
Uncovering the Unusual Long Chains of Vegetative Cells within Single Colonies of the Dryland Nitrogen-Fixing Cyanobacterium Nostoc flagelliforme


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Microscopic Observation
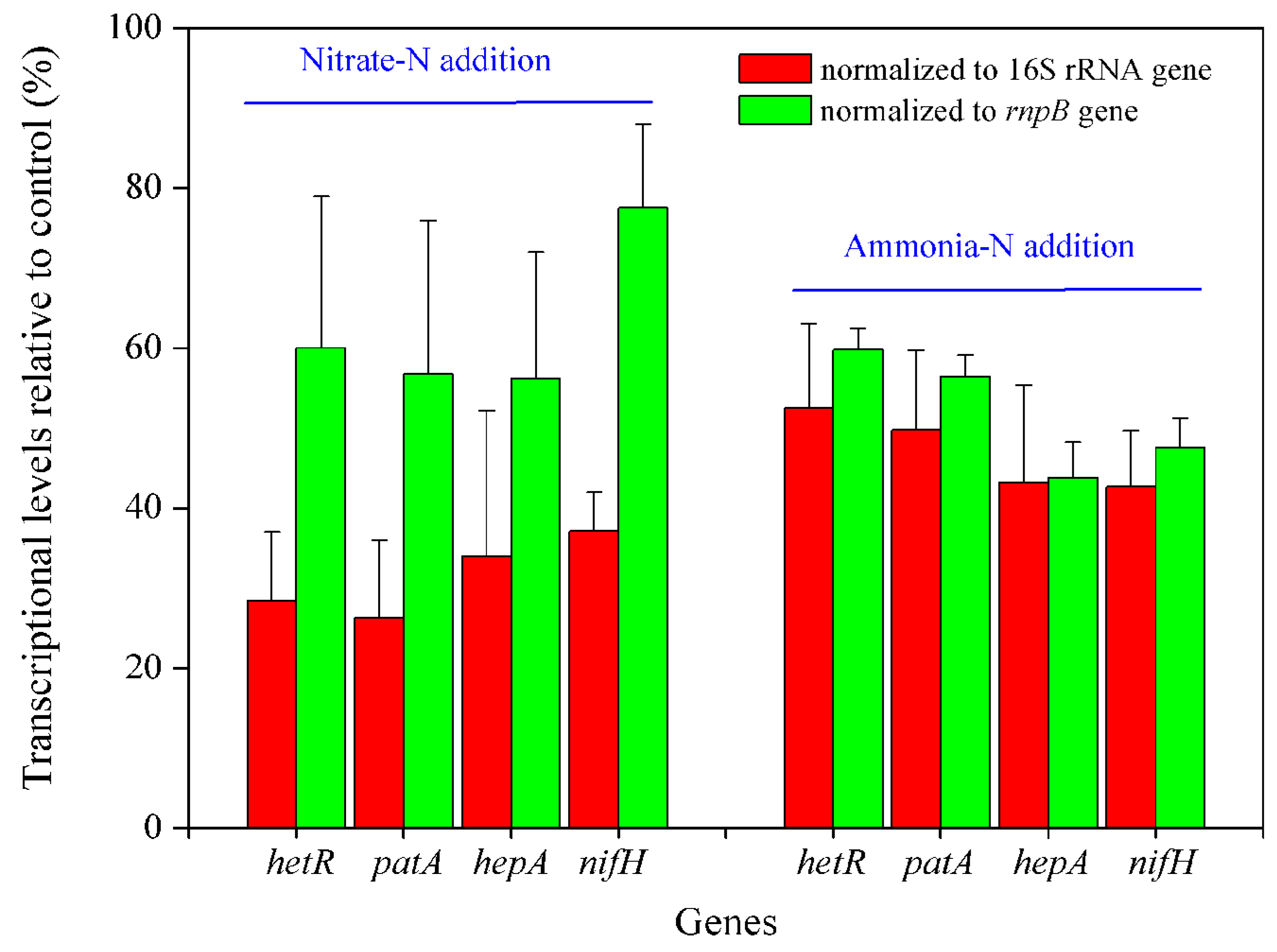
2.2. Transcriptional Analysis
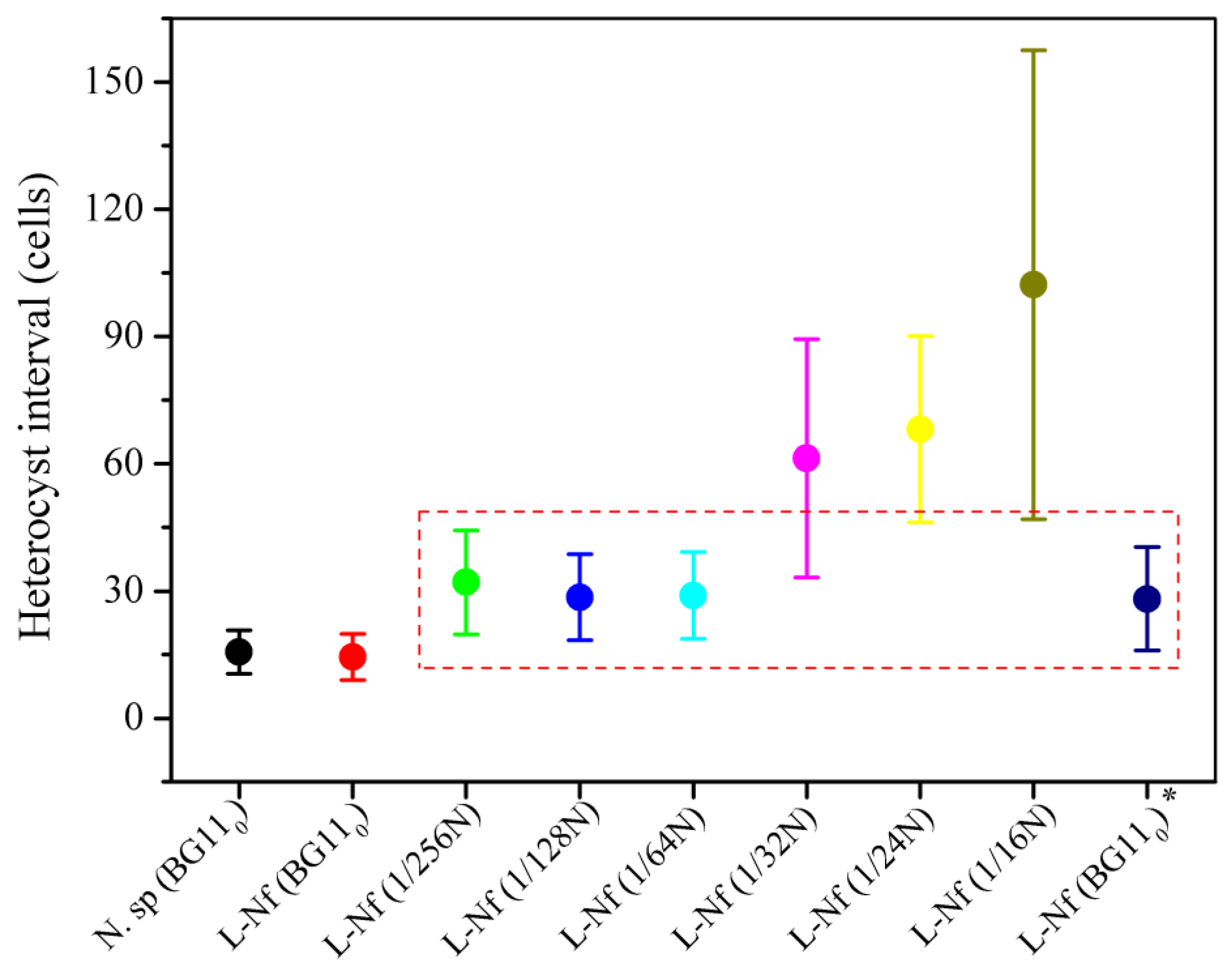
2.3. Testing of Heterocyst Frequency upon Nitrogen Availability
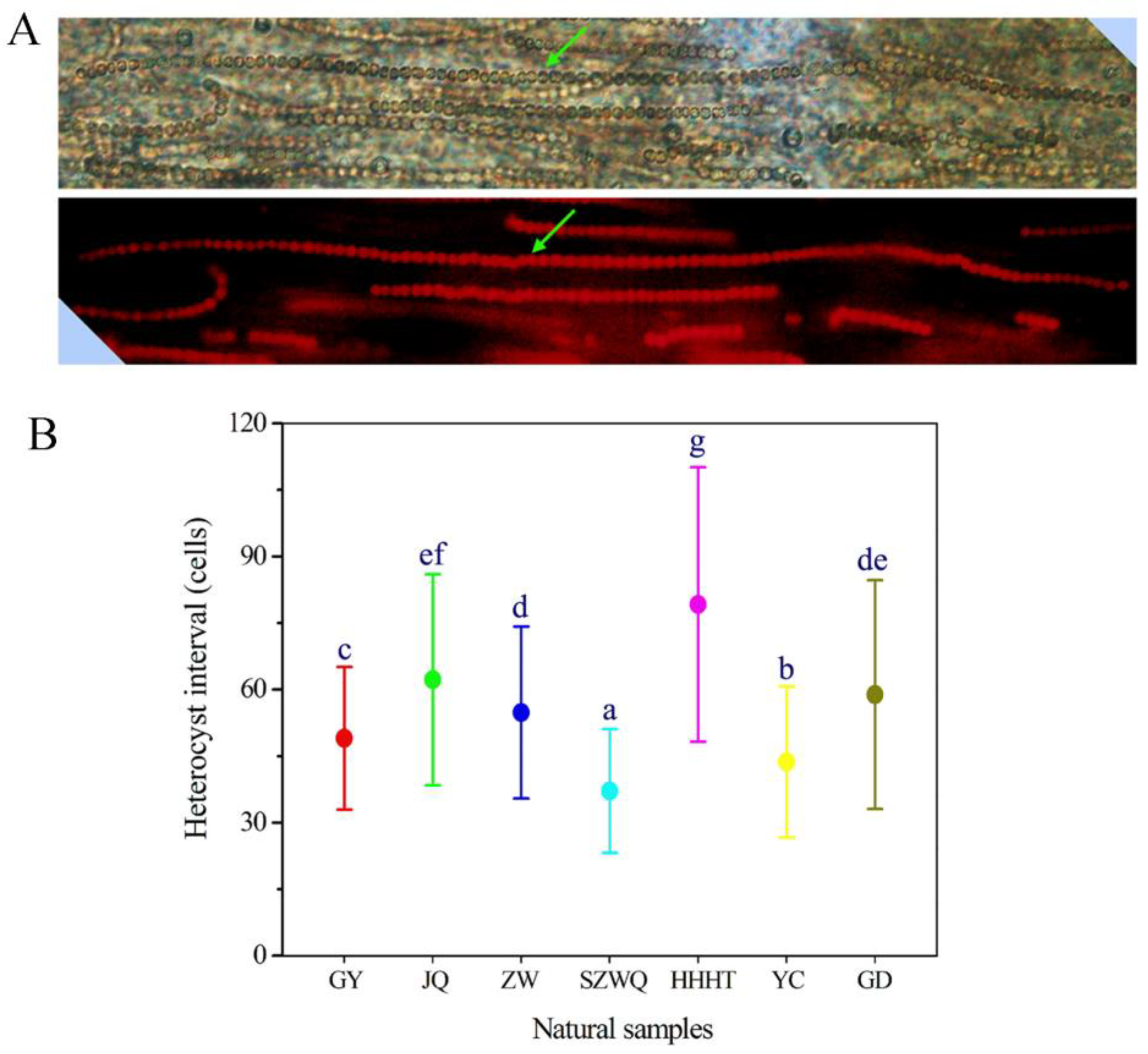
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Su, Y.G.; Zhao, X.; Li, A.X.; Li, X.R.; Huang, G. Nitrogen fixation in biological soil crusts from the Tengger desert, northern China. Eur. J. Soil Biol. 2011, 47, 182–187. [Google Scholar] [CrossRef]
- Rossi, F.; Li, H.; Liu, Y.D.; De Philippis, R. Cyanobacterial inoculation (cyanobacterisation): Perspectives for the development of a standardized multifunctional technology for soil fertilization and desertification reversal. Earth Sci. Rev. 2017, 171, 28–43. [Google Scholar] [CrossRef]
- Rossi, F.; Mugnai, G.; De Philippis, R. Cyanobacterial biocrust induction: A comprehensive review on a soil rehabilitation-effective biotechnology. Geoderma 2022, 415, 115766. [Google Scholar] [CrossRef]
- Baldauf, S.; Cantón, Y.; Tietjen, B. Biocrusts intensify water redistribution and improve water availability to dryland vegetation: Insights from a spatially-explicit ecohydrological model. Front. Microbiol. 2023, 14, 1179291. [Google Scholar] [CrossRef]
- Scherer, S.; Zhong, Z.P. Desiccation independence of terrestrial Nostoc commune ecotypes (cyanobacteria). Microb. Ecol. 1991, 22, 271–283. [Google Scholar] [CrossRef]
