
Biodiversity-Based Empirical Critical Loads of Nitrogen Deposition in the Athabasca Oil Sands Region


Abstract
1. Introduction
2. Methods
2.1. Study Sites and Data Sources
2.2. Site Sampling Methods
2.3. Laboratory Analysis Methods
2.4. Environmental Data
2.5. Statistical Analyses
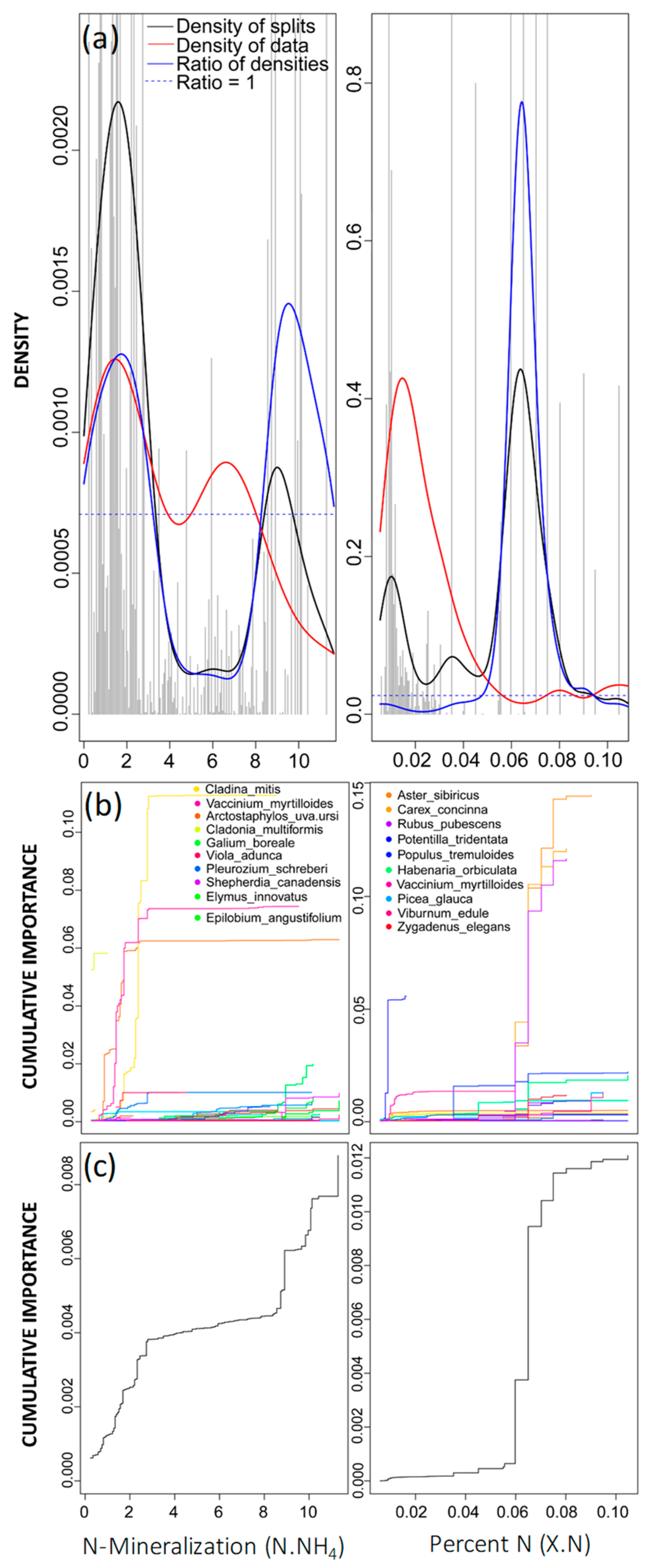
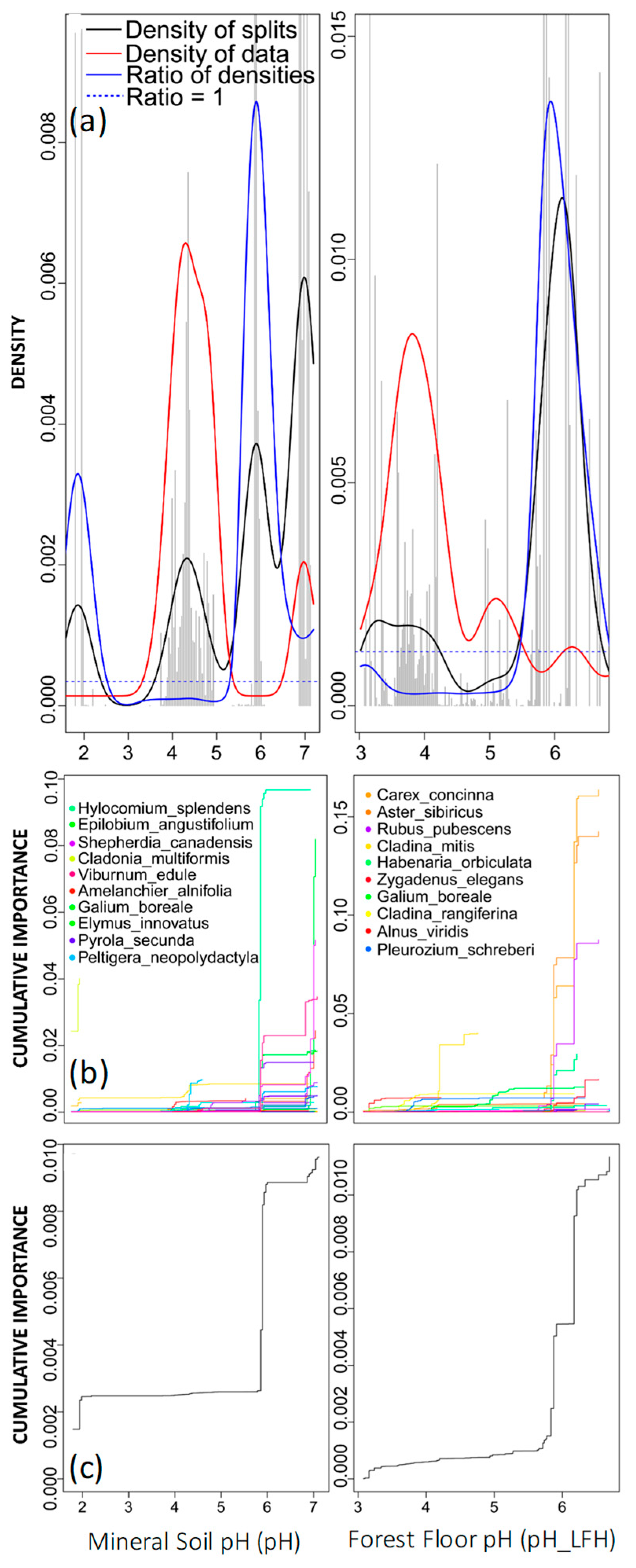
2.6. GradientForest Outputs
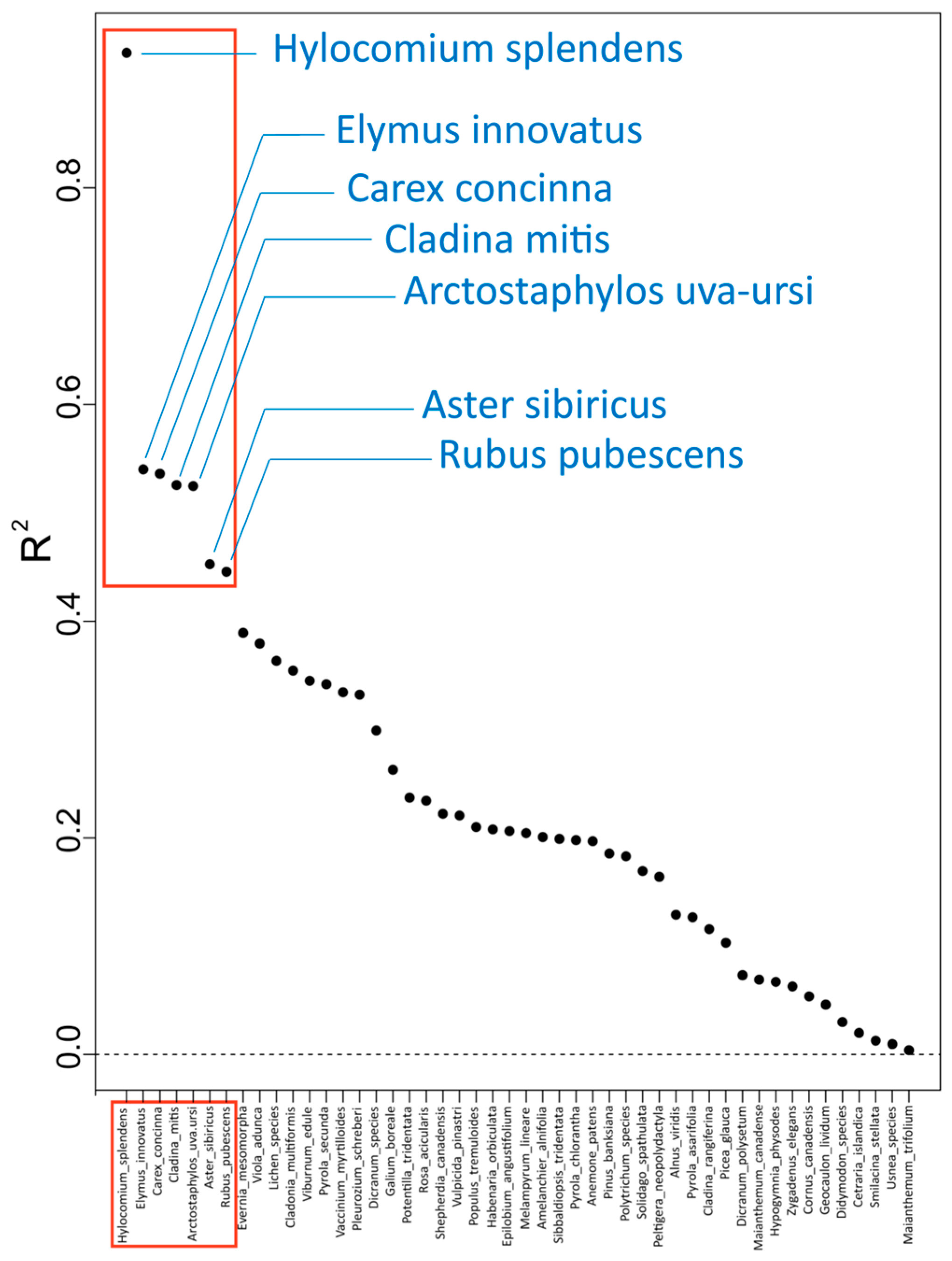
3. Results
4. Discussion
4.1. Variance Partitioning and Joint Effects
4.2. Drivers of Community Thresholds for Jack Pine Forests in the AOSR
4.3. Biodiversity-Based Empirical Critical Loads
4.4. Limitations and Uncertainties
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aber, J.D.; Nadelhoffer, K.J.; Steudler, P.; Melillo, J.M. Nitrogen saturation in Northern forest ecosystems. Bioscience 1989, 39, 378–386. [Google Scholar] [CrossRef]
- Bobbink, R.; Hornung, M.; Roelofs, J.G.M. The effects of air-borne nitrogen pollutants on species diversity in natural and semi-natural European vegetation. J. Ecol. 1998, 86, 717–738. [Google Scholar] [CrossRef]
- Dise, N.B.; Ashmore, M.; Belyazid, S.; Bleeker, A.; Bobbink, R.; de Vries, W.; Erisman, J.W.; Spranger, T.; Stevens, C.J.; van den Berg, L. Nitrogen as a threat to European terrestrial biodiversity. In The European Nitrogen Assessment; Sutton, M.A., Howard, C.M., Erisman, J.W., Billen, G., Bleeker, A., Grennfelt, P., van Grinsven, H., Grizzetti, B., Eds.; Cambridge University Press: Cambridge, UK, 2011; Chapter 20. [Google Scholar]
- Fenn, M.E.; Haeuber, R.; Tonnesen, G.S.; Baron, J.S.; Grossman-Clarke, S.; Hope, D.; Jaffe, D.A.; Copeland, S.; Geiser, L.; Rueth, H.M.; et al. Nitrogen emissions, deposition and monitoring in the Western United States. Bioscience 2003, 53, 391–403. [Google Scholar] [CrossRef]
- Galloway, J.N.; Aber, J.D.; Erisman, J.W.; Seitzinger, S.P.; Howarth, R.W.; Cowling, E.B.; Cosby, B.J. The nitrogen cascade. BioScience 2003, 53, 341–356. [Google Scholar] [CrossRef]
- Bobbink, R.; Loran, C.; Tomassen, H. Review and Revision of Empirical Critical Loads of Nitrogen for Europe; German Environment Agency: Dessau-Roßlau, Germany, 2022; p. 358. [Google Scholar]
- Fenn, M.E.; Baron, J.S.; Allen, E.B.; Rueth, H.M.; Nydick, K.R.; Geiser, L.; Bowman, W.D.; Sickman, J.O.; Meixner, T.; Johnson, D.W.; et al. Ecological effects of nitrogen deposition in the Western United States. BioScience 2004, 53, 404–420. [Google Scholar] [CrossRef]
- Geiser, L.H.; Jovan, S.E.; Glavich, D.A.; Porter, M.K. Lichen-based critical loads for atmospheric nitrogen deposition in Western Oregon and Washington forests, USA. Environ. Pollut. 2010, 158, 2412–2421. [Google Scholar] [CrossRef] [PubMed]
- Maskell, L.C.; Smart, S.M.; Bullock, J.M.; Thompson, K.; Stevens, C.J. Nitrogen deposition causes widespread loss of species richness in British habitats. Glob. Change Biol. 2010, 16, 671–679. [Google Scholar] [CrossRef]
