Cárdenas, L.M.; Hawkins, J.M.B.; Chadwick, D.; Scholefield, D. Biogenic gas emissions from soils measured using a new automated laboratory incubation system. Soil Biol. Biochem. 2003, 35, 867–870. [Google Scholar] [CrossRef]
- Loick, N.; Dixon, E.; Abalos, D.; Vallejo, A.; Matthews, P.; McGeough, K.; Watson, C.; Baggs, E.M.; Cardenas, L.M. “Hot spots” of N and C impact nitric oxide, nitrous oxide and nitrogen gas emissions from a UK grassland soil. Geoderma 2017, 305, 336–345. [Google Scholar] [CrossRef]
- Bergstermann, A.; Cárdenas, L.; Bol, R.; Gilliam, L.; Goulding, K.; Meijide, A.; Scholefield, D.; Vallejo, A.; Well, R. Effect of antecedent soil moisture conditions on emissions and isotopologue distribution of N2O during denitrification. Soil Biol. Biochem. 2011, 43, 240–250. [Google Scholar] [CrossRef]
- Meijide, A.; Cardenas, L.M.; Bol, R.; Bergstermann, A.; Goulding, K.; Well, R.; Vallejo, A.; Scholefield, D. Dual isotope and isotopomer measurements for the understanding of N2O production and consumption during denitrification in an arable soil. Eur. J. Soil Sci. 2010, 61, 364–374. [Google Scholar] [CrossRef]
- Burns, L.C.; Stevens, R.J.; Laughlin, R.J. Determination of the simultaneous production and consumption of soil nitrite using 15N. Soil Biol. Biochem. 1995, 27, 839–844. [Google Scholar] [CrossRef]
- Burns, L.C.; Stevens, R.J.; Laughlin, R.J. Production of nitrite in soil by simultaneous nitrification and denitrification. Soil Biol. Biochem. 1996, 28, 609–616. [Google Scholar] [CrossRef]
- Paul, E.A.; Clark, F.E. Soil Microbiology and Biochemistry; Academic Press: Cambridge, MA, USA, 1989. [Google Scholar]
- Russow, R.; Stange, C.F.; Neue, H.U. Role of nitrite and nitric oxide in the processes of nitrification and denitrification in soil: Results from 15N tracer experiments. Soil Biol. Biochem. 2009, 41, 785–795. [Google Scholar] [CrossRef]
- Bremner, J.M.; Keeney, D.R. Determination and Isotope-Ratio Analysis of Different Forms of Nitrogen in Soils: 3. Exchangeable Ammonium, Nitrate, and Nitrite by Extraction-Distillation Methods. Soil Sci. Soc. Am. J. 1966, 30, 577–582. [Google Scholar] [CrossRef]
- Krol, D.J.; Carolan, R.; Minet, E.; McGeough, K.L.; Watson, C.J.; Forrestal, P.J.; Lanigan, G.J.; Richards, K.G. Improving and disaggregating N2O emission factors for ruminant excreta on temperate pasture soils. Sci. Total Environ. 2016, 568, 327–338. [Google Scholar] [CrossRef] [PubMed]
- Clough, T.J.; Sherlock, R.R.; Mautner, M.N.; Milligan, D.B.; Wilson, P.F.; Freeman, C.G.; McEwan, M.J. Emission of nitrogen oxides and ammonia from varying rates of applied synthetic urine and correlations with soil chemistry. Soil Res. 2003, 41, 421–438. [Google Scholar] [CrossRef]
- Linn, D.M.; Doran, J.W. Effect of Water-Filled Pore Space on Carbon Dioxide and Nitrous Oxide Production in Tilled and Nontilled Soils. Soil Sci. Soc. Am. J. 1984, 48, 1267–1272. [Google Scholar] [CrossRef]
- Ambus, P.; Petersen, S.O.; Soussana, J.F. Short-term carbon and nitrogen cycling in urine patches assessed by combined carbon-13 and nitrogen-15 labelling. Agric. Ecosyst. Environ. 2007, 121, 84–92. [Google Scholar] [CrossRef]