- Aboal, M.; Werner, O.; García-Fernández, M.E.; Palazón, J.A.; Cristóbal, J.C.; Williams, W. Should ecomorphs be conserved? The case of Nostoc flagelliforme, an endangered extremophile cyanobacteria. J. Nat. Conserv. 2016, 30, 52–64. [Google Scholar] [CrossRef][Green Version]
- Shang, J.L.; Chen, M.; Hou, S.W.; Li, T.; Yang, Y.W.; Li, Q.; Jiang, H.B.; Dai, G.Z.; Zhang, Z.C.; Hess, W.R.; et al. Genomic and transcriptomic insights into the survival of the subaerial cyanobacterium Nostoc flagelliforme in arid and exposed habitats. Environ. Microbiol. 2019, 21, 845–863. [Google Scholar] [CrossRef]
- Xu, H.F.; Dai, G.Z.; Bai, Y.; Shang, J.L.; Zheng, B.; Ye, D.M.; Shi, H.Z.; Kaplan, A.; Qiu, B.S. Coevolution of tandemly repeated hlips and RpaB-like transcriptional factor confers desiccation tolerance to subaerial Nostoc species. Proc. Natl. Acad. Sci. USA 2022, 119, e2211244119. [Google Scholar] [CrossRef]
- Tamaru, Y.; Takani, Y.; Yoshida, T.; Sakamoto, T. Crucial role of extracellular polysaccharides in desiccation and freezing tolerance in the terrestrial cyanobacterium Nostoc commune. Appl. Environ. Microbiol. 2005, 71, 7327–7333. [Google Scholar] [CrossRef]
- Gao, X. Scytonemin plays a potential role in stabilizing the exopolysaccharidic matrix in terrestrial cyanobacteria. Microb. Ecol. 2017, 73, 255–258. [Google Scholar] [CrossRef]
- Liu, W.; Cui, L.J.; Xu, H.Y.; Zhu, Z.X.; Gao, X. Flexibility-rigidity coordination of the dense exopolysaccharide matrix in terrestrial cyanobacteria acclimated to periodic desiccation. Appl. Environ. Microbiol. 2017, 83, e01619-17. [Google Scholar] [CrossRef]
