Abstract
The global spread of antimicrobial resistance genes (ARGs) is a major public health concern. Mobile genetic elements (MGEs) are the main drivers of this spread by horizontal gene transfer (HGT). Escherichia coli is widespread in various environments and serves as an indicator for monitoring antimicrobial resistance (AMR). Therefore, the objective of this work was to evaluate the whole genome of multidrug-resistant E. coli strains isolated from human clinical, animal, and environmental sources. Four E. coli strains previously isolated from human urine (n = 2), retail meat (n = 1), and water from the Rio Grande River (n = 1) collected in northern Tamaulipas, Mexico, were analyzed. E. coli strains were evaluated for antimicrobial susceptibility, followed by whole genome sequencing and bioinformatic analysis. Several ARGs were detected, including blaCTX-M-15, blaOXA-1, blaTEM-1B, blaCMY-2, qnrB, catB3, sul2, and sul3. Additionally, plasmid replicons (IncFIA, IncFIB, IncFII, IncY, IncR, and Col) and intact prophages were also found. Insertion sequences (ISs) were structurally linked with resistance and virulence genes. Finally, these findings indicate that E. coli strains have a large repertoire of resistance determinants, highlighting a high pathogenic potential and the need to monitor them.
1. Introduction
Antimicrobial resistance (AMR) represents one of the most urgent public health problems [1,2]. Excessive and inappropriate use of antimicrobials has led to the emergence of resistant bacteria and their subsequent dissemination among bacteria in different environments. AMR leads to antimicrobial treatment failure in both humans and animals [3]. In 2019, infections due to multidrug-resistant (MDR) bacteria were responsible for 1.27 million deaths mainly attributed to Escherichia coli (E. coli) [1].
Due to its ubiquity and genomic plasticity, E. coli [4] is one of the main bacteria involved in the spread of AMR in communities, foods, farms, animals, the environment, and in clinical settings [5] since it can constantly exchange external genetic material such as ARGs through mobile genetic elements (MGEs) [3,6]. Thus, this bacterium is used as an excellent index not only to monitor AMR but also prevalence, types, and movement of ARGs [5,7].
E. coli represents a threat to public health due to AMR and the versatility of pathotypes, which lead to intestinal and extraintestinal infections [7,8]. Extraintestinal pathogenic E. coli (ExPEC) strains are opportunists that remain asymptomatically in the intestine to subsequently colonize extraintestinal sites [9].
Previous studies [10,11,12] in the Tamaulipas state have evidenced by molecular techniques the circulation and high prevalence of MDR E. coli strains among different environments. However, the MGEs responsible for the spread of AGRs have not been investigated and consequently, it is unknown what elements are involved in the dispersal of MDR in the region, as well as the genetic diversity they present.
The implementation of WGS is an alternative to improve surveillance of MDR pathogens of health concern by overcoming the limitation of analyzing only a small part of the genome and providing faster management and monitoring of the emergence of new antibiotic-resistant strains and their evolution [13].
Therefore, in support of the One-Health program as well as the health concerns generated by antimicrobial resistance, the objective of this work was to evaluate whole genome multidrug-resistant E. coli strains from human clinical and environmental sources as well as to characterize the mobilome and resistome that contribute to the spread of antimicrobial resistance genes.
2. Materials and Methods
2.1. E. coli Strains
E. coli strains used in this study are part of a bacterial collection from the Environment-Microorganism Interaction Laboratory of the Centro de Biotecnología Genómica of the Instituto Politécnico Nacional and are also part of a larger project on antimicrobial resistance (Table 1). Two E. coli strains were collected from routine nosocomial pathogen testing specimens at a tertiary care hospital in Reynosa, Tamaulipas, Mexico. No patient data were requested. Therefore, no institutional review board (IRB) approval or informed consent was required.

Table 1.
E. coli strains included in this study.
2.2. Isolation of Strains
E. coli strains (n = 4) were collected between 2015 and 2018 and were previously identified by standard biochemical tests (lactose, indole, methyl red, Voges–Proskauer, Simmons Citrate, Christensen’s urea, and H2S production). The strains were grown in TSA (MCD LAB) and EMB (Eosin Methylene Blue) medium (MCD LAB) at 37 °C for 24 h. In addition, they were confirmed by ChromAgar orientation.
2.3. Antimicrobial Susceptibility Testing
Antimicrobial susceptibility analysis was performed using the disk diffusion method, following established guidelines [14]. Sixteen antibiotics were tested: tetracycline (TET, 30 µg), doxycycline (DOX, 30 µg), minocycline (MIN, 30 µg), ampicillin (AMP, 10 µg), amoxicillin/clavulanic acid (AMC, 20/10 µg), ciprofloxacin (CIP, 5 µg), trimethoprim-sulfamethoxazole (SXT, 25 µg), levofloxacin (LEV, 5 µg), streptomycin (STR, 30 µg), gentamicin (GE, 10 µg), cephalothin (CF, 30 µg), cefepime (FEP, 30 µg), cefotaxime (CTX, 30 µg), amikacin (AK, 30 µg), ceftriaxone (CRO, 30 µg), and chloramphenicol (CHL, 30 µg). E. coli ATCC 25,922 was used as a quality control strain.
2.4. Whole Genome Sequencing
For genomic DNA extraction, the strains were grown in LB broth (Condalab, Madrid, Spain) under agitation at 37 °C for 24 h. Subsequently, DNA was obtained using the Promega Wizard Genomics extraction kit (Promega Corp., Madison, WI, USA) and QIAmp® DNA Mini Kit (QIAGEN, Hilden, Germany). Whole genome sequencing was performed at the National Laboratory of Animal Digestive Nutrigenomics and Microbiomics (LANMDA-IPN) and DNA quantification was performed using the Qubit dsDNA HS Assay kit on the Qubit 3.0 fluorometer (Thermo Scientific, Waltham, MA, USA). Libraries were constructed using the Nextera Flex library kit. Libraries were sequenced using the MiniSeq™ sequencing system (150 bp paired-end reads).
2.5. Bioinformatic Analysis
Reads quality was assessed through FastQC v0.11.3 (https://www.bioinformatics.babraham.ac.uk/projects/fastqc/, accessed on 10 October 2021). Cleaning of raw reads was performed with Trim Galore v0.6.6 (https://www.bioinformatics.babraham.ac.uk/projects/trim_galore/, accessed on 12 October 2021). Assembly was performed using SPAdes v3.15.2 with settings —isolate and –k 21,31,41,51,61,71,81,91. The quality of the assemblies was assessed with QUAST v5.0.2 (https://github.com/ablab/quast, accessed on 12 October 2021). Contigs smaller than 500 bp were removed. Sequences were deposited in GenBank under bioproject PRJNA749581. Moreover, 31HGR E. coli strain was previously published as a draft genome [15] and it has been deposited in GenBank under the accession number JAKJKJ000000000.
