Perennial Trees Associating with Nitrogen-Fixing Symbionts Differ in Leaf After-Life Nitrogen and Carbon Release


Abstract
1. Introduction
2. Materials and Methods
2.1. Litter Decomposition
2.2. Soil Respiration
2.3. Mineralization
3. Results
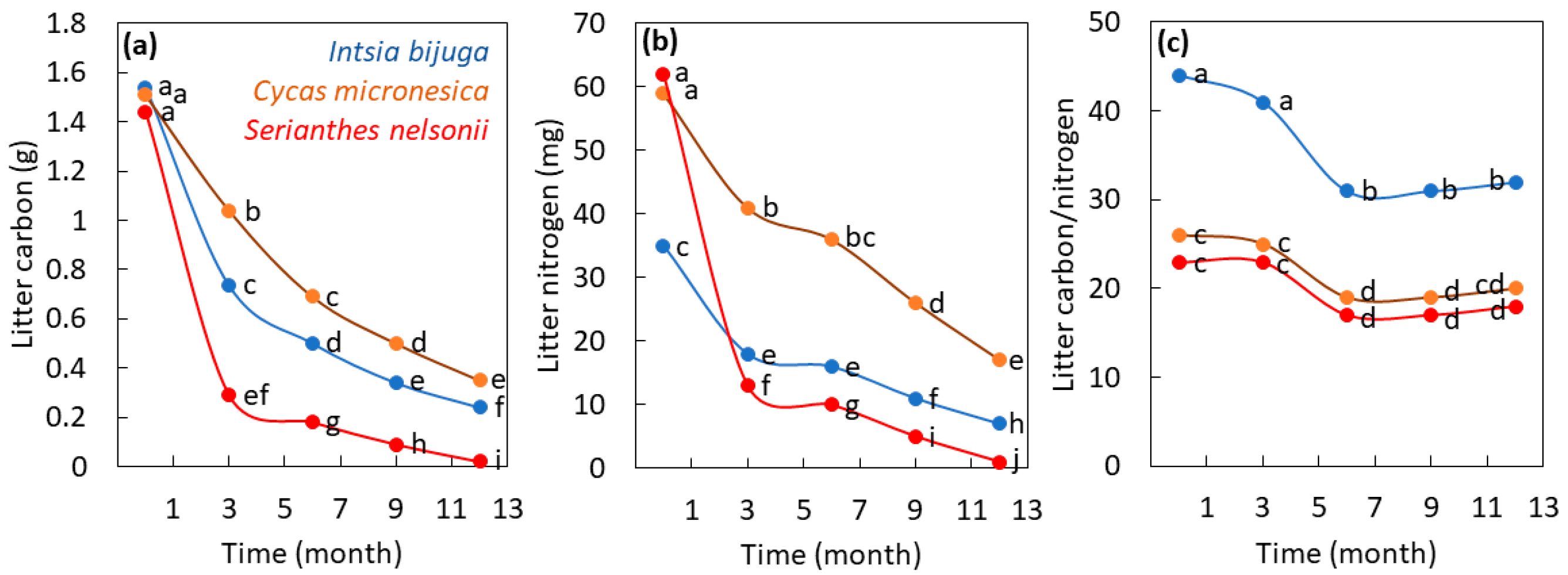
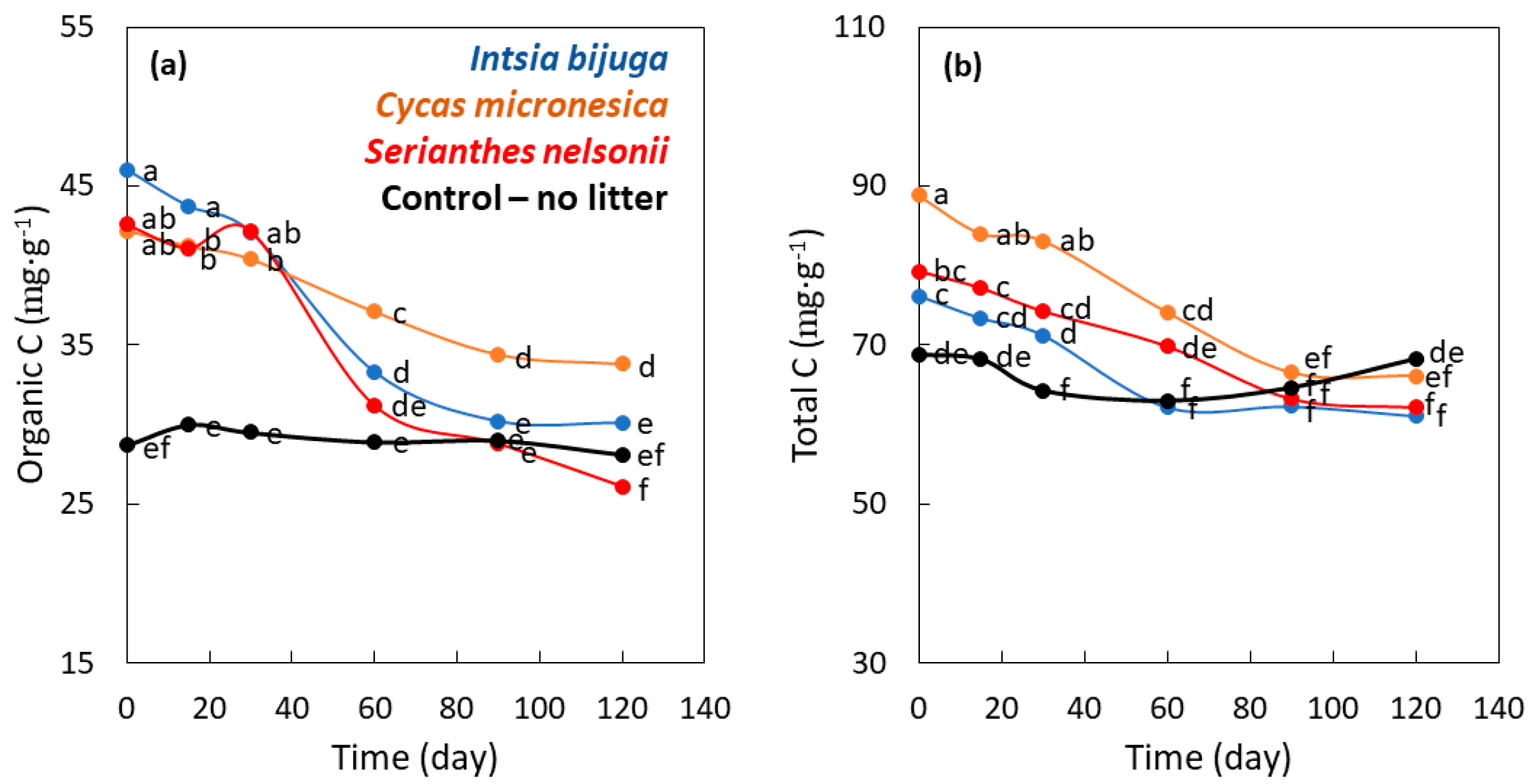
3.1. Litter Decomposition
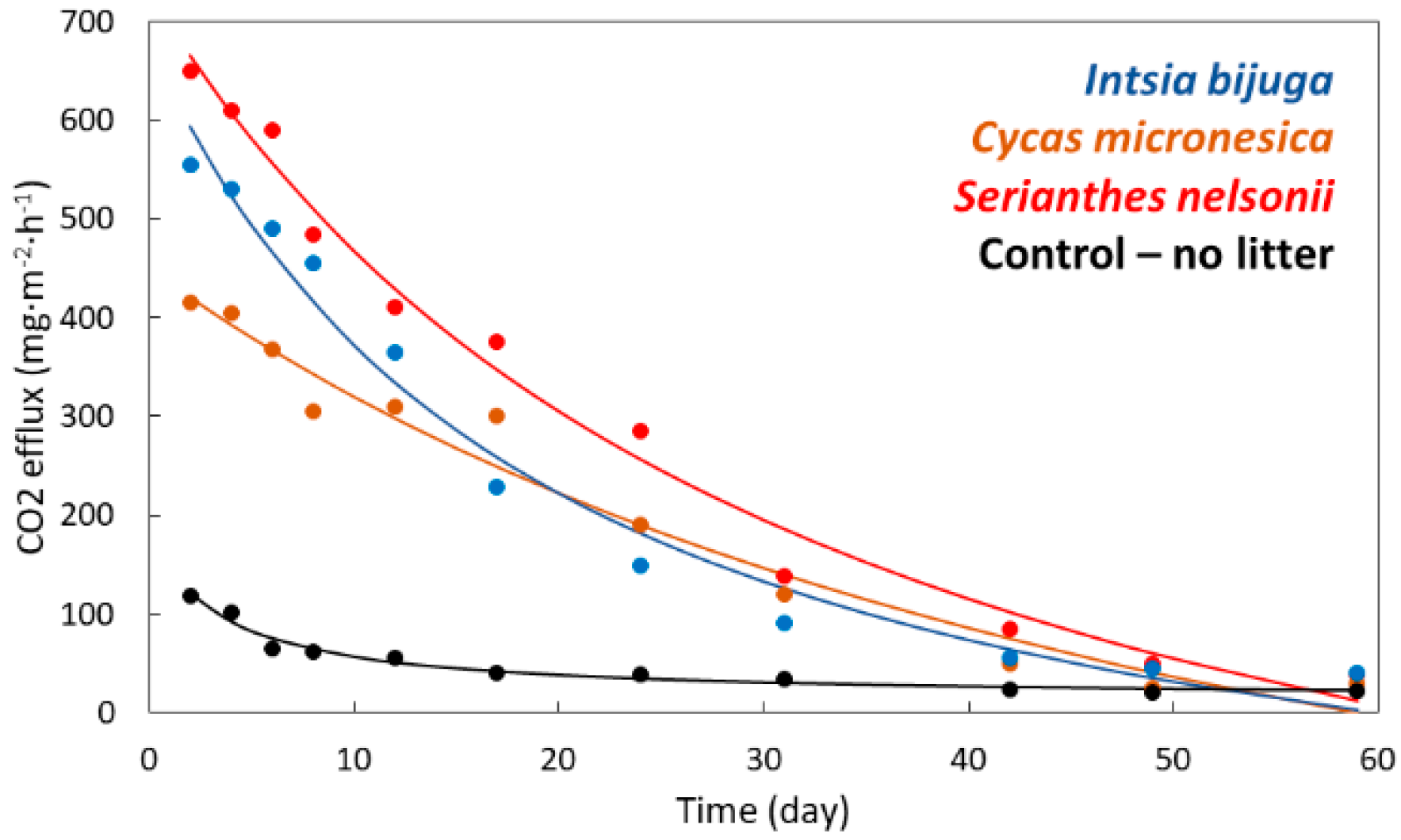
3.2. Soil Respiration
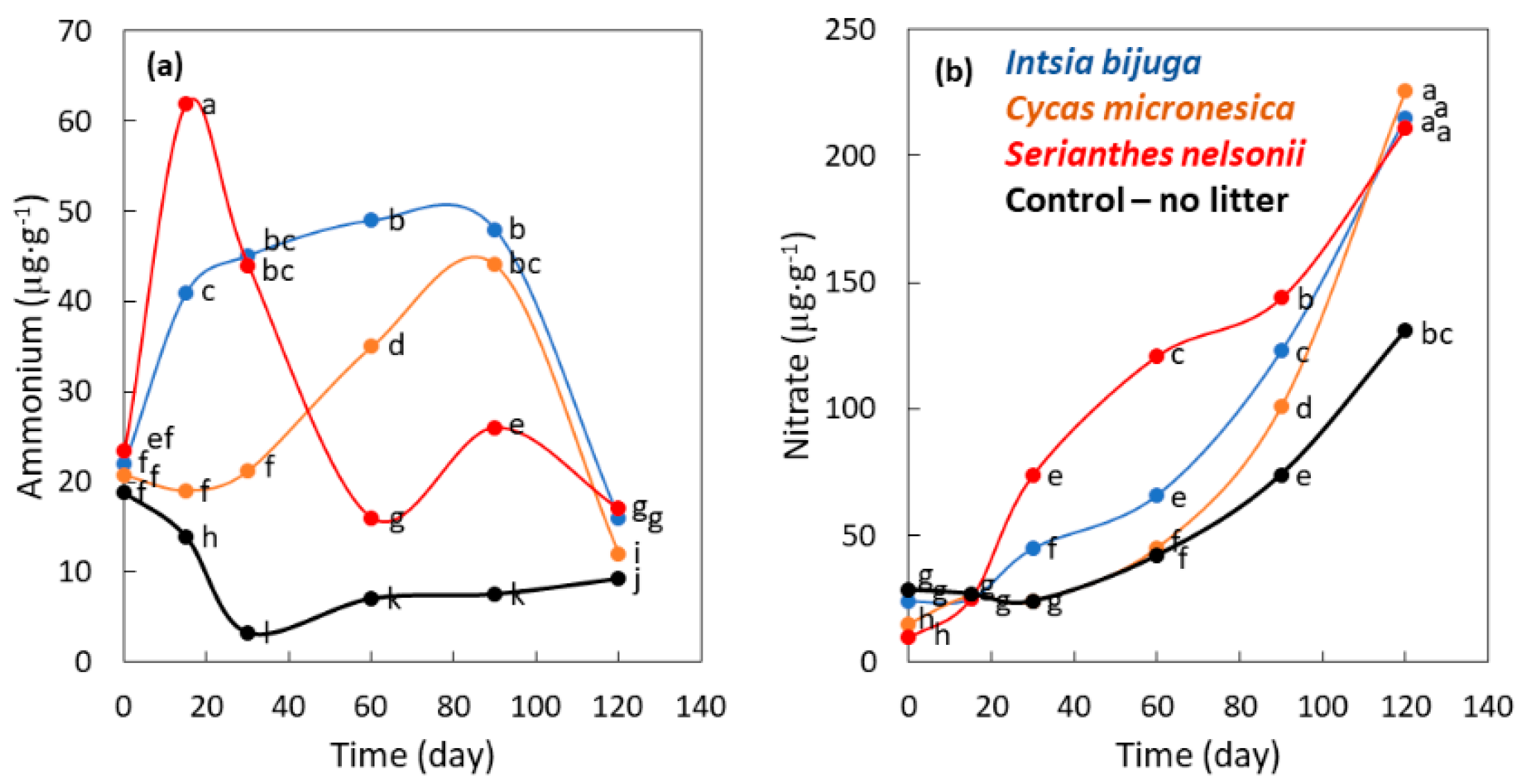
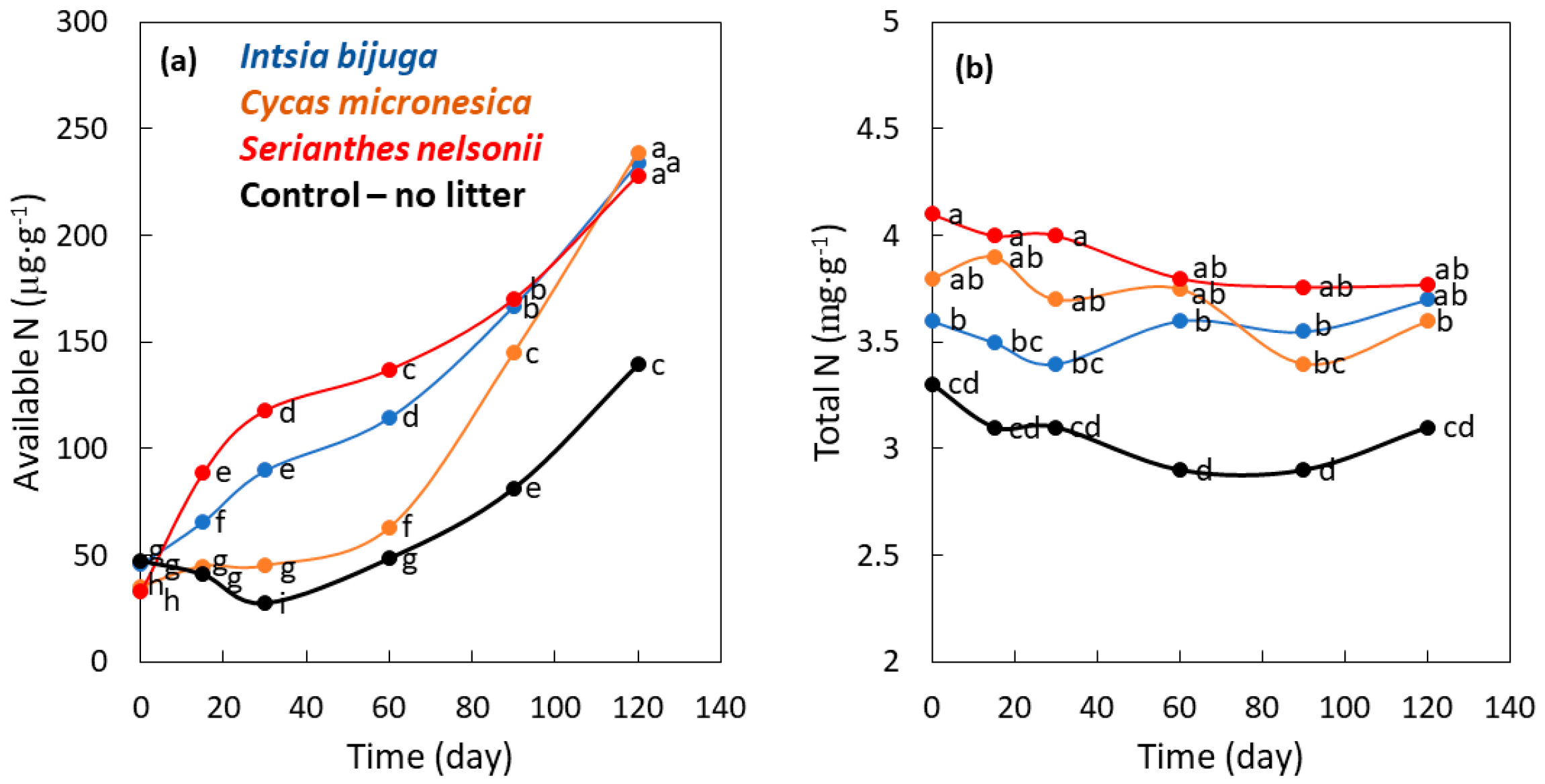
3.3. Mineralization
4. Discussion
Funding
Acknowledgments
Conflicts of Interest
References
- Postgate, J. Nitrogen Fixation, 3rd ed.; Cambridge University Press: Cambridge, UK, 1988. [Google Scholar]
- Smil, V. Cycles of Life: Civilization and the Biosphere; W.H. Freeman: New York, NJ, USA, 2000. [Google Scholar]