- Gao, X.; Xu, H.Y.; Yuan, X.L. The overlooked genetic diversity in the dryland soil surface-dwelling cyanobacterium Nostoc flagelliforme as revealed by the marker gene wspA. Microb. Ecol. 2021, 81, 828–831. [Google Scholar] [CrossRef]
- Ferroni, L.; Klisch, M.; Pancaldi, S.; Häder, D.-P. Complementary UV-absorption of mycosporine-like amino acids and scytonemin is responsible for the UV-Insensitivity of photosynthesis in Nostoc flagelliforme. Mar. Drugs 2010, 8, 106–121. [Google Scholar] [CrossRef]
- Arima, H.; Horiguchi, N.; Takaichi, S.; Kofuji, R.; Ishida, K.-I.; Wada, K.; Sakamoto, T. Molecular genetic and chemotaxonomic characterization of the terrestrial cyanobacterium Nostoc commune and its neighboring species. FEMS Microbiol. Ecol. 2011, 79, 34–45. [Google Scholar] [CrossRef]
- Shaw, E.; Hill, D.R.; Brittain, N.; Wright, D.J.; Tauber, U.; Marand, H.; Helm, R.F.; Potts, M. Unusual water flux in the extracellular polysaccharide of the cyanobacterium Nostoc commune. Appl. Environ. Microbiol. 2003, 69, 5679–5684. [Google Scholar] [CrossRef]
- Golden, J.W.; Yoon, H.-S. Heterocyst development in Anabaena. Curr. Opin. Microbiol. 2003, 6, 557–563. [Google Scholar] [CrossRef]
- Zeng, X.L.; Zhang, C.C. The making of a heterocyst in cyanobacteria Annu. Rev. Microbiol. 2022, 76, 597–618. [Google Scholar] [CrossRef]
- Meeks, J.C.; Campbell, E.L.; Summers, M.L.; Wong, F.C. Cellular differentiation in the cyanobacterium Nostoc punctiforme. Arch. Microbiol. 2002, 178, 395–403. [Google Scholar] [CrossRef]
- Zhang, C.C.; Laurent, S.; Sakr, S.; Peng, L.; Bedu, S. Heterocyst differentiation and pattern formation in cyanobacteria: A chorus of signals. Mol. Microbiol. 2006, 59, 367–375. [Google Scholar] [CrossRef]