Automatic annotation was performed in Rapid Annotation using Subsystem Technology from the Pathosystems Resource Integration Center (PATRIC) v3.6.9 [16] (now Bacterial and Viral Bioinformatics Resource Center https://www.bv-brc.org/, accessed on 15 October 2021). Manual annotation was executed in the Center for Genomic Epidemiology (https://www.genomicepidemiology.org/, accessed on 1 December 2021) bioinformatics tools such as ResFinder v4.1 (https://cge.food.dtu.dk/services/ResFinder/, accessed 1 December 2021), PlasmidFinder v2.1 (https://cge.food.dtu.dk/services/PlasmidFinder/, 1 December 2021), and SerotypeFinder v.2.0 (https://cge.food.dtu.dk/services/SerotypeFinder/, 3 December 2021). Other MGEs such as ISs and phage were analyzed by ISSaga v2.0 (https://issaga.biotoul.fr/, accessed on 2 December 2021) and PHASTER (https://phaster.ca/, accessed on 3 December 2021) [17], respectively. ISs that could not be identified by ISSaga were searched by BLASTn. Sequence types (STs) were determined in silico using the PubMLST database (https://pubmlst.org/, accessed on 3 January 2022). Default parameters were used for all software unless otherwise specified.
2.6. Phylogenomic Analysis and Genomic Comparison
Using the Similar Genome Finder of PATRIC web resources v3.6.12, a search for close reference genomes was performed. The sequences of these genomes, in FASTA format, were deposited in the CSIPhylogeny v1.4 pipeline (https://cge.cbs.dtu.dk/services/CSIPhylogeny/, accessed on 6 April 2022). Default parameters were used: minimum depth at SNP positions: 10×; minimum relative depth at SNP positions: 10%, minimum distance between SNPs: 10 bp; minimum SNP quality: 30; minimum read mapping quality: 25, and minimum Z-score: 1.96. E. coli K12 substr. MG1655 (GenBank: U00096.3) was used as the reference strain. Subsequently, to infer phylogeny from SNPs, the program MEGA X v10.0.5 [18] was employed using the maximum likelihood method and based on the Tamura–Nei model. Genomic comparisons were performed in GView Server v3.0 (https://server.gview.ca/, accessed on 29 April 2023) in which the four genomes that were sequenced in this study were combined with a few related genomes based on the data obtained from the Similar Genome Finder. The reference genome used was E. coli K12 substr. MG1655 (Genbank: U00096.3). The genomic context of regions containing resistance genes was analyzed using EasyFig v2.2.2 [19].
OrthoVenn2 web server [20] (https://orthovenn2.bioinfotoolkits.net/home, accessed on 28 April 2023) was used to predict orthologous gene clusters among the E. coli strains and default parameters were used.
3. Results
3.1. Antibiotic Susceptibility
Phenotypic resistance was identified mainly to TET, AMP, STR, SXT, CHL, and GE (Table 2). All four strains exhibited resistance to TET, while resistance to AMP was present in three strains as well as STR, SXT, CHL, and GE. Quinolone resistance was observed in E. coli 31HGR, 47C, and 87CLU. All E. coli strains were identified as MDR.
Table 2.
Phenotypic resistance profiles identified in E. coli strains.
3.2. Genomic Characteristics
The total genome size on average ranged between 4.72 Mb and 5.31 Mb, and the GC content was found to be between 50.80% and 50.95%. The number of contigs and N50 values are described in Table 3.
Table 3.
General genome features of E. coli strains.
3.3. Resistome
All strains had multiple acquired antibiotic resistance genes. Aminoglycoside resistance genes included aadA1, aadA2, aadA5, aac3-IIa, aph6-Id, aph(3″)-Ia, aph(3″)-Ib, and aac(6’)-Ib-cr. Resistance genes to β-lactams (blaOXA-1, blaTEM-1B, blaCMY-2, and blaCTX-M-15), sulfonamides (sul1, sul2, and sul3), trimethoprim (dfra12, drfa14, and drfa17), phenicols (catB3, cmlA1, and floR), tetracyclines (tet(A), and tet(B)), quaternary ammonium compounds (QCA) (qacE and qacL), macrolides (mdfA and mphA), quinolones (qnrB), and bacitracin (bacA) were also found (Table 4, Tables S1 and S2).
Table 4.
Resistome, mobilome, and virulome profiles of E. coli strains.
The tet(A), aadA2, aph(3″)-Ib, and aph(6)-Id genes were the most common among the genomes. The clinic strain 31HGR was found to have the blaCTX-M-15 and blaOXA-1 genes, among others. Additionally, gene qnrB was found in the surface water isolate (3AS) of the Rio Grande/Rio Bravo River. The sul2 and sul3, blaTEM-1B, and blaCMY-2 genes were the most relevant resistance genes identified in 87CLU and 47C.
Quinolone resistance was detected by mutations in three genes: gyrA, parC, and parE. Two mutations were found in gyrA (S83L, D87N), one mutation in parC (S80I), and two mutations in parE (S458T, S458A). All mutations in gyrA and parC were present in strains 31HGR, 47C, and 87CLU. The parE (S458T) mutation was identified only in strain 31HGR, while the parE (S458A) mutation was present in strains 87CLU and 47C. Additionally, strain 3AS E. coli did not exhibit any of these mutations (Table 5).
Table 5.
Mutational and plasmid-mediated quinolone resistance in E. coli strains.
3.4. Mobilome of E. coli
3.4.1. Plasmids
Multiple plasmid replicons were identified in all genomes (Table 4). The conjugative IncFIB replicon was the most prevalent in all strains. IncFIA and IncFII replicons were detected in 31HGR, 87CLU, and 47C. Meanwhile, IncY (phage-like plasmid) was present in 87CLU and 47C. The mobilizable replicons IncR and Col were identified in 3AS. All genomes evaluated had four plasmids.
3.4.2. Phages
A total of 18 intact prophages were identified; strain 47C harbored the most elements of this type (seven), followed by 87CLU which had five, while 31HGR had four and 3AS harbored three (Table 6). All the prophages encoded mainly hypothetical and structural proteins, except for the blaCMY-2 gene found exclusively in the IncY phage-like plasmid of 47C and 87CLU. In addition, IncY was found as an intact prophage in 47C E. coli strains.
Table 6.
Distribution of intact prophages and their detected E. coli strains.
