

Leptospira borgpetersenii Leucine-Rich Repeat Proteins and Derived Peptides in an Indirect ELISA Development for the Diagnosis of Canine Leptospiral Infections

 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Dog Plasma Samples
2.2. Recombinant Proteins
2.3. Epitope Peptide Prediction
2.4. Rabbit Hyperimmune Sera
2.5. Identification of Leptospira in Plasma Samples by Real-Time PCR
2.6. Line Blot Immunodetection of rKU_Sej_LRR_2012M Protein by Rabbit Hyperimmune Sera
2.7. Immunodetection of rKU_Sej_LRR_2012M and Its Derived Peptides by ELISA
2.8. Canine Plasma Samples Leptospira Detection by an Indirect Peptide-Based ELISA
2.9. Microscopic Agglutination Test (MAT)
2.10. Statistical Analysis
3. Results
3.1. PCR Detection of Leptospira in Dog Plasma Samples
3.2. Line Blots Immunodetection against the Recombinant Protein by Rabbit Hyperimmune Sera
3.3. Prediction on LRR Epitope Peptides
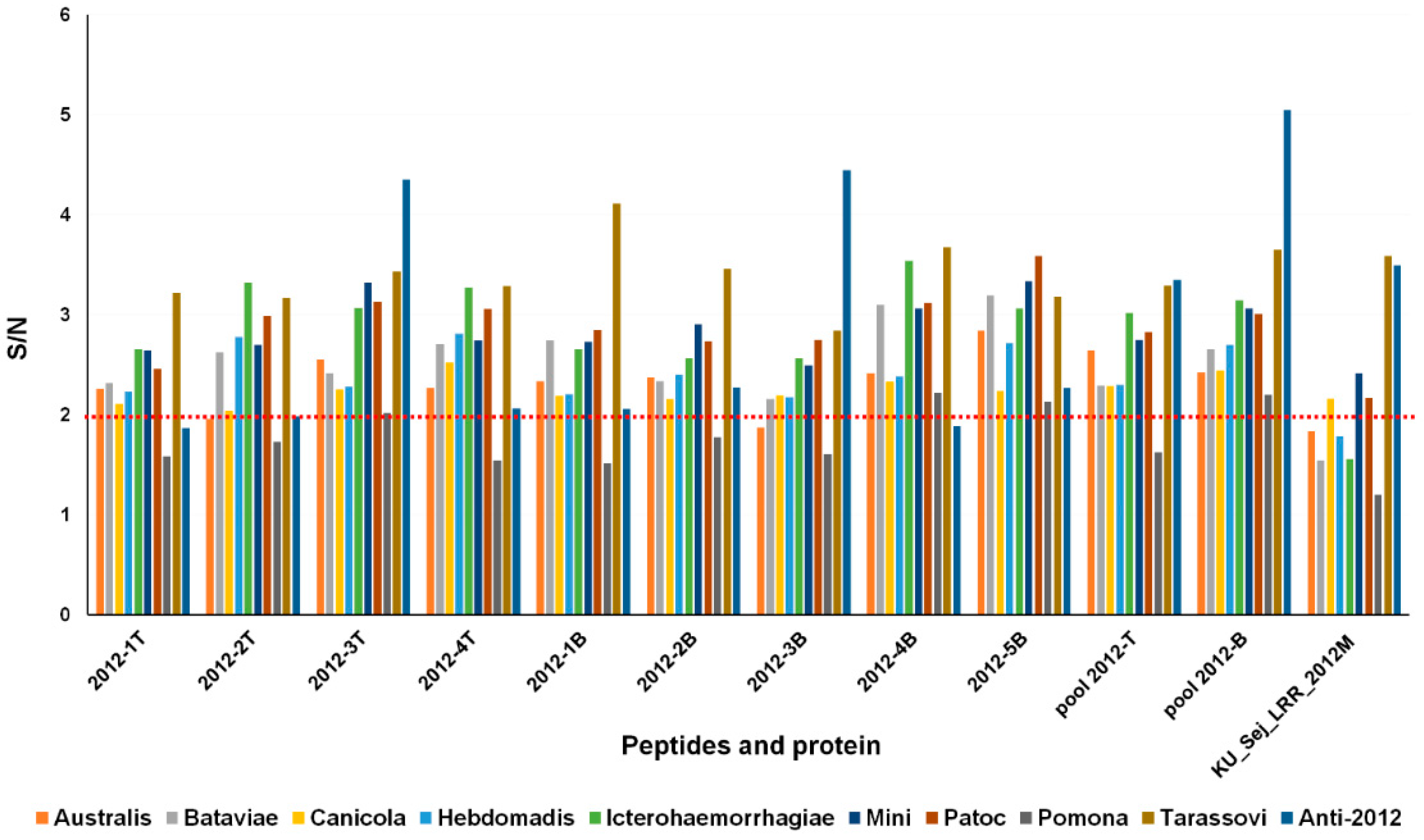
3.4. Selection of Antigenic Peptides
3.5. Peptide-Based ELISA of Canine Antisera
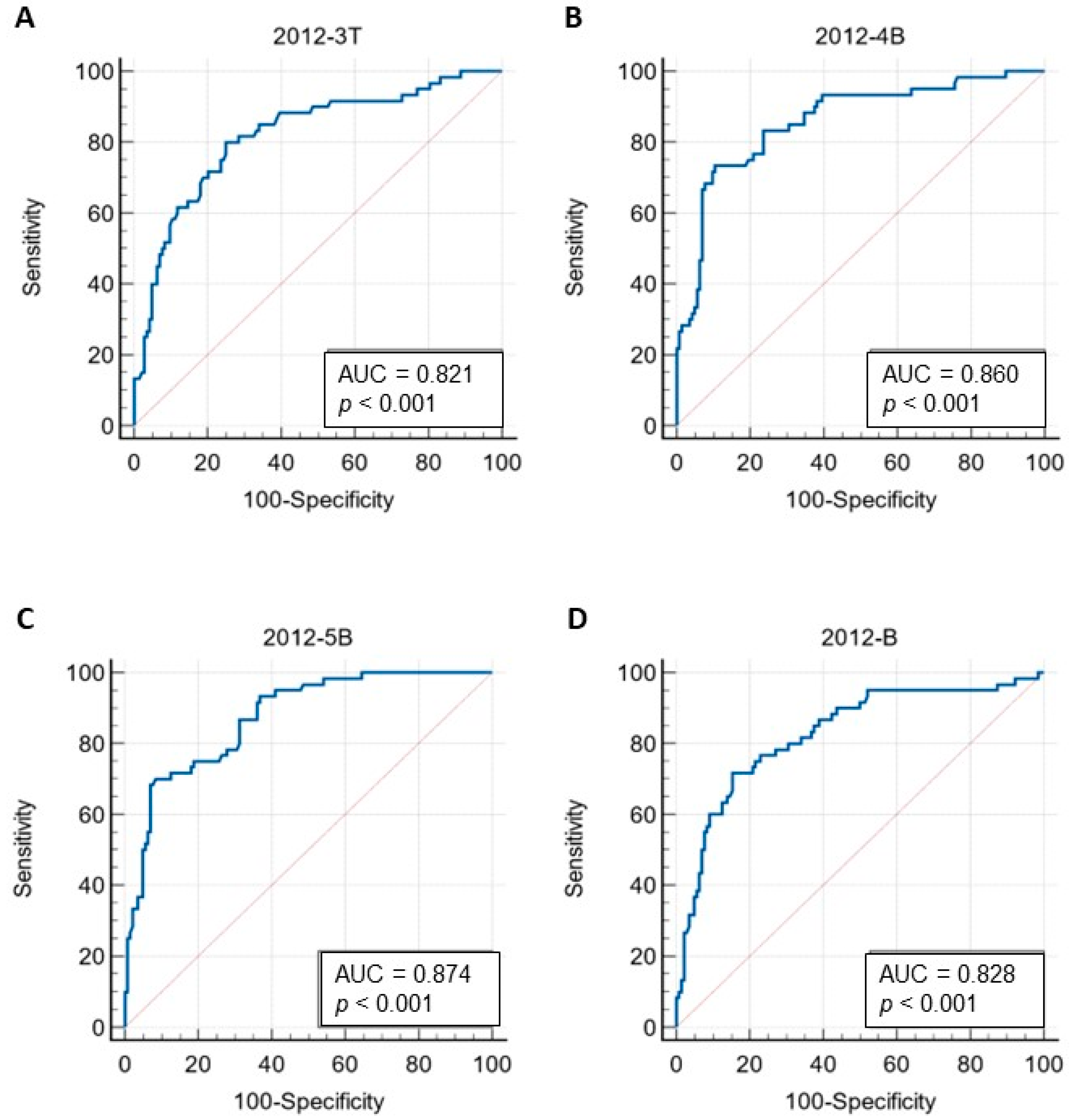
3.6. The MAT Validation on the Developed ELISAs Results
3.7. Association between Type of Dogs and Leptospirosis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Samrot, A.V.; Sean, T.C.; Bhavya, K.S.; Sahithya, C.S.; Chan-Drasekaran, S.; Palanisamy, R.; Robinson, E.R.; Subbiah, S.K.; Mok, P.L. Leptospiral Infection, Pathogenesis and Its Diagnosis–A Review. Pathogens 2021, 10, 145. [Google Scholar] [CrossRef]
- Vincent, A.T.; Schiettekatte, O.; Goarant, C.; Neela, V.K.; Bernet, E.; Thibeaux, R.; Ismail, N.; Mohd Khalid, M.K.N.; Amran, F.; Masuzawa, T.; et al. Revisiting the taxonomy and evolution of pathogenicity of the genus Leptospira through the prism of genomics. PLoS Negl. Trop. Dis. 2019, 13, e0007270. [Google Scholar] [CrossRef]
- Brown, K.; Prescott, J. Leptospirosis in the family dog: A public health perspective. Cmaj 2008, 178, 399–401. [Google Scholar] [CrossRef]
- Costa, F.; Hagan, J.E.; Calcagno, J.; Kane, M.; Torgerson, P.; Martinez-Silveira, M.S.; Stein, C.; Abela-Ridder, B.; Ko, A.I. Global Morbidity and Mortality of Leptospirosis: A Systematic Review. PLoS Negl. Trop. Dis. 2015, 9, e0003898. [Google Scholar] [CrossRef] [PubMed]