- Gao, K. Chinese studies on the edible blue-green alga, Nostoc flagelliforme: A review. J. Appl. Phycol. 1998, 10, 37–49. [Google Scholar] [CrossRef]
- Stanier, R.Y.; Kunisawa, R.; Mandel, M.; Cohen-Bazire, G. Purification and properties of unicellular blue-green algae (order Chroococcales). Bacteriol. Rev. 1971, 35, 171–205. [Google Scholar] [CrossRef]
- Gao, K.; Ye, C. Culture of the terrestrial cyanobacterium, Nostoc flagelliforme (Cyanophyceae), under aquatic conditions. J. Phycol. 2003, 39, 617–623. [Google Scholar] [CrossRef]
- Flores, E.; Herrero, A. Nitrogen assimilation and nitrogen control in cyanobacteria. Biochem. Soc. Trans. 2005, 33, 164–167. [Google Scholar] [CrossRef]
- Glibert, P.M.; Bronk, D.A. Release of dissolved organic nitrogen by marine diazotrophic cyanobacteria, Trichodesmium spp. Appl. Environ. Microbiol. 1994, 60, 3996–4000. [Google Scholar] [CrossRef]
- Hill, D.R.; Hladun, S.L.; Scherer, S.; Potts, M. Water stress proteins of Nostoc commune (Cyanobacteria) are secreted with UV-A/B-absorbing pigments and associate with 1,4-beta-D-xylanxylanohydrolase activity. J. Biol. Chem. 1994, 269, 7726–7734. [Google Scholar] [CrossRef]
- Subramanian, G.; Shanmugasundaram, S. Uninduced ammonia release by the nitrogen-fixing cyanobacterium Anabaena. FEMS Microbiol. Lett. 1986, 37, 151–154. [Google Scholar] [CrossRef][Green Version]
- Stuart, R.K.; Mayali, X.; Lee, J.Z.; Craig Everroad, R.; Hwang, M.; Bebout, B.M.; Weber, P.K.; Pett-Ridge, J.; Thelen, M.P. Cyanobacterial reuse of extracellular organic carbon in microbial mats. ISME J. 2016, 10, 1240–1251. [Google Scholar] [CrossRef]