3.4.3. Other EGMs
All strains possessed the intI1 gene, which codes for the integrase of the class 1 integron. Due to the characteristics of the reads, only the intI1 gene was recovered intact and adjacent to its resistance gene cassette in strains 31HGR and 3AS, while in strains 47C and 87CLU, it was found in separate contigs.
Moreover, multiple ISs associated with resistance or virulence genes were detected (Table S3); IS91 was detected close to aph(6)-Id, tet(A), sul2, and floR. ISEcp1 was detected adjacent to blaCTX-M-15 and blaCMY-2. IS26 was also identified by BLASTn adjacent to resistance genes (Table S4).
Common co-resistance patterns were detected in 47C and 87CLU, which have the arrangement sul2::IS91::floR::IS91 and IS91::floR::IS91::IS91::sul2, respectively, where the difference is in the position of genes involved in resistance to sulfonamides and phenicols. Furthermore, in these two strains, the aadA1::aadA2:cmlA:aadA1:qacL:IS256 arrangement in 87CLU is very similar to the aadA2:cmlA:aadA1:qacL:IS256:sul3 of 47C. These arrangements confer co-resistance to aminoglycosides, chloramphenicol, quaternary ammonium compounds, and sulfonamides.
3.5. Virulome
A total of 18 virulence-associated genes were identified (Table 4). The strains belonging to ST224 had the highest number of these genes (14), 31HGR had 12, and 3AS had the lowest number of genes (6). The csg, fimH, ompA, and hlyE genes were identified in all four strains, the iutA, iucA, and sitA genes were common in E. coli 31HGR, 47C, and 87CLU, which harbored the highest number of resistance genes. Genes from type II, III, IV, and VI secretion systems were also identified.
3.6. Phylogenomic Analysis
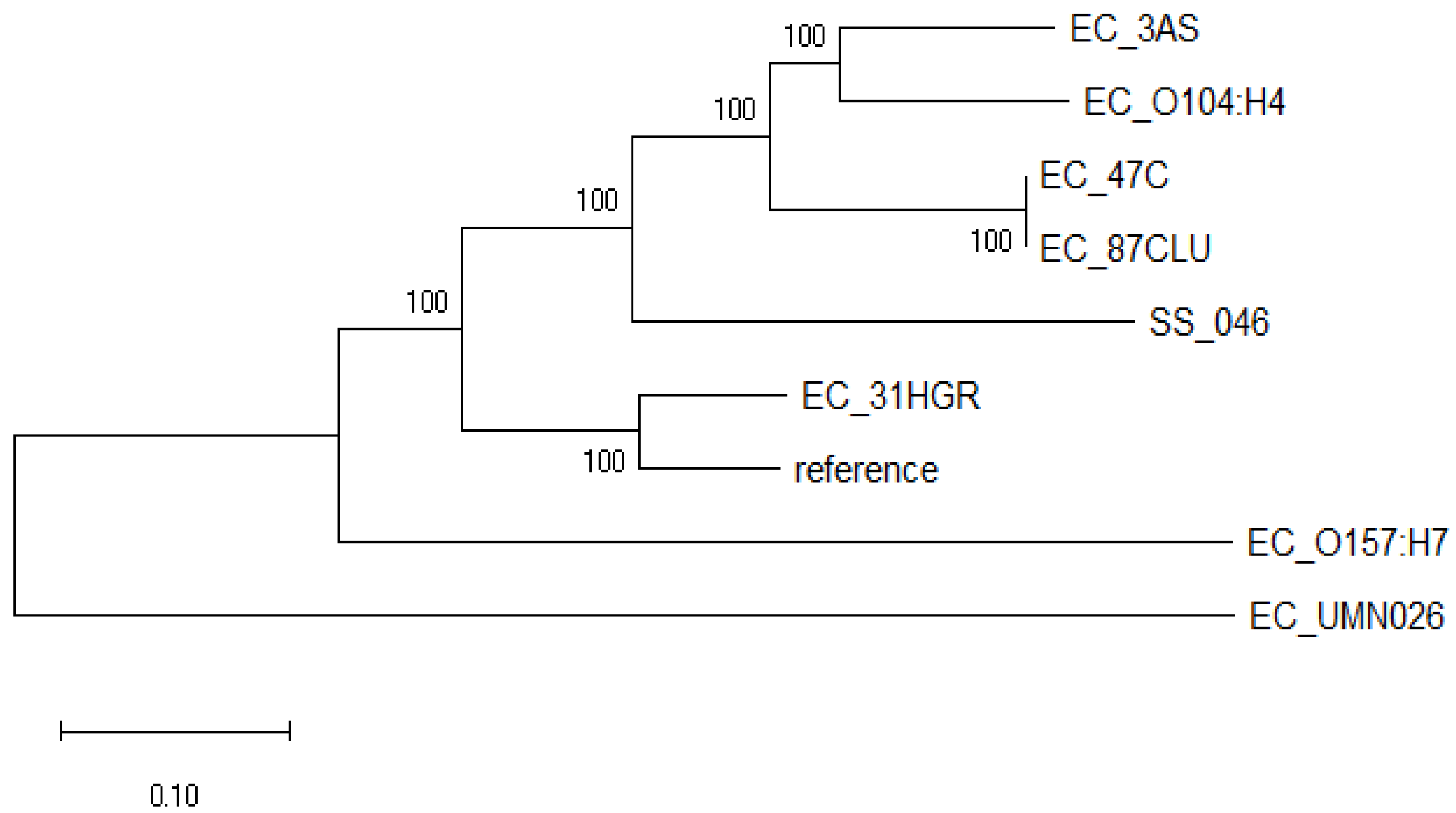
This analysis showed that the E. coli strains evaluated diverged into two well-defined clades (Figure 1). One clade consisted only of EC_UMN026 (CU928163.2), an MDR uropathogenic E. coli. On the other hand, the second clade was subdivided into two clearly differentiated groups: one comprised only by shigatoxigenic E. coli EC_O157:H7 (BA000007.2), and the other consisted of the strains sequenced in this study and those previously reported EC_O104:H4 (CP003289.1), EC_ K12 (U00096.3) and SS_046 (CP000038.1); 31HGR showed a divergence from the rest of the strains as it grouped only with the reference strain. Meanwhile, 3AS exhibited a close relationship with the intestinal pathogen EC_O104:H4 (CP003289.1).
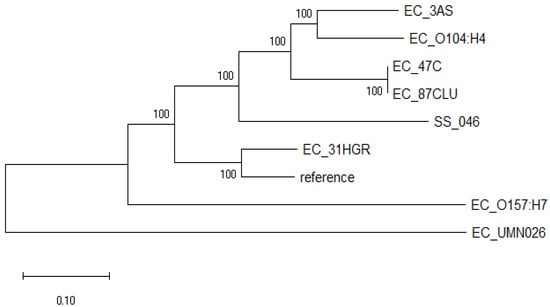
Figure 1.