- Sigurdarson, J.J.; Svane, S.; Karring, H. The molecular processes of urea hydrolysis in relation to ammonia emissions from agriculture. Rev. Environ. Sci. Bio/Technol. 2018, 17, 241–258. [Google Scholar] [CrossRef]
- Marsden, K.A.; Jones, D.L.; Chadwick, D.R. The urine patch diffusional area: An important N2O source? Soil Biol. Biochem. 2016, 92, 161–170. [Google Scholar] [CrossRef]
- Cardenas, L.M.; Misselbrook, T.M.; Hodgson, C.; Donovan, N.; Gilhespy, S.; Smith, K.A.; Dhanoa, M.S.; Chadwick, D. Effect of the application of cattle urine with or without the nitrification inhibitor DCD, and dung on greenhouse gas emissions from a UK grassland soil. Agric. Ecosyst. Environ. 2016, 235, 229–241. [Google Scholar] [CrossRef]
- van der Weerden, T.J.; Rutherford, A.J.; de Klein, C.A.M.; Ganasamurthy, S.; Morales, S.E. Elevating soil pH does not reduce N2O emissions from urine deposited onto pastoral soils. N. Z. J. Agric. Res. 2021, 65, 484–506. [Google Scholar] [CrossRef]
- Barneze, A.S.; Minet, E.P.; Cerri, C.C.; Misselbrook, T. The effect of nitrification inhibitors on nitrous oxide emissions from cattle urine depositions to grassland under summer conditions in the UK. Chemosphere 2015, 119, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Wrage, N.; Demirbağ, N.S.; Hofmann, M.; Isselstein, J. Influence of stocking density on plant species richness and diversity in permanent grassland. In Grassland Farming and Land Management Systems in Mountainous Regions. Proceedings of the 16th Symposium of the European Grassland Federation, Gumpenstein, Austria, 29–31 August 2011; Agricultural Research and Education Center (AREC) Raumberg-Gumpenstein: Gumpenstein, Austria, 2011. [Google Scholar]
- Malhi, S.S.; McGill, W.B. Nitrification in three Alberta soils: Effect of temperature, moisture and substrate concentration. Soil. Biol. Biochem. 1982, 14, 393–399. [Google Scholar] [CrossRef]
- Selbie, D.R.; Cameron, K.C.; Di, H.J.; Moir, J.L.; Lanigan, G.J.; Richards, K.G. The effect of urinary nitrogen loading rate and a nitrification inhibitor on nitrous oxide emissions from a temperate grassland soil. J. Agric. Sci. 2014, 152, 159–171. [Google Scholar] [CrossRef]
- Clough, T.J.; Cardenas, L.M.; Friedl, J.; Wolf, B. Nitrous oxide emissions from ruminant urine: Science and mitigation for intensively managed perennial pastures. Curr. Opin. Environ. Sustain. 2020, 47, 21–27. [Google Scholar] [CrossRef]
- Parkin, T.B.; Doran, J.W.; Franco-Vizcaíno, E. Field and Laboratory Tests of Soil Respiration. Methods Assess. Soil Qual. 1997, 49, 231–245. [Google Scholar]


{kind=link}
| Property | Units | Soil |
|---|---|---|
| Soil type | Silandic Andosol; Eutric, Siltic (IUSS Working Group 2006) | |
| Texture | Silty clay loam—silt loam (FAO 2006) | |
| pH | 5.55 ± 0.50 | |
| Phosphorus (P) | mg kg−1 dry soil | 26.11 ± 7.14 |
| Potassium (K) | mmol kg−1 dry soil | 3.3 ± 1.6 |
| Magnesium (Mg) | mmol kg−1 dry soil | 5.6 ± 2.4 |