- Oren, N.; Raanan, H.; Murik, O.; Keren, N.; Kaplan, A. Dawn illumination prepares desert cyanobacteria for dehydration. Curr. Biol. 2017, 27, R1056–R1057. [Google Scholar] [CrossRef]
- Xu, H.F.; Dai, G.Z.; Qiu, B.S. Weak red light plays an important role in awakening the photosynthetic machinery following desiccation in the subaerial cyanobacterium Nostoc flagelliforme. Environ. Microbiol. 2019, 21, 2261–2272. [Google Scholar] [CrossRef]
- Gao, X.; Liu, L.T.; Liu, B. Dryland cyanobacterial exopolysaccharides show protection against acid deposition damage. Environ. Sci. Pollut. Res. 2019, 26, 24300–24304. [Google Scholar] [CrossRef]
- Brauer, V.S.; Stomp, M.; Rosso, C.; van Beusekom, S.A.M.; Emmerich, B.; Stal, L.J.; Huisman, J. Low temperature delays timing and enhances the cost of nitrogen fixation in the unicellular cyanobacterium Cyanothece. ISME J. 2013, 7, 2105–2115. [Google Scholar] [CrossRef]
- Zapomelová, E.; Hrouzek, P.; Reháková, K.; Sabacká, M.; Stibal, M.; Caisová, L.; Komárková, J.; Lukesová, A. Morphological variability in selected heterocystous cyanobacterial strains as a response to varied temperature, light intensity and medium composition. Folia Microbiol. 2008, 53, 333–341. [Google Scholar] [CrossRef]
- Gollan, P.J.; Muth-Pawlak, D.; Aro, E.-M. Rapid transcriptional reprogramming triggered by alteration of the carbon/nitrogen balance has an impact on energy metabolism in Nostoc sp. PCC 7120. Life 2020, 10, 297. [Google Scholar] [CrossRef]
- Schwabedissen, S.G.; Lohse, K.A.; Reed, S.C.; Aho, K.A.; Magnuson, T.S. Nitrogenase activity by biological soil crusts in cold sagebrush steppe ecosystems. Biogeochemistry 2017, 134, 57–76. [Google Scholar] [CrossRef]