Phylogenomic tree of E. coli strains sequenced in this work (31HGR, 87CLU, 47C, and 3AS) and close reference genomes based on single nucleotide polymorphism (SNPs) differences and constructed with the maximum likelihood method. Evolutionary distances were calculated using the Tamura–Nei model at 100 replicates.
3.7. Comparative Genomics
The genomic environment of the class 1 integron resistance gene cassettes of strains 31HGR and 3AS, which had the most resistance gene cassettes in the same contig, was analyzed (Figure 2). The comparison was performed against the sequences of p32-4_A (CP048311.1) and pCTXM15_000200 (CP022227.3).
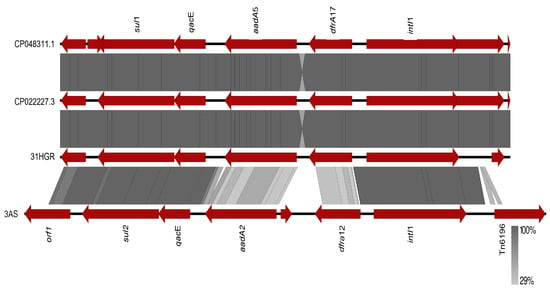
Figure 2.
Comparison of the genomic environment of the intI1 gene of reported plasmids CP048311.1 and CP022227.3 against the sequences of E. coli strains 31HGR and 3AS. The gray shaded region between the sequences indicates the similarity according to BLASTx (29–100%).
A 100% similarity was observed between E. coli 31HGR and p32-4_A (CP048311.1) and pCTXM15_000200 (CP022227.3), whereas 3AS presented a lower similarity since, although they have the same resistance gene families, they have different variants.
Orthologous gene analysis identified that strain 31HGR contained 4686 proteins and 4257 clusters, 5046 proteins and 5000 clusters in 47C, 5047 proteins and 4992 in 87CLU, and 4477 proteins and 4094 clusters; the reference strain EC_ K12 (U00096.3) consisted of 4305 proteins and 4055 clusters. In addition, among the orthologous gene clusters, the core genome consisted of 2595 clusters (Figure 3). The species formed 5313 clusters, 1741 orthologous clusters, and 3572 single-copy gene clusters. Strains 87CLU and 47C shared the most gene clusters and these bacteria belong to the same ST. Strains 31HGR and 3AS had the most single-copy gene clusters (357).
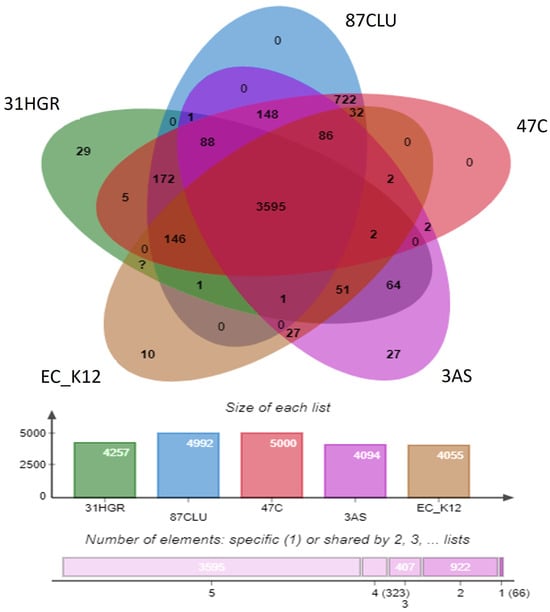
Figure 3.
Venn diagram of orthologous gene clusters present in E. coli strains 31HGR, 87CLU, 47C, 3AS, and the reference strain.
The GView analysis showed the similarity between the different genomes sequenced in the present work. It is also possible to visualize the variations in GC content in different regions; those areas that are not shared with those of the reference genome show a higher content with respect to the genome average, which is evidence of genetic material acquired by HGT (Figure 4).
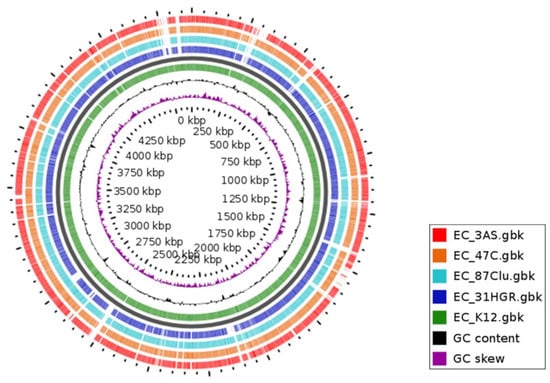
Figure 4.
Comparative analysis of E. coli genomes with the reference strain. The BLAST atlas was obtained using GView Server. Blanks indicate areas where there is no similarity. Colored rings match the color of the strains. E. coli K12 (U00096.3) was used as a reference genome.
4. Discussion
The findings of this work show a diversity of the resistome, mobilome, and virulome of E. coli strains isolated from different sources in Tamaulipas. Most of the ARGs were detected associated with the class 1 integron or ISs. To our knowledge, this is the first study that provides a genomic evaluation of antimicrobial resistance and MGEs in E. coli strains in northeastern Tamaulipas.
In this work, resistance to antibiotics that are currently of clinical relevance such as quinolone resistance mediated by chromosomal mutations in the gyrA (S83L, D87N), parC (S80I), and parE (S458T, S458A) genes were detected in the genomes of E. coli from human isolates and retail meat. These findings correlate with the phenotypic resistance observed to ciprofloxacin, levofloxacin, and nalidixic acid, which are relevant antibiotics for the treatment of a wide range of infections, mainly in urinary tract infections (UTIs) [21]. In addition, plasmid-mediated quinolone resistance (PMQR) was also identified by the presence of qnrB and aac(6’)-Ib-cr genes. These genes by themselves confer low levels of resistance [22]; in this work, the 3AS strain harboring qnrB did not exhibit resistance to quinolones, which is consistent with that previously reported [23], and also did not present chromosomal mutations that confer resistance to this family.
Aminoglycoside resistance genes were common; aph(6)-Id and aph(3″)-Ib were present in three strains (31HGR, 87CLU and 47C) both encoding phosphotransferases. However, only 87CLU and 47C exhibit phenotypic resistance to two members of this antibiotic family: gentamicin and streptomycin, which still have clinical relevance against Gram-positive and Gram-negative infections, as they act synergistically with β-lactams and glycopeptides [24].
