
Incident Light Level Influences Elemental Budgets of Serianthes nelsonii Merr. Leaf Development


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
3. Results
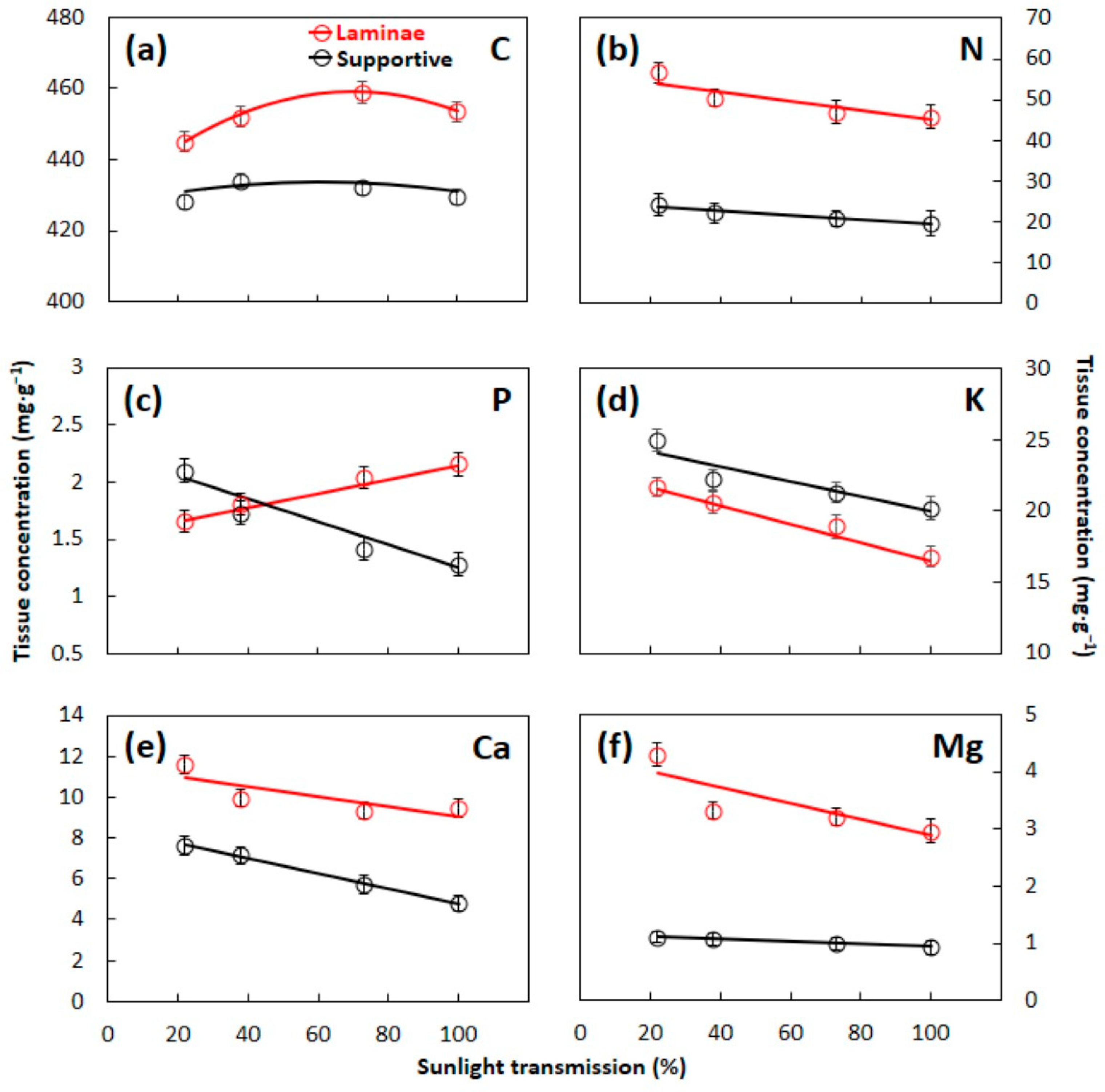
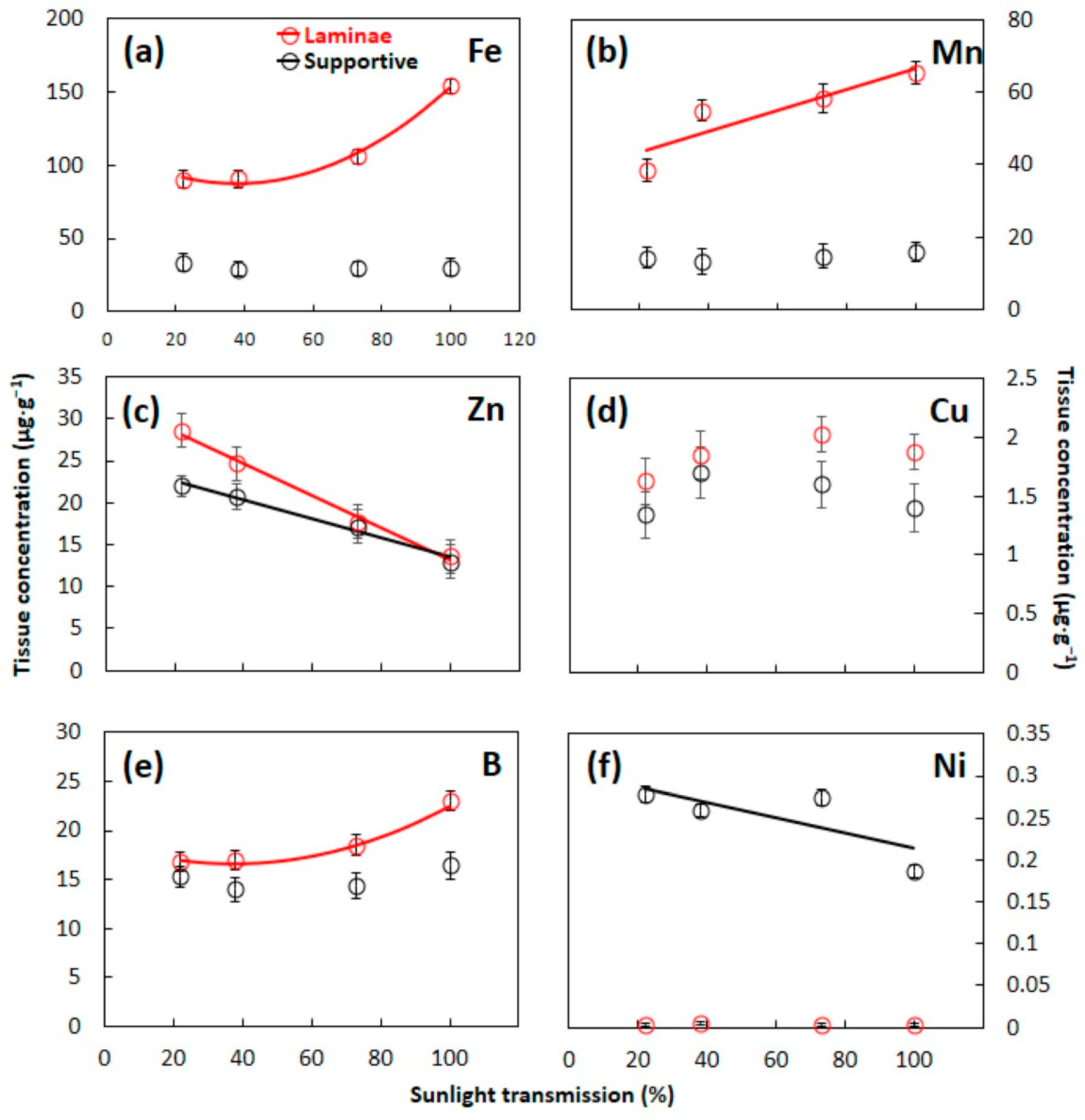
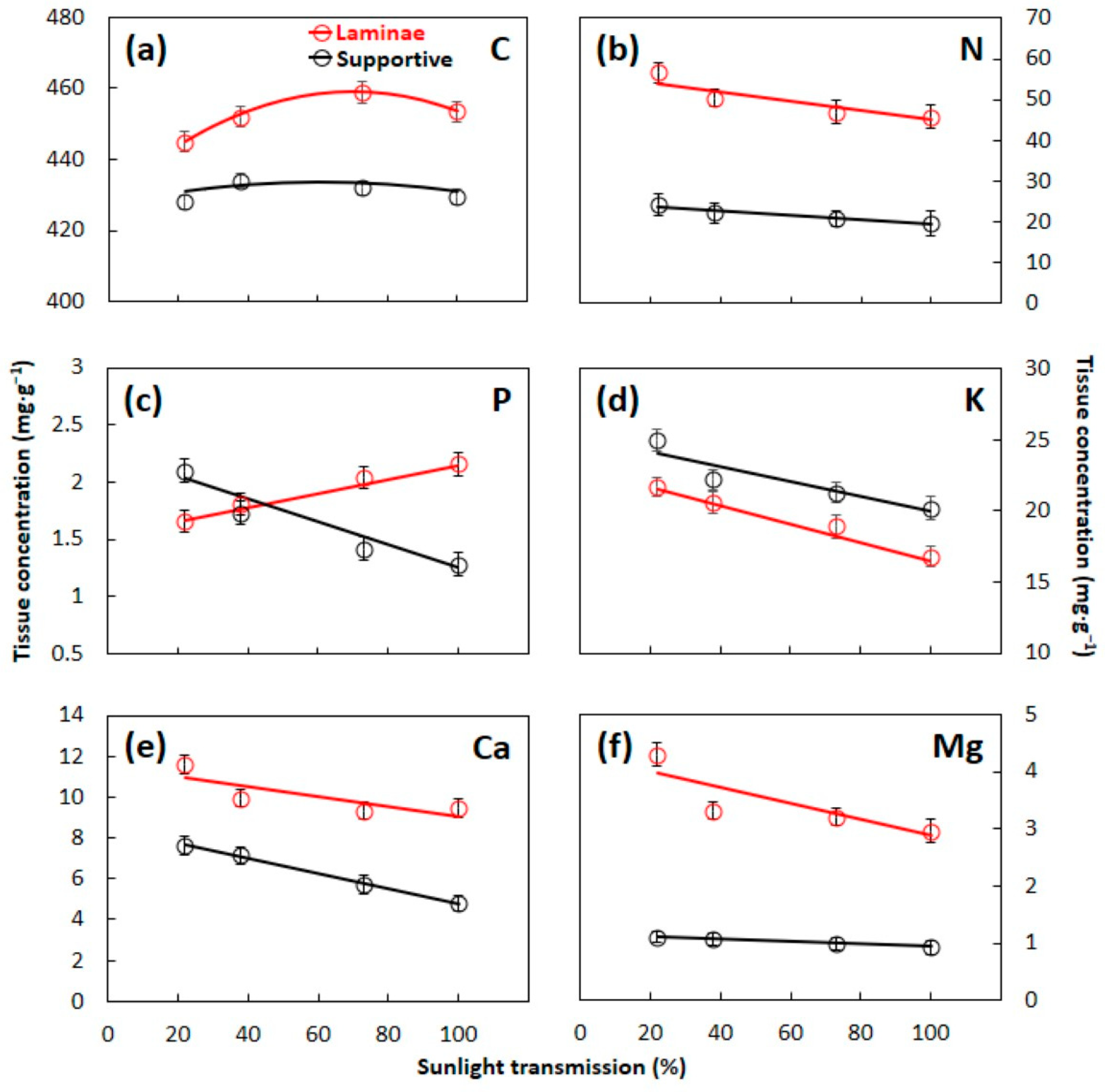
3.1. Essential Element Concentrations
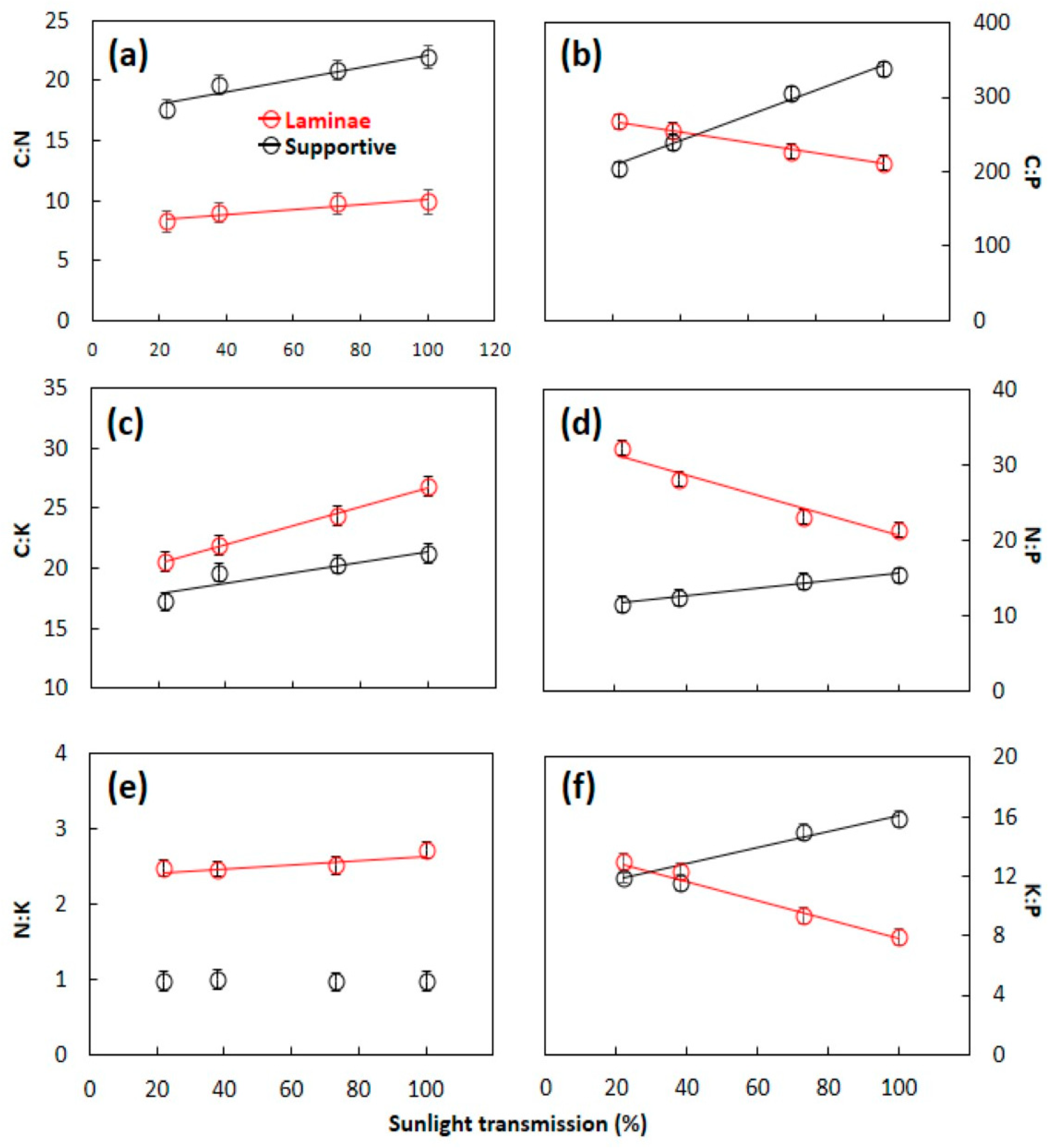
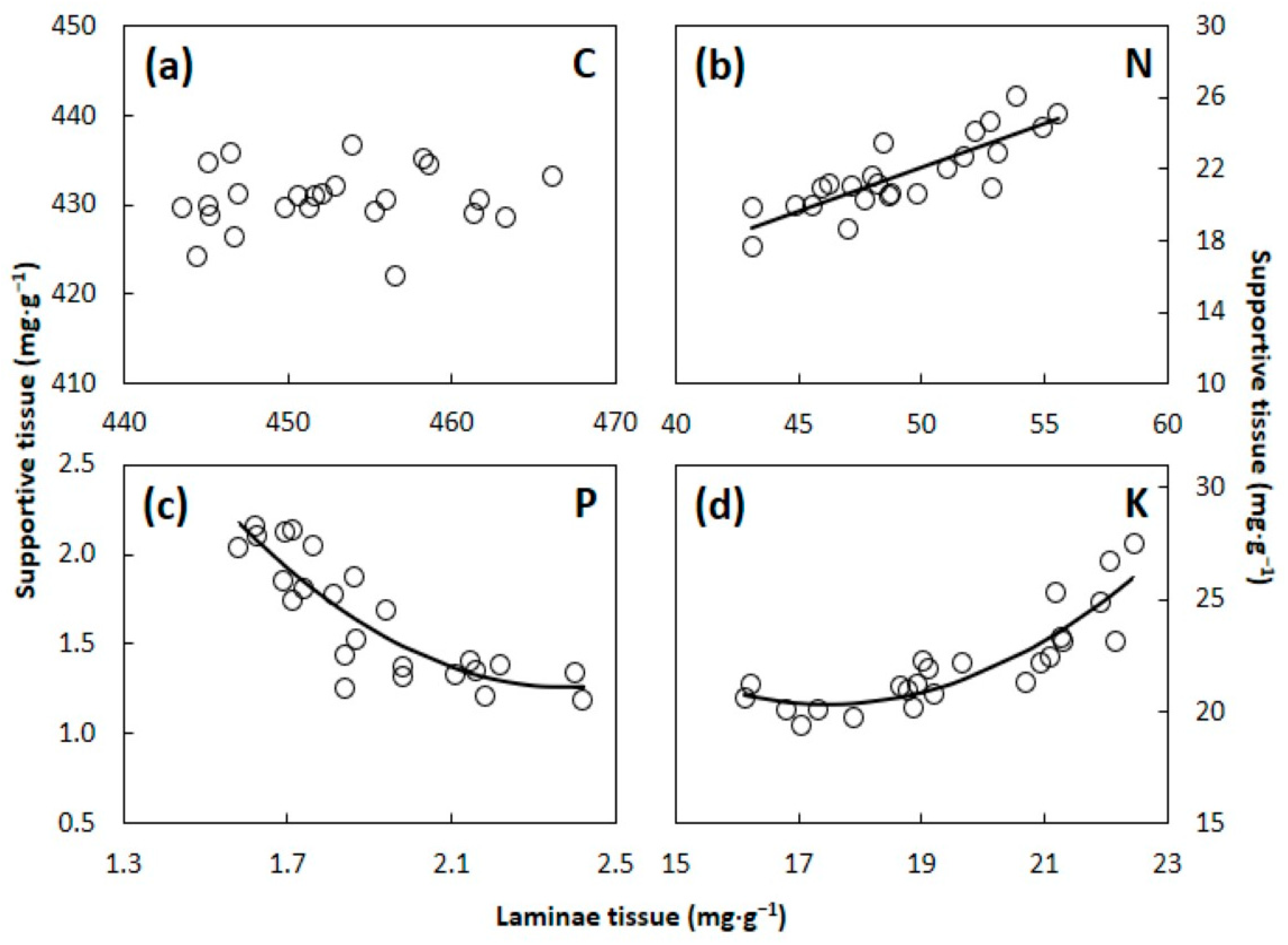
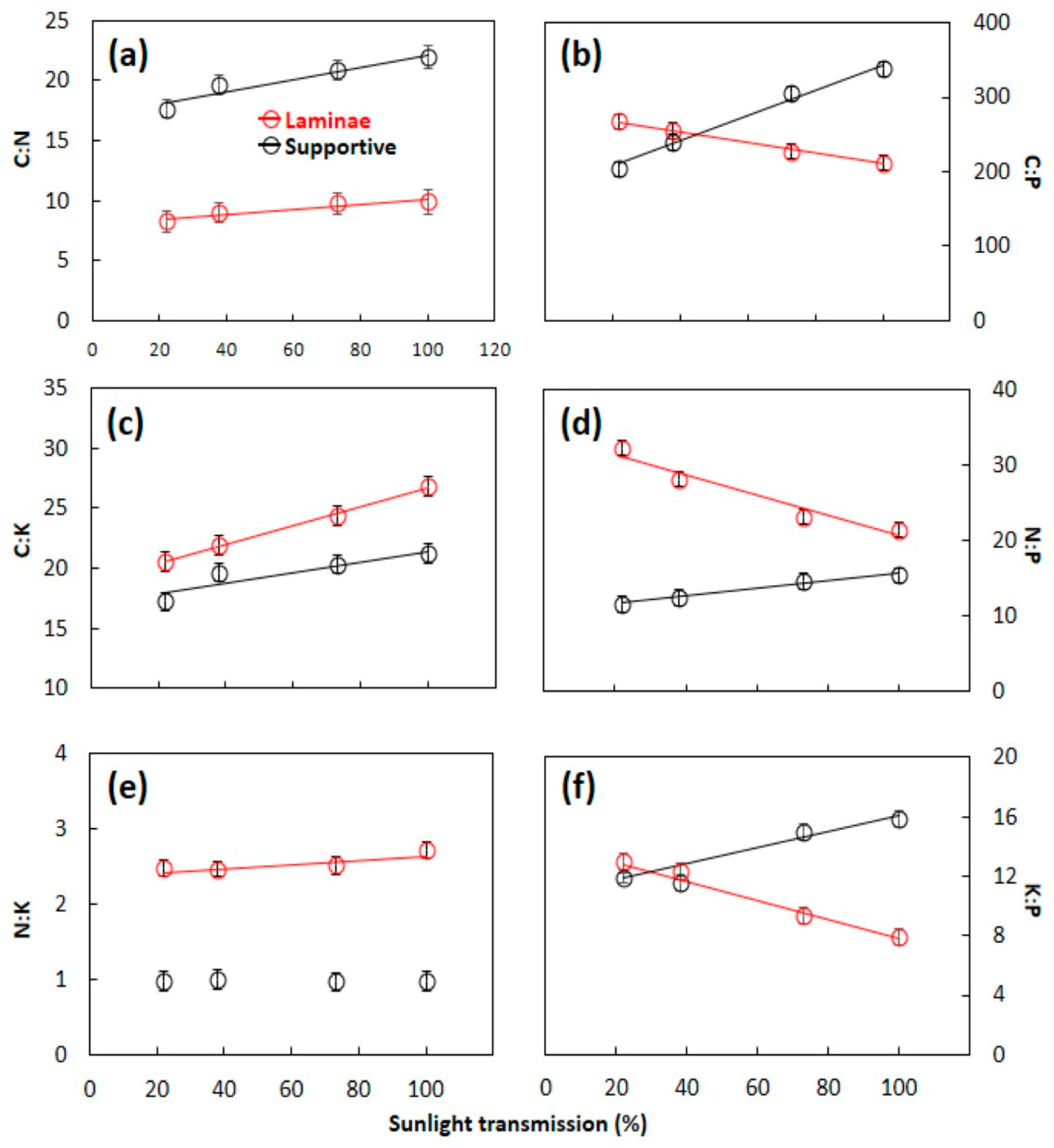
3.2. C:N:P:K Stoichometry
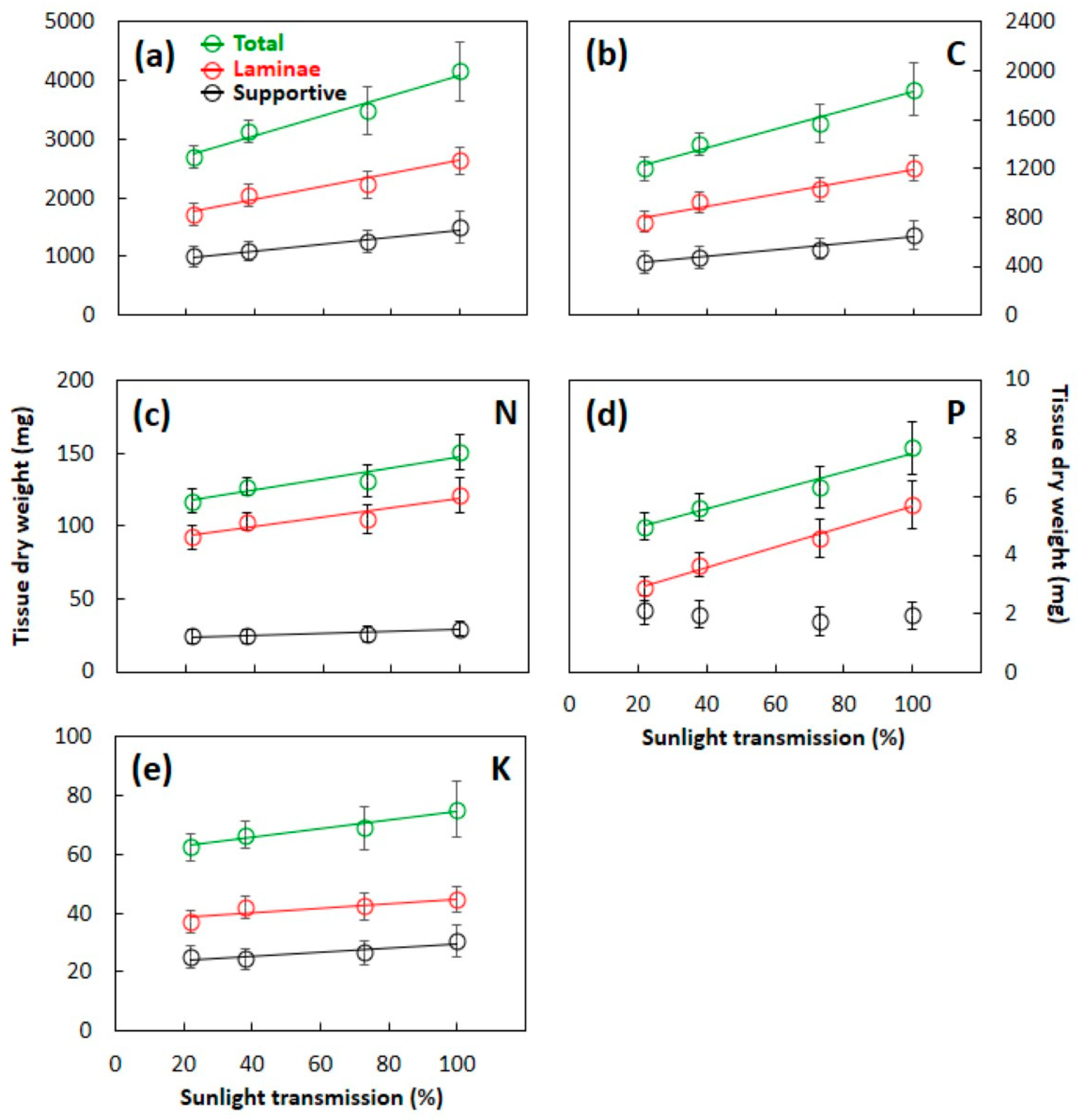
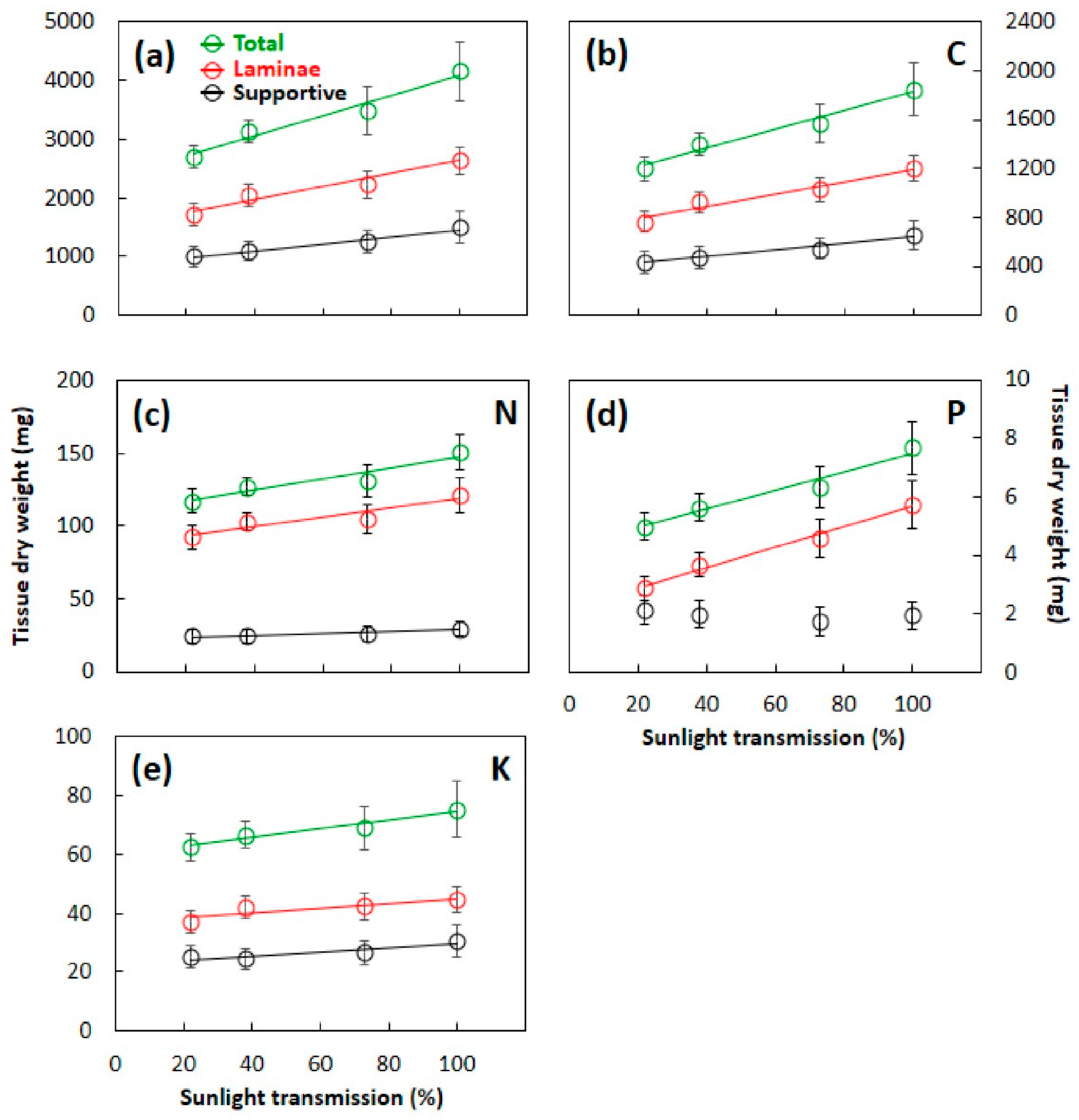
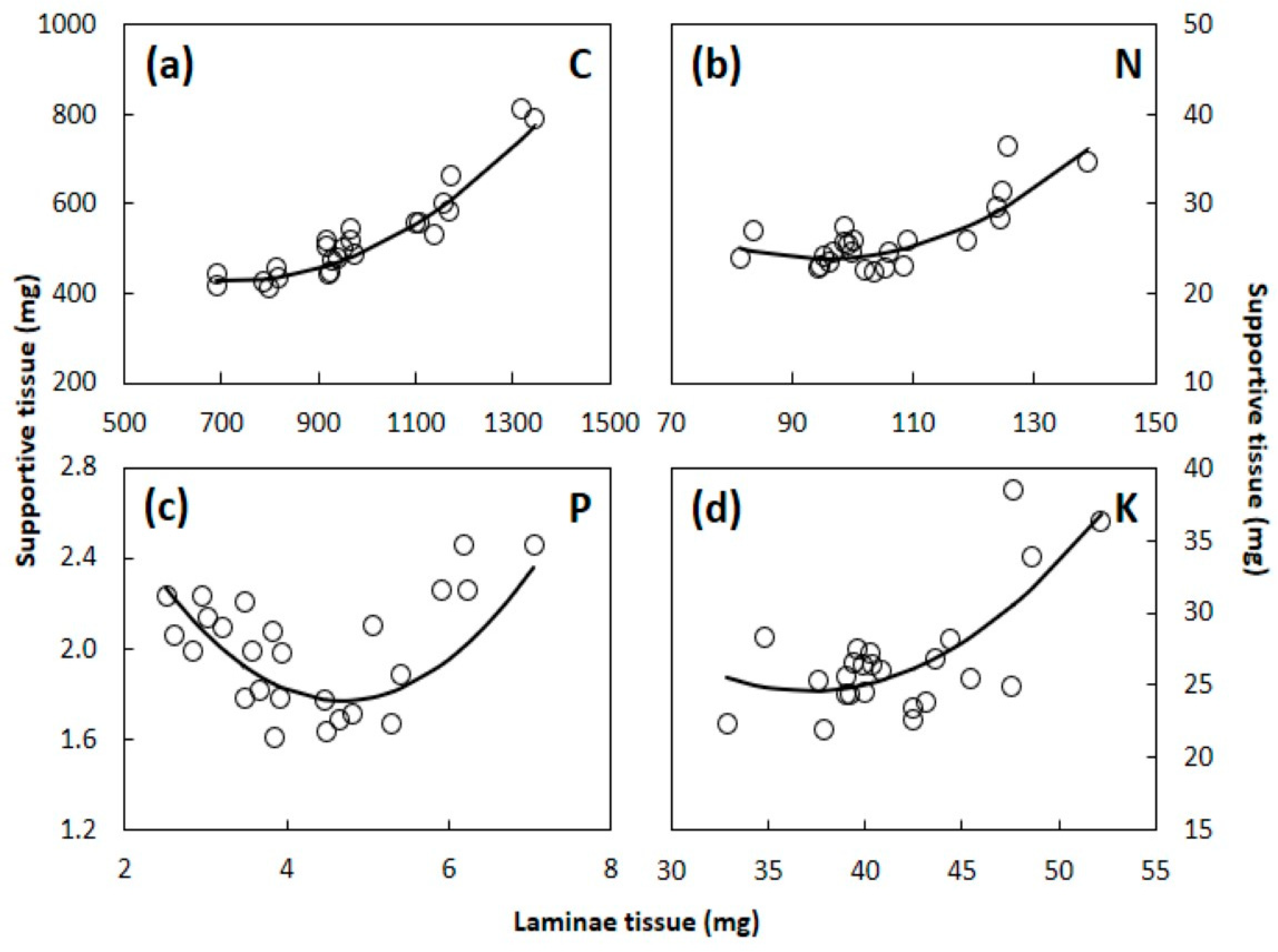
3.3. Total Leaf Mass and Element Content
4. Discussion
4.1. Leaf Construction Needs
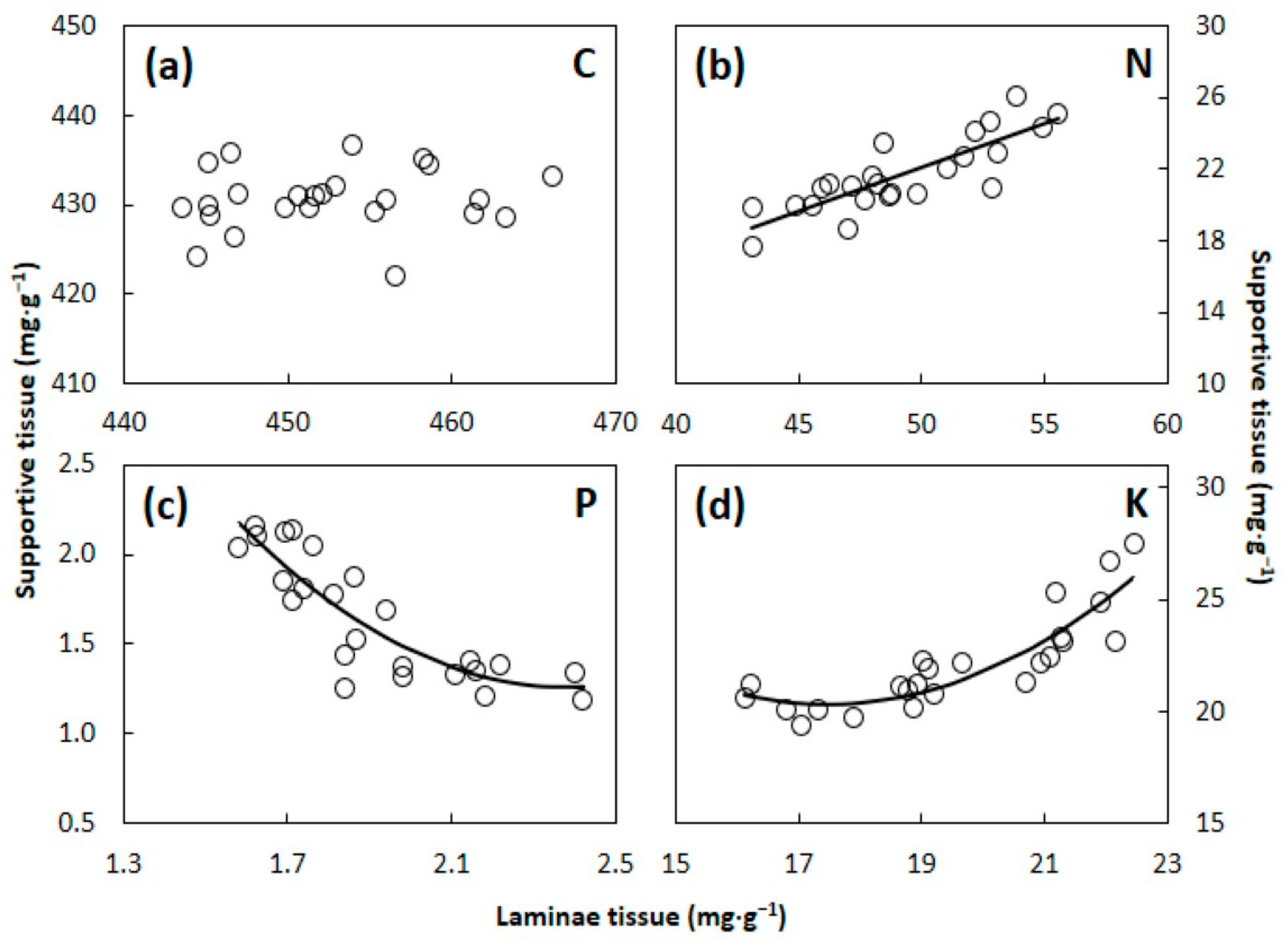
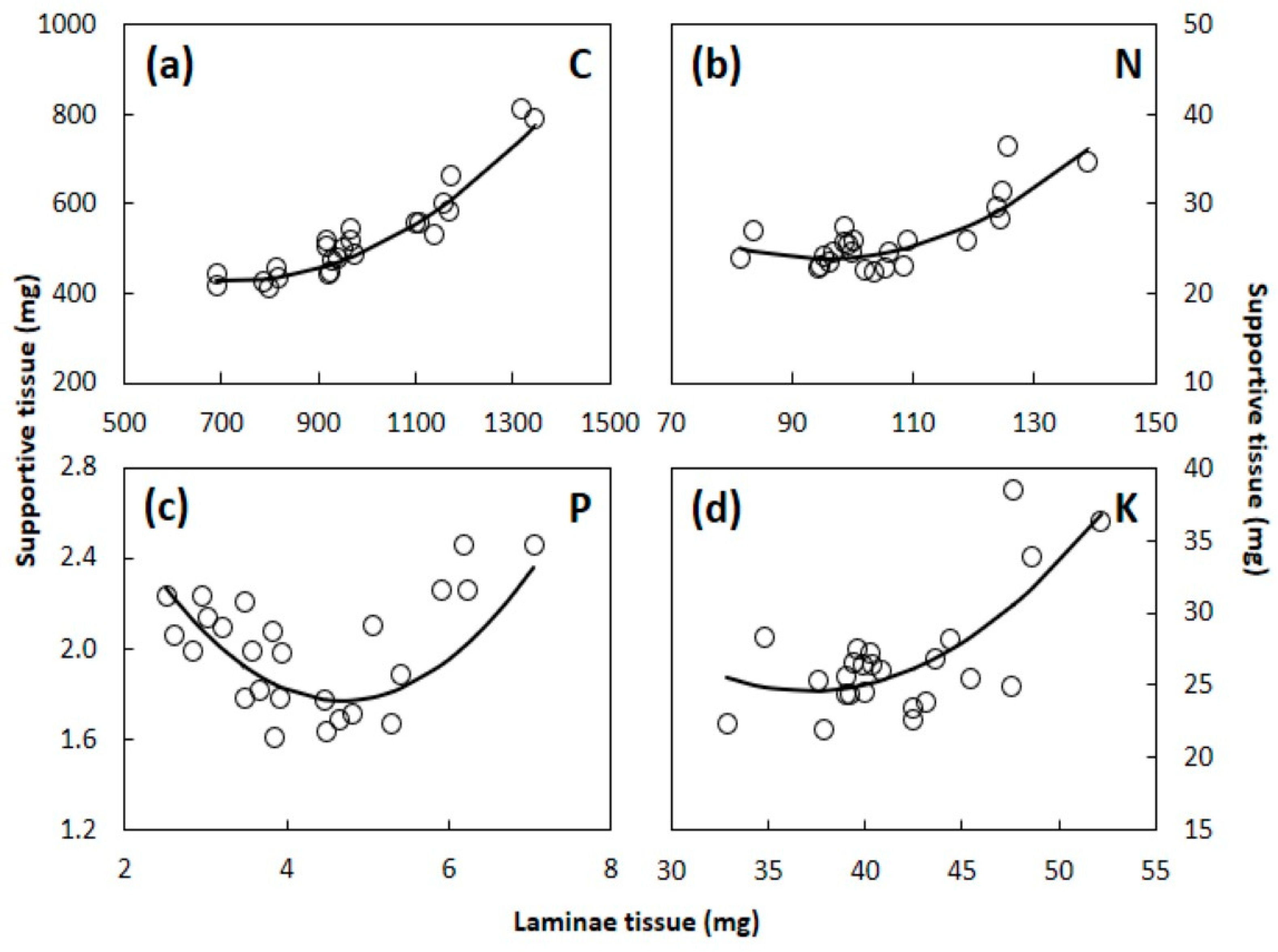
4.2. Laminae versus Supportive Tissue
4.3. Previously Published Lamina Data
4.4. Conservation Implications
4.5. Future Directions
5. Conclusions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
- Figure 4a: NS
- Figure 5e supportive: NS
Appendix B
- Figure 6d supportive: NS
References
- Wiles, G.; Williams, E. Serianthes nelsonii. IUCN Red List Threat. Species 2017, e.T30437A98715973. Available online: https://doi.org/10.2305/IUCN.UK.2017-3.RLTS.T30437A98715973.en (accessed on 15 February 2024).