- Qi, J.; Li, X.; Jia, R.; Yang, H.; Shi, Y.; Sun, J.; Fu, T. Response of biocrust nitrogenase activity to the variation of rainfall regime in the Tengger Desert, northern China. CATENA 2022, 212, 106093. [Google Scholar] [CrossRef]
- Zhang, Q.; Li, Y.; Wang, M.; Wang, K.; Meng, F.; Liu, L.; Zhao, Y.; Ma, L.; Zhu, Q.; Xu, W.; et al. Atmospheric nitrogen deposition: A review of quantification methods and its spatial pattern derived from the global monitoring networks. Ecotox. Environ. Saf. 2021, 216, 112180. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, X.; Liu, C.; Liang, W. Uncovering the Unusual Long Chains of Vegetative Cells within Single Colonies of the Dryland Nitrogen-Fixing Cyanobacterium Nostoc flagelliforme. Nitrogen 2024, 5, 144-151. https://doi.org/10.3390/nitrogen5010009
Gao X, Liu C, Liang W. Uncovering the Unusual Long Chains of Vegetative Cells within Single Colonies of the Dryland Nitrogen-Fixing Cyanobacterium Nostoc flagelliforme. Nitrogen. 2024; 5(1):144-151. https://doi.org/10.3390/nitrogen5010009
Chicago/Turabian StyleGao, Xiang, Chang Liu, and Wensheng Liang. 2024. "Uncovering the Unusual Long Chains of Vegetative Cells within Single Colonies of the Dryland Nitrogen-Fixing Cyanobacterium Nostoc flagelliforme" Nitrogen 5, no. 1: 144-151. https://doi.org/10.3390/nitrogen5010009
APA StyleGao, X., Liu, C., & Liang, W. (2024). Uncovering the Unusual Long Chains of Vegetative Cells within Single Colonies of the Dryland Nitrogen-Fixing Cyanobacterium Nostoc flagelliforme. Nitrogen, 5(1), 144-151. https://doi.org/10.3390/nitrogen5010009