Regarding resistance to β-lactams, blaOXA-1, blaCTX-M-15, blaTEM-1B, and blaCMY-2 genes were presented in this study of which CTX-M-15 is the most prevalent and globally distributed ESBL in E. coli [25] and is associated with UPEC ST131 [26].
Furthermore, blaCMY-2 encodes for a plasmid-mediated AmpC-type β-lactamase [23] that confers resistance to all β-lactams except the fourth generation of extended-spectrum cephalosporins and carbapenems [27]. These β-lactamases are more concerning than ESBLs because they are typically resistant to inhibitors such as clavulanic acid, tazobactam, and sulbactam, thereby becoming clinically relevant [5,28]. Moreover, the combination of ESBL and AmpC genes with the loss of outer membrane porin can cause resistance to carbapenems [27].
Due to its spread by plasmids from different groups, the blaCMY-2 gene is commonly found in E. coli worldwide from human, animal, and environmental sources [29]. In North America, this gene is commonly found in E. coli from cattle [30] Additionally, it has been previously identified in pediatric patients in Mexico [31]. However, in this study, blaCMY-2 was identified not only in retail meat (47C) but also in E. coli that was isolated from humans (87CLU), emphasizing the transmission of blaCMY-2 from animals to humans.
Interestingly, PHASTER revealed that in the two strains harboring blaCMY-2, it was embedded in a phage-like plasmid, IncY; in both cases, blaCMY-2 was adjacent to ISEcp1, which explains its mobility by THG between plasmids and between bacteria from different environments.
In this work, the presence of the mphA gene in 31HGR also is noteworthy; although phenotypic resistance was not observed, its presence is of concern since macrolides are usually used mainly as therapeutics against Gram-positives [32]. Of this family of antibiotics, azithromycin has shown greater activity and offers an alternative treatment against Shigella, Salmonella, and enterotoxigenic E. coli (ETEC) [33,34]. Therefore, plasmid-mediated macrolide resistance is alarming; mphA is the most relevant resistance mechanism since it has been observed to increase MIC in E. coli, and another aspect of concern is that E. coli is serving as a reservoir for this gene and could transfer it to clinically important bacteria [5,33].
Additionally, all E. coli strains that exhibited phenotypic resistance to tetracyclines harbored the tet(A) gene, tet(B) gene, or both. Notably, the tet(A) gene is readily dispersed between different environments [35] which is largely because such a gene is frequently harbored by plasmids of a wide host range and a different incompatibility group [36].
On the other hand, in this study, the genes for resistance to sulfonamides, trimethoprim-sulfamethoxazole and aminoglycosides, were identified forming part of the class 1 integron, which is congruent since these cassettes are frequently associated with such MGEs in commensal and pathogenic E. coli from animal, environmental, and human sources [37,38].
Currently, an increase in phenotypic resistance to multiple antibiotics has been reported in different types of sequences around the world and obtained from different environments; this situation can lead to therapeutic failures. In this work, it was detected that E. coli 31HGR belongs to ST44, which is part of CC10, as is ST131, and is well known for being opportunistic, MDR, and associated with intestinal and extraintestinal infections in both humans and animals [9]. Whereas 47C and 87CLU belong to ST224, which is considered pandemic and related to MDR, mainly to β-lactams and carbapenems [39], in this work, the two ST224 harbored blaTEM-1B and blaCMY-2 genes. Similarly, this study identified ST155 in E. coli strain 3AS, which originates from animals and is linked to the transmission of multidrug resistance (MDR) via plasmids [23,40].
It is noteworthy that the E. coli strains 47C and 87CLU, which belong to the same ST and have a similar genomic content, showed different phenotypic resistance profiles; this situation may be influenced by the fact that the bacteria were isolated from different sources (human and retail meat), so the exposure to selective pressures in their environment are different and could influence the expression of resistance genes.
Regarding phenotypic quinolone resistance, here, 87CLU and 47C had a different profile. Zhang et al. showed that isolates with the combination S83L + D87N mutation had resistance to statistically different drugs between ciprofloxacin and levofloxacin [41]. Resistance to quinolones is a major concern because they are some of the most potent classes of antibiotics that are currently available. Consequently, the World Health Organization (WHO) has prioritized combating quinolone resistance [42].
In addition, different plasmid replicons were also identified in the present study with IncF being the most common; this element is widely dispersed in Enterobacterales and is known to be epidemic and to harbor ARGs such as blaCTX-M-15 in addition to virulence genes [43]. Interestingly, in strain 3AS, the Col-like mobilizable replicon, a small 6 to 10 Kb, colicin-producing plasmid also associated with the spread of qnr family genes, was detected, which is consistent with the findings reported here, as qnrB was identified in strain 3AS [44].
Multiple transposable elements were identified; in some cases, they were found adjacent or close to ARGs, which raises concern for their ability to mobilize genetic material such as ISEcp1 of the IS1380 family related to the mobilization of a wide variety of ARGs, mainly β-lactam resistance [45] as identified in this work, adjacent to blaCMY-2 and blaCTX-M-15. In the present study, IS91 was among the most associated with antibiotic resistance genes, [5] and [46] reported similar results in E. coli; these types of sequences can mobilize adjacent genes through a single-end transposition process [46]. Detection of multiple insertion sequences and compound transposons flanking antibiotic resistance genes or located near them emphasizes the significance of these elements in mobilizing the genes within E. coli strains isolated in our region.
The spread and increase of clinically relevant UTIs in different surroundings is an alarming health and environmental concern [47]. In Mexico, treatment of urinary tract infections generally involves trimethoprim with sulfamethoxazole, quinolones, second- and third-generation cephalosporins, nitrofurantoin, and fosfomycin [48,49]. In our work, resistance to some of the first-choice treatment antibiotics was found, which is of concern because such strains could successfully establish in different environments and subsequently lead to difficult-to-treat infections. In Mexico, similar findings were reported in UPEC [22,38,50].
Moreover, numerous virulence genes were identified, especially those related to iron uptake. Notably, the strain 87CLU from phylogroup B1 harbored the highest number of virulence genes, so these findings are important because these bacteria and their clones could reach other environments closely related to humans.
Phylogenetic analysis showed a close relationship between E. coli strains regardless of the isolation site. The 31HGR, 47C, 87CLU, and 3AS genomes were related to previously reported strains that are of intestinal origins (either pathogenic or commensal) such as E. coli O157:H7 and E. coli 0104:H4. It is noteworthy that 31HGR, 47C, and 87CLU harbored genes that are common to ExPECs, especially 31HGR, which was the strain that exhibited divergence with respect to the other genomes sequenced in this work; this evidences the evolution of intestinal bacteria given the need to adapt to different environments [51] by acquiring genetic material from ExPECs.