- Disease Notification Report (Leptospirosis). Available online: http://www.boe.moph.go.th/boedb/surdata/disease.php?ds=43 (accessed on 1 September 2021).
- Population of Dog and Cat in 2016. Available online: http://dcontrol.dld.go.th/webnew/index.php/th/news-menu/2018-07-02-08-24-32/360-dogpop2016 (accessed on 9 August 2016).
- Jittapalapong, S.; Sittisan, P.; Sakpuaram, T.; Kabeya, H.; Maruyama, S.; Inpankaew, T. Coinfection of Leptospira spp and Toxoplasma gondii among stray dogs in Bangkok, Thailand. Southeast Asian J. Trop. Med. Public Health 2009, 40, 247–252. [Google Scholar] [PubMed]
- Aslantaş, O.; Ozdemir, V.; Kiliç, S.; Babür, C. Seroepidemiology of leptospirosis, toxoplasmosis, and leishmaniosis among dogs in Ankara, Turkey. Vet. Parasitol. 2005, 129, 187–191. [Google Scholar] [CrossRef] [PubMed]
- Weekes, C.C.; Everard, C.O.; Levett, P.N. Seroepidemiology of canine leptospirosis on the island of Barbados. Vet. Microbiol. 1997, 57, 215–222. [Google Scholar] [CrossRef]
- De Paula Dreer, M.K.; Gonçalves, D.D.; da Silva Caetano, I.C.; Gerônimo, E.; Menegas, P.H.; Bergo, D.; Ruiz Lopes-Mori, F.M.; Benitez, A.; de Freitas, J.C.; Evers, F.; et al. Toxoplasmosis, leptospirosis and brucellosis in stray dogs housed at the shelter in Umuarama municipality, Paraná, Brazil. J. Venom. Anim. Toxins Incl. Trop. Dis. 2013, 19, 23. [Google Scholar] [CrossRef]
- Ellis, W.A. Animal leptospirosis. In Leptospira and Leptospirosis; Adler, B., Ed.; Springer: Berlin/Heidelberg, Germany, 2015; pp. 99–137. [Google Scholar]
- Goldstein, R.E. Canine leptospirosis. Vet. Clin. N. Am. Small Anim. Pract. 2010, 40, 1091–1101. [Google Scholar] [CrossRef]
- Altheimer, K.; Jongwattanapisan, P.; Luengyosluechakul, S.; Pusoonthornthum, R.; Prapasarakul, N.; Kurilung, A.; Broens, E.M.; Wagenaar, J.A.; Goris, M.G.A.; Ahmed, A.A.; et al. Leptospira infection and shedding in dogs in Thailand. BMC Vet. Res. 2020, 16, 89. [Google Scholar] [CrossRef]
- Ngasaman, R.; Saechan, V.; Prachantasena, S.; Yingkajorn, M.; Sretrirutchai, S. Investigation of Leptospira Infection in Stray Animals in Songkhla, Thailand: Leptospirosis Risk Reduction in Human. Vector Borne Zoonotic Dis. 2020, 20, 432–435. [Google Scholar] [CrossRef] [PubMed]
