
Overlapping of Pulmonary Fibrosis of Postacute COVID-19 Syndrome and Tuberculosis in the Helminth Coinfection Setting in Sub-Saharan Africa

 ,
, 
{kind=link}
Abstract
:1. Introduction
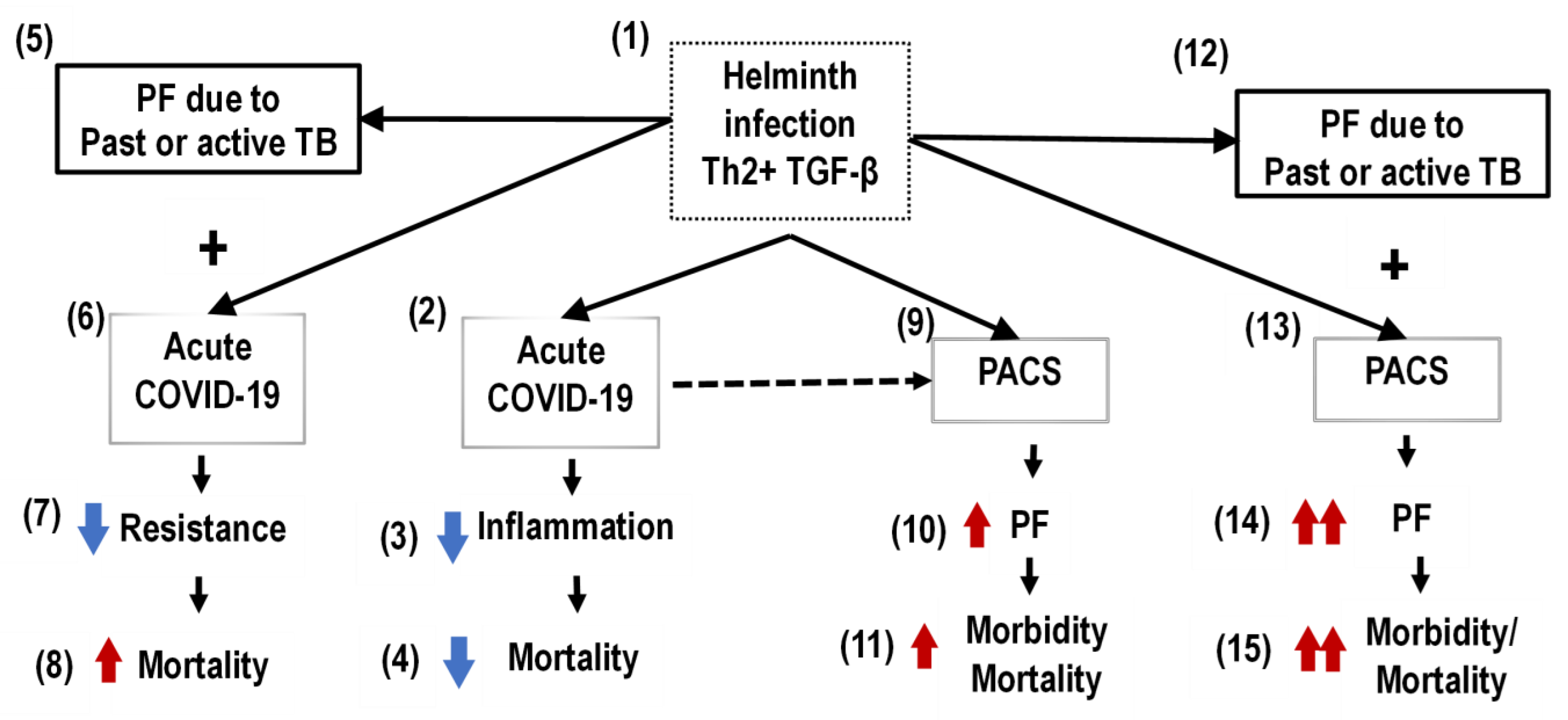
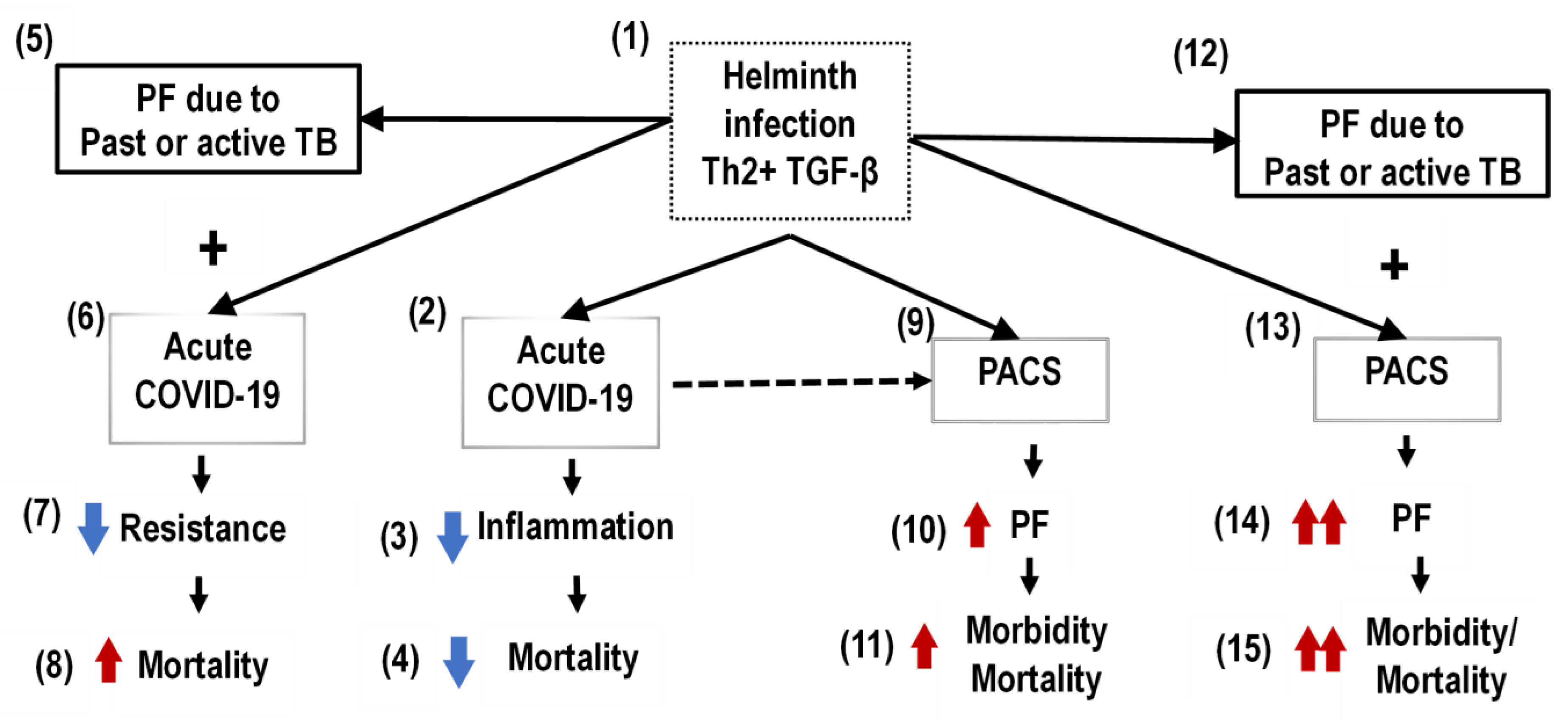
2. Pulmonary Fibrosis (PF) of Postacute COVID-19 Syndrome (PACS)
3. Overlapping of Pulmonary Fibrosis (PF) of Postacute COVID-19 Syndrome (PACS) and Tuberculosis (TB)
4. Pulmonary Fibrosis (PF) in the Helminth Coinfection Setting in Sub-Saharan Africa (SSA)
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vianello, A.; Guarnieri, G.; Braccioni, F.; Lococo, S.; Molena, B.; Cecchetto, A.; Giraudo, C.; De Marchi, L.B.; Caminati, M.; Senna, G. The pathogenesis, epidemiology and biomarkers of susceptibility of pulmonary fibrosis in COVID-19 survivors. Clin. Chem. Lab. Med. 2022, 60, 307–316. [Google Scholar] [CrossRef] [PubMed]
- Spagnolo, P.; Balestro, E.; Aliberti, S.; Cocconcelli, E.; Biondini, D.; Della Casa, G.; Sverzellati, N.; Maher, T.M. Pulmonary fibrosis secondary to COVID-19: A call to arms? Lancet 2020, 8, 750–752. [Google Scholar] [CrossRef]
- Aggarwal, A.N.; Agarwal, R.; Dhooria, S.; Prasad, K.T.; Sehgal, I.S.; Muthu, V. Active pulmonary tuberculosis and coronavirus disease 2019: A systematic review and meta-analysis. PLoS ONE 2021, 16, e0259006. [Google Scholar] [CrossRef] [PubMed]
- Tamuzi, J.L.; Lulendo, G.; Mbuesse, P. The incidence and mortality of COVID-19 related TB infection in Sub-Saharan Africa: A systematic review and meta-analysis. Int. Clin. Img. Med. Rew. 2022, 1, 1036. Available online: https://ijcimr.org/articles/IJCIMR-V1-1036.pdf (accessed on 18 May 2022).