- United States Fish and Wildlife Service. Determination of endangered status for Serianthes nelsonii Merr. (Hayun lagu or Tronkon Guafi). Fed. Regist. 1987, 52, 4907–4910. [Google Scholar]
- United States Fish and Wildlife Service. Recovery Plan for Serianthes nelsonii; USFWS: Portland, OR, USA, 1994. [Google Scholar]
- Wiles, G.J.; Aguon, C.F.; Davis, G.W.; Grout, D.J. The status and distribution of endangered animals and plants in northern Guam. Micronesica 1995, 28, 31–49. [Google Scholar]
- Demeulenaere, E.; Ickert-Bond, S.M. Guam’s last håyun lågu tree (Serianthes nelsonii) in peril. Conserv. Sci. Practice 2023, 5, e13019. [Google Scholar] [CrossRef]
- Marler, T.E.; Musser, C.; Cascasan, A.N.J.; Cruz, G.N.; Deloso, B.E. Adaptive management lessons for Serianthes nelsonii conservation. Horticulturae 2021, 7, 43. [Google Scholar] [CrossRef]
- Niinemets, Ü. Are compound-leaved woody species inherently shade-intolerant? An analysis of species ecological requirements and foliar support costs. Plant Ecol. 1998, 134, 1–11. [Google Scholar] [CrossRef]
- Deloso, B.E.; Marler, T.E. Bi-pinnate compound Serianthes nelsonii leaf-level plasticity magnifies leaflet-level plasticity. Biology 2020, 9, 333. [Google Scholar] [CrossRef] [PubMed]
- Horn, H.S. The Adaptive Geometry of Trees; Princeton University Press: Princeton, NJ, USA, 1971; p. 144. [Google Scholar]
