


Preparation and Preliminary Application of Epitope Peptide-Based Antibody against Toxoplasma gondii GRA3


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Parasites
2.2. Plasmids and Protein Expression
2.3. Main Reagents
2.4. Rabbit Immunization and Polyclonal Antibody Production
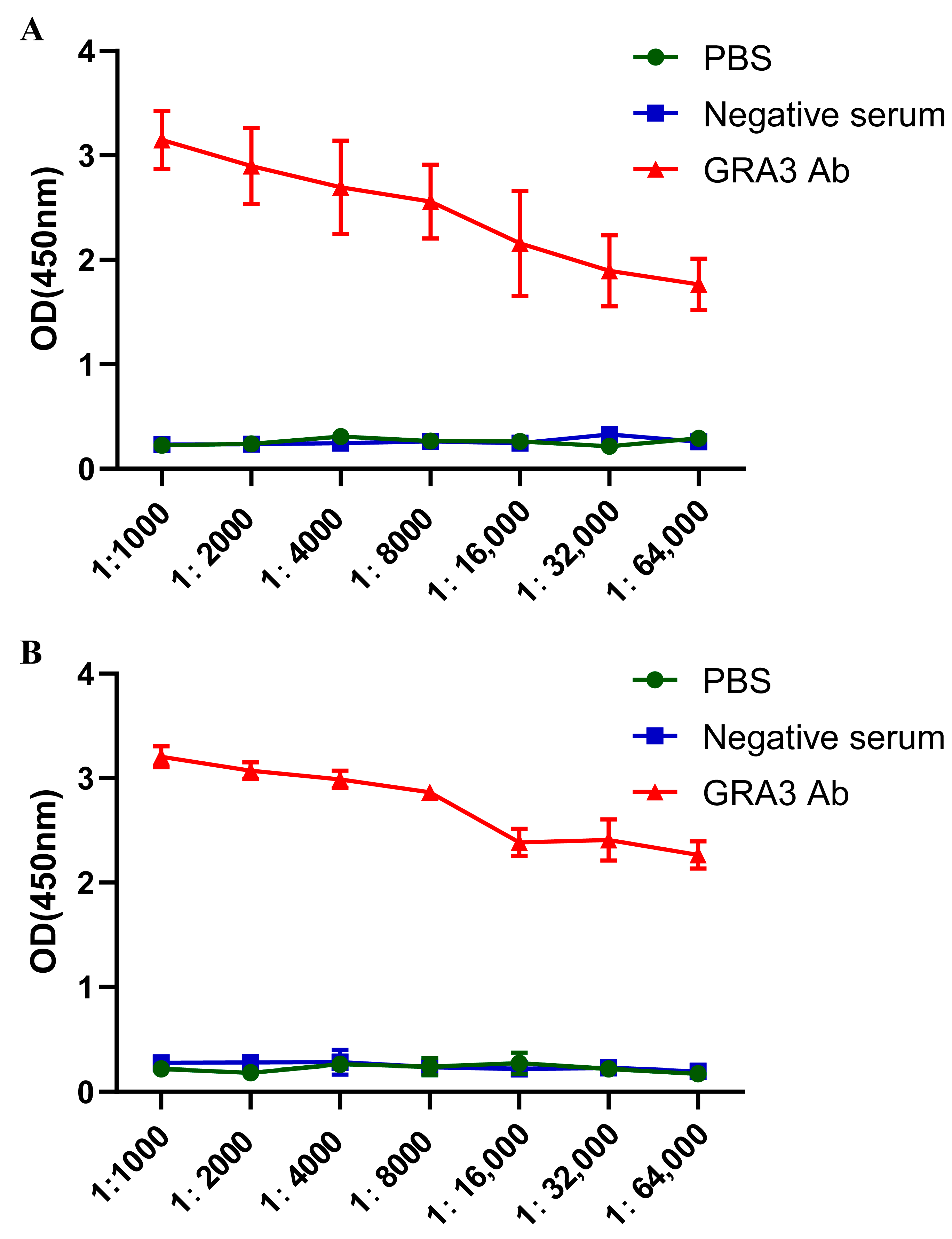
2.5. Determination of Antibody Potency
2.6. Antibody Specificity Assay
2.7. Application in Indirect Immunofluorescence Assay
2.8. Immunoprecipitation Assay
2.9. Statistical Analysis
3. Results
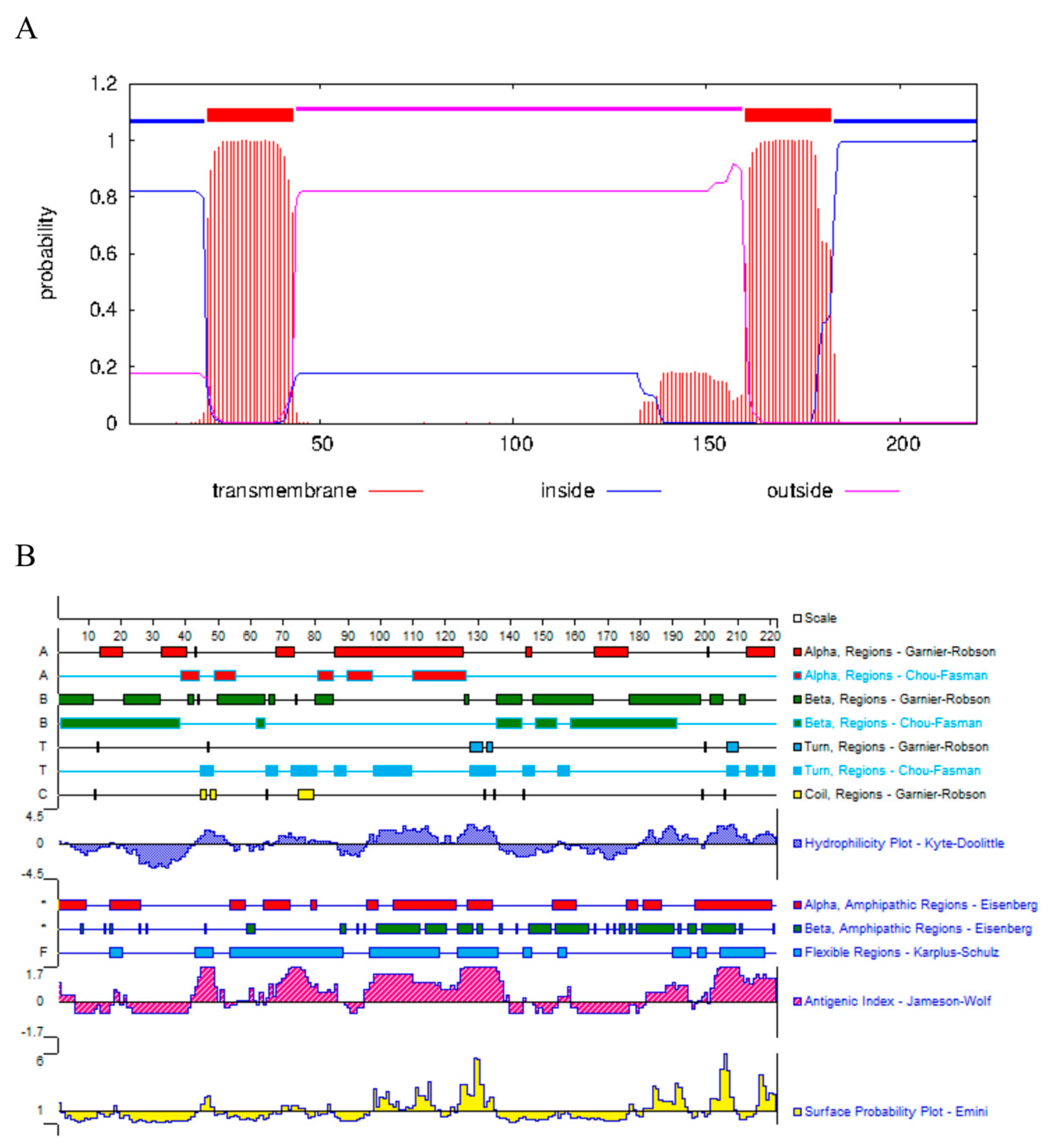
3.1. Peptide Design and Synthesis
3.2. Plasmid Construction and Determination of Antibody Potency
3.3. Determination of Antibody Potency
3.4. Antibody Specificity Assay
3.5. Application in Indirect Immunofluorescence Assay
3.6. Immunoprecipitation Assay
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lima, T.S.; Lodoen, M.B. Mechanisms of human innate immune evasion by Toxoplasma gondii. Front. Cell Infect. Microbiol. 2019, 9, 103. [Google Scholar] [CrossRef] [PubMed]
- Harker, K.S.; Ueno, N.; Lodoen, M.B. Toxoplasma gondii dissemination: A parasite’s journey through the infected host. Parasite Immunol. 2015, 37, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Inacio, P.; Zuzarte-Luis, V.; Ruivo, M.T.; Falkard, B.; Nagaraj, N.; Rooijers, K.; Mann, M.; Mair, G.; Fidock, D.A.; Mota, M.M. Parasite-induced er stress response in hepatocytes facilitates Plasmodium liver stage infection. EMBO Rep. 2015, 16, 955–964. [Google Scholar] [CrossRef]
- Hetz, C.; Mollereau, B. Disturbance of endoplasmic reticulum proteostasis in neurodegenerative diseases. Nat. Rev. Neurosci. 2014, 15, 233–249. [Google Scholar] [CrossRef]
- Kim, J.Y.; Ahn, H.J.; Ryu, K.J.; Nam, H.W. Interaction between parasitophorous vacuolar membrane-associated gra3 and calcium modulating ligand of host cell endoplasmic reticulum in the parasitism of Toxoplasma gondii. Korean J. Parasitol. 2008, 46, 209–216. [Google Scholar] [CrossRef]