- Meeyam, T.; Tablerk, P.; Petchanok, B.; Pichpol, D.; Padungtod, P. Seroprevalence and risk factors associated with leptospirosis in dogs. Southeast Asian J. Trop Med. Public Health 2006, 37, 148–153. [Google Scholar]
- Chotivanich, S.; Pinprasong, B.; Suwancharoen, D. Serologic survey of leptospirosis in pigs, dogs, beef cattle, buffaloes and dairy cattle in Chaiyaphum Province. DLD Tech. Paps. 2000, 5, 1–13. [Google Scholar]
- Pumipuntu, N.; Suwannarong, K. Seroprevalence of Leptospira Spp. in Cattle and Dogs in Mahasarakham Province, Thailand. J. Health Res. 2016, 30, 223–226. [Google Scholar]
- Niwetpathomwat, A.; Assarasakorn, S. Preliminary investigation of canine leptospirosis in a rural area of Thailand. Med. Weter. 2007, 63, 59–61. [Google Scholar]
- Sakpuaram, T.; Wajjwalku, W.; Sirinarumithra, T.; Kasemsuwan, S.; Sithisara, P.; Chanthick, N. Monitoring of Leptospirosis in healthy vaccinated dogs in Hua-Hin district, Prachuapkhirikhan province between January to December, 2004. In Proceedings of the Kasetsart University Annual Academic Conference, Bangkok, Thailand, 30 January–2 February 2006; p. 586. [Google Scholar]
- Kurilung, A.; Chanchaithong, P.; Lugsomya, K.; Niyomtham, W.; Wuthiekanun, V.; Prapasarakul, N. Molecular detection and isolation of pathogenic Leptospira from asymptomatic humans, domestic animals and water sources in Nan province, a rural area of Thailand. Res. Vet. Sci. 2017, 115, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Adler, B. Vaccines against leptospirosis. Curr. Top. Microbiol. Immunol. 2015, 387, 251–272. [Google Scholar] [CrossRef]
- Cao, Y.; Faisal, S.M.; Yan, W.; Chang, Y.-C.; McDonough, S.P.; Zhang, N.; Akey, B.L.; Chang, Y.-F. Evaluation Of Novel Fusion Proteins Derived From Extracellular Matrix Binding Domains Of Ligb As Vaccine Candidates Against Leptospirosis In A Hamster Model. Vaccine 2011, 29, 7379–7386. [Google Scholar] [CrossRef]
- Deveson Lucas, D.S.; Lo, M.; Bulach, D.M.; Quinsey, N.S.; Murray, G.L.; Allen, A.; Adler, B. Recombinant LipL32 stimulates interferon-gamma production in cattle vaccinated with a monovalent Leptospira borgpetersenii serovar Hardjo subtype Hardjobovis vaccine. Vet. Microbiol. 2014, 169, 163–170. [Google Scholar] [CrossRef]