Genomic analysis of E. coli MDR strains from different environments revealed a diversity of ARGs and virulence genes, as well as multiple MGEs involved in their propagation that confer to the bacterium advantages not only to colonize, but also to persist in different environments [38]. An important limitation of this study is the small number of sequenced isolates. Consequently, it does not offer a comprehensive overview of the wide range of clonal groups circulating within our area, including the ARGs and MGEs. In addition, the reconstruction of certain important MGEs, such as plasmids and their genomic content, is difficult due to the short reads. Therefore, larger-scale epidemiological studies are required to monitor multidrug-resistant bacteria and their MGEs to determine the possible routes and fates of ARGs.
5. Conclusions
Finally, the findings of this work evidence the circulation in the environment and in the community of bacteria with a diversity of antimicrobial resistance determinants and virulence genes, as well as multiple MGEs involved in their propagation, which represent a health concern because they could lead to therapeutic failures. Therefore, epidemiological monitoring is of great importance to know and control the spread of these MDR bacteria.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/2414-6366/8/10/458/s1, Table S1: ARGs identified by ResFinder; Table S2: ARGs identified by BV-BRC (PATRIC) in E. coli strains; Table S3: Insertion sequences; Table S4: Synteny ARG and IS.
Author Contributions
Conceptualization, J.L.O.-B., A.G. and V.B.-G.; methodology, J.L.O.-B., A.G., A.V.M.-V., M.A.C.-H., E.d.J.d.L.-S., E.A.-C. and I.P.R.-S.; resources, A.V.M.-V. and M.A.C.-H.; writing—original draft preparation, J.L.O.-B. and G.C.-E.; writing—review and editing, G.R. and V.B.-G.; supervision, G.C.-E. and V.B.-G.; funding acquisition, G.R. and V.B.-G. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All the analyzed data are included in this published article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Murray, C.J.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Robles Aguilar, G.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global Burden of Bacterial Antimicrobial Resistance in 2019: A Systematic Analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef]
- O’Neill, J. Tackling Drug-Resistant Infections Globally: Final Report and Recommendations. Rev. Antimicrob. Resist. 2016. Available online: https://amr-review.org/sites/default/files/160518_Final%20paper_with%20cover.pdf (accessed on 13 September 2023).
- Aworh, M.K.; Kwaga, J.K.P.; Hendriksen, R.S.; Okolocha, E.C.; Thakur, S. Genetic Relatedness of Multidrug Resistant Escherichia coli Isolated from Humans, Chickens and Poultry Environments. Antimicrob. Resist. Infect. Control 2021, 10, 58. [Google Scholar] [CrossRef] [PubMed]
- van Elsas, J.D.; Semenov, A.V.; Costa, R.; Trevors, J.T. Elsas Survival of Escherichia coli in the Environment: Fundamental and Public Health Aspects. ISME J. 2011, 5, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Mbelle, N.M.; Feldman, C.; Sekyere, J.O.; Maningi, N.E.; Modipane, L.; Essack, S.Y. The Resistome, Mobilome, Virulome and Phylogenomics of Multidrug-Resistant Escherichia coli Clinical Isolates from Pretoria, South Africa. Sci. Rep. 2019, 9, 16457. [Google Scholar] [CrossRef] [PubMed]
- Massella, E.; Giacometti, F.; Bonilauri, P.; Reid, C.J.; Djordjevic, S.P.; Merialdi, G.; Bacci, C.; Fiorentini, L.; Massi, P.; Bardasi, L.; et al. Antimicrobial Resistance Profile and ExPEC Virulence Potential in Commensal Escherichia coli of Multiple Sources. Antibiotics 2021, 10, 351. [Google Scholar] [CrossRef]
- Shin, S.W.; Shin, K.; Jung, M.; Belaynehe, M.; Yoo, S. Prevalence of Antimicrobial Resistance and Transfer of Tetracycline Resistance Genes in Escherichia coli Isolates from Beef Cattle. Appl. Environ. Microbiol. 2015, 81, 5560–5566. [Google Scholar] [CrossRef]
- Reid, C.J.; Blau, K.; Jechalke, S.; Smalla, K.; Djordjevic, S.P.; Campo, R. Del Whole Genome Sequencing of Escherichia coli From Store-Bought Produce. Front. Microbiol. 2020, 10, 3050. [Google Scholar] [CrossRef]
- Reid, C.J.; Wyrsch, E.R.; Chowdhury, P.R.; Zingali, T.; Liu, M.; Darling, A.E.; Chapman, T.A.; Djordjevic, S.P. Porcine Commensal Escherichia coli: A Reservoir for Class 1 Integrons Associated with IS 26. Microb. Genom. 2017, 3, e000143. [Google Scholar] [CrossRef]
- Martínez-Vázquez, A.V.; Rivera-Sánchez, G.; Lira-Méndez, K.; Reyes-López, M.A.; Bocanegra-García, V. Prevalence, Antimicrobial Resistance and Virulence Genes of Escherichia coli Isolated from Retail Meat in Tamaulipas, Mexico. J. Glob. Antimicrob. Resist. 2018, 14, 266–272. [Google Scholar] [CrossRef]
- Martínez-Vázquez, A.V.; Vázquez-Villanueva, J.; Leyva-Zapata, L.M.; Barrios-García, H.; Rivera, G.; Bocanegra-García, V. Multidrug Resistance of Escherichia coli Strains Isolated From Bovine Feces and Carcasses in Northeast Mexico. Front. Vet. Sci. 2021, 8, 643802. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Vázquez, A.V.; Mandujano, A.; Cruz-Gonzalez, E.; Guerrero, A.; Vazquez, J.; Cruz-Pulido, W.L.; Rivera, G.; Bocanegra-García, V. Evaluation of Retail Meat as a Source of ESBL Escherichia coli. Antibiotics 2022, 11, 1795. [Google Scholar] [CrossRef] [PubMed]