- Bazzaz, F.A. The physiological ecology of plant succession. Annu. Rev. Ecol. Syst. 1979, 10, 351–371. [Google Scholar] [CrossRef]
- Givnish, T.J. Adaptation to sun and shade: A whole-plant perspective. Aust. J. Plant Physiol. 1988, 15, 63–92. [Google Scholar] [CrossRef]
- Niinemets, Ü.; Valladares, F. Tolerance to shade, drought and waterlogging of temperate northern hemisphere trees and shrubs. Ecol. Monogr. 2006, 76, 521–547. [Google Scholar] [CrossRef]
- Valladares, F.; Niinemets, Ü. Shade tolerance, a key plant feature of complex nature and consequences. Annu. Rev. Ecol. Evol. Syst. 2008, 39, 237–257. [Google Scholar] [CrossRef]
- Demmig-Adams, B.; Adams, W.W., III. Photoprotection in an ecological context: The remarkable complexity of thermal energy dissipation. New Phytol. 2006, 172, 11–21. [Google Scholar] [CrossRef]
- Dumas, J.B.A. Procedes de L’analyse Organique. Ann. Chim. Phys. 1831, 47, 198–205. [Google Scholar]
- Hou, X.; Jones, B.T. Inductively coupled plasma/optical emission spectrometry. In Encyclopedia of Analytical Chemistry; Meyers, R.A., Ed.; John Wiley & Sons: Chichester, UK, 2000; pp. 9468–9485. [Google Scholar]
- Lin, J.; Harcombe, P.A.; Fulton, M.R.; Hall, R.W. Sapling growth and survivorship as a function of light in a mesic forest of southeast Texas, USA. Oecologia 2002, 132, 428–435. [Google Scholar] [CrossRef] [PubMed]