- Haake, D.A.; Mazel, M.K.; McCoy, A.M.; Milward, F.; Chao, G.; Matsunaga, J.; Wagar, E.A. Leptospiral outer membrane proteins OmpL1 and LipL41 exhibit synergistic immunoprotection. Infect. Immun. 1999, 67, 6572–6582. [Google Scholar] [CrossRef] [PubMed]
- Yan, W.W.; Faisal, S.M.; McDonough, S.P.; Divers, T.J.; Barr, S.C.; Chang, C.F.; Pan, M.J.; Chang, Y.F. Immunogenicity and protective efficacy of recombinant Leptospira immunoglobulin-like protein B (rLigB) in a hamster challenge model. Microbes Infect. 2009, 11, 230–237. [Google Scholar] [CrossRef]
- Matsui, M.; Soupé, M.-E.; Becam, J.; Goarant, C. Differential In Vivo Gene Expression of Major Leptospira Proteins in Resistant or Susceptible Animal Models. Appl. Environ. Microbiol. 2012, 78, 6372–6376. [Google Scholar] [CrossRef] [PubMed]
- Haake, D.A.; Matsunaga, J. Leptospiral Immunoglobulin-Like Domain Proteins: Roles in Virulence and Immunity. Front. Immunol. 2021, 11, 579907. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, J.S.; Fouts, D.E.; Haft, D.H.; Cannella, A.P.; Ricaldi, J.N.; Brinkac, L.; Harkins, D.; Durkin, S.; Sanka, R.; Sutton, G.; et al. Pathogenomic Inference of Virulence-Associated Genes in Leptospira interrogans. PLoS Negl. Trop. Dis. 2013, 7, e2468. [Google Scholar] [CrossRef]
- Doxey, A.C.; McConkey, B.J. Prediction of molecular mimicry candidates in human pathogenic bacteria. Virulence 2013, 4, 453–466. [Google Scholar] [CrossRef] [PubMed]
- Hsu, S.-H.; Chou, L.-F.; Hong, C.-H.; Chang, M.-Y.; Tsai, C.-Y.; Tian, Y.-C.; Yang, H.-Y.; Yang, C.-W. Crosstalk between E-Cadherin/β-Catenin and NF-κB Signaling Pathways: The Regulation of Host-Pathogen Interaction during Leptospirosis. Int. J. Mol. Sci. 2021, 22, 13132. [Google Scholar] [CrossRef] [PubMed]
- Hsu, S.-H.; Yang, C.-W. Insight into the Structure, Functions, and Dynamics of the Leptospira Outer Membrane Proteins with the Pathogenicity. Membranes 2022, 12, 300. [Google Scholar] [CrossRef] [PubMed]
- Kobe, B.; Kajava, A.V. The leucine-rich repeat as a protein recognition motif. Curr. Opin. Struct. Biol. 2001, 11, 725–732. [Google Scholar] [CrossRef]