- Moghnia, O.H.; Al-Sweih, N.A. Whole Genome Sequence Analysis of Multidrug Resistant Escherichia coli and Klebsiella pneumoniae Strains in Kuwait. Microorganisms 2022, 10, 507. [Google Scholar] [CrossRef]
- CLSI. M100 Performance Standards for Antimicrobial; CLSI: Malvern, PA, USA, 2018; ISBN 156238838X. [Google Scholar]
- Ortega-Balleza, J.L.; Guerrero, A.; Castro-Escarpulli, G.; Cruz-González, E.; Rivera, G.; Bocanegra-García, V. Draft Genome Sequence of a Uropathogenic Escherichia coli. Microbiol. Resour. Annouc. 2022, 11, e0093121. [Google Scholar]
- Davis, J.J.; Wattam, A.R.; Aziz, R.K.; Brettin, T.; Butler, R.; Butler, R.M.; Chlenski, P.; Conrad, N.; Dickerman, A.; Dietrich, E.M.; et al. The PATRIC Bioinformatics Resource Center: Expanding Data and Analysis Capabilities. Nucleic Acids Res. 2020, 48, 606–612. [Google Scholar] [CrossRef] [PubMed]
- Arndt, D.; Grant, J.R.; Marcu, A.; Sajed, T.; Pon, A.; Liang, Y.; Wishart, D.S. PHASTER: A Better, Faster Version of the PHAST Phage Search Tool. Nucleic Acids Res. 2016, 44, 16–21. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Sullivan, M.J.; Petty, N.K.; Beatson, S.A. Easyfig: A Genome Comparison Visualizer. Bioinformatics 2011, 27, 1009–1010. [Google Scholar] [CrossRef]
- Xu, L.; Dong, Z.; Fang, L.; Luo, Y.; Wei, Z.; Guo, H.; Zhang, G.; Gu, Y.Q.; Coleman-Derr, D.; Xia, Q.; et al. OrthoVenn2: A Web Server for Whole-Genome Comparison and Annotation of Orthologous Clusters across Multiple Species. Nucleic Acids Res. 2019, 47, W52–W58. [Google Scholar] [CrossRef]
- Kot, B. Antibiotic Resistance Among Uropathogenic Escherichia coli. Pol. J. Microbiol. 2019, 68, 403–415. [Google Scholar] [CrossRef]
- Ramírez-Castillo; Castillo, F.Y.R.; Flores, A.C.M.; González, F.J.A.; Díaz, F.M.; Harel, J.; Barrera, A.L.G. An Evaluation of Multidrug—Resistant Escherichia coli Isolates in Urinary Tract Infections from Aguascalientes, Mexico: Cross—Sectional Study. Ann. Clin. Microbiol. Antimicrob. 2018, 17, 34. [Google Scholar] [CrossRef] [PubMed]
- Gomi, R.; Matsuda, T.; Matsumura, Y.; Yamamoto, M.; Tanaka, M.; Ichiyama, S.; Yoneda, M. Crossm Resistant and Extraintestinal Pathogenic Escherichia coli in River Water. Appl. Environ. Microbiol. 2017, 83, e02703-16. [Google Scholar] [CrossRef] [PubMed]
- Soleimani, N.; Aganj, M.; Ali, L.; Shokoohizadeh, L.; Sakinc, T. Frequency Distribution of Genes Encoding Aminoglycoside Modifying Enzymes in Uropathogenic E. coli Isolated from Iranian Hospital. BMC Res. Nores 2014, 7, 842. [Google Scholar] [CrossRef]
- Rocha-Gracia, R.C.; Lozano-Zarain, P.; Cázarez, Z.G.; Andrea, C.; Brambila, E.; Torres, C.; Cortés-Cortés, G. IncFIB Plasmids Carrying the Resistance Gene Bla CTX-M-15 in ESBL-Producing Escherichia coli Clones from Pediatric Patients. J. Infect. Dev. Ctries. 2022, 16, 500–506. [Google Scholar] [CrossRef] [PubMed]
- Mazumder, R.; Abdullah, A.; Ahmed, D.; Hussain, A. High Prevalence of blaCTX-M-15 Gene among Escherichia coli Isolates Causing Extraintestinal Infections in Bangladesh. Antibiotics 2020, 9, 796. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Kader, F.; Hamza, E.; Abdel-Moein, K.A.; Sabry, M.A. Retail Chicken Giblets Contaminated with Extended-Spectrum Cephalosporin-and Carbapenem-Resistant Salmonella enterica Carrying blaCMY-2. Vet. World 2022, 15, 1297–1304. [Google Scholar] [CrossRef]
- Jouini, A.; Klibi, A.; Kmiha, S.; Hamrouni, S.; Ghram, A.; Maaroufi, A. Lineages, Virulence Gene Associated and Integrons among Extended Spectrum β-Lactamase (ESBL) and CMY-2 Producing Enterobacteriaceae from Bovine Mastitis, in Tunisia. Pathogens 2022, 11, 948. [Google Scholar] [CrossRef]
- Fang, L.X.; Li, X.P.; Li, L.; Chen, M.Y.; Wu, C.Y.; Li, L.L.; Liao, X.-P.; Liu, Y.-H.; Sun, J. ISEcp1 -Mediated Transposition of Chromosome- Borne BlaCMY-2 into an Endogenous ColE1-like Plasmid in Escherichia coli. Infect. Drug Resist. 2018, 11, 995–1005. [Google Scholar] [CrossRef]
- Awosile, B.; Reyes-velez, J.; Cuesta-Astroz, Y.; Rodríguez-Lecompte, J.C.; Saab, M.E.; Heider, L.C.; Keefe, G.; Sánchez, J.; Mcclure, J.T. Whole-Genome Sequence Analysis of 4 Fecal blaCMY-2 -Producing Escherichia coli Isolates from Holstein Dairy Calves. J. Dairy Sci. 2020, 103, 877–883. [Google Scholar] [CrossRef]
- Merida-Vieyra, J.; De Colsa-Ranero, A.; Calderón-Castañeda, Y.; Aquino-Andrade, A. Detection of CMY-Type Beta-Lactamases in Escherichia coli Isolates from Paediatric Patients in a Tertiary Care Hospital in Mexico. Antimicrob. Resist. Infect. Control 2020, 9, 168. [Google Scholar] [CrossRef]
- Gomes, C.; Ruiz-Roldán, L.; Mateu, J.; Ochoa, T.J.; Ruiz, J. Azithromycin Resistance Levels and Mechanisms in Escherichia coli. Sci. Rep. 2019, 9, 6089. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, M.C.P.; Woerther, P.L.; Bouvet, M.; Andremont, A.; Leclercq, R.; Canu, A. Escherichia coli as Reservoir for Macrolide. Emerg. Infect. Dis. 2009, 15, 14–16. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Y.; Wu, F.; Chai, Y.; Xu, X.; Yang, L.; Tian, S.; Zhang, H.; Li, Y.; Yang, C.; Liu, H.; et al. A New Plasmid Carrying MphA Causes Prevalence of Azithromycin Resistance in Enterotoxigenic Escherichia coli Serogroup. BMC Microbiol. 2020, 20, 247. [Google Scholar] [CrossRef] [PubMed]
- Reza, H.; Wang, P.; Robertson, I.; Abraham, S.; Sahibzada, S.; Habib, I. Antimicrobial Resistance and Genomic Characterisation of Escherichia coli Isolated from Caged and Non-Caged Retail Table Eggs in Western Australia. Int. J. Food Microbiol. 2021, 340, 109054. [Google Scholar] [CrossRef]
- Adesoji, A.; Ogunjobi, A.; Olatoye, I.; Douglas, D. Prevalence of Tetracycline Resistance Genes among Multi-Drug Resistant Bacteria from Selected Water Distribution Systems in Southwestern Nigeria. Ann. Clin. Microbiol. Antimicrob. 2015, 14, 35. [Google Scholar] [CrossRef]
- Awad, A.; Arafat, N.; Elhadidy, M. Genetic Elements Associated with Antimicrobial Resistance among Avian Pathogenic Escherichia coli. Ann. Clin. Microbiol. Antimicrob. 2016, 15, 59. [Google Scholar] [CrossRef]
- Ochoa, S.A.; Cruz-Cordova, A.; Lune-Pineda, V.M.; Reyes-Grajeda, J.P.; Cázares-Domínguez, V.; Escalona, G.; Sépulbeda-González, M.E.; López-Montiel, F.; Arellano-Galinedo, J.; López-Martínez, B.; et al. Multidrug- and Extensively Drug-Resistant Uropathogenic Escherichia coli Clinical Strains: Phylogenetic Groups Widely Associated with Integrons Maintain High Genetic Diversity. Front. Microbiol. 2016, 7, 2042. [Google Scholar] [CrossRef]
- Delgado-Blas, J.F.; Ovejero, C.M.; David, S.; Montero, N.; Calero-Caceres, W.; Garcillan-Barcia, M.P.; de la Cruz, F.; Muniesa, M.; Aanensen, D.M.; Gonzalez-Zorn, B. Population Genomics and Antimicrobial Resistance Dynamics of Escherichia coli in Wastewater and River Environments. Commun. Biol. 2021, 4, 457. [Google Scholar] [CrossRef]
- Neumann, B.; Rackwitz, W.; Hunfeld, K.P.; Fuchs, S.; Werner, G.; Pfeifer, Y. Genome Sequences of Two Clinical Escherichia coli Isolates Harboring the Novel. Gut Pathog. 2020, 12, 40. [Google Scholar] [CrossRef]
- Fu, Y.; Zhang, W.; Wang, H.; Zhao, S.; Chen, Y.; Meng, F.; Zhang, Y.; Xu, H.; Chen, X.; Zhang, F. Specific Patterns of GyrA Mutations Determine the Resistance Difference to Ciprofloxacin and Levofloxacin in Klebsiella pneumoniae and Escherichia coli. BMC Infect. Dis. 2013, 13, 2–7. [Google Scholar] [CrossRef]
- Astorga, F.; Navarrete-Talloni, M.J.; Miró, M.P.; Bravo, V.; Toro, M.; Blondel, C.J.; Hervé-Claude, L.P. Antimicrobial Resistance in E. coli Isolated from Dairy Calves and Bedding Material. Heliyon 2019, 5, e02773. [Google Scholar] [CrossRef] [PubMed]
- Pitout, J.D.D.; Chen, L. The Significance of Epidemic Plasmids in the Success of Multidrug-Resistant Drug Pandemic Extraintestinal Pathogenic Escherichia coli. Infect. Dis. Ther. 2023, 12, 1029–1041. [Google Scholar] [CrossRef] [PubMed]
- Rozwandowicz, M.; Brouwer, M.S.M.; Fischer, J.; Wagenaar, J.A.; Gonzalez-Zorn, B.; Guerra, B.; Mevius, D.J.; Hordijk, J. Plasmids Carrying Antimicrobial Resistance Genes in Enterobacteriaceae. J. Antimicrob. Chemother. 2018, 73, 1121–1137. [Google Scholar] [CrossRef] [PubMed]
- Partridge, S.R.; Kwong, S.M.; Firth, N.; Jensen, S.O. Crossm Mobile Genetic Elements Associated with Antimicrobial Resistance. Clin. Microbiol. Rev. 2018, 31, e00088-17. [Google Scholar] [CrossRef] [PubMed]
- Mbanga, J.; Amoako, D.G.; Abia, A.L.K.; Allam, M. Genomic Insights of Multidrug-Resistant Escherichia coli From Wastewater Sources and Their Association With Clinical Pathogens in South Africa. Front. Vet. Sci. 2021, 8, 636715. [Google Scholar] [CrossRef] [PubMed]
- Furlan, R.; Lopes, R.; Santana, M.; David, L.; Rosa, S.; Angelino, E.; Stehling, G. Infection, Genetics and Evolution blaCTX-M-2 and blaCMY-2 Recovered from an Urban Stream. Infect. Genet. Evol. 2021, 96, 105156. [Google Scholar] [CrossRef]
- Garza-Montúfar, M.E.; Treviño-Valdez, P.D.; De La Garza-Salina, L. Comorbidities and Antimicrobial Resistance in Urological Outpatients with Positive Urine Culture. Rev. Med. Inst. Mex. Seguro Soc. 2018, 56, 347–353. [Google Scholar]
- Ballesteros-Monrreal, M.G.; Arenas-Hernández, M.M.P.; Enciso-Martínez, Y.; Martínez-de la Peña, C.F.; Rocha-Gracia, R.C.; Lozano-Zaraín, P.; Navarro-Ocaña, A.; Martínez-Laguna, Y. Virulence and Resistance Determinants of Uropathogenic Escherichia coli Strains Isolated from Pregnant and Non-Pregnant Women from Two States in Mexico. Infect. Drug Resist. 2020, 13, 295–310. [Google Scholar] [CrossRef]
- Paniagua-Contreras, G.L.; Monroy-Pérez, E.; Díaz-Velásquez, C.E.; Uribe-García, A.; Labastida, A.; Peñaloza-Figueroa, F.; Domínguez-Trejo, P.; García, L.R.; Vaca-Paniagua, F.; Vaca, S. Whole-Genome Sequence Analysis of Multidrug-Resistant Uropathogenic Strains of Escherichia coli from Mexico. Infect. Drug Resist. 2019, 12, 2363–2377. [Google Scholar] [CrossRef]
- Zhang, X.; Xiao, S.; Jiang, X.; Li, Y.; Fan, Z.; Yu, Y.; Wang, P.; Li, D. Genomic Characterization of Escherichia coli LCT—EC001, an Extremely Multidrug—Resistant Strain with an Amazing Number of Resistance Genes. Gut Pathog. 2019, 11, 25. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).