Abstract
This study investigated structural changes in microbial community of biological nutrient removal (BNR) in response to changes in substrate composition (ammonium and phosphate), redox condition, and morphological characteristics (flocs to granules), with a focus on nitrification and phosphate removal. Analyzing treatment performance and 16S rRNA phylogenetic gene sequencing data suggested that heterotrophic nitrification (HN) and autotrophic nitrification (AN) potentially happened in aerobic organic-rich (HN_AS) and aerobic organic-deficient (AN_AS) activated sludge batch reactors, respectively. However, phosphate release and uptake were not observed under alternating anaerobic/aerobic regime. Phosphate release could not be induced even when anaerobic phase was extended, although Accumulibacter existed in the inoculum (5.1% of total bacteria). Some potential HN (e.g., Thauera, Acinetobacter, Flavobacterium), AN (e.g., Nitrosomonas (3.2%) and Nitrospira), and unconventional phosphate-accumulating organisms (PAOs) were identified. Putative HN bacteria (i.e., Thauera (29–36%) and Flavobacterium (18–25%)) were enriched in aerobic granular sludge (AGS) regardless of the granular reactor operation mode. Enrichment of HN organisms in the AGS was suspected to be mainly due to granulation, possibly due to the floc-forming ability of HN species. Thus, HN is likely to play a role in nitrogen removal in AGS reactors. This study is supposed to serve as a starting point for the investigation of the microbial communities of AS- and AGS-based BNR processes. It is recommended that the identified roles for the isolated bacteria are further investigated in future works.
1. Introduction
Molecular techniques have identified numerous bacteria from diverse genera and phyla, and sometimes organisms from distinct kingdoms (i.e., Archaea and Eukarya) in wastewater treatment processes [1,2]. Biological functions of most wastewater organisms have remained unexplored, with the attention focused on certain organisms that are believed to play fundamental functional roles in selected series of transformations such as autotrophic nitrification (AN), anoxic denitrification (AXD), and classical anaerobic phosphorus (P) release/aerobic P uptake.
Recently, heterotrophic nitrification (HN) has been attributed to diverse bacteria distributing across ten genera, including Arthrobacter [3], Acinetobacter [4,5], Pseudomonas [6,7], Alcaligenes [8], Bacillus [9,10], Thiosphaera [11], Comamonas [12], and Thauera [1]. In most of these studies, it is reported that the identified HN species can simultaneously perform HN and aerobic denitrification (AD). Integrating HN–AD process by a single species is of practical interest, because it can result in convenient simultaneous nitrification–denitrification (SND) in a single biological wastewater treatment reactor. The SND can be accomplished in a single bioreactor using emerging technologies such as anaerobic ammonium oxidation (anammox) and aerobic granular sludge (AGS), albeit with difficulty, because certain operating conditions should be maintained to allow culturing organisms with incompatible metabolic requirements, i.e., anammox, AN, and AXD bacteria, and polyphosphate-accumulating organisms (PAOs) [13,14].
The HN capacity and optimal growth conditions of HN bacteria have been mainly investigated in pure cultures so far [3,10,15]. Due to pure-culture biases, derived conclusions from these studies cannot be extrapolated to real-time conditions in wastewater treatment processes where mixed microbial consortia are present. This necessitates further research to shed light on the contribution of HN bacteria to wastewater treatment in biological nutrient removal (BNR) processes.
In addition, there are uncertainties about efficiency and microbiology of enhanced biological phosphate removal (EBPR), a BNR process developed based on classical anaerobic release–aerobic uptake of orthophosphate (ortho-P) [16]. Nonetheless, it has been suggested that the PAOs can maintain their metabolism under continuously aerobic condition as well [17,18]. Ortho-P removal mechanism under such circumstance is not well understood.
Through lab- and full-scale studies, Rhodocyclus-related bacteria, namely “Candidatus Accumulibacter phosphatis” (in short, Accumulibater), is recognized as major PAO species [19]. Yet, Acinetobacterial PAOs are found more abundantly in full-scale EBPR processes [20,21,22], implying the potential role of these organisms in EBPR. Moreover, the possible role of some other species in phosphate accumulation has recently been pointed out, such as Dechloromonas belonging to Betaproteobacteria, Acinetobacterial filament “Candidatus Microthrix parvicella” (hereinafter, Microthrix), Corynebacterium, Pseudomonas, and Acinetobacteria [20,21,23]. Accumulation of poly-P granules has been observed in the cellular tissues of some of these species [21,24]. These uncertainties and ambiguities warrant further research on EBPR microbiology coupled with its performance.
Therefore, this study attempted to investigate structural changes in functional populations of BNR microbial community in response to different selection pressures (i.e., substrate composition, redox condition, and physical transformation). Studying the effect of these changes on BNR microbial community is important to enable the operators and researchers to investigate and understand the link between process performance and operational changes. Raw wastewater shows fluctuations in substrate composition and loadings on daily and seasonal bases. For example, a Canadian domestic wastewater sample has shown large variations in biochemical oxygen demand (BOD), chemical oxygen demand (COD), and total suspended solids (TSS) throughout a year, in ranges of 80–1000 mg/L, 100–1650 mg/L, and 120–1330 mg/L TSS, respectively (author’s personal experience). Variations in redox conditions are the essence of the design of AS-based BNR processes. Different redox conditions can happen in a single AGS granule. Physical transformation from suspended microbial cells to AS flocs has been constantly observed in AS basins, and it is the foundation of AGS processes. It has been hypothesized that BNR microbial community responds to these changes, which can therefore reflect in BNR process performance.
As such, the objectives of this study were to (i) determine evolution in activated sludge (AS) microbial structure under given incubation conditions deemed suitable for HN, in order to identify putative HN species; (ii) demonstrate whether dissolved oxygen (DO) enrichment can sustain PAO population and EBPR performance of AS under DO-rich condition; and (iii) study EBPR microbiology and performance under anaerobic/aerobic operating regime. The scope of the study was defined to address a few gaps in knowledge identified in the literature as outlined above, including understanding the contribution of HN bacteria to total nitrogen (TN) removal in BNR processes, and microbiology and performance of biological P removal under different redox conditions (i.e., aerobic heterotrophic, aerobic autotrophic, heterotrophic anaerobic/aerobic, and heterotrophic extended anaerobic media). Four AS batch studies were performed with these media, and the information collected from these batch studies were extrapolated to AGS bioreactors, to identify functional species in AGS.
It is hoped that this study can start a conversation about microbial communities of AS- and AGS-based BNR processes under different circumstances and can, hence, help further practical knowledge pertaining to BNR processes and their optimization.
2. Materials and Methods
2.1. Experimental Setups and Operational Conditions
To perform AS experiments, geometrically-identical cylindrical bioreactors ABDOSTM P50607 Large Capacity PP Beakers, 5000 mL (Cole-Parmer, Montreal, QC, Canada) were used, with the specifications described in Pishgar et al. [25,26]. Briefly, the dimensions of these bioreactors included internal diameter (ID) of 18.7 cm, height (H) of 20 cm, and working volume (WV) of 5 L.
Synthetic wastewater with given substrate compositions, which would stimulate the growth of microorganisms of interest, was prepared and added to the relevant bioreactor one time at the beginning of the operation (Table 1). The experiments were continued until observing a decay phase in the microbial growth pattern, to allow the completion of substrate conversions and the associated bioreactions. This would ensure that the expected changes in the microbial communities had occurred. In aerobic experiments, continuous aeration was provided using fine pore diffusers (Pentair AS40, Minneapolis, MN, USA), alongside mixing for homogenization. In anaerobic experiments, mechanical mixers (laboratory mixer Stir-Pak Model 04555-00 (Cole-Parmer, Montreal, QC, Canada), equipped with three-bladed propeller were used. DO concentration was monitored in situ intermittently (i.e., at the sampling times), to ensure achieving the target environmental conditions as delineated in Table 1. The experiments were conducted in triplicate.

Table 1.
Media composition.
To perform AGS experiments, the granules were cultivated and the treatment performance was investigated in two geometrically identical, bubble-column, granular sequencing batch reactors (GSBRs), with the specifications described in Pishgar et al. [27,28,29] and Pishgar [30]. Briefly, the AGS reactors’ dimensions included ID of 13.7 cm, H of 150 cm, and WV of 18 L. The GSBRs were operated under two different modes: (i) alternating anaerobic/aerobic/anoxic/aerobic mode (AN/O/AX/O_SBR bioreactor): 8 min fill, 55 min anaerobic (effluent recirculation for mixing), 104 min aeration, 1 min fill, 30 min anoxic (effluent recirculation for mixing), 30 min aeration, 10 min settle, and 2 min decant; (ii) purely aerobic mode (O_SBR bioreactor): 8 min fill, 220 min aeration, 10 min settle, 2 min decant. Cycle duration, volumetric exchange ratio, hydraulic retention time (HRT), and aeration intensities in the GSBRs were identical (4 h, 60%, and 6.7 h, and 2.7 cm/s, respectively).
2.2. Inoculum and Media
The BNR return activated sludge (RAS) was collected from Pine Creek BNR WWTP, Calgary, AB, Canada, and used as seed for both AS and AGS experiments. The inoculum’s mixed liquor suspended solids (MLSS) concentration was 7746 ± 498 mg/L and had a volatile content of 81 ± 1.2%. Median floc size of the inoculum was 126 ± 7 µm. The volumes of 10 and 30% (v/v) were inoculated into AS bioreactors and GSBRs, resulting in initial biomass concentrations (MLSSin) of 700 and 2800 mg MLSS/L, respectively. Due to limited mixing capacity of AS bioreactors, relatively diluted initial mixed liquor concentration was chosen for AS experiments to avoid biomass settling at the farthest zones (from the mixer and/or diffuser) of the bioreactors. It would otherwise cause having less active or completely inactive biomass in local zones of the bioreactors, which could distort the results of microbial community analysis.
The compositions of the used media are summarized in Table 1. Experimental design and operating conditions and objective of each batch study are defined in Table 2. In AS experiments, medium compositions and metabolic conditions were adjusted to encourage proliferation of certain group of bacterial species in each bioreactor.
Table 2.
Experiment design and operating conditions.
In heterotrophic nitrification AS experiment (HN_AS bioreactor), the biomass was exposed to high concentrations of organic matter (5500 mg chemical oxygen demand (COD)/L) and organic nitrogen (100 mg total nitrogen (TN)/L, solely supplemented by peptone) to satisfy the growth requirement of heterotrophic nitrifiers. Carbon to nitrogen (C/N) ratio is reported as a critical factor for HN. Optimum range of C/N ratio (5–12) could promote growth and enrichment of HN bacteria [5,7,9,10,31,32,33]. As such, in HN_AS medium, initial concentrations of total organic carbon (TOC) and TN were adjusted to achieve C/N ratio of 10 which, as suggested by van Niel et al. [34], could cause HN species to outcompete autotrophic ammonium oxidizers (e.g., Nitrosomonas europaea). Different sources of organic nitrogen can serve HN as sole nitrogen source. Examples of organic nitrogenous substrates for HN are oximes (e.g., pyruvic oxime and acetaldoxime), hydroxamic acids, amino acids (e.g., glycine, and L-glutamic acid), urea, maize straw, yeast extract, peptone, and casein [35,36,37,38,39]. In this study, peptone was selected.
In the autotrophic nitrification AS experiment (AN_AS bioreactor), ammonium was the sole nitrogen source and bicarbonate was added as inorganic carbon source required for the growth of autotrophic nitrifiers. The AH_AS experiment was supposed to serve as a negative control to justify the ecophysiological rationale used to identify potential heterotrophic nitrifiers.
To investigate P removal, the AS experiment was performed under alternating anaerobic/aerobic regime (AN/O_AS bioreactor). A purely anaerobic AS experiment for achieving an extended anaerobic period (ExAN_AS bioreactor) was conducted to ensure that anaerobic P release would have enough time to emerge. Biological P removal could also be monitored in organic-rich HN_AS and organic-deficient AN_AS media, because phosphate was added to HN_AS and AN_AS bioreactor as supplementary nutrient (see Table 1).
The AGS experiments were designed to investigate the microbial diversity of AGS developed under distinct incubation conditions. High COD concentration was adopted in these experiments (Table 1) to rapidly grow aerobic granules under high organic loading condition (i.e., 7 kg COD/m3/d). The N and P were supplemented to support aerobic growth requirement of bacteria (i.e., COD:N:P = 100:5:1). Wastewater composition and substrate loading were fixed to impose certain selection pressure by nourishment condition on the AGS in the GSBRs. To read more about AGS development, maturation, and characteristics, as well as AGS treatment performance, refer to Pishgar et al. [27,28,29] and Pishgar [30].
2.3. Water Quality and Microbial Community Analyses
Water quality analyses included pH and temperature (YSI MultiLab IDS 4010-3). In situ DO concentration was carried out using portable probe (YSI ProComm II, Yellow Springs, OH, USA). MLSS concentration was determined based on the mean of four replicates, following standard methods [40]. MLSS measurement of AS reactors was performed using a calibration curve and optical density measurements at 620 nm (OD620), to avoid significant biomass loss and disruption in the process. Samples were centrifuged (6000 rpm for 10 min in Hermle Z 206 A, Wehingen, Germany) and filtered (0.45 µm Hydrophobic Teflon®, SCP Science, Mississauga, ON, Canada) prior to the subsequent analyses. COD and NH4–N concentrations were measured in accordance with standard methods [40], using Hach® kits (Mississauga, ON, Canada). Anions (NO3−, NO2−, PO43−) were analyzed using ion chromatography (Metrohm Enviro Pro IC, the column Metrosep A Supp 5-100/4.0, Mississauga, ON, Canada). A TOC/TN analyzer (Shimadzu TOC-LCPH and TNM-L with ASI-L autosampler, 720 °C catalytic thermal decomposition/chemiluminescence method, West Chicago, IL, USA) was used for combined TOC and TN measurements.
In AS experiments, the data points were obtained using the mean of a total of six readings (two measurements from each of the three replicates). The AGS data were taken from a typical SBR cycle during maturation phases (day 59 in AN/O/AX/O_SBR and day 53 in O_SBR). Samples were withdrawn at proper time intervals to reflect the dynamics of substrate conversions.
Microbial community analysis was performed using 16S rRNA phylogenetic gene sequencing and constructing the corresponding clone library. This analysis is described in Pishgar et al. [26,27,28,29] and Pishgar [30] in detail. Briefly, genomic DNA of samples was extracted using DNeasy PowerSoil Kit from QIAGEN, Inc. (Calgary, AB, Canada). Paired-end sequencing was conducted (Illumina MiSeq platform, San Diego, CA, USA) using two primers (357wF and 785R) to identify 16S rRNA phylogenetic genes. The microbial analyses were performed at the Centre for Health Genomics and Informatics in the Cumming School of Medicine, University of Calgary, which deposits the data generated from MiSeq and runs on its own server and on Illumina’s BaseSpace Sequence Hub. Sampling for the microbial analysis is described in Pishgar et al. [26] and Pishgar [30].
3. Results and Discussion
3.1. Treatment Performance
3.1.1. Heterotrophic Nitrification
HN_AS biomass showed different nitrogen removal abilities (Figure 1a). Initial TN concentration reduced to 12.8 mg/L within 43, which corresponded to 87% removal efficiency. Accumulation of small concentrations of ammonium and nitrite were observed in 42–67 h which, respectively, declined to trace concentrations of 0.89 and 0.23 mg N/L in 116 h. Nitrate accumulation started at hour 24 and continued with a constant rate of 0.35 mg NO3–N/L/h.
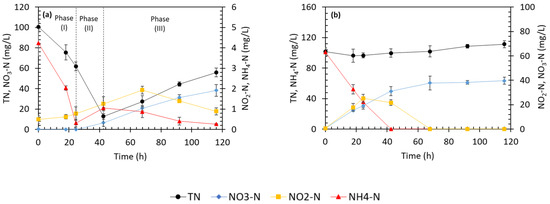
Figure 1.
Treatment performance of (a) heterotrophic nitrification (HN_AS) and (b) autotrophic nitrification activated sludge (AN_AS) bioreactors.
Accumulations of NH4+, NO2−, and, especially, NO3− suggested that HN might happen in this bioreactor. In organic HN pathway (also known as ammonification), organic nitrogen (general formula of R-NH2) is hydrolyzed to NH4+. In the consecutive inorganic HN steps, NH4+ is converted to NH2OH, NO2−, and then NO3− [4]. In direct oxidative pathway (another type of organic nitrification), HN bacteria directly oxidize organic nitrogen to the end products (i.e., NH2OH or NO3−), with minor or no intermediate metabolite production. Examples include Alcaligenes faecalis [8], Pseudomonas stutzeri [41], and Acinetobacter sp. [4]. Accumulation of trace concentrations of intermediate products, i.e., NH4+ and NO2−, could propose greater share of direct oxidative pathway to overall N removal.
Biological assimilation and HN could simultaneously contribute to TN removal during the first 24 h (phase I, Figure 1a). Organic HN could convert organic nitrogen to ammonium which could be taken up rapidly for growth. Conversion of any type of nitrogen to glutamate before incorporation into cell mass is a must. Ammonium is the precursor of glutamate and thus is the preferred source of nitrogen for assimilation. If available, it is primarily used for cell synthesis [31,42]. The exponential growth of biomass (Figure S1a) suggested that within the first 43 h when TN removal was under progress, biomass experienced a drastic growth and MLSS reached a maximal value of 1260 mg/L at 43 h. During this time, TN treatment was accompanied by mineralization of organic matter (compare Figure 1a and Figure S1b); COD concentration of 5500 mg/L was degraded by 99% within the first 43 h of the operation, with an estimated degradation rate of 128 mg COD/L/h. Rapid biodegradation of high COD concentration caused rapid depletion of available oxygen, and thus, limited DO was observed during the first 24 h (Figure S1c), although air was constantly diffused into the bioreactor. When organic matter became scarce at 24 h, biomass autolysis and decay commenced (Figure S1a), nitrate concentration increased, and TN profile showed an increasing trend instead of being further reduced (Figure 1a).
Co-occurrence of nitrogen assimilation and HN seems to be a common phenomenon. For instance, Alcaligenes faecalis could remove 38–50% of 500–1000 mg NH4–N/L, which was attributed to incorporation into cellular tissues [43]. According to Sun et al. [42], ammonium assimilation is a crucial accessory for AD which typically follows HN. Assimilation of ammonium affects HN positively through increasing the count of HN bacteria [42]. However, it seems that contribution of assimilation to overall nitrogen removal would increase if high C/N ratio (>10) is maintained. Thus, optimal C/N ratio of 5–7 has been anticipated for HN [33,42].
In Phase II, HN possibly became the principal nitrogen removal pathway when COD concentration became limited at 24 h and C/N ratio reduced to 7 (Figure 1a). In this phase, the process was not accompanied by exponential biomass growth anymore. A stationary phase occurred from 24 h onward (Figure S1a), at which TN removal rate of 2.7 mg TN/L/h was estimated.
Accumulation of NO3− in phase III (Figure 1a) could suggest that the activity of HN bacteria could continue during death phase (Figure S1a). Cell decay and decomposition could discharge intracellular nitrogen into the surrounding environment in the form of ammonium, which could be immediately oxidized to nitrate via HN.
3.1.2. Autotrophic Nitrification
As mentioned in the Section 2, AN_AS bioreactor could serve as a negative control to justify the rationale used to identify putative HN bacteria (see previous section). Ammonium concentration of 100 mg NH4–N/L was completely removed in 43 h (Figure 1b), with a removal rate of 2.4 mg NH4–N/L/h. Nitrite and nitrate accumulated concurrently. Nitrite reached maximum concentration of 25 mg N/L within 24 h and subsequently reduced during the following 36 h. Nitrite was not detected after 68 h. Nitrate gradually accumulated from the start of the operation. Its production rate sustained a fast pace (1.14 mg NO3–N/L/h) up to 43 h, whereas it significantly slowed down during the rest of the operation, leading to a plateau from 68 h onward.
TN concentration gradually increased slightly toward the end of the operation, which could be due to the combined effect of ammonium transformation to other nitrogen forms and cell decay. Under aerobic condition, initial ammonium concentration (i.e., 100 mg NH4–N/L) was converted to nitrite, and then nitrate, as described above. In addition, according to cell decay stoichiometry shown in Equation (1), decrease in biomass concentration (Figure S1d), interpreted as biomass decay, could contribute to increase in ammonia concentration:
Anoxic denitrification could not proceed due to aerobic redox condition, hence the constant nitrate nitrogen concentration in the medium toward the end of operation. Consecutive AN steps were identical to those of inorganic HN pathways. It is because inorganic nitrogen sources (i.e., NH4+, NO2−, and NO3−) serve as substrates for both AN and HN [4,41]. Thus, HN was likely to mutually contribute to ammonium removal in AN_AS medium. However, the trend of biomass concentration suggested that the role of HN, if any, was insignificant. In the AN_AS bioreactor, MLSS continuously reduced from the beginning of operation, possibly due to lack of readily biodegradable organic matter for growth (Figure S1d). Constant biomass loss could testify that the consumption of removed ammonium was mainly for energy conservation, confirming the occurrence of catabolic AN reactions. Unlike AN, ammonium conversion via HN is not linked to energy generation [44]. Contrasting microbial community structure of AN_AS and HN_AS (see Section 3.2) could support this argument.
3.1.3. Biological Phosphate Removal
The AS biomass was subjected to shifting anaerobic/aerobic condition by switching aerator on–off (see DO concentration profile in Figure S2c). This oxygenation strategy was adopted to subject the microbial community to different redox conditions to investigate the AS biological phosphate removal capacity in the AN/O_AS bioreactor. Since a single PAO should experience anaerobic and aerobic conditions in sequence, such oxygenation pattern was defined to trigger biological phosphate removal by PAOs if they existed in the microbial culture. Between 1.3–2.3 h, the P profile showed an increasing trend (Figure 1a). However, the P release was not considerable and could not increase the initial P concentration of 38 mg/L more than 3%. Minor anaerobic release could be due to significantly low population of Accumulibacter in the inoculum (5.1%, see Section 3.2 and Figure S3a), compared to its population in robust EBPR processes (e.g., 20–60% of total bacteria [17,45]). During the rest of the anaerobic phase, reduction in P concentration happened, which is not characteristic of EBPR biomass [46]. During the subsequent aerobic phase (hours 4 to 5), the P concentration remained almost constant.
In a second attempt, sparging air into the bioreactor was stopped from hour 9 to 48 (Figure 2a), to possibly encourage the activation of PAOs under an extended anaerobic phase. Yet, the biomass constantly showed P uptake activity. Upon resuming aeration at hour 48, rapid reduction in P started which lasted up to 72 h, during which COD mineralization was under progress as well (Figure S2b). Upon complete depletion of biodegradable organic matter, the P removal pace slowed down afterward, which suggested that the P removal was potentially linked to bacterial assimilation in the AN/O_AS bioreactor. Biomass growth occurred during the second anaerobic phase (from 24 to 48 h) and the following aerobic phase, until COD was depleted (Figure S2a).
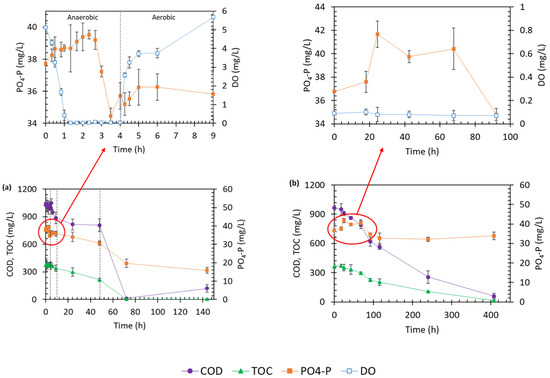
Figure 2.
Treatment performance of (a) alternating anaerobic/aerobic (AN/O_AS) and (b) extended anaerobic activated sludge (ExAN_AS) bioreactors.
The AS experiment was repeated under an extended anaerobic condition (i.e., ExAN_AS bioreactor) to provide further evidence regarding P removal. Figure 2b illustrates the changes in P and organic matter concentrations. A slight P release was observed initially; P concentration rose from 36.8 to 41.7 mg/L in 24 h, which corresponded to a 13.3% increase. However, P release did not continue during the rest of the operation. Once again, such an increase was not comparable to anaerobic P release capacity of biomass with high PAO population and tangible EBPR capacity [17,46]. Reduction in P concentration coincided with COD and NH4–N removal, implying a relation between P removal and biomass growth in this reactor as well. Biomass concentration gradually increased up to hour 116 (Figure S2d). Degradation of about half (42%) of COD concentration and partial removal (32%) of NH4–N concentration coincided with biomass growth during the first 116 h. Removal of ammonium and P stopped when biomass growth ceased at 116 h. Thus, the P removal in the ExAN_AS reactor could be attributed to biomass assimilation.
Overall, an ultimate P removal efficiency of 11% was achieved in the ExAN_AS bioreactor, while in the AN/O_AS bioreactor, initial P concentration of 37.7 mg/L was reduced to 15.8 mg/L, with a removal efficiency of 58%. Under aerobic conditions (i.e., HN_AS and AN_AS bioreactors where P was provided as a supplementary nutrient for nitrification), the AS could remove 27 mg P/L at high organic loading in HN_AS and 10 mg P/L in the COD-deficient environment of AN_AS, corresponding to 70 and 30% removal efficiencies, respectively. Consumption of 10 mg P/L in AN_AS, considering the organic-deficient environment of this reactor alongside no biomass growth (see Figure S1d), could reject the possibility that P removal was solely due to biomass growth in either all of these bioreactors or, at least, only in AN_AS.
3.1.4. Aerobic Granular Sludge Performance
AGS performance is discussed elsewhere [26,30]. Briefly, in a single cycle of the GSBR bioreactors, similar nitrogen removal performances were observed despite distinct operation modes. The NOx− (i.e., NO3− + NO2−) did not accumulate entirely, and thus the profiles of TN and NH4–N were intertwined. Identical TN and NH4–N removal efficiencies were therefore achieved (61 and 67% in AN/O/AX/O_SBR and 48 and 49% in O_SBR, respectively). P release/uptake did not happen in the GSBRs. Continuous P reduction occurred during the entire cycle, even during the anaerobic phase of AN/O/AX/O_SBR, a behavior which was shown by the AS experiments (see the previous section). The P decrease followed the same trend as those of COD and ammonium profiles, suggesting a connection between P removal and heterotrophic activity in the GSBRs. COD and P removal efficiencies were 99 and 60 in AN/O/AX/O_ SBR and 97 and 41% in O_ SBR, respectively.
3.2. Changes in Microbial Community Structure and Population
Phylogenetic analysis showed substantial structural changes in microbial communities under different selection pressures applied in this study (Figure 3). The readers are recommended to refer to the pie charts presented in Supplementary Materials for convenient chart reading and data extraction. The results are summarized in Table 3, where putative HN, AN, and PAOs are identified and important observations and supporting evidence are highlighted. Drawing from HN_AS and AN_AS results, dominant nitrifying organisms were identified in the AGS. However, this approach is speculative and requires further study on BNR processes to gather comprehensive, large databases which can allow establishing a solid relation between process performance and microbial structure.
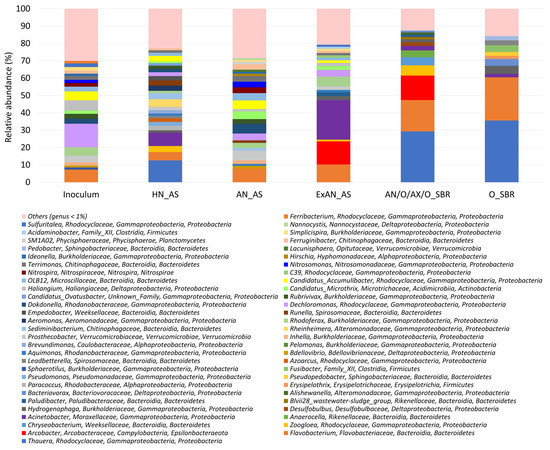
Figure 3.
Microbial selection and population dynamics at genus-level inoculum, heterotrophic nitrification (HN_AS), autotrophic nitrification (AN_AS), extended anaerobic activated sludge (ExAN_AS) bioreactors, as well as granular biomass bioreactors (i.e., AN/O/AX/O_SBR and O_SBR). The legend shows genus, family, and phylum.
Table 3.
Dominant bacterial species under different selection pressures.
4. Conclusions and Directions for Future Research
The HN and AN potentially occurred in aerobic organic-rich (HN_AS) and organic-deficient (AN_AS) environments, respectively. This speculation was supported by treatment performance, enrichment of certain functional bacterial groups, and microbial growth patterns of these bioreactors.
Thauera, Acinetobacter, Flavobacterium, and Rheinheimera (4–13%) were identified as potential HN bacteria, and Nitrosomonas (3.2%) and Nitrospira (3.4%) as potential AN bacteria. Some unconventional PAOs were isolated from ExAN_AS biomass, such as Acinetobacter (23%), Dechloromonas (3.9%), Microthirx (2.1%), and Psuedomonas (0.2%), whose PAO abilities are still open to debate. Conventional PAO Accumulibacter did not enrich in any of the bioreactors in this study.
The HN_AS biomass could simultaneously remove high COD (99%) and organic nitrogen (87%) concentrations within 43 h. As such, it was suspected that bacterial assimilation was a co-contributor to nitrogen removal in HN reactions, and that HN became the principal nitrogen removal pathway (2.7 mg TN/L/h) when COD concentration became relatively scarce (C/N = 7).
Putative HN bacteria (i.e., Thauera (29–36%) and Flavobacterium (18–25%)) were enriched in AGS regardless of the granular reactor operation mode. Enrichment of HN organisms in the AGS was suspected to be mainly due to granulation, possibly because of the floc-forming ability of HN species.
This study should be seen as a starting point for the investigation of microbial structure of AS- and AGS-based BNR processes, and thus, the roles attributed to the isolated species warrant further research through systematic experimentation (e.g., factorial experiment design, multivariate statistical analysis, and total Kjeldahl nitrogen (TKN) testing).
Data collected using metagenomic analyses adopted in this study (16S rRNA phylogenetic gene sequencing) should be complemented with functional analysis using metaproteomic studies, especially focusing on functional genes responsible for BNR, to provide greater insight and add depth to the conclusions of this study. The scope of this study was mainly focused on short-term batch assays to investigate changes in microbial structure, using seed microbial culture as a benchmark, in response to the imposed operating conditions. Monitoring bacterial community in long-term experiments can be considered in future investigations to add the factor of time as an additional independent variable and to observe temporal variations in the microbial composition (i.e., structure and populations). Monitoring temporal changes in microbial composition of BNR processes can provide grounding for further statistical analysis, where the main and combined effects of time and operating conditions on microbial composition can be established.
In this study, changes in microbial communities under different conditions were based on single analysis of 16S rRNA gene. Functional gene analysis alongside long-term experiments can help in collecting a larger database, which can help in verifying the conclusions of this study.
Moreover, incorporating TKN testing in the sampling and analysis regiment can provide additional insight about transformation of organic nitrogen to inorganic nitrogen species. TKN analysis in combination with functional gene analyses can provide solid evidence regarding heterotrophic nitrification.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/nitrogen2040032/s1. Figure S1. Concentration profiles of soluble constituents, biomass, and environmental factors in (a–c) heterotrophic nitrification (HN) and (d–f) autotrophic nitrification (AN) suspended-biomass batch bioreactors; Figure S2. Concentration profiles of soluble constituents, biomass, and environmental factors in (a–c) alternating anaerobic/aerobic (BioP_AN/O) and (d–f) extended biomass (BioP_ExAN) suspended-biomass batch bioreactors; Figure S3. Bacterial assembly of (a) inoculum; (b) heterotrophic nitrification (HN) AS biomass; (c) autotrophic nitrification (AN) AS biomass; (d) extended anaerobic (BioP_ExAN) AS biomass; (e) alternating (AN/O/AX/O_SBR) granular (AGS) biomass; and (f) purely aerobic (O_SBR) granular (AGS) biomass at genus level (relative abundances 1%).
Author Contributions
R.P.: conceptualization, data curation, investigation, methodology, visualization, writing—original draft, writing—review and editing; J.A.D.: conceptualization, investigation, validation, writing—review and editing; J.H.T.: funding acquisition, supervision; A.C.: supervision. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Natural Sciences and Engineering Research Council of Canada (NSERC, grant # 10011088).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are mainly available in this article and its Supplementary Materials. Additional information and data can be found in Pishgar et al. [25,26,27,28,29] and Pishgar [30].
Acknowledgments
This study was funded by Natural Sciences and Engineering Research Council of Canada (NSERC) and the revision process was supported by Associated Engineering Alberta Ltd. The authors would like to thank the Centre for Health Genomics and Informatics in the Cumming School of Medicine, University of Calgary for their assistance in the microbial analysis.
Conflicts of Interest
Hereby, the authors declare that the content of this article is subject to no conflict of interest.
Abbreviations
| AS | Activated sludge |
| AGS | Aerobic granular sludge |
| BNR | Biological nutrient removal |
| N | Nitrogen |
| P | Phosphorus |
| AN | Autotrophic nitrification |
| AXD | Anoxic denitrification |
| PAOs | Polyphosphate accumulating organisms |
| HN | Heterotrophic nitrification |
| Anammox | Anaerobic ammonium oxidation |
| AD | Aerobic denitrification |
| SND | Simultaneous nitrification–denitrification |
| PHAs | Polyhydroxyalkanoates |
| Poly-P | Polyphosphate |
| Ortho-P | Orthophosphate |
| EBPR | Enhanced biological phosphate removal |
| DO | Dissolved oxygen |
| AN/O/AX/O_SBR | Alternating anaerobic/aerobic/anoxic/aerobic granular sequencing batch reactor |
| GSBR | Granular sequencing batch reactor |
| SBR | Sequencing batch reactor |
| O_SBR | purely aerobic granular sequencing batch reactors |
| HN_AS | Heterotrophic nitrification activated sludge reactor |
| AN_AS | Autotrophic nitrification activated sludge reactor |
| AN/O_AS | Alternating anaerobic/aerobic activated sludge reactor |
| ExAN_AS | Extended anaerobic activated sludge reactor |
| COD | Chemical oxygen demand |
| TOC | Total organic carbon |
| TN | Total nitrogen |
| C/N | Carbon to nitrogen |
| MLSS | Mixed liquor suspended solids |
References
- Zhao, Y.; Huang, J.; Zhao, H.; Yang, H. Microbial community and N removal of aerobic granular sludge at high COD and N loading rates. Bioresour. Technol. 2013, 143, 439–446. [Google Scholar] [CrossRef]
- Limpiyakorn, T.; Sonthiphand, P.; Rongsayamanont, C.; Polprasert, C. Abundance of amoA genes of ammonia-oxidizing archaea and bacteria in activated sludge of full-scale wastewater treatment plants. Bioresour. Technol. 2011, 102, 3694–3701. [Google Scholar] [CrossRef]
- He, T.; Xie, D.; Li, Z.; Ni, J.; Sun, Q. Ammonium stimulates nitrate reduction during simultaneous nitrification and denitrification process by Arthrobacter arilaitensis Y-10. Bioresour. Technol. 2017, 239, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Su, J.; Zhang, K.; Huang, T.; Wen, G.; Guo, L.; Yang, S. Heterotrophic nitrification and aerobic denitrification at low nutrient conditions by a newly isolated bacterium, Acinetobacter sp. SYF26. Microbiology 2015, 161, 829–837. [Google Scholar] [CrossRef]
- Ren, Y.X.; Yang, L.; Liang, X. The characteristics of a novel heterotrophic nitrifying and aerobic denitrifying bacterium, Acinetobacter junii YB. Bioresour. Technol. 2014, 171, 1–9. [Google Scholar] [CrossRef]
- He, T.; Li, Z.; Sun, Q.; Xu, Y.; Ye, Q. Heterotrophic nitrification and aerobic denitrification by Pseudomonas tolaasii Y-11 without nitrite accumulation during nitrogen conversion. Bioresour. Technol. 2016, 200, 493–499. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Yang, J.; Wang, X.; Wang, E.; Li, B.; He, R.; Yuan, H. Removal of nitrogen by heterotrophic nitrification-aerobic denitrification of a phosphate accumulating bacterium Pseudomonas stutzeri YG-24. Bioresour. Technol. 2015, 182, 18–25. [Google Scholar] [CrossRef]
- Zhao, B.; An, Q.; He, Y.L.; Guo, J.S. N2O and N2 production during heterotrophic nitrification by Alcaligenes faecalis strain NR. Bioresour. Technol. 2012, 116, 379–385. [Google Scholar] [CrossRef]
- Yang, X.P.; Wang, S.M.; Zhang, D.W.; Zhou, L.X. Isolation and nitrogen removal characteristics of an aerobic heterotrophic nitrifying-denitrifying bacterium, Bacillus subtilis A1. Bioresour. Technol. 2011, 102, 854–862. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Pan, L.; Lv, N.; Tang, X. Characterization of novel Bacillus strain N31 from mariculture water capable of halophilic heterotrophic nitrification–aerobic denitrification. J. Biosci. Bioeng. 2017, 124, 564–571. [Google Scholar] [CrossRef]
- Su, J.J.; Liu, B.Y.; Liu, C.Y. Comparison of aerobic denitrification under high oxygen atmosphere by Thiosphaera pantotropha ATCC 35512 and Pseudomonas stutzeri SU2 newly isolated from the activated sludge of a piggery wastewater treatment system. J. Appl. Microbiol. 2001, 90, 457–462. [Google Scholar] [CrossRef] [PubMed]
- Cui, M.; Chen, F.; Fu, J.; Sheng, G.; Sun, G. Microbial metabolism of quinoline by Comamonas sp. World J. Microbiol. Biotechnol. 2004, 20, 539–543. [Google Scholar] [CrossRef]
- de Kreuk, M.K.; Heijnen, J.J.; van Loosdrecht, M.C.M. Simultaneous COD, nitrogen, and phosphate removal by aerobic granular sludge. Biotechnol. Bioeng. 2005, 90, 761–769. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Ji, M.; Li, R.; Qin, F. Organic and nitrogen removal from landfill leachate in aerobic granular sludge sequencing batch reactors. Waste Manag. 2012, 32, 448–455. [Google Scholar] [CrossRef]
- Zhang, S.; Sun, X.; Fan, Y.; Qiu, T.; Gao, M.; Wang, X. Heterotrophic nitrification and aerobic denitrification by Diaphorobacter polyhydroxybutyrativorans SL-205 using poly(3-hydroxybutyrate-co-3-hydroxyvalerate) as the sole carbon source. Bioresour. Technol. 2017, 241, 500–507. [Google Scholar] [CrossRef]
- Mino, T.; van Loosdrecht, M.C.M.; Heijnen, J.J. Microbiology and biochemistry of the enhanced biological phosphate removal process. Water Res. 1998, 32, 3193–3207. [Google Scholar] [CrossRef]
- Pijuan, M.; Guisasola, A.; Baeza, J.A.; Carrera, J.; Casas, C.; Lafuente, J. Aerobic phosphorus release linked to acetate uptake: Influence of PAO intracellular storage compounds. Biochem. Eng. J. 2005, 26, 184–190. [Google Scholar] [CrossRef]
- Guisasola, A.; Pijuan, M.; Baeza, J.A.; Carrera, J.; Casas, C.; Lafuente, J. Aerobic phosphorus release linked to acetate uptake in bio-P sludge: Process modeling using oxygen uptake rate. Biotechnol. Bioeng. 2004, 85, 722–733. [Google Scholar] [CrossRef] [PubMed]
- Oehmen, A.; Lemos, P.C.; Carvalho, G.; Yuan, Z.; Keller, J.; Blackall, L.L.; Reis, M.A.M. Advances in enhanced biological phosphorus removal: From micro to macro scale. Water Res. 2007, 41, 2271–2300. [Google Scholar] [CrossRef]
- Nguyen, H.T.T.; Le, V.Q.; Hansen, A.A.; Nielsen, J.L.; Nielsen, P.H. High diversity and abundance of putative polyphosphate-accumulating Tetrasphaera-related bacteria in activated sludge systems. FEMS Microbiol. Ecol. 2011, 76, 256–267. [Google Scholar] [CrossRef]