- Wolf, A.A.; Funk, J.L.; Menge, D.N.L. The symbionts made me do it: Legumes are not hardwired for high nitrogen concentrations but incorporate more nitrogen when inoculated. New Phytol. 2017, 213, 690–699. [Google Scholar] [CrossRef]
- Portsmuth, A.; Niinemets, U. Structural and physiological plasticity in response to light and nutrients in five temperate deciduous woody species of contrasting shade tolerance. Funct. Ecol. 2007, 21, 61–77. [Google Scholar] [CrossRef]
- Marler, T.E. Leaf elemental concentrations, stoichiometry, and resorption in Guam’s coastal karst forests. Diversity 2021, 13, 545. [Google Scholar] [CrossRef]
- Niinemets, Ü. Differences in chemical composition relative to functional differentiation between petioles and laminas of Fraxinus excelsior. Tree Physiol. 1999, 19, 39–45. [Google Scholar] [CrossRef]
- Niinemets, Ü.; Kull, O. Biomass investment in leaf lamina versus lamina support in relation to growth irradiance and leaf size in temperate deciduous trees. Tree Physiol. 1999, 19, 349–358. [Google Scholar] [CrossRef]
- Niinemets, Ü.; Portsmuth, A.; Tena, D.; Tobias, M.; Matesanz, S.; Valladares, F. Do we underestimate the importance of leaf size in plant economics? Disproportional scaling of support costs within the spectrum of leaf physiognomy. Annals Bot. 2007, 100, 283–303. [Google Scholar] [CrossRef]
- Niinemets, Ü.; Fleck, S. Petiole mechanics, leaf inclination, morphology, and investment in support in relation to light availability in the canopy of Liriodendron tulipifera. Oecologia 2002, 132, 21–33. [Google Scholar] [CrossRef]
- Sims, D.A.; Gebauer, R.L.E.; Pearcy, R.W. Scaling sun and shade photosynthetic acclimation of Alocasia macrorrhiza to whole-plant performance—II. Simulation of carbon balance and growth at different photon flux densities. Plant Cell Environ. 1994, 17, 889–900. [Google Scholar] [CrossRef]
- Marler, T.E. NPK fertilization of Serianthes plants influences growth and stoichiometry of leaf nutrients. Horticulturae 2022, 8, 717. [Google Scholar] [CrossRef]
- Marler, T.E. Soil from Serianthes rhizosphere influences growth and leaf nutrient content of Serianthes plants. Agronomy 2022, 12, 1938. [Google Scholar] [CrossRef]
- Marler, T.E. Foliar nutrition of Serianthes nelsonii seedlings as a conservation tool. HortScience 2022, 57, 389–390. [Google Scholar] [CrossRef]
- Lawson, S.S.; Michler, C.H. Afforestation, restoration and regeneration—Not all trees are created equal. J. For. Res. 2014, 25, 3–20. [Google Scholar] [CrossRef]
- Löf, M.; Madsen, P.; Metslaid, M.; Witzell, J.; Jacobs, D.F. Restoring forests: Regeneration and ecosystem function for the future. New For. 2019, 50, 139–152. [Google Scholar] [CrossRef]
- Antonelli, A.; Fry, C.; Smith, R.J.; Eden, J.; Govaerts, R.H.A.; Kersey, P.; Nic Lughadha, E.; Onstein, R.E.; Simmonds, M.S.J.; Zizka, A.; et al. State of the World’s Plants and Fungi 2023; Royal Botanic Gardens, Kew: Richmond, UK, 2023. [Google Scholar] [CrossRef]
- Lindenmayer, D.; Laurance, W.; Franklin, J. Global decline in large old trees. Science 2012, 338, 1305–1306. [Google Scholar] [CrossRef] [PubMed]
- Niinemets, Ü.; Kull, O.; Tenhunen, J.D. An analysis of light effects on foliar morphology, physiology, and light interception in temperate deciduous woody species of contrasting shade tolerance. Tree Physiol. 1998, 18, 681–696. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.R.; Poorter, H. Photosynthetic acclimation of plants to growth irradiance: The relative importance of specific leaf area and nitrogen partitioning in maximizing carbon gain. Plant Cell Environ. 2001, 24, 755–767. [Google Scholar] [CrossRef]
- Yu, Z.-C.; Zheng, X.-T.; He, W.; Lin, W.; Yan, G.-Z.; Zhu, H.; Peng, C.-L. Different responses of macro- and microelement contents of 41 subtropical plants to environmental changes in the wet and dry seasons. J. Plant Ecol. 2023, 16, rtad027. [Google Scholar] [CrossRef]
- Lusk, C.H. Leaf area and growth of juvenile temperate evergreens in low light: Species of contrasting shade tolerance change rank during ontogeny. Funct. Ecol. 2004, 18, 820–828. [Google Scholar] [CrossRef]
- Rozendaal, D.M.A.; Hurtado, V.H.; Poorter, L. Plasticity in leaf traits of 38 tropical tree species in response to light; relationships with light demand and adult stature. Funct. Ecol. 2006, 20, 207–216. [Google Scholar] [CrossRef]
- Li, X.-B.; Liu, C.-C.; Chen, J.-X.; Zhang, M.-M.; Zhang, J.-H.; Tong, Z.-K.; Yang, Q. Leaf plasticity of the subtropical evergreen tree Phoebe bournei increases with ontogeny in response to sun and shade. Forests 2023, 14, 1683. [Google Scholar] [CrossRef]
- Barton, K.E. The ontogenetic dimension of plant functional ecology. Funct. Ecol. 2024, 38, 1–16. [Google Scholar] [CrossRef]
- Guo, X.; Schrader, J.; Shi, P.; Jiao, Y.; Miao, Q.; Xue, J.; Niklas, K.J. Leaf-age and petiole biomass play significant roles in leaf scaling theory. Front. Plant Sci. 2023, 14, 1322245. [Google Scholar] [CrossRef]
- Poorter, H.; Niinemets, Ü.; Poorter, L.; Wright, I.J.; Villar, R. Causes and consequences of variation in leaf mass per area (LMA): A meta-analysis. New Phytol. 2009, 182, 565–588. [Google Scholar] [CrossRef] [PubMed]
- Markesteijn, L.; Poorter, L.; Bongers, F. Light-dependent leaf trait variation in 43 tropical dry forest tree species. Am. J. Bot. 2007, 94, 515–525. [Google Scholar] [CrossRef] [PubMed]
- Poorter, H.; Niklas, K.J.; Reich, P.B.; Oleksyn, J.; Poot, P.; Mommer, L. Biomass allocation to leaves, stems and roots: Meta-analyses of interspecific variation and environmental control. New Phytol. 2012, 193, 30–50. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marler, T.E. Incident Light Level Influences Elemental Budgets of Serianthes nelsonii Merr. Leaf Development. Nitrogen 2024, 5, 177-190. https://doi.org/10.3390/nitrogen5010012
Marler TE. Incident Light Level Influences Elemental Budgets of Serianthes nelsonii Merr. Leaf Development. Nitrogen. 2024; 5(1):177-190. https://doi.org/10.3390/nitrogen5010012
Chicago/Turabian StyleMarler, Thomas E. 2024. "Incident Light Level Influences Elemental Budgets of Serianthes nelsonii Merr. Leaf Development" Nitrogen 5, no. 1: 177-190. https://doi.org/10.3390/nitrogen5010012
APA StyleMarler, T. E. (2024). Incident Light Level Influences Elemental Budgets of Serianthes nelsonii Merr. Leaf Development. Nitrogen, 5(1), 177-190. https://doi.org/10.3390/nitrogen5010012