| Calcium (Ca) | mmol kg−1 dry soil | 42.7 ± 22.4 |
| Sodium (Na) | mmol kg−1 dry soil | 1.6 ± 0.4 |
| Aluminium cation (Al) | mmol kg−1 dry soil | 5.4 ± 3.5 |
| Organic matter | g kg−1 dry soil | 168 ± 15 |
| Particle density | g cm−3 | 2.24 |
| Water content at core packing | % w/w | 39 |
| Treatment | CO2 (g C m−2) | N2O (g N m−2) | N2 (g N m−2) | N Loss via N2O (% of Applied N) | N2O/(N2O + N2) |
|---|---|---|---|---|---|
| 0N | 47.41 ± 0.11 a | 0.059 ± 0.029 a | 0.681 ± 0.257 ab | 0.059 ± 0.013 a | |
| 25N | 113.44 ± 1.05 b | 0.100 ± 0.004 a | 0.670 ± 0.325 ab | 0.17 ± 0.02 a | 0.086 ± 0.028 a |
| 50N | 139.06 ± 0.82 c | 0.690 ± 0.091 b | 0.660 ± 0.030 b | 1.39 ± 0.20 c | 0.069 ± 0.006 a |
| 100N | 184.98 ± 2.04 d | 0.855 ± 0.051 b * | 0.268 ± 0.188 a | 0.88 ± 0.06 b * | 0.183 ± 0.069 a |
| Treatment | Before Amendment Application | After Incubation | ||||
|---|---|---|---|---|---|---|
| Parameter | Native Soil | 0N | 25N | 50N | 100N | |
| TON | Top | 0.059 ± 0.002 A | 0.108 ± 0.002 A | 0.605 ± 0.079 B | 1.223 ± 0.027 * C | 1.451 ± 0.078 C |
| g N kg−1 soil | Bottom | 0.059 ± 0.002 a | 0.108 ± 0.001 a | 0.500 ± 0.078 b | 0.873 ± 0.034 * c | 1.167 ± 0.069 d |
| NH4+ | Top | 0.010 ± 0.002 A | 0.004 ± 0.000 A | 0.004 ± 0.000 A | 0.007 ± 0.001 A | 1.016 ± 0.114 * B |
| g N kg−1 soil | Bottom | 0.010 ± 0.002 a | 0.004 ± 0.000 a | 0.004 ± 0.000 a | 0.007 ± 0.000 a | 0.241 ± 0.034 * b |
| SOC | Top | 0.877 ± 0.011 AB | 0.931 ± 0.034 AB | 0.984 ± 0.026 B | 0.798 ± 0.039 A | 0.988 ± 0.019 * B |
| g C kg−1 soil | Bottom | 0.877 ± 0.011 a | 0.874 ± 0.054 a | 0.892 ± 0.023 a | 0.838 ± 0.014 a | 0.791 ± 0.016 * a |
| H2O | Top | 79.32 ± 0.28 A | 83.27 ± 0.44 * A | 80.32 ± 1.78 A | 82.53 ± 0.85 * A | 81.39 ± 1.26 A |
| % | Bottom | 79.32 ± 0.28 a | 79.84 ± 0.51 * a | 79.01 ± 1.29 a | 78.41 ± 0.20 * a | 78.10 ± 0.22 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramírez-Sandoval, M.A.; Loick, N.; Pinochet, D.E.; López-Aizpun, M.; Rivero, M.J.; Cárdenas, L.M. N Losses from an Andisol via Gaseous N2O and N2 Emissions Increase with Increasing Ruminant Urinary–N Deposition Rate. Nitrogen 2024, 5, 254-265. https://doi.org/10.3390/nitrogen5020017
Ramírez-Sandoval MA, Loick N, Pinochet DE, López-Aizpun M, Rivero MJ, Cárdenas LM. N Losses from an Andisol via Gaseous N2O and N2 Emissions Increase with Increasing Ruminant Urinary–N Deposition Rate. Nitrogen. 2024; 5(2):254-265. https://doi.org/10.3390/nitrogen5020017
Chicago/Turabian StyleRamírez-Sandoval, Magdalena A., Nadine Loick, Dante E. Pinochet, Maria López-Aizpun, M. Jordana Rivero, and Laura M. Cárdenas. 2024. "N Losses from an Andisol via Gaseous N2O and N2 Emissions Increase with Increasing Ruminant Urinary–N Deposition Rate" Nitrogen 5, no. 2: 254-265. https://doi.org/10.3390/nitrogen5020017
APA StyleRamírez-Sandoval, M. A., Loick, N., Pinochet, D. E., López-Aizpun, M., Rivero, M. J., & Cárdenas, L. M. (2024). N Losses from an Andisol via Gaseous N2O and N2 Emissions Increase with Increasing Ruminant Urinary–N Deposition Rate. Nitrogen, 5(2), 254-265. https://doi.org/10.3390/nitrogen5020017