- Udwadia, Z.F.; Vora, A.; Tripathi, A.R.; Malu, K.N.; Lange, C.; Raju, R.S. COVID-19 -Tuberculosis interactions: When dark forces collide. Indian J. Tuberc. 2020, 67, S155–S162. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Y.; Chen, Y.; Qin, Q. Unique epidemiological and clinical features of the emerging 2019 novel coronavirus pneumonia (COVID-19) implicate special control measures. J. Med. Virol. 2020, 92, 568–576. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Coronavirus Disease 2019 (COVID-19): Weekly Epidemiological Update, 8 March 2022; WHO: Geneva, Switzerland, 2022; Available online: https://www.who.int/publications/m/item/weekly-epidemiological-update-on-covid-19---8-march-2022 (accessed on 10 March 2022).
- Nalbandian, A.; Sehgal, K.; Gupta, A.; Madhavan, M.V.; McGroder, C.; Stevens, J.S.; Cook, J.R.; Nordvig, A.S.; Shalev, D.; Sehrawat, T.S.; et al. Post-acute COVID-19 syndrome. Nat. Med. 2021, 27, 601–615. [Google Scholar] [CrossRef]
- Datta, S.D.; Talwar, A.; Lee, J.T. A Proposed Framework and Timeline of the Spectrum of Disease Due to SARS-CoV-2 Infection: Illness beyond acute infection and public health implications. JAMA 2020, 324, 2251–2252. [Google Scholar] [CrossRef] [PubMed]
- Wilson, M.S.; A Wynn, T. Pulmonary fibrosis: Pathogenesis, etiology and regulation. Mucosal. Immunol. 2009, 2, 103–121. [Google Scholar] [CrossRef] [Green Version]
- Gause, W.C.; Wynn, T.A.; Allen, J.E. Type 2 immunity and wound healing: Evolutionary refinement of adaptive immunity by helminths. Nat. Rev. Immunol. 2013, 13, 607–614. [Google Scholar] [CrossRef] [Green Version]
- Tanni, S.E.; Fabro, A.T.; de Albuquerque, A.; Ferreira, E.V.M.; Verrastro, C.G.Y.; Sawamura, M.V.Y.; Ribeiro, S.M.; Baldi, B.G. Pulmonary fibrosis secondary to COVID-19: A narrative review. Expert Rev. Respir. Med. 2021, 15, 791–803. [Google Scholar] [CrossRef]
- Liang, J.; Liu, G.; Yu, S.; Yang, Y.; Li, Y.; Tian, H.; Chen, Z.; Gong, W. Dynamic Changes in Chest CT Images Over 167 Days in 11 Patients with COVID-19: A Case Series and Literature Review. Zoonoses 2021, 1, 1–11. [Google Scholar] [CrossRef]
- Safont, B.; Tarraso, J.; Rodriguez-Borja, E.; Fernández-Fabrellas, E.; Sancho-Chust, J.N.; Molina, V.; Lopez-Ramirez, C.; Lope-Martinez, A.; Cabanes, L.; Andreu, A.L.; et al. Lung Function, Radiological Findings and Biomarkers of Fibrogenesis in a Cohort of COVID-19 Patients Six Months After Hospital Discharge. Arch. Bronconeumol. 2021, 58, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Thille, A.W.; Esteban, A.; Fernández-Segoviano, P.; Rodriguez, J.-M.; Aramburu, J.-A.; Vargas-Errázuriz, P.; Martín-Pellicer, A.; Lorente, J.A.; Frutos-Vivar, F. Chronology of histological lesions in acute respiratory distress syndrome with diffuse alveolar damage: A prospective cohort study of clinical autopsies. Lancet Respir. Med. 2013, 1, 395–401. [Google Scholar] [CrossRef]