- Gaillard, J.L.; Berche, P.; Frehel, C.; Gouln, E.; Cossart, P. Entry of L. monocytogenes into cells is mediated by internalin, a repeat protein reminiscent of surface antigens from gram-positive cocci. Cell 1991, 65, 1127–1141. [Google Scholar] [CrossRef]
- Parida, S.K.; Domann, E.; Rohde, M.; Müller, S.; Darji, A.; Hain, T.; Wehland, J.; Chakraborty, T. Internalin B is essential for adhesion and mediates the invasion of Listeria monocytogenes into human endothelial cells. Mol. Microbiol. 1998, 28, 81–93. [Google Scholar] [CrossRef] [PubMed]
- Sabet, C.; Lecuit, M.; Cabanes, D.; Cossart, P.; Bierne, H. LPXTG Protein InlJ, a Newly Identified Internalin Involved in Listeria monocytogenes Virulence. Infect. Immun. 2005, 73, 6912–6922. [Google Scholar] [CrossRef] [PubMed]
- Sawyer, R.T.; Drevets, D.A.; Campbell, P.A.; Potter, T.A. Internalin A can mediate phagocytosis of Listeria monocytogenes by mouse macrophage cell lines. J. Leukoc. Biol. 1996, 60, 603–610. [Google Scholar] [CrossRef] [PubMed]
- McDonald, C.; Vacratsis, P.O.; Bliska, J.B.; Dixon, J.E. The Yersinia Virulence Factor YopM Forms a Novel Protein Complex with Two Cellular Kinases. J. Biol. Chem. 2003, 278, 18514–18523. [Google Scholar] [CrossRef] [PubMed]
- Ye, Z.; Kerschen, E.J.; Cohen, D.A.; Kaplan, A.M.; van Rooijen, N.; Straley, S.C. Gr1+ Cells Control Growth of YopM-Negative Yersinia pestis during Systemic Plague. Infect. Immun. 2009, 77, 3791–3806. [Google Scholar] [CrossRef] [PubMed]
- Helft, L.; Reddy, V.; Chen, X.; Koller, T.; Federici, L.; Fernández-Recio, J.; Gupta, R.; Bent, A. LRR Conservation Mapping to Predict Functional Sites within Protein Leucine-Rich Repeat Domains. PLoS ONE 2011, 6, e21614. [Google Scholar] [CrossRef]
- Hniman, A.; Prapong, S. Development of leptospira molecular markers by using bioinformation from predicted Leucine-Rich Repeat (LRR) protein genes. J. Thai Vet. Med. Assoc. 2007, 58, 65–78. [Google Scholar]
- Nitipan, S.; Sritrakul, T.; Kunjantarachot, A.; Prapong, S. Identification of epitopes in Leptospira borgpetersenii leucine-rich repeat proteins. Infect. Genet. Evol. 2013, 14, 46–57. [Google Scholar] [CrossRef]