- Nilsson, J.; Grennfelt, P. Critical Loads for Sulphur and Nitrogen; Report from a workshop held at Skokloster, Sweden; UNECE and the Nordic Council of Ministers: Sweden, 1988. [Google Scholar]
- Wilkins, K.; Clark, C.; Aherne, J. Ecological thresholds under atmospheric nitrogen deposition for 1200 herbaceous species and 24 communities across the United States. Glob. Chang. Biol. 2022, 28, 2381–2395. [Google Scholar] [CrossRef]
- Bobbink, R. Plant Species Richness and the Exceedance of Empirical Nitrogen Critical Loads: An Inventory; Report Landscape Ecology; Utrecht University/RIVM: Utrecht, The Netherlands, 2004; p. 19. [Google Scholar]
- Environment and Climate Change Canada. Canadian Environmental Sustainability Indicators: Air Pollutant Emissions. 2019. Available online: www.canada.ca/en/environment-climate-change/services/environmental-indicators/air-pollutant-emissions.html (accessed on 19 June 2019).
- Clair, T.A.; Percy, K.E. Assessing Forest Health in the Athabasca Oil Sands Region; WBEA Technical Report 2015-05-25; Wood Buffalo Environmental Association: Fort McMurray, AB, Canada, 2015; p. 180 + Appendices. [Google Scholar]
- Aherne, J.; Shaw, P. Impacts of sulphur and nitrogen deposition in western Canada. J. Limnol. 2010, 69, 1–3. [Google Scholar] [CrossRef]
- de Vries, W.; Hettelingh, J.-P.; Posch, M. Critical Loads and Dynamic Risk Assessments: Nitrogen, Acidity and Metals in Terrestrial and Aquatic Ecosystems; Environmental Pollution Series; Springer: Dordrecht, The Netherlands, 2015; Volume 25. [Google Scholar]
- Aherne, J.; Posch, M. Impacts of nitrogen and sulphur deposition on forest ecosystem services in Canada. Curr. Opin. Environ. Sustain. 2013, 5, 108–115. [Google Scholar] [CrossRef]
- de Vries, W.; Wamelink, G.W.W.; van Dobben, H.; Kros, J.; Reinds, G.J.; Mol-Dijkstra, J.P.; Smart, S.M.; Evans, C.D.; Rowe, E.C.; Belyazid, S.; et al. Use of dynamic soil-vegetation models to assess impacts of nitrogen deposition on plant species composition: An overview. Ecol. Appl. 2010, 20, 69–79. [Google Scholar] [CrossRef]
- Bobbink, R.; Hettelingh, J. Review and Revision of Empirical Critical Loads and Dose-Response Relationships: Proceedings of An Expert Workshop; Noordwijkerhout, Coordination Centre for Effects, National Institute for Public Health and the Environment (RIVM): The Netherlands, 2011. [Google Scholar]
- Henry, J.; Aherne, J. Nitrogen deposition and exceedance of critical loads for nutrient nitrogen in Irish grasslands. Sci. Total Environ. 2014, 470–471, 216–223. [Google Scholar] [CrossRef] [PubMed]
- Pardo, L.H.; Fenn, M.E.; Goodale, C.L.; Geiser, L.H.; Driscoll, C.T.; Allen, E.B.; Baron, J.S.; Bobbink, R.; Bowman, W.D.; Clark, C.M.; et al. Effects of nitrogen deposition and empirical nitrogen critical loads for ecoregions of the United States. Ecol. Appl. 2011, 21, 3049–3082. [Google Scholar] [CrossRef]
- Blake, L.; Goulding, K.W.T.; Mott, C.J.B.; Johnston, A.E. Changes in soil chemistry accompanying acidification over more than 100 years under woodland and grass at Rothamstead Experimental Station, UK. Eur. J. Soil Sci. 1999, 50, 401–412. [Google Scholar] [CrossRef]
- Duprè, C.; Stevens, C.J.; Ranke, T.; Bleekers, A.; Peppler-Lisbach, C.; Gowing, D.J.G.; Dise, N.B.; Dorland, E.; Bobbink, R.; Diekmann, M. Changes in species richness and composition in European acidic grasslands over the past 70 years: The contribution of cumulative atmospheric nitrogen deposition. Glob. Chang. Biol. 2010, 16, 344–357. [Google Scholar] [CrossRef]
- Clark, C.M.; Cleland, E.E.; Collings, S.L.; Fargione, J.E.; Gough, L.; Gross, K.L.; Pennings, S.C.; Suding, K.N.; Grace, J.B. Environmental and plant community determinants of species loss following nitrogen enrichment. Ecol. Lett. 2007, 10, 596–607. [Google Scholar] [CrossRef] [PubMed]
- Stevens, C.J.; Dise, N.B.; Mountford, J.O.; Gowing, D.J. Impact of nitrogen deposition on the species richness of grasslands. Science 2004, 303, 1876–1879. [Google Scholar] [CrossRef]
- Stevens, C.J.; Duprè, C.; Dorland, E.; Gaudnik, C.; Gowing, D.J.G.; Bleeker, A.; Diekmann, M.; Alard, D.; Bobbink, R.; Fowler, D.; et al. Nitrogen deposition threatens species richness of grasslands across Europe. Environ. Pollut. 2010, 158, 2940–2945. [Google Scholar] [CrossRef] [PubMed]
- Henrys, P.A.; Stevens, C.J.; Smart, S.M.; Maskell, L.C.; Walker, K.J.; Preston, C.D.; Crowe, A.; Rowe, E.C.; Gowing, D.J.; Emmett, B.A. Impacts of nitrogen deposition on vascular plants in Britain: An analysis of two national observation networks. Biogeosciences 2011, 8, 3501–3518. [Google Scholar] [CrossRef]
- Baker, M.E.; King, R.S. A new method for detecting and interpreting biodiversity and ecological community thresholds. Methods Ecol. Evol. 2010, 1, 25–37. [Google Scholar] [CrossRef]
- Payne, R.J.; Dise, N.B.; Stevens, C.J.; Gowin, D.J.; BEGIN Partners. Impact of nitrogen deposition at the species level. Proc. Natl. Acad. Sci. USA. 2013, 110, 984–987. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, K.; Aherne, J.; Bleasdale, A. Vegetation community change points suggest that critical loads of nutrient nitrogen may be too high. Atmos. Environ. 2016, 146, 324–331. [Google Scholar] [CrossRef]
- Ellis, N.; Smith, S.J.; Pitcher, C.R. Gradient forests: Calculating importance gradients on physical predictors. Ecology 2012, 93, 156–168. [Google Scholar] [CrossRef]
- Pitcher, C.R.; Lawton, P.; Ellis, N.; Smith, S.J.; Incze, L.S.; Wei, C.-L.; Greenlaw, M.E.; Wolff, N.H.; Sameoto, J.A.; Snelgrove, P.V.R. Exploring the role of environmental variables in shaping patterns of seabed biodiversity composition in regional-scale ecosystems. J. Appl. Ecol. 2012, 49, 670–679. [Google Scholar] [CrossRef]
- McLaughlan, M.S.; Wright, R.A.; Jiricka, R.D. Field Guide to the Ecosites of Saskatchewan’s Provincial Forests; Saskatchewan Ministry of Environment, Forest Service: Prince Albert, SK, Canada, 2010; p. 343. [Google Scholar]
- Ecodynamics Consulting Group International Inc. 2004 Northwest Territories Forest Ecosystem Classification Pilot Project (Final Report); Government of the Northwest Territories, Department of Resources, Wildlife, and Economic Development: Prince Albert, SK, Canada, 2004. [Google Scholar]
- Foster, K.R.; Baines, D.; Percy, K.; Legge, A.; Maynard, D.; Chisholm, V. WBEA TEEM Forest Health Monitoring Program; Draft Procedures Manual. Version 1; Wood Buffalo Environmental Association: Fort McMurray, AB, Canada, 2015; p. 201. [Google Scholar]
- Makar, P.A.; Akingunola, A.; Aherne, J.; Cole, A.S.; Aklilu, Y.-A.; Zhang, J.; Wong, I.; Hayden, K.; Li, S.-M.; Kirk, J.; et al. Estimates of exceedances of critical loads for acidifying deposition in Alberta and Saskatchewan. Atmos. Chem. Phys. 2018, 18, 9897–9927. [Google Scholar] [CrossRef]
- King, J.R.; Jackson, D.A. Variable selection in large environmental data sets using principal component analysis. Environmetrics 1999, 10, 67–77. [Google Scholar] [CrossRef]
- Graham, M.H. Confronting multicollinearity in ecological multiple regression. Ecology 2003, 84, 2809–2815. [Google Scholar] [CrossRef]
- Ramette, A. Multivariate analyses in microbial ecology. FEMS Microbiol. Ecol. 2007, 62, 142–160. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Community Ecology Package: Package ‘Vegan’. R Vignette. 2016. Available online: https://cran.r-project.org (accessed on 12 April 2023).
- Borcard, D.; Gillet, F.; Legendre, P. Numerical Ecology with R; Springer: New York, NY, USA, 2011. [Google Scholar]
- ter Braak, C.J.F. Canonical Correspondence Analysis: A new eigenvector technique for multivariate direct gradient analysis. Ecol. Soc. Am. 1986, 67, 1167–1179. [Google Scholar] [CrossRef]
- Compton, T.J.; Bowden, D.A.; Pitcher, C.R.; Hewitt, J.E.; Ellis, N. Biophysical patterns in benthic assemblage composition across contrasting continental margins off New Zealand. J. Biogeogr. 2012, 40, 75–89. [Google Scholar] [CrossRef]
- Pitcher, C.R.; Ellis, N.; Smith, S.J. Example Analysis of Biodiversity Survey Data with R Package gradientForest. R Vignette. 2011. Available online: http://gradientforest.r-forge.r-project.org/biodiversity-survey.pdf (accessed on 12 April 2023).
- Roubeix, V.; Danis, P.-A.; Feret, T.; Baudoin, J.-M. Identification of ecological thresholds from variations in phytoplankton communities among lakes: Contribution to the definition of environmental standards. Environ. Monit. Assess. 2016, 188, 246. [Google Scholar] [CrossRef]
- Tang, T.; Tang, T.; Tan, L.; Gu, Y.; Jiang, W.; Cai, Q. Identifying community thresholds for lotic benthic diatoms in response to human disturbance. Sci. Rep. 2017, 7, 4134. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.J.; Ellis, N.; Pitcher, C.R. Conditional Variable Importance in R Package Extended Forest. 2014. Available online: http://gradientforest.r-forge.r-project.org/Conditional-importance.pdf (accessed on 12 April 2023).