- Hu, Z.-J.; Xu, J.; Yin, J.-M.; Li, L.; Hou, W.; Zhang, L.-L.; Zhou, Z.; Yu, Y.-Z.; Li, H.-J.; Feng, Y.-M.; et al. Lower Circulating Interferon-Gamma Is a Risk Factor for Lung Fibrosis in COVID-19 Patients. Front. Immunol. 2020, 11, 585647. [Google Scholar] [CrossRef]
- Furuhashi, M.; Moniwa, N.; Takizawa, H.; Ura, N.; Shimamoto, K. Potential differential effects of renin-angiotensin system inhibitors on SARS-CoV-2 infection and lung injury in COVID-19. Hypertens. Res. 2020, 43, 837–840. [Google Scholar] [CrossRef] [PubMed]
- Zuo, W.; Zhao, X.; Chen, Y.-G. SARS Coronavirus and Lung Fibrosis. In Molecular Biology of the SARS-Coronavirus; Lal, S.K., Ed.; Springer: Berlin/Heidelberg, Gremany, 2010. [Google Scholar] [CrossRef] [Green Version]
- Rübe, C.E.; Uthe, D.; Schmid, K.W.; Richter, K.D.; Wessel, J.; Schuck, A.; Willich, N.; Rübe, C. Dose-dependent induction of transforming growth factor β (TGF-β) in the lung tissue of fibrosis-prone mice after thoracic irradiation. Int. J. Radiat. Oncol. Biol. Phys. 2000, 47, 1033–1042. [Google Scholar] [CrossRef]
- Liu, F.; Matsuura, I. Inhibition of Smad Antiproliferative Function by CDK Phosphorylation. Cell Cycle 2005, 4, 63–66. [Google Scholar] [CrossRef] [Green Version]
- Derynck, R.; Akhurst, R. Differentiation plasticity regulated by TGF-β family proteins in development and disease. Nat. Cell Biol. 2007, 9, 1000–1004. [Google Scholar] [CrossRef]
- Barry, C.E., III; Boshoff, H.I.; Dartois, V.; Dick, T.; Ehrt, S.; Flynn, J.; Schnappinger, D.; Wilkinson, R.; Young, D. The spectrum of latent tuberculosis: Rethinking the biology and intervention strategies. Nat. Rev. Microbiol. 2009, 7, 845–855. [Google Scholar] [CrossRef]
- Cadmus, S.I.; Akinseye, V.O.; Taiwo, B.O.; Pinelli, E.O.; van Soolingen, D.; Rhodes, S.G. Interactions between helminths and tuberculosis infections: Implications for tuberculosis diagnosis and vaccination in Africa. PLOS Neglected Trop. Dis. 2020, 14, e0008069. [Google Scholar] [CrossRef] [PubMed]
- Global Tuberculosis Report 2021. Available online: https://www.who.int/publications/digital/global-tuberculosis-report-2021/tb-disease-burden/incidence (accessed on 28 March 2022).
- Chen, Q.; Ghilardi, N.; Wang, H.; Baker, T.; Xie, M.-H.; Gurney, A.; Grewal, I.; De Sauvage, F.J. Development of Th1-type immune responses requires the type I cytokine receptor TCCR. Nature 2000, 407, 916–920. [Google Scholar] [CrossRef] [PubMed]
- Hunter, R.L. Pathology of post primary tuberculosis of the lung: An illustrated critical review. Tuberculosis 2011, 91, 497–509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsenova, L.; Singhal, A. Effects of host-directed therapies on the pathology of tuberculosis. J. Pathol. 2020, 250, 636–646. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Information Note: Tuberculosis and COVID-19. 12 May 2020. Available online: https://www.who.int/docs/default-source/documents/tuberculosis/infonote-tb-covid-19.pdf (accessed on 8 March 2022).