- Holloway, M.P.; Bram, R.J. Co-localization of calcium-modulating cyclophilin ligand with intracellular calcium pools. J. Biol. Chem. 1998, 273, 16346–16350. [Google Scholar] [CrossRef]
- Craver, M.P.; Knoll, L.J. Increased efficiency of homologous recombination in Toxoplasma gondii dense granule protein 3 demonstrates that gra3 is not necessary in cell culture but does contribute to virulence. Mol. Biochem. Parasitol. 2007, 153, 149–157. [Google Scholar] [CrossRef]
- Mercier, C.; Adjogble, K.D.; Daubener, W.; Delauw, M.F. Dense granules: Are they key organelles to help understand the parasitophorous vacuole of all apicomplexa parasites? Int. J. Parasitol. 2005, 35, 829–849. [Google Scholar] [CrossRef]
- Black, M.W.; Boothroyd, J.C. Lytic cycle of Toxoplasma gondii. Microbiol. Mol. Biol. Rev. 2000, 64, 607–623. [Google Scholar] [CrossRef]
- Coppens, I.; Dunn, J.D.; Romano, J.D.; Pypaert, M.; Zhang, H.; Boothroyd, J.C.; Joiner, K.A. Toxoplasma gondii sequesters lysosomes from mammalian hosts in the vacuolar space. Cell 2006, 125, 261–274. [Google Scholar] [CrossRef]
- Sinai, A.P.; Webster, P.; Joiner, K.A. Association of host cell endoplasmic reticulum and mitochondria with the Toxoplasma gondii parasitophorous vacuole membrane: A high affinity interaction. J. Cell Sci. 1997, 110 (Pt 17), 2117–2128. [Google Scholar] [CrossRef] [PubMed]
- Meng, M.; Zhou, A.; Lu, G.; Wang, L.; Zhao, G.; Han, Y.; Zhou, H.; Cong, H.; Zhao, Q.; Zhu, X.Q.; et al. DNA prime and peptide boost immunization protocol encoding the Toxoplasma gondii gra4 induces strong protective immunity in balb/c mice. BMC Infect. Dis. 2013, 13, 494. [Google Scholar] [CrossRef] [PubMed]
- Min, J.; Qu, D.; Li, C.; Song, X.; Zhao, Q.; Li, X.A.; Yang, Y.; Liu, Q.; He, S.; Zhou, H. Enhancement of protective immune responses induced by Toxoplasma gondii dense granule antigen 7 (gra7) against toxoplasmosis in mice using a prime-boost vaccination strategy. Vaccine 2012, 30, 5631–5636. [Google Scholar] [CrossRef] [PubMed]
- Mercier, C.; Cesbron-Delauw, M.F. Toxoplasma secretory granules: One population or more? Trends Parasitol. 2015, 31, 60–71. [Google Scholar] [CrossRef]
- Scorza, T.; D’Souza, S.; Laloup, M.; Dewit, J.; De Braekeleer, J.; Verschueren, H.; Vercammen, M.; Huygen, K.; Jongert, E. A gra1 DNA vaccine primes cytolytic cd8(+) t cells to control acute Toxoplasma gondii infection. Infect. Immun. 2003, 71, 309–316. [Google Scholar] [CrossRef]
- Yin, H.; Zhao, L.; Wang, T.; Zhou, H.; He, S.; Cong, H. A Toxoplasma gondii vaccine encoding multistage antigens in conjunction with ubiquitin confers protective immunity to balb/c mice against parasite infection. Parasit. Vectors 2015, 8, 498. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, G.; Cai, J.; Yin, H. Review on the identification and role of Toxoplasma gondii antigenic epitopes. Parasitol. Res. 2016, 115, 459–468. [Google Scholar] [CrossRef]
- Xia, J.; Kong, L.; Zhou, L.J.; Wu, S.Z.; Yao, L.J.; He, C.; He, C.Y.; Peng, H.J. Genome-wide bimolecular fluorescence complementation-based proteomic analysis of Toxoplasma gondii rop18’s human interactome shows its key role in regulation of cell immunity and apoptosis. Front. Immunol. 2018, 9, 61. [Google Scholar] [CrossRef]
- Donnes, P.; Elofsson, A. Prediction of mhc class i binding peptides, using svmhc. BMC Bioinform. 2002, 3, 25. [Google Scholar] [CrossRef]
- Rosowski, E.E.; Lu, D.; Julien, L.; Rodda, L.; Gaiser, R.A.; Jensen, K.D.; Saeij, J.P. Strain-specific activation of the nf-kappab pathway by gra15, a novel Toxoplasma gondii dense granule protein. J. Exp. Med. 2011, 208, 195–212. [Google Scholar] [CrossRef] [PubMed]
- Ahn, H.J.; Kim, S.; Kim, H.E.; Nam, H.W. Interactions between secreted gra proteins and host cell proteins across the paratitophorous vacuolar membrane in the parasitism of Toxoplasma gondii. Korean J. Parasitol. 2006, 44, 303–312. [Google Scholar] [CrossRef]
- Henriquez, F.L.; Nickdel, M.B.; McLeod, R.; Lyons, R.E.; Lyons, K.; Dubremetz, J.F.; Grigg, M.E.; Samuel, B.U.; Roberts, C.W. Toxoplasma gondii dense granule protein 3 (gra3) is a type i transmembrane protein that possesses a cytoplasmic dilysine (kkxx) endoplasmic reticulum (er) retrieval motif. Parasitology 2005, 131, 169–179. [Google Scholar] [CrossRef]
- Dubey, J.P. The history of Toxoplasma gondii--the first 100 years. J. Eukaryot. Microbiol. 2008, 55, 467–475. [Google Scholar] [CrossRef]
- Su, C.; Evans, D.; Cole, R.H.; Kissinger, J.C.; Ajioka, J.W.; Sibley, L.D. Recent expansion of Toxoplasma through enhanced oral transmission. Science 2003, 299, 414–416. [Google Scholar] [CrossRef]
- Chen, M.; Yao, L.; Zhou, L.; Yang, P.; Zou, W.; Xu, L.; Li, S.; Peng, H. Toxoplasma gondii rop18i inhibits host innate immunity through cgas-sting signaling. FASEB J. 2022, 36, e22171. [Google Scholar]
- Sibley, L.D.; Krahenbuhl, J.L.; Adams, G.M.; Weidner, E. Toxoplasma modifies macrophage phagosomes by secretion of a vesicular network rich in surface proteins. J. Cell Biol. 1986, 103, 867–874. [Google Scholar] [CrossRef]
- Michelin, A.; Bittame, A.; Bordat, Y.; Travier, L.; Mercier, C.; Dubremetz, J.F.; Lebrun, M. Gra12, a Toxoplasma dense granule protein associated with the intravacuolar membranous nanotubular network. Int. J. Parasitol. 2009, 39, 299–306. [Google Scholar] [CrossRef]
- Achbarou, A.; Mercereau-Puijalon, O.; Sadak, A.; Fortier, B.; Leriche, M.A.; Camus, D.; Dubremetz, J.F. Differential targeting of dense granule proteins in the parasitophorous vacuole of Toxoplasma gondii. Parasitology 1991, 103 Pt 3, 321–329. [Google Scholar] [CrossRef]
- Linder, E.; Thors, C.; Edberg, F.; Haglund, S.; von Bonsdorff, C.H. Generation of antibodies against Toxoplasma gondii antigen associated with dense granules and the parasitophorous vacuole of the host cell. Parasitol. Res. 1992, 78, 175–178. [Google Scholar] [CrossRef]
- Bermudes, D.; Joiner, K.A. The role of parasites in generating evolutionary novelty. Parasitol. Today 1993, 9, 458–463. [Google Scholar] [CrossRef]
- Leriche, M.A.; Dubremetz, J.F. Exocytosis of Toxoplasma gondii dense granules into the parasitophorous vacuole after host cell invasion. Parasitol. Res. 1990, 76, 559–562. [Google Scholar] [CrossRef]
- Bermudes, D.; Dubremetz, J.F.; Achbarou, A.; Joiner, K.A. Cloning of a cdna encoding the dense granule protein gra3 from Toxoplasma gondii. Mol. Biochem. Parasitol. 1994, 68, 247–257. [Google Scholar] [CrossRef]
- Li, M.; Mo, X.W.; Wang, L.; Chen, H.; Luo, Q.L.; Wen, H.Q.; Wei, W.; Zhang, A.M.; Du, J.; Lu, F.L.; et al. Phylogeny and virulence divergency analyses of Toxoplasma gondii isolates from china. Parasit. Vectors 2014, 7, 133. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, R.; Wu, M.; Cai, H.; An, R.; Chen, Y.; Wang, J.; Zhou, N.; Du, J. Preparation and Preliminary Application of Epitope Peptide-Based Antibody against Toxoplasma gondii GRA3. Trop. Med. Infect. Dis. 2023, 8, 143. https://doi.org/10.3390/tropicalmed8030143
Wang R, Wu M, Cai H, An R, Chen Y, Wang J, Zhou N, Du J. Preparation and Preliminary Application of Epitope Peptide-Based Antibody against Toxoplasma gondii GRA3. Tropical Medicine and Infectious Disease. 2023; 8(3):143. https://doi.org/10.3390/tropicalmed8030143
Chicago/Turabian StyleWang, Ru, Minmin Wu, Haijian Cai, Ran An, Ying Chen, Jie Wang, Nan Zhou, and Jian Du. 2023. "Preparation and Preliminary Application of Epitope Peptide-Based Antibody against Toxoplasma gondii GRA3" Tropical Medicine and Infectious Disease 8, no. 3: 143. https://doi.org/10.3390/tropicalmed8030143
APA StyleWang, R., Wu, M., Cai, H., An, R., Chen, Y., Wang, J., Zhou, N., & Du, J. (2023). Preparation and Preliminary Application of Epitope Peptide-Based Antibody against Toxoplasma gondii GRA3. Tropical Medicine and Infectious Disease, 8(3), 143. https://doi.org/10.3390/tropicalmed8030143