- Marler, T.E.; Terral, O. It is what it is, but it shouldn’t be: The science of ambiguity. HortScience 2014, 49, 1234–1236. [Google Scholar]
- Zahran, H.H. Rhizobium-legume symbiosis and nitrogen fixation under severe conditions and in an arid climate. Microbiol. Mol. Biol. Rev. 1999, 63, 968–989. [Google Scholar] [PubMed]
- Kuypers, M.M.M.; Marchant, H.K.; Kartal, B. The Microbial Nitrogen-Cycling Network. Nat. Rev. Microbiol. 2011, 16, 263–276. [Google Scholar]
- Latysheva, N.; Junker, V.L.; Palmer, W.J.; Codd, G.A.; Barker, D. The evolution of nitrogen fixation in cyanobacteria. Bioinformatics 2012, 28, 603–606. [Google Scholar]
- Norstog, K.J.; Nicholls, T.J. The Biology of the Cycads; Cornell University Press: Ithaca, NY, USA, 1997. [Google Scholar]
- Rai, A.N.; Söderbäck, E.; Bergman, B. Cyanobacterium-plant symbioses. N. Phytol. 2000, 147, 449–481. [Google Scholar]
- Khanna, P.K.; Raison, R.J. In situ core methods for estimating soil mineral-N fluxes: Re-evaluation based on 25 years of application and experience. Soil Biol. Biochem. 2013, 64, 203–210. [Google Scholar]
- Xie, Y. A meta-analysis of critique of litterbag method used in examining decomposition of leaf litters. J. Soils Sediments 2020, 20, 1881–1886. [Google Scholar]
- Pei, G.; Liu, J.; Peng, B.; Wang, C.; Jiang, P.; Bai, E. Nonlinear coupling of carbon and nitrogen release during litter decomposition and its responses to nitrogen addition. J. Geophys. Res. Biogeosci. 2020, 125, e2019JG005462. [Google Scholar] [CrossRef]
- Chapin, I.F.S.; Matson, P.A.; Vitousek, P. Principles of Terrestrial Ecosystem Ecology; Springer Science & Business Media: Berlin, Germany, 2011. [Google Scholar]
- World Conservation Monitoring Centre. Intsia bijuga. The IUCN Red List of Threatened Species. 1998. Available online: https://dx.doi.org/10.2305/IUCN.UK.1998.RLTS.T32310A9694485.en (accessed on 17 September 2020).
- Marler, T.E. Balancing growth and wood quality of Intsia bijuga under management: Complexity of silviculture conservation decisions. J. Trop. Forest Sci. 2015, 27, 429–434. [Google Scholar]
- Safford, W.E. The Useful Plants of the Island of Guam; Government Printing Office: Washington, DC, USA, 1905.
- Marler, T.; Haynes, J.; Lindstrom, A. Cycas Micronesica. The IUCN Red List of Threatened Species. 2010. Available online: https://dx.doi.org/10.2305/IUCN.UK.2010-3.RLTS.T61316A12462113.en (accessed on 17 September 2020).
- Marler, T.E.; Lawrence, J.H. Demography of Cycas micronesica on Guam following introduction of the armoured scale Aulacaspis yasumatsui. J. Tropical Ecol. 2012, 28, 233–242. [Google Scholar]
- Marler, T.E.; Krishnapillai, M.V. Longitude, forest fragmentation, and plant size influence Cycas micronesica mortality following island insect invasions. Diversity 2020, 12, 194. [Google Scholar] [CrossRef]
- Wiles, G.; Williams, E. Serianthes Nelsonii. The IUCN Red List of Threatened Species. 2017. Available online: https://dx.doi.org/10.2305/IUCN.UK.2017-3.RLTS.T30437A98715973.en (accessed on 17 September 2020).
- Merrill, E.D. Additions to the flora of Guam. Philipp. J. Sci. 1919, 15, 539–544. [Google Scholar]
- Marler, T.; Musser, C. Potential stressors leading to seedling mortality in the endemic Håyun lågu tree (Serianthes nelsonii Merr.) in the island of Guam. Trop. Conserv. Sci. 2015, 8, 738–744. [Google Scholar] [CrossRef]
- Marler, T.E.; Cascasan, A.N. Number of emerged seedlings and seedling longevity of the non-recruiting, critically endangered Håyun lågu tree Serianthes nelsonii Merr. (Fabales: Leguminosae) are influenced by month of emergence. J. Threatened Taxa 2015, 7, 8221–8225. [Google Scholar] [CrossRef]
- Marler, T.E.; Cruz, G.N. Extreme wind events influence seed rain and seedling dynamics of Guam’s Serianthes nelsonii Merr. Trop. Conserv. Sci. 2019, 12, 1–6. [Google Scholar] [CrossRef]
- Marler, T.E.; Lindström, A.J. The value of research to selling the conservation of threatened species: The case of Cycas micronesica (Cycadopsida: Cycadales: Cycadaceae). J. Threat. Taxa 2014, 6, 6523–6528. [Google Scholar] [CrossRef]
- Berkes, F.; Folke, C. (Eds.) Linking Social and Ecological Systems: Management Practices and Social Mechanisms for Building Resilience; Cambridge University Press: Cambridge, UK, 1998. [Google Scholar]
- Van Oudenhoven, A.P.E.; Martín-López, B.; Schröter, M.; de Groot, R. Advancing science on the multiple connections between biodiversity, ecosystems and people. Int. J. Biodivers. Sci. Ecosyst. Serv. Manag. 2018, 14, 127–131. [Google Scholar] [CrossRef]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.; Diemer, M.; et al. The world-wide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef]
- Wright, I.J.; Reich, P.B.; Cornelissen, J.H.C.; Falster, D.S.; Garnier, E.; Hikosaka, K.; Lamont, B.B.; Lee, W.; Oleksyn, J.; Osada, N.; et al. Assessing the generality of global leaf trait relationships. N. Phytol. 2005, 166, 485–496. [Google Scholar] [CrossRef]
- Young, F.J. Soil Survey of Territory of Guam; U. S. Dept. of Agric. Soil Conservation Service: Washington, DC, USA, 1988.
- Marler, T.E. Cycad aulacaspis scale invades the Mariana Islands. Mem. N. Y. Bot. Garden 2012, 106, 20–35. [Google Scholar]
- Dumas, J.B.A. Procedes de L’analyse Organique. Ann. Chim. Phys. 1831, 47, 198–205. [Google Scholar]
- Iiyama, K.; Wallis, A.F.A. Determination of lignin in herbaceous plants by an improved acetyl bromide procedure. J. Sci. Food Agric. 1990, 51, 145–161. [Google Scholar] [CrossRef]
- AOAC. Official Method 973. 18 fibre (acid detergent) and lignin in animal feed. In Official Methods of Analysis of AOAC International, 16th ed.; AOAC International: Arlington, VA, USA, 1997; pp. 28–29. [Google Scholar]
- Ayres, E.; Steltzer, H.; Simmons, B.L.; Simpson, R.T.; Steinweg, J.M.; Wallenstein, M.D.; Mellor, N.; Parton, W.J.; Moore, J.C.; Wall, D.H. Home-field advantage accelerates leaf litter decomposition in forests. Soil Biol. Biochem. 2009, 41, 606–610. [Google Scholar] [CrossRef]
- Lin, D.; Dou, P.; Yang, G.; Qian, S.; Wang, H.; Zhao, L.; Yang, Y.; Mi, X.; Ma, K.; Fanin, N. Home-field advantage of litter decomposition differs between leaves and fine roots. N. Phytol. 2020, 227, 995–1000. [Google Scholar] [CrossRef]
- Olsen, S.R.; Cole, C.V.; Watanabe, F.S.; Dean, L.A. Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate; U. S. Dept. of Agric.: Washington, DC, USA, 1954; p. 19.
- Berghage, R.D.; Krauskopf, D.M.; Warncke, D.D.; Widders, I. Micronutrient Testing of Plant Growth Media Extractant, Identification and Evaluation. Commun. Soil Sci. Plant Anal. 1987, 18, 1089–1109. [Google Scholar] [CrossRef]
- Eno, C.F. Nitrate production in the field by incubating the soil in polyethylene bags. Proc. Soil Sci. Soc. Amer. 1960, 24, 277–279. [Google Scholar] [CrossRef]
- Marler, T.E.; Dongol, N.; Cruz, G.N. Leucaena leucocephala and adjacent native limestone forest habitats contrast in soil properties on Tinian Island. Commun. Integr. Biol. 2016, 9, e1212792. [Google Scholar] [CrossRef]
- Nelson, D.W.; Sommers, L.E. Total carbon, organic carbon, and organic matter. In Methods of Soil Analysis. Part 3. Chemical Methods; Sparks, D.L., Ed.; SSSA Book Series No. 5; SSSA and ASA: Madison, WI, USA, 1996; pp. 961–1010. [Google Scholar]
- Cataldo, D.A.; Haroon, M.; Schrader, L.E.; Youngs, V.L. Rapid colorimetric determination of nitrate in plant tissue by nitration of salicylic acid. Commun. Soil Sci. Plant Anal. 1975, 6, 71–80. [Google Scholar] [CrossRef]
- Cornwell, W.K.; Cornelissen, J.H.C.; Amatangelo, K.; Dorrepaal, E.; Eviner, V.T.; Godoy, O.; Hobbie, S.E.; Hoorens, B.; Kurokawa, H.; Pérez-Harguindeguy, N.; et al. Plant species traits are the predominant control on litter decomposition rates within biomes worldwide. Ecol. Lett. 2008, 11, 1065–1071. [Google Scholar] [CrossRef]
- Zhang, D.; Hui, D.; Luo, Y.; Zhou, G. Rates of litter decomposition in terrestrial ecosystems: Global patterns and controlling factors. J. Plant Ecol. 2008, 1, 85–93. [Google Scholar] [CrossRef]
- Makkonen, M.; Berg, M.P.; Handa, I.T.; Hattenschwiler, S.; van Ruijven, J.; van Bodegom, P.M.; Aerts, R. Highly consistent effects of plant litter identity and functional traits on decomposition across a latitudinal gradient. Ecol. Lett. 2012, 15, 1033–1041. [Google Scholar] [CrossRef] [PubMed]
- Marler, T.E. Tropical cyclones and perennial species in the Mariana Islands. Hortscience 2001, 36, 264–268. [Google Scholar] [CrossRef]
- Marler, T.E. Pacific island tropical cyclones are more frequent and globally relevant, yet less studied. Front. Environ. Sci. 2014, 2, 42. [Google Scholar] [CrossRef]
- Stone, B.C. America’s Asiatic flora: The plants of Guam. Amer. Sci. 1971, 59, 308–319. [Google Scholar]
- Wilcots, M.E.; Taylor, B.N.; Kuprewicz, E.K.; Menge, D.N.L. Small traits with big consequences: How seed traits of nitrogen-fixing plants might influence ecosystem nutrient cycling. Oikos 2019, 128, 668–679. [Google Scholar] [CrossRef]
- Marler, T.E. Late successional tree species in Guam create biogeochemical niches. Commun. Integr. Biol. 2019, 12, 86–90. [Google Scholar] [CrossRef][Green Version]
- Marler, T.E.; Krishnapillai, M.V. Cycas micronesica trees alter local soil traits. Forests 2018, 9, 565. [Google Scholar] [CrossRef]
- Marler, T.E.; Calonje, M. Two cycad species affect the carbon, nitrogen, and phosphorus content of soils. Horticulturae 2020, 6, 24. [Google Scholar] [CrossRef]
- Hartmann, H.; Trumbore, S. Understanding the roles of nonstructural carbohydrates in forest trees—From what we can measure to what we want to know. New Phytol. 2016, 211, 386–403. [Google Scholar] [CrossRef]
- Phillips, C.L.; Bond-Lamberty, B.; Desai, A.R.; Lavoie, M.; Risk, D.; Tang, J.; Todd-Brown, K.; Vargas, R. The value of soil respiration measurements for interpreting and modeling terrestrial carbon cycling. Plant Soil 2017, 413, 1–25. [Google Scholar] [CrossRef]
- Cassart, B.; Basia, A.A.; Jonard, M.; Ponette, Q. Average leaf litter quality drives the decomposition of single-species, mixed-species and transplanted leaf litters for two contrasting tropical forest types in the Congo Basin (DRC). Ann. For. Sci. 2020, 77, 33. [Google Scholar] [CrossRef]
- Liu, J.; Liu, X.; Song, Q.; Compson, Z.G.; LeRoy, C.J.; Luan, F.; Wang, H.; Hu, Y.; Yang, Q. Synergistic effects: A common theme in mixed-species litter decomposition. New Phytol. 2020, 227, 757–765. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Butenschoen, O.; Barantal, S.; Handa, I.T.; Makkonen, M.; Vos, V.; Aerts, R.; Berg, M.P.; McKie, B.; Van Ruijven, J.; et al. Decomposition of leaf litter mixtures across biomes: The role of litter identity, diversity and soil fauna. J. Ecol. 2020. [Google Scholar] [CrossRef]
- Steinwandter, M.; Schlick-Steiner, B.C.; Steiner, F.M.; Seeber, J. One plus one is greater than two: Mixing litter types accelerates decomposition of low-quality alpine dwarf shrub litter. Plant Soil 2019, 438, 405–419. [Google Scholar] [CrossRef]
- Lodge, D.; Scatena, F.; Asbury, C.; Sanchez, M. Fine litterfall and related nutrient inputs resulting from Hurricane Hugo in subtropical wet and lower montane rain forests of Puerto Rico. Biotropica 1991, 23, 336–342. [Google Scholar] [CrossRef]
- Xu, X. Nutrient dynamics in decomposing needles of Pinus luchuensis after typhoon disturbance in a subtropical environment. Ann. For. Sci. 2006, 63, 707–713. [Google Scholar] [CrossRef]
- Liu, X.; Zeng, X.; Zou, X.; Lodge, D.J.; Stankavich, S.; González, G.; Cantrell, S.A. Responses of soil labile organic carbon to a simulated hurricane disturbance in a tropical wet forest. Forests 2018, 9, 420. [Google Scholar] [CrossRef]
- Marler, T.E.; Ferreras, U.F. Disruption of leaf nutrient remobilization in coastal Cycas trees by tropical cyclone damage. J. Geogr. Nat. Disasters 2015, 5, 1421–1427. [Google Scholar]
- Luo, Y.; Weng, E. Dynamic disequilibrium of the terrestrial carbon cycle under global change. Trends Ecol. Evol. 2011, 26, 96–104. [Google Scholar] [CrossRef]
- Marler, T.E.; Dongol, N. Three invasive insects alter Cycas micronesica leaf chemistry and predict changes in biogeochemical cycling. Commun. Integr. Biol. 2016, 9, e1208324. [Google Scholar] [CrossRef]
- Gandhi, K.J.K.; Herms, D.A. Direct and indirect effects of alien insect herbivores on ecological processes and interactions on forests of eastern North America. Biol. Invas. 2010, 12, 389–405. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiment | Nitrogen (mg·g−1) | Carbon (mg·g−1) | Phosphorus (µg·g−1) | Potassium (µg·g−1) | Calcium (mg·g−1) | Magnesium (µg·g−1) |
|---|---|---|---|---|---|---|
| Decomposition and mineralization | 12.3 | 177.3 | 60.3 | 571.2 | 12.1 | 1021.1 |
| Soil respiration | 12.6 | 174.2 | 61.5 | 579.8 | 11.8 | 1053.3 |
| Trait | Cycas micronesica | Intsia bijuga | Serianthes nelsonii | F | p |
|---|---|---|---|---|---|
| Nitrogen (mg·g−1) | 19.5 ± 0.7 a 1 | 11.4 ± 0.8 b | 20.6 ± 0.7 a | 45.52 | <0.001 |
| Carbon (mg·g−1) | 509.0 ± 9.9 | 495.2 ± 25.9 | 472.2 ± 22.4 | 0.82 | 0.461 |
| Lignin (mg·g−1) | 284.5 ± 7.3 a | 207.7 ± 14.5 b | 148.3 ± 7.0 c | 44.82 | <0.001 |
| Cellulose (mg·g−1) | 226.5 ± 8.1 a | 148.0 ± 10.4 b | 137.7 ± 6.7 b | 32.25 | <0.001 |
| Carbon/Nitrogen | 26.3 ± 1.2 b | 44.5 ± 4.5 a | 23.0 ± 0.8 c | 18.04 | <0.001 |
| Lignin/Nitrogen | 14.7 ± 0.4 b | 18.6 ± 1.8 a | 7.2 ± 0.2 c | 28.69 | <0.001 |
| Cellulose/Nitrogen | 11.7 ± 0.7 a | 13.2 ± 1.1 a | 6.7 ± 0.4 b | 17.72 | <0.001 |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marler, T.E. Perennial Trees Associating with Nitrogen-Fixing Symbionts Differ in Leaf After-Life Nitrogen and Carbon Release. Nitrogen 2020, 1, 111-124. https://doi.org/10.3390/nitrogen1020010
Marler TE. Perennial Trees Associating with Nitrogen-Fixing Symbionts Differ in Leaf After-Life Nitrogen and Carbon Release. Nitrogen. 2020; 1(2):111-124. https://doi.org/10.3390/nitrogen1020010
Chicago/Turabian StyleMarler, Thomas E. 2020. "Perennial Trees Associating with Nitrogen-Fixing Symbionts Differ in Leaf After-Life Nitrogen and Carbon Release" Nitrogen 1, no. 2: 111-124. https://doi.org/10.3390/nitrogen1020010
APA StyleMarler, T. E. (2020). Perennial Trees Associating with Nitrogen-Fixing Symbionts Differ in Leaf After-Life Nitrogen and Carbon Release. Nitrogen, 1(2), 111-124. https://doi.org/10.3390/nitrogen1020010