- Gao, Y.; Liu, M.; Chen, Y.; Shi, S.; Geng, J.; Tian, J. Association between tuberculosis and COVID-19 severity and mortality: A rapid systematic review and meta-analysis. J. Med. Virol. 2020, 93, 194–196. [Google Scholar] [CrossRef] [PubMed]
- Oh, T.K.; Song, I.-A. Impact of coronavirus disease-2019 on chronic respiratory disease in South Korea: An NHIS COVID-19 database cohort study. BMC Pulm. Med. 2021, 21, 12. [Google Scholar] [CrossRef]
- Tadolini, M.; Codecasa, L.; García-García, J.-M.; Blanc, F.-X.; Borisov, S.; Alffenaar, J.-W.; Andréjak, C.; Bachez, P.; Bart, P.-A.; Belilovski, E.; et al. Active tuberculosis, sequelae and COVID-19 co-infection: First cohort of 49 cases. Eur. Respir. J. 2020, 56, 2001398. [Google Scholar] [CrossRef]
- Kumar, M.S.; Surendran, D.; Manu, M.S.; Rakesh, P.S.; Balakrishnan, S. Mortality due to TB-COVID-19 coinfection in India. Int. J. Tuberc. Lung Dis. 2021, 25, 250–251. [Google Scholar] [CrossRef] [PubMed]
- Hotez, P.J.; Kamath, A. Neglected Tropical Diseases in Sub-Saharan Africa: Review of Their Prevalence, Distribution, and Disease Burden. PLOS Negl. Trop. Dis. 2009, 3, e412. [Google Scholar] [CrossRef] [Green Version]
- Harris, N.L.; Loke, P. Recent Advances in Type-2-Cell-Mediated Immunity: Insights from Helminth Infection. Immunity 2017, 47, 1024–1036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maizels, R.M. Regulation of immunity and allergy by helminth parasites. Allergy 2020, 75, 524–534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, J.; Jackson, J.A.; Faulkner, H.; Behnke, J.; Else, K.; Kamgno, J.; Boussinesq, M.; Bradley, J.E. Intensity of Intestinal Infection with Multiple Worm Species Is Related to Regulatory Cytokine Output and Immune Hyporesponsiveness. J. Infect. Dis. 2008, 197, 1204–1212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Liu, L.; Liu, X.; Zhang, Y.; Shi, H.; Jia, W.; Zhu, H.; Jia, H.; Liu, M.; Bai, X. Extracellular Vesicles Derived From Trichinella spiralis Muscle Larvae Ameliorate TNBS-Induced Colitis in Mice. Front. Immunol. 2020, 11, 1174. [Google Scholar] [CrossRef]
- Ryan, S.; Shiels, J.; Taggart, C.C.; Dalton, J.P.; Weldon, S. Fasciola hepatica-Derived Molecules as Regulators of the Host Immune Response. Front. Immunol. 2020, 11, 2182. [Google Scholar] [CrossRef] [PubMed]
- Napoli, P.E.; Nioi, M. Global Spread of Coronavirus Disease 2019 and Malaria: An Epidemiological Paradox in the Early Stage of A Pandemic. J. Clin. Med. 2020, 9, 1138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gursel, M.; Gursel, I. Is global BCG vaccination coverage relevant to the progression of SARS-CoV-2 pandemic? Allergy 2020, 75, 1815–1819. [Google Scholar] [CrossRef] [PubMed]
- Hays, R.; Pierce, D.; Giacomin, P.; Loukas, A.; Bourke, P.; McDermott, R. Helminth coinfection and COVID-19: An alternate hypothesis. PLOS Neglected Trop. Dis. 2020, 14, e0008628. [Google Scholar] [CrossRef]
- Fonte, L.; Acosta, A.; Sarmiento, M.E.; Ginori, M.; García, G.; Norazmi, M.N. COVID-19 Lethality in Sub-Saharan Africa and Helminth Immune Modulation. Front. Immunol. 2020, 11, 574910. [Google Scholar] [CrossRef]
- Acosta, A.; Fonte, L.; Sarmiento, M.E.; Norazmi, M.N. Does our Mycobacteriome Influence COVID-19 Morbidity and Lethality? Front. Microbiol. 2021, 12, 589165. [Google Scholar] [CrossRef]
- Zhou, Y.; Fu, B.; Zheng, X.; Wang, D.; Zhao, C.; Qi, Y.; Sun, R.; Tian, Z.; Xu, X.; Wei, H. Pathogenic T-cells and inflammatory monocytes incite inflammatory storms in severe COVID-19 patients. Natl. Sci. Rev. 2020, 7, nwaa041. [Google Scholar] [CrossRef] [Green Version]
- Tcheutchoua, D.N.; Tankeu, A.T.; Angong, D.L.W.; Agoons, B.B.; Nguemnang, N.Y.Y.; Djeunga, H.C.N.; Kamgno, J. Unexpected low burden of coronavirus disease 2019 (COVID-19) in sub-Saharan Africa region despite disastrous predictions: Reasons and perspectives. Pan Afr. Med. J. 2020, 37, 352. [Google Scholar] [CrossRef] [PubMed]
- Abdoli, A. Helminths and COVID-19 Co-Infections: A Neglected Critical Challenge. ACS Pharmacol. Transl. Sci. 2020, 3, 1039–1041. [Google Scholar] [CrossRef] [PubMed]
- Cepon-Robins, T.J.; Gildner, T.E. Old friends meet a new foe. Evol. Med. Public Health 2020, 2020, 234–248. [Google Scholar] [CrossRef] [PubMed]
- Parker, W.; Sarafian, J.T.; A Broverman, S.; Laman, J.D. Between a hygiene rock and a hygienic hard place. Evol. Med. Public Health 2021, 9, 120–130. [Google Scholar] [CrossRef] [PubMed]
- Wolday, D.; Gebrecherkos, T.; Arefaine, Z.G.; Kiros, Y.K.; Gebreegzabher, A.; Tasew, G.; Abdulkader, M.; Abraha, H.E.; Desta, A.A.; Hailu, A.; et al. Effect of co-infection with intestinal parasites on COVID-19 severity: A prospective observational cohort study. eClinicalMedicine 2021, 39, 101054. [Google Scholar] [CrossRef] [PubMed]
- Dheda, K.; Perumal, T.; Moultrie, H.; Perumal, R.; Esmail, A.; Scott, A.J.; Udwadia, Z.; Chang, K.C.; Peter, J.; Pooran, A.; et al. The intersecting pandemics of tuberculosis and COVID-19: Population-level and patient-level impact, clinical presentation, and corrective interventions. Lancet Respir. Med. 2022, 10, 603–622. [Google Scholar] [CrossRef]
- Wait, L.F.; Dobson, A.P.; Graham, A.L. Do parasite infections interfere with immunisation? A review and meta-analysis. Vaccine 2020, 38, 5582–5590. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fonte, L.; Acosta, A.; Sarmiento, M.E.; Norazmi, M.N.; Ginori, M.; de Armas, Y.; Calderón, E.J. Overlapping of Pulmonary Fibrosis of Postacute COVID-19 Syndrome and Tuberculosis in the Helminth Coinfection Setting in Sub-Saharan Africa. Trop. Med. Infect. Dis. 2022, 7, 157. https://doi.org/10.3390/tropicalmed7080157
Fonte L, Acosta A, Sarmiento ME, Norazmi MN, Ginori M, de Armas Y, Calderón EJ. Overlapping of Pulmonary Fibrosis of Postacute COVID-19 Syndrome and Tuberculosis in the Helminth Coinfection Setting in Sub-Saharan Africa. Tropical Medicine and Infectious Disease. 2022; 7(8):157. https://doi.org/10.3390/tropicalmed7080157
Chicago/Turabian StyleFonte, Luis, Armando Acosta, María E. Sarmiento, Mohd Nor Norazmi, María Ginori, Yaxsier de Armas, and Enrique J. Calderón. 2022. "Overlapping of Pulmonary Fibrosis of Postacute COVID-19 Syndrome and Tuberculosis in the Helminth Coinfection Setting in Sub-Saharan Africa" Tropical Medicine and Infectious Disease 7, no. 8: 157. https://doi.org/10.3390/tropicalmed7080157
APA StyleFonte, L., Acosta, A., Sarmiento, M. E., Norazmi, M. N., Ginori, M., de Armas, Y., & Calderón, E. J. (2022). Overlapping of Pulmonary Fibrosis of Postacute COVID-19 Syndrome and Tuberculosis in the Helminth Coinfection Setting in Sub-Saharan Africa. Tropical Medicine and Infectious Disease, 7(8), 157. https://doi.org/10.3390/tropicalmed7080157