- Breiman, L. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Large, S.I.; Fay, G.; Friedland, K.D.; Link, J.S. Quantifying patterns of change in marine ecosystem response to multiple pressures. PLoS ONE 2015, 10, e0119922. [Google Scholar] [CrossRef]
- Vandinther, N.; Aherne, J. Ecological Risks from Atmospheric Deposition of Nitrogen and Sulphur in Jack Pine forests of Northwestern Canada. Nitrogen 2023, 4, 102–124. [Google Scholar] [CrossRef]
- Field, R.; Hawkins, B.A.; Cornell, H.V.; Currie, D.J.; Diniz-Filho, A.F.; Guegan, J.-F.; Kaufman, D.M.; Kerr, J.T.; Mittelbach, G.G.; Oberdorff, T.; et al. Spatial species-richness gradients across scales: A meta-analysis. J. Biogeogr. 2009, 36, 132–147. [Google Scholar] [CrossRef]
- Aber:, J.; McDowell, W.; Nadelhoffer, K.; Magill, A.; Berntson, G.; Kamakea, M.; McNulty, S.; Currie, W.; Rustad, L.; Fernandez, I. Nitrogen saturation in temperate forest ecosystems. BioScience 1998, 48, 921–934. [Google Scholar] [CrossRef]
- Lovett, G.M.; Goodale, C.L. A new conceptual model of nitrogen saturation based on experimental nitrogen addition to an Oak forest. Ecosystems 2011, 14, 615–631. [Google Scholar] [CrossRef]
- van Breeman, N.; Driscoll, C.T.; Mulder, J. Acidic deposition and internal proton sources in acidification of soils and water. Nature 1984, 307, 599–604. [Google Scholar] [CrossRef]
- Stevens, C.J.; Duprè, C.; Gaudnik, C.; Dorland, E.; Dise, N.B.; Gowing, D.; Bleeker, A.; Alard, D.; Bobbink, R.; Fowler, D.; et al. Changes in species composition of European acid grasslands observed along a gradient of nitrogen deposition. J. Veg. Sci. 2011, 22, 207–215. [Google Scholar] [CrossRef]
- van Dobben, H.; de Vries, W. Relationship between forest vegetation, atmospheric deposition and site conditions at regional and European scales. Environ. Pollut. 2010, 158, 921–933. [Google Scholar] [CrossRef] [PubMed]
- Pinho, P.; Branquinho, C.; Cruz, C.; Tang, Y.S.; Dias, T.; Rosa, A.P.; Máguas, C.; Martins-Loução, M.-A.; Sutton, M.A. Assessment of critical levels of atmospheric ammonia for lichen diversity in Cork-Oak Woodland, Portugal. In Results of an Expert Workshop Under the Convention on Long-Range Transboundary Air Pollution; Sutton, M., Reis, S., Baker, S., Eds.; Springer: Berlin/Heidelberg, Germany, 2008; Chapter 10; p. 490. [Google Scholar]
- Pitcairn, C.E.R.; Fowler, D.; Leith, I.D.; Sheppard, L.J.; Sutton, M.A.; Kennedy, V.; Okello, E. Bioindicators of enhanced nitrogen deposition. Environ. Pollut. 2003, 126, 353–361. [Google Scholar] [CrossRef] [PubMed]
- Galloway, J.N.; Townsend, A.R.; Erisman, J.W.; Bekunda, M.; Cai, Z.; Freney, J.R.; Martinelli, L.A.; Seitzinger, S.P.; Sutton, M.A. Transformation of the nitrogen cycle: Recent trends, questions and potential solutions. Science 2008, 320, 889–892. [Google Scholar] [CrossRef] [PubMed]
- Allen, E.; Bayley, S.E. Effects of Nitrogen Deposition on Forests and Peatlands: A Literature Review and Discussion of the Potential Impacts of Nitrogen Deposition in the Alberta Oil Sands Region; Wood Buffalo Environmental Association: Fort McMurray, AB, Canada, 2007. [Google Scholar]
- Sutton, M.; Reis, S.; Baker, S. Atmospheric ammonia: Detecting emission changes and environmental impact. In Results of an Expert Workshop Under the Convention on Long-Range Transboundary Air Pollution; Springer: Berlin/Heidelberg, Germany, 2009; p. 490. [Google Scholar]
- Dominati, E.; Patterson, M.; Mackay, A. A framework for classifying and quantifying the natural capital and ecosystem services of soils. Ecol. Econ. 2010, 69, 1858–1868. [Google Scholar] [CrossRef]
- Bobbink, R.; Hicks, W.K. Factors affecting nitrogen deposition impacts on biodiversity: An Overview. In Nitrogen Deposition, Critical Loads and Biodiversity; Sutton, M.A., Mason, K.E., Shepphard, L.H., Sverdrup, H., Haueber, R., Hicks, W.K., Eds.; Springer: Dordrecht, The Netherlands, 2014; Chapter 14; pp. 127–138. [Google Scholar]
- Stevens, C.J.; Duprè, C.; Dorland, E.; Gaudnik, C.; Gowing, D.J.G.; Bleeker, A.; Diekmann, M.; Alard, D.; Bobbink, R.; Fowler, D.; et al. The impact of nitrogen deposition on acid grasslands in the Atlantic region of Europe. Environ. Pollut. 2011, 159, 2243–2250. [Google Scholar] [CrossRef]
- Brunet, J.; Diekmann, M.; Falkengren-Grerup, U. Effects of nitrogen deposition on field layer vegetation in South Swedish oak forests. Environ. Pollut. 1998, 120S, 35–40. [Google Scholar] [CrossRef]
- Clark, C.M.; Phelan, J.; Doraiswamy, P.; Buckley, J.; Cajka, J.C.; Dennis, R.L.; Lynch, J.; Nolte, C.G.; Spero, T.L. Atmospheric deposition and exceedances of critical loads from 1800–2025 for the conterminous United States. Ecol. Appl. 2018, 28, 978–1002. [Google Scholar] [CrossRef]
- Emmett, B.A. Nitrogen saturation of terrestrial ecosystems: Some recent findings and their implications for our conceptual framework. In Acid Rain—Deposition to Recovery; Brimblecombe, P., Hara, H., Houle, D., Novak, M., Eds.; Springer: Dordrecht, The Netherlands, 2007. [Google Scholar]
- Hutchinson, T.F.; Boerner, R.E.J.; Iverson, L.R.; Sutherland, S.; Sutherland, E.K. Landscape patterns of understory composition and richness across a moisture and nitrogen mineralization gradient in Ohio (U.S.A.) Quercus forests. Plant Ecol. 1999, 144, 177–189. [Google Scholar] [CrossRef]
- Stevens, C.J.; Dise, N.B.; Gowing, D.J. Regional trends in soil acidification and exchangeable metal concentrations in relation to acid deposition rates. Environ. Pollut. 2009, 157, 313–319. [Google Scholar] [CrossRef]
- Fenn, M.E.; Geiser, L.; Bachman, R.; Blubaugh, T.J.; Bytnerowicz, A. Atmospheric deposition inputs and effects on lichen chemistry and indicator species in the Columbia River Gorge, USA. Environ. Pollut. 2007, 146, 77–91. [Google Scholar] [CrossRef]
- Horswill, P.; O’Sullivan, O.; Phoenix, G.K.; Lee, J.A.; Leake, J.R. Base cation depletion, eutrophication and acidification of species-rich grasslands in response to long-term simulated nitrogen deposition. Environ. Pollut. 2008, 155, 336–349. [Google Scholar] [CrossRef] [PubMed]
- De Graaf, M.C.C.; Bobbink, R.; Verbeek, P.J.M.; Roelofs, J.G.M. Aluminium toxicity and tolerance in three health land species. Water Air Soil Pollut. 1996, 98, 229–239. [Google Scholar] [CrossRef]
- Bowman, W.D.; Cleveland, C.C.; Halada, L.; Hreško, J.; Baron, J.S. Negative impact of nitrogen deposition on soil buffering capacity. Nature 2008, 1, 767–770. [Google Scholar] [CrossRef]
- Driscoll, C.T.; Lawrence, G.B.; Bulger, A.J.; Butler, T.J.; Cronan, C.S.; Eagar, C.; Lambert, K.F.; Likens, G.E.; Stoddard, J.L.; Weathers, K.C. Acidic deposition in the northeastern United States: Sources and inputs, ecosystem effects, and management strategies. BioScience 2001, 51, 180–198. [Google Scholar] [CrossRef]
- Falkengren-Grerup, U.; Brunet, J.; Diekmann, M. Nitrogen mineralisation in deciduous forest soils in south Sweden in gradients of soil acidity and deposition. Environ. Pollut. 1998, 102, 415–420. [Google Scholar] [CrossRef]
- van den Berg, L.; Vergeer, P.; Rich, T.; Smart, S.; Guest, D.; Ashmore, M.R. Direct and indirect effects of nitrogen deposition on species composition change in calcareous grasslands. Glob. Change Biol. 2010, 17, 1871–1883. [Google Scholar] [CrossRef]
- Bobbink, R.; Hicks, K.; Galloway, J.; Spranger, T.; Alkemade, R.; Ashmore, M.; Bustamante, M.; Cinderby, S.; Davidson, E.; Dentener, F.; et al. Global assessment of nitrogen deposition effects on terrestrial plant diversity: A synthesis. Ecol. Appl. 2010, 20, 30–59. [Google Scholar] [CrossRef]
- Falkengren-Grerup, U.; Diekmann, M. Use of a gradient of N-deposition to calculate effect-related soil and vegetation measures in deciduous forests. For. Ecol. Manag. 2003, 180, 113–124. [Google Scholar] [CrossRef]
- Mäkipää, R. Sensitivity of forest-floor mosses in boreal forests to nitrogen and sulphur deposition. Water Air Soil Pollut. 1995, 85, 1239–1244. [Google Scholar] [CrossRef]
- Nordin, A.; Strengbom, J.; Witzell, J.; Näsholm, T.; Ericson, L. Nitrogen deposition and the biodiversity of boreal forests: Implications for the nitrogen critical load. Ambio: A J. Hum. Environ. 2005, 34, 20–24. [Google Scholar] [CrossRef]
- Phoenix, G.K.; Emmett, B.A.; Britton, A.J.; Caporn, S.J.M.; Dise, N.B.; Helliwell, R.; Jones, L.; Leake, J.R.; Leith, I.D.; Sheppard, L.J.; et al. Impacts of atmospheric nitrogen deposition: Responses of multiple plant and soil parameters across contrasting ecosystems in long-term field experiments. Glob. Change Biol. 2012, 18, 1197–1215. [Google Scholar] [CrossRef]
- Olsson, B.A.; Kellner, O. Long-term effects of nitrogen fertilization on ground vegetation in coniferous forests. For. Ecol. Manag. 2006, 237, 458–470. [Google Scholar] [CrossRef]
- Mäkipää, R. Sensitivity of understorey vegetation to nitrogen and sulphur deposition in a spruce stand. Ecol. Eng. 1998, 10, 87–95. [Google Scholar] [CrossRef]
- Strengbom, J.; Nordin, A.; Näsholm, T.; Ericson, L. Slow recovery of boreal forest ecosystem following decreased nitrogen input. Funct. Ecol. 2001, 15, 451–457. [Google Scholar] [CrossRef]
- Bobbink, R.; Ashmore, M.; Braun, S.; Flückiger, W.; Van den Wyngaert, I.J.J. Empirical nitrogen loads for natural and semi-natural ecosystems: 2002 update. In Empirical Critical Loads for Nitrogen, Environmental Documentation No. 164 Air; Achermann, B., Bobbink, R., Eds.; Swiss Agency for Environment, Forest and Landscape SAEFL: Berne, Switzerland, 2003. [Google Scholar]
- Aerts, R.; de Caluwe, H. Effects of nitrogen supply on canopy structure and leaf nitrogen distribution in Carex species. Ecology 1994, 75, 1482–1490. [Google Scholar] [CrossRef]
- Cunha, A.; Power, S.A.; Ashmore, M.R.; Green, P.R.S.; Haworth, B.J.; Bobbink, R. Whole Ecosystem Nitrogen Manipulation: Updated Review; JNCC Report 331; JNCC: Peterborough, UK, 2002. [Google Scholar]
- Pitcairn, C.E.R.; Leith, I.D.; Sheppard, L.J.; Sutton, M.A.; Fowler, D.; Munro, R.C.; Tang, S.; Wilson, D. The relationship between nitrogen deposition, species composition and foliar nitrogen concentrations in woodland flora in the vicinity of livestock farms. Environ. Pollut. 1998, 102, 41–48. [Google Scholar] [CrossRef]
- Roubeix, V.; Daufresne, M.; Argillier, C.; Dublon, J.; Maire, A.; Nicolas, D.; Raymond, J.-C. and danis, P.-A. Physico-chemical thresholds in the distribution of fish species among French lakes. Knowl. Manag. Aquat. Syst. 2017, 418, 41. [Google Scholar] [CrossRef]
- Turkington, R.; John, E.; Watson, S.; Seccombe-Hett, P. The effects of fertilization and herbivory on the herbaceous vegetation of the boreal forest in Northwestern Canada: A 10-year study. J. Ecol. 2002, 90, 325–337. [Google Scholar] [CrossRef]
- Boonstra, R.; Krebs, C.J.; Cowcill, K. Responses of key understory plants in the boreal forests of western North America to natural versus anthropogenic nitrogen levels. For. Ecol. Manag. 2017, 401, 45–54. [Google Scholar] [CrossRef]
- Gillam, F.S. Response of the herbaceous layer of forest ecosystems to excess nitrogen deposition. J. Ecol. 2006, 94, 1176–1191. [Google Scholar] [CrossRef]
- Stevens, C.J.; Dise, N.B.; Gowing, D.J.G.; Mountford, J.O. Loss of forb diversity in relation to nitrogen deposition in the UK: Regional trends and potential controls. Glob. Change Biol. 2006, 12, 1823–1833. [Google Scholar] [CrossRef]
- Southon, G.E.; Field, C.; Caporn, S.J.M.; Britton, A.J.; Power, S.A. Nitrogen deposition reduces plant diversity and alters ecosystem functioning: Field-scale evidence from a nationwide survey of UK heathlands. PLoS ONE 2013, 8, e59031. [Google Scholar] [CrossRef]
- Geiser, L.H.; Nelson, P.R.; Jovan, P.R.; Root, H.T.; Clark, C.M. Assessing ecological risks from atmospheric deposition of nitrogen and sulfur to US forests using epiphytic macrolichens. Diversity 2019, 11, 87. [Google Scholar] [CrossRef] [PubMed]
- WallisDeVries, M.; Bobbink, R. Nitrogen deposition impacts on biodiversity in terrestrial ecosystems: Mechanisms and perspectives for restoration. Biol. Conserv. 2017, 212, 387–389. [Google Scholar] [CrossRef]
- Jones, F.C.; Plewes, R.; Murison, L.; MacDougall, M.J.; Sinclair, S.; Davies, C.; Bailey, J.L.; Richardson, M.; Gunn, J. Random forests as a cumulative effects models: A case study of lakes and rivers in Muskoka, Canada. J. Environ. Manag. 2017, 201, 407–424. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Environmental Variable | Range | Median | Mean | SD |
|---|---|---|---|---|---|
| TDN | Total Deposited Nitrogen (eq ha−1 yr−1) | 37.21–597.01 | 136.70 | 176.45 | 131.60 |
| WNO3 | Wet Nitrate (eq ha−1 yr−1) | 3.34–28.77 | 18.36 | 17.36 | 6.51 |
| TDS | Total Deposited Sulphur (eq ha−1 yr−1) | 23.21–1182.20 | 99.83 | 179.41 | 212.08 |
| WSO4 | Wet Sulphate (eq ha−1 yr−1) | 11.74–57.67 | 27.56 | 29.23 | 9.82 |
| LATITUDE | Latitude (°) | 56.25–60.88 | 57.08 | 57.54 | 1.31 |
| LONGITUDE | Longitude (°) | −113.74–108.89 | −110.45 | −110.68 | 1.20 |
| ELEVATION | Elevation (m) | 231–622.52 | 460.34 | 425.50 | 108.37 |
| bio_12 | Annual Precipitation (mm) | 359–483 | 451.50 | 441.87 | 31.95 |
| sg_01 | Julian day number at start of growing season | 115–136 | 123.5 | 124.33 | 6.20 |
| sg_04 | Total Precipitation for period 1 (mm) | 50–70.7 | 63.60 | 63.07 | 5.46 |
| sg_06 | Total precipitation for period 3 (mm) | 209.40–324.1 | 304 | 293.27 | 31.26 |
| sg_09 | Gdd above base temperature for Period 2 | 194–245 | 219 | 218.06 | 14.17 |
| sg_12 | Annual Mean Temperature (°C) | −2.8–1.64 | 0.44 | 0.026 | 1.28 |
| pH | pH—Soil | 3.59–7.19 | 4.46 | 4.75 | 0.96 |
| LOI | Loss on Ignition (%) − Soil | 0.40–41.72 | 1.15 | 2.78 | 6.19 |
| X._Moisture | % Moisture − Soil | 0.096–8.25 | 0.38 | 0.69 | 1.28 |
| Bulk_Density | Bulk Density (g cm−3) − Soil | 0–1.64 | 0 | 0.58 | 0.65 |
| N_NH4 | NH4 N-Mineralization (μg L−1) − Soil | 0–11.65 | 2.86 | 4.09 | 3.20 |
| PO4_H2O | PO4 Water Extractions (mg L−1) − Soil | 0–0.46 | 0 | 3.81 × 10−2 | 8.64 × 10−2 |
| Al_EC | Al Exchangeable Cations (mg L−1) − Soil | 0.03–2.43 | 0.23 | 0.44 | 0.48 |
| Fe_EC | Fe Exchangeable Cations (mg L−1) − Soil | −8.80 × 10−2–0.39 | 4.12 × 10−2 | 8.23 × 10−2 | 0.12 |
| Mn_EC | Mn Exchangeable Cations (mg L−1) − Soil | 1.90 × 10−2–0.54 | 0.15 | 0.17 | 0.12 |
| Na_EC | Na Exchangeable Cations (mg L−1) − Soil | 3.00 × 10−2–0.70 | 0.10 | 0.14 | 0.12 |
| X.N | CNS − % Nitrogen − Soil | 5.33 × 10−3–0.10 | 1.77 × 10−2 | 2.49 × 10−2 | 2.19 × 10−2 |
| X.S | CNS − % Sulphur − Soil | 0–2.0 × 10−2 | 2.81 × 10−3 | 3.84 × 10−3 | 4.28 × 10−3 |
| C/N_Ratio | Carbon to Nitrogen Ratio − Soil | 16.9–43.40 | 26.32 | 27.32 | 5.82 |
| pH_LFH | pH–LFH | 3.02–7.01 | 3.96 | 4.23 | 0.90 |
| Fl_LFH_H2O | Fl Water Extractions (mg L−1) − LFH | 0–0.98 | 0.14 | 0.20 | 0.21 |
| Cl_LFH_H2O | Cl Water Extractions (mg L−1) − LFH | 0–10.32 | 0.79 | 1.34 | 1.80 |
| NO2_LFH_H2O | NO2 Water Extractions (mg L−1) − LFH | 0–3.86 × 10−2 | 1.05 × 10−2 | 1.27 × 10−2 | 1.04 × 10−2 |
| NO3_LFH_H2O | NO3 Water Extractions (mg L−1) − LFH | 0–0.30 | 7.11 × 10−2 | 7.67 × 10−2 | 6.71 × 10−2 |
| PO4_LFH_H2O | PO4 Water Extractions (mg L−1) − LFH | 0–11.52 | 4.36 | 4.22 | 3.84 |
| X.N_LFH | CNS − % Nitrogen − LFH | 0–1.48 | 0.99 | 0.95 | 0.28 |
| X.S_LFH | CNS − % Sulphur − LFH | 0–0.22 | 6.37 × 10−2 | 7.76 × 10−2 | 4.25 × 10−2 |
| C/N_Ratio_LFH | Carbon to Nitrogen Ratio − LFH | 0–59.91 | 38.17 | 37.21 | 10.31 |
| Environmental Variable | R2 Weighted Importance | Environmental Variable | R2 Weighted Importance |
|---|---|---|---|
| Longitude | 0.0147 | Al_EC | 0.0091 |
| Latitude | 0.0146 | N.NH4 | 0.0087 |
| sg_09 | 0.0138 | TDN | 0.0084 |
| sg_12 | 0.0135 | WSO4 | 0.0079 |
| sg_01 | 0.0128 | Mn_EC | 0.0075 |
| X.S_LFH 1 | 0.0125 | LOI | 0.0066 |
| X.N 1 | 0.0121 | WNO3 | 0.0061 |
| pH_LFH | 0.0113 | sg_06 | 0.0059 |
| TDS | 0.0111 | bio_12 | 0.0053 |
| pH | 0.0096 | X._Moisture | 0.0044 |
| Mean R2 Weighted Importance | 0.00685 | ||
| Species Information | R2 Overall Importance | Specific R2S | Environmental Variable |
|---|---|---|---|
| Hylocomium splendens, bryophyte | 0.925 | 0.1100 | sg_01 |
| 0.0967 | pH | ||
| 0.0908 | Latitude | ||
| 0.0851 | TDN | ||
| 0.0820 | Longitude | ||
| Elymus innovatus, vascular | 0.540 | 0.1570 | TDS |
| 0.0960 | WSO4 | ||
| 0.0607 | TDN | ||
| 0.0270 | sg_01 | ||
| 0.0256 | Longitude | ||
| Carex concinna, vascular | 0.536 | 0.1640 | pH_LFH |
| 0.1210 | X.N | ||
| 0.0495 | X._Moisture | ||
| 0.0444 | sg_12 | ||
| 0.0376 | Latitude | ||
| Cladina mitis, lichen | 0.526 | 0.1120 | N.NH4 |
| 0.1040 | sg_09 | ||
| 0.0459 | sg_06 | ||
| 0.0400 | pH_LFH | ||
| 0.0340 | PO4_LFH_H2O | ||
| Arctostaphylos-uva-ursi, vascular | 0.525 | 0.1210 | sg_01 |
| 0.1120 | X.S_LFH | ||
| 0.0759 | sg_12 | ||
| 0.0626 | N.NH4 | ||
| 0.0538 | LOI | ||
| Aster sibiricus, vascular | 0.453 | 0.1440 | X.N |
| 0.1420 | pH_LFH | ||
| 0.0291 | sg_01 | ||
| 0.0263 | X._Moisture | ||
| 0.0196 | Latitude | ||
| Rubus pubescens, vascular | 0.446 | 0.1160 | X.N |
| 0.0949 | sg_12 | ||
| 0.0873 | pH_LFH | ||
| 0.0303 | X._Moisture | ||
| 0.0205 | sg_01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vandinther, N.; Aherne, J. Biodiversity-Based Empirical Critical Loads of Nitrogen Deposition in the Athabasca Oil Sands Region. Nitrogen 2023, 4, 169-193. https://doi.org/10.3390/nitrogen4020012
Vandinther N, Aherne J. Biodiversity-Based Empirical Critical Loads of Nitrogen Deposition in the Athabasca Oil Sands Region. Nitrogen. 2023; 4(2):169-193. https://doi.org/10.3390/nitrogen4020012
Chicago/Turabian StyleVandinther, Nicole, and Julian Aherne. 2023. "Biodiversity-Based Empirical Critical Loads of Nitrogen Deposition in the Athabasca Oil Sands Region" Nitrogen 4, no. 2: 169-193. https://doi.org/10.3390/nitrogen4020012
APA StyleVandinther, N., & Aherne, J. (2023). Biodiversity-Based Empirical Critical Loads of Nitrogen Deposition in the Athabasca Oil Sands Region. Nitrogen, 4(2), 169-193. https://doi.org/10.3390/nitrogen4020012