- Tarayre, C.; Charlier, R.; Delepierre, A.; Brognaux, A.; Bauwens, J.; Francis, F.; Dermience, M.; Lognay, G.; Taminiau, B.; Daube, G.; et al. Looking for phosphate-accumulating bacteria in activated sludge processes: A multidisciplinary approach. Environ. Sci. Pollut. Res. 2017, 24, 8017–8032. [Google Scholar] [CrossRef] [PubMed]
- Kong, Y.; Xia, Y.; Nielsen, J.L.; Nielsen, P.H. Ecophysiology of a group of uncultured Gammaproteobacterial glycogen-accumulating organisms in full-scale enhanced biological phosphorus removal wastewater treatment plants. Environ. Microbiol. 2006, 8, 479–489. [Google Scholar] [CrossRef]
- Ferrera, I.; Sánchez, O. Insights into microbial diversity in wastewater treatment systems: How far have we come? Biotechnol. Adv. 2016, 34, 790–802. [Google Scholar] [CrossRef]
- Han, Y.-H.; Fu, T.; Wang, S.-S.; Yu, H.-T.; Xiang, P.; Zhang, W.-X.; Chen, D.-L.; Li, M. Efficient phosphate accumulation in the newly isolated Acinetobacter junii strain LH4. 3 Biotech 2018, 8, 1–12. [Google Scholar] [CrossRef]
- Pishgar, R.; Hamza, R.A.; Tay, J.H. Augmenting Lagoon Process Using Reactivated Freeze-dried Biogranules. Appl. Biochem. Biotechnol. 2017, 183, 137–154. [Google Scholar] [CrossRef] [PubMed]
- Pishgar, R.; Dominic, J.A.; Sheng, Z.; Tay, J.H. Denitrification performance and microbial versatility in response to different selection pressures. Bioresour. Technol. 2019, 281, 72–83. [Google Scholar] [CrossRef]
- Pishgar, R.; Dominic, J.A.; Sheng, Z.; Tay, J.H. Influence of operation mode and wastewater strength on aerobic granulation at pilot scale: Startup period, granular sludge characteristics, and effluent quality. Water Res. 2019, 160, 81–96. [Google Scholar] [CrossRef]
- Pishgar, R.; Dominic, J.A.; Tay, J.H.; Chu, A. Pilot-scale investigation on nutrient removal characteristics of mineral-rich aerobic granular sludge: Identification of uncommon mechanisms. Water Res. 2019, 168, 115151. [Google Scholar] [CrossRef] [PubMed]
- Pishgar, R.; Kanda, A.; Gress, G.R.; Gong, H.; Dominic, J.A.; Tay, J.H. Effect of aeration pattern and gas distribution during scale-up of bubble column reactor for aerobic granulation. J. Environ. Chem. Eng. 2018, 6, 6431–6443. [Google Scholar] [CrossRef]
- Pishgar, R. Nutrient Removal (Ammonium and Phosphate) Using Aerobic Granulation at Pilot Scale. Ph.D. Thesis, University of Calgary, Calgary, AB, Canada, 2019. [Google Scholar]
- Kim, M.; Jeong, S.-Y.; Yoon, S.J.; Cho, S.J.; Kim, Y.H.; Kim, M.J.; Ryu, E.Y.; Lee, S.-J. Aerobic Denitrification of Pseudomonas putida AD-21 at Different C/N Ratios. J. Biosci. Bioeng. 2008, 106, 498–502. [Google Scholar] [CrossRef]
- Joong, K.K.; Kyoung, J.P.; Kyoung, S.C.; Nam, S.W.; Park, T.J.; Bajpai, R. Aerobic nitrification-denitrification by heterotrophic Bacillus strains. Bioresour. Technol. 2005, 96, 1897–1906. [Google Scholar] [CrossRef]
- Joo, H.-S.; Hirai, M.; Shoda, M. Characteristics of ammonium removal by heterotrophic nitrification-aerobic denitrification by Alcaligenes faecalis No. 4. J. Biosci. Bioeng. 2005, 100, 184–191. [Google Scholar] [CrossRef]
- van Niel, E.W.J.; Arts, P.A.M.; Wesselink, B.J.; Robertson, L.A.; Kuenen, J.G. Competition between heterotrophic and autotrophic nitrifiers for ammonia in chemostat cultures. FEMS Microbiol. Lett. 1993, 102, 109–118. [Google Scholar] [CrossRef]
- Castignetti, D.; Hollocher, T.C. Heterotrophic nitrification among denitrifiers. Appl. Environ. Microbiol. 1984, 47, 620–623. [Google Scholar] [CrossRef]
- Castignetti, D.; Forgue, C.J.; Sclafan, L.A. Pathway of Oxidation of Pyruvic Oxime by a Heterotrophic Nitrifier of the Genus Alcaligenes: Evidence against Nitroethane as an Intermediate. Arch. Biochem. Biophys. 1986, 250, 228–232. [Google Scholar] [CrossRef]
- Eylar, O.R.; Schmidt, E.L. A survey of heterotrophic micro-organisms from soil for ability to form nitrite and nitrate. J. Gen. Microbiol. 1959, 20, 473–481. [Google Scholar] [CrossRef] [PubMed]
- Wrage, N.; Velthof, G.L.; Van Beusichem, M.L.; Oenema, O. Role of nitrifier denitrification in the production of nitrous oxide. Soil Biol. Biochem. 2001, 33, 1723–1732. [Google Scholar] [CrossRef]
- Zhang, J.; Sun, W.; Zhong, W.; Cai, Z. The substrate is an important factor in controlling the significance of heterotrophic nitrification in acidic forest soils. Soil Biol. Biochem. 2014, 76, 143–148. [Google Scholar] [CrossRef]
- American Public Health Association (APHA); American Water Works Association (AWWA); Water Environment Federation (WEF). Standard Methods for the Examination of Water and Wastewater, 22nd ed.; American Public Health Association: Washington, DC, USA, 2012. [Google Scholar]
- Zhang, J.; Wu, P.; Hao, B.; Yu, Z. Heterotrophic nitrification and aerobic denitrification by the bacterium Pseudomonas stutzeri YZN-001. Bioresour. Technol. 2011, 102, 9866–9869. [Google Scholar] [CrossRef]
- Sun, Y.; Feng, L.; Li, A.; Zhang, X.; Yang, J.; Ma, F. Ammonium assimilation: An important accessory during aerobic denitrification of Pseudomonas stutzeri T13. Bioresour. Technol. 2017, 234, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Joo, H.S.; Hirai, M.; Shoda, M. Piggery wastewater treatment using Alcaligenes faecalis strain No. 4 with heterotrophic nitrification and aerobic denitrification. Water Res. 2006, 40, 3029–3036. [Google Scholar] [CrossRef] [PubMed]
- Moir, J.W.B.; Wehrfritz, J.; Spiro, S.; Richardson, D.J. The biochemical characterization of a nover non-haem-iron hydroxylamine oxidase from Paracococcus denitrifiecans GB17. Biochem. J. 1996, 319, 823–827. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Gil, G.; Holliger, C. Dynamics of microbial community structure of and enhanced biological phosphorus removal by aerobic granules cultivated on propionate or acetate. Appl. Environ. Microbiol. 2011, 77, 8041–8051. [Google Scholar] [CrossRef]
- Bin, Z.; Ji, M.; Qiu, Z.; Liu, H.; Wang, J.; Li, J. Microbial population dynamics during sludge granulation in an anaerobic-aerobic biological phosphorus removal system. Bioresour. Technol. 2011, 102, 2474–2480. [Google Scholar] [CrossRef]
- Liu, B.; Zhang, F.; Feng, X.; Liu, Y.; Yan, X.; Zhang, X.; Wang, L.; Zhao, L. Thauera and Azoarcus as functionally important genera in a denitrifying quinoline-removal bioreactor as revealed by microbial community structure comparison. FEMS Microbiol. Ecol. 2006, 55, 274–286. [Google Scholar] [CrossRef]
- Liang, S.C.; Zhao, M.; Lu, L.; Wang, C.L.; Zhao, L.Y.; Liu, W.J. Isolation and characteristic of an aerobic denitrifier with high nitrogen removal efficiency. Afr. J. Biotechnol. 2011, 10, 10648–10656. [Google Scholar] [CrossRef]
- Strand, S.E.; McDonnell, A.J.; Unz, R.F. Oxygen and nitrate reduction kinetics of a nonflocculating strain of Zoogloea ramigera. Antonie Van Leeuwenhoek 1988, 255, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.L.; Zhou, S.L.; Zhang, H.H.; Bai, S.Y.; He, X.X.; Yang, X. Nitrogen removal characteristics of a newly isolated indigenous aerobic denitrifier from oligotrophic drinking water reservoir, Zoogloea sp. N299. Int. J. Mol. Sci. 2015, 16, 10038–10060. [Google Scholar] [CrossRef]
- Nielsen, P.H.; Mielczarek, A.T.; Kragelund, C.; Nielsen, J.L.; Saunders, A.M.; Kong, Y.; Hansen, A.A.; Vollertsen, J. A conceptual ecosystem model of microbial communities in enhanced biological phosphorus removal plants. Water Res. 2010, 44, 5070–5088. [Google Scholar] [CrossRef]
- Wells, G.F.; Park, H.D.; Yeung, C.H.; Eggleston, B.; Francis, C.A.; Criddle, C.S. Ammonia-oxidizing communities in a highly aerated full-scale activated sludge bioreactor: Betaproteobacterial dynamics and low relative abundance of Crenarchaea. Environ. Microbiol. 2009, 11, 2310–2328. [Google Scholar] [CrossRef]
- Berg, P.E.R.; Klemedtssn, L.; Rosswall, T. Inhibitory effect of low partial pressures of acetylene on nitrification. Soil Biol. Biochem. 1982, 14, 301–303. [Google Scholar] [CrossRef]
- Aravind, J.; Saranya, T.; Kanmani, P. Optimizing the production of Polyphosphate from Acinetobacter towneri. Glob. J. Environ. Sci. Manag. 2015, 1, 63–70. [Google Scholar] [CrossRef]
- Kong, Y.; Xia, Y.; Nielsen, J.L.; Nielsen, P.H. Structure and function of the microbial community in a full-scale enhanced biological phosphorus removal plant. Microbiology 2007, 153, 4061–4073. [Google Scholar] [CrossRef]
- Kim, J.M.; Lee, H.J.; Lee, D.S.; Jeon, C.O. Characterization of the denitrification-associated phosphorus uptake properties of “Candidatus Accumulibacter phosphatis” clades in sludge subjected to enhanced biological phosphorus removal. Appl. Environ. Microbiol. 2013, 79, 1969–1979. [Google Scholar] [CrossRef] [PubMed]
- Coats, E.R.; Brinkman, C.K.; Lee, S. Characterizing and contrasting the microbial ecology of laboratory and full-scale EBPR systems cultured on synthetic and real wastewaters. Water Res. 2017, 108, 124–136. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.; Schroeder, S.; Beer, M.; McIlroy, S.; Bayly, R.C.; May, J.W.; Vasiliadis, G.; Seviour, R.J. Ecology of the microbial community removing phosphate from wastewater under continuously aerobic conditions in a sequencing batch reactor. Appl. Environ. Microbiol. 2007, 73, 2257–2270. [Google Scholar] [CrossRef][Green Version]
- Horn, M.A.; Ihssen, J.; Matthies, C.; Schramm, A.; Acker, G.; Drake, H.L. Dechloromonas denitrificans sp. nov., Flavobacterium denitrificans sp. nov., Paenibacillus anaericanus sp. nov. and Paenibacillus terrae strain MH72, N2O-producing bacteria isolated from the gut of the earthworm Aporrectodea caliginosa. Int. J. Syst. Evol. Microbiol. 2005, 55, 1255–1265. [Google Scholar] [CrossRef]
- Yilmaz, G.; Lemaire, R.; Keller, J.; Yuan, Z. Simultaneous nitrification, denitrification, and phosphorus removal from nutrient-rich industrial wastewater using granular sludge. Biotechnol. Bioeng. 2008, 100, 529–541. [Google Scholar] [CrossRef] [PubMed]
- Bassin, J.P.; Kleerebezem, R.; Dezotti, M.; van Loosdrecht, M.C.M. Simultaneous nitrogen and phosphate removal in aerobic granular sludge reactors operated at different temperatures. Water Res. 2012, 46, 3805–3816. [Google Scholar] [CrossRef]
- Wang, J.; Qi, R.; Liu, M.; Li, Q.; Bao, H.; Li, Y.; Wang, S.; Tandoi, V.; Yang, M. The potential role of “Candidatus Microthrix parvicella” in phosphorus removal during sludge bulking in two full-scale enhanced biological phosphorus removal plants. Water Sci. Technol. 2014, 70, 367–375. [Google Scholar] [CrossRef]
- McIlroy, S.J.; Kristiansen, R.; Albertsen, M.; Karst, S.M.; Rossetti, S.; Nielsen, J.L.; Tandoi, V.; Seviour, R.J.; Nielsen, P.H. Metabolic model for the filamentous “Candidatus Microthrix parvicella” based on genomic and metagenomic analyses. ISME J. 2013, 7, 1161–1172. [Google Scholar] [CrossRef] [PubMed]
- Rashid, M.H.; Rumbaugh, K.; Passador, L.; Davies, D.G.; Hamood, A.N.; Iglewski, B.H.; Kornberg, A. Polyphosphate kinase is essential for biofilm development, quorum sensing, and virulence of Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2000, 97, 9636–9641. [Google Scholar] [CrossRef]
- Rashid, M.H.; Kornberg, A. Inorganic polyphosphate is needed for swimming, swarming, and twitching motilities of Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2002, 97, 4885–4890. [Google Scholar] [CrossRef]
- Kundu, P.; Pramanik, A.; Dasgupta, A.; Mukherjee, S.; Mukherjee, J. Simultaneous heterotrophic nitrification and aerobic denitrification by Chryseobacterium sp. R31 isolated from abattoir wastewater. Biomed. Res. Int. 2014, 2014, 436056. [Google Scholar] [CrossRef] [PubMed]
- Patureau, D.; Davison, J.; Bernet, N.; Moletta, R. Dentrification under various aeration conditions in Comomonas sp., strain SGLY2. FEMS Microbiol. Lett. 1994, 14, 71–78. [Google Scholar] [CrossRef]
- Wan, C.; Lee, D.-J.; Yang, X.; Wang, Y.; Wang, X.; Liu, X. Calcium precipitate induced aerobic granulation. Bioresour. Technol. 2015, 176, 32–37. [Google Scholar] [CrossRef]
- Wan, C.; Zhang, P.; Lee, D.J.; Yang, X.; Liu, X.; Sun, S.; Pan, X. Disintegration of aerobic granules: Role of second messenger cyclic di-GMP. Bioresour. Technol. 2013, 146, 330–335. [Google Scholar] [CrossRef]
- Adav, S.S.; Lee, D.J.; Lai, J.Y. Potential cause of aerobic granular sludge breakdown at high organic loading rates. Appl. Microbiol. Biotechnol. 2010, 85, 1601–1610. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).