- Nitipan, S. Molecular Cloning and Sequence Analysis of Gene Encoding Leucine-Rich Repeat proteins of Pathogenic Leptospira borgpetersenii. Ph.D. Thesis, Kasetsart University, Bangkok, Thailand, 2013. [Google Scholar]
- Suphatpahirapol, C.; Nguyen, T.-H.; Tansiri, Y.; Yingchutrakul, Y.; Roytrakul, S.; Nitipan, S.; Wajjwalku, W.; Haltrich, D.; Prapong, S.; Keawsompong, S. Expression of a leptospiral leucine-rich repeat protein using a food-grade vector in Lactobacillus plantarum, as a strategy for vaccine delivery. 3 Biotech 2019, 9, 324. [Google Scholar] [CrossRef]
- Suphatpahirapol, C. Leptospiral Protein Expression using Lactic Acid Bacteria-based Expression Systems for Mucosal Vaccine Delivery. Ph.D. Thesis, Kasetsart University, Bangkok, Thailand, 2017. [Google Scholar]
- Sritrakul, T.; Nitipan, S.; Wajjwalku, W.; La-ard, A.; Suphatpahirapol, C.; Petkarnjanapong, W.; Ongphiphadhanakul, B.; Prapong, S. Leptospira borgpetersenii hybrid leucine-rich repeat protein: Cloning and expression, immunogenic identification and molecular docking evaluation. J. Microbiol. Methods 2017, 142, 52–62. [Google Scholar] [CrossRef] [PubMed]
- Vita, R.; Mahajan, S.; Overton, J.A.; Dhanda, S.K.; Martini, S.; Cantrell, J.R.; Wheeler, D.K.; Sette, A.; Peters, B. The Immune Epitope Database (IEDB): 2018 update. Nucleic Acids Res. 2019, 47, D339–D343. [Google Scholar] [CrossRef]
- Saha, S.; Raghava, G.P.S. BcePred: Prediction of continuous B-cell epitopes in antigenic sequences using physico-chemical properties. In Artificial Immune Systems, Proceedings of the Third International Conference, ICARIS 2004, Catania, Italy, 13–16 September 2004; Springer: Berlin/Heidelberg, Germany, 2004; pp. 197–204. [Google Scholar]
- Jespersen, M.C.; Peters, B.; Nielsen, M.; Marcatili, P. BepiPred-2.0: Improving sequence-based B-cell epitope prediction using conformational epitopes. Nucleic Acids Res. 2017, 45, W24–W29. [Google Scholar] [CrossRef] [PubMed]
- Reche, P.A.; Lafuente, E.M.; Perosanz, M.G.; Sanchez, J.L.; Ras-Carmona, A.; Alipour, T.F.; Pelaez, H.F. Predicted Antigenic Peptides. Available online: http://imed.med.ucm.es/Tools/antigenic.pl (accessed on 13 May 2020).
- Saha, S.; Raghava, G.P. Prediction of continuous B-cell epitopes in an antigen using recurrent neural network. Proteins 2006, 65, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Saha, S.; Raghava, G.P. Prediction methods for B-cell epitopes. Methods Mol. Biol. 2007, 409, 387–394. [Google Scholar] [CrossRef] [PubMed]
- Rice, P.; Longden, I.; Bleasby, A. EMBOSS: The European Molecular Biology Open Software Suite. Trends Genet. 2000, 16, 276–277. [Google Scholar] [CrossRef]
- Yao, B.; Zhang, L.; Liang, S.; Zhang, C. SVMTriP: A Method to Predict Antigenic Epitopes Using Support Vector Machine to Integrate Tri-Peptide Similarity and Propensity. PLoS ONE 2012, 7, e45152. [Google Scholar] [CrossRef]
- Guan, P.; Doytchinova, I.A.; Zygouri, C.; Flower, D.R. MHCPred: Bringing a quantitative dimension to the online prediction of MHC binding. Appl. Bioinform. 2003, 2, 63–66. [Google Scholar]
- Guan, P.; Doytchinova, I.A.; Zygouri, C.; Flower, D.R. MHCPred: A server for quantitative prediction of peptide-MHC binding. Nucleic Acids Res. 2003, 31, 3621–3624. [Google Scholar] [CrossRef]
- Hattotuwagama, C.K.; Guan, P.; Doytchinova, I.A.; Zygouri, C.; Flower, D.R. Quantitative online prediction of peptide binding to the major histocompatibility complex. J. Mol. Graph. Model 2004, 22, 195–207. [Google Scholar] [CrossRef]
- Lata, S.; Bhasin, M.; Raghava, G.P. Application of machine learning techniques in predicting MHC binders. Methods Mol. Biol. 2007, 409, 201–215. [Google Scholar] [CrossRef]
- Nielsen, M.; Lundegaard, C.; Worning, P.; Lauemøller, S.L.; Lamberth, K.; Buus, S.; Brunak, S.; Lund, O. Reliable prediction of T-cell epitopes using neural networks with novel sequence representations. Protein Sci. 2003, 12, 1007–1017. [Google Scholar] [CrossRef]
- Andreatta, M.; Nielsen, M. Gapped sequence alignment using artificial neural networks: Application to the MHC class I system. Bioinformatics 2016, 32, 511–517. [Google Scholar] [CrossRef] [PubMed]
- Combet, C.; Blanchet, C.; Geourjon, C.; Deléage, G. NPS@: Network protein sequence analysis. Trends Biochem. Sci. 2000, 25, 147–150. [Google Scholar] [CrossRef]
- Zhang, Y. I-TASSER server for protein 3D structure prediction. BMC Bioinform. 2008, 9, 40. [Google Scholar] [CrossRef]
- Roy, A.; Kucukural, A.; Zhang, Y. I-TASSER: A unified platform for automated protein structure and function prediction. Nat. Protoc. 2010, 5, 725–738. [Google Scholar] [CrossRef]
- Yang, J.; Yan, R.; Roy, A.; Xu, D.; Poisson, J.; Zhang, Y. The I-TASSER Suite: Protein structure and function prediction. Nat. Methods 2015, 12, 7–8. [Google Scholar] [CrossRef]
- Doytchinova, I.A.; Flower, D.R. VaxiJen: A server for prediction of protective antigens, tumour antigens and subunit vaccines. BMC Bioinform. 2007, 8, 4. [Google Scholar] [CrossRef]
- Doytchinova, I.A.; Flower, D.R. Identifying candidate subunit vaccines using an alignment-independent method based on principal amino acid properties. Vaccine 2007, 25, 856–866. [Google Scholar] [CrossRef]
- Doytchinova, I.; Flower, D. Bioinformatic Approach for Identifying Parasite and Fungal Candidate Subunit Vaccines. Open Vaccine J. 2008, 1, 22–26. [Google Scholar] [CrossRef]
- Mérien, F.; Amouriaux, P.; Perolat, P.; Baranton, G.; Saint Girons, I. Polymerase chain reaction for detection of Leptospira spp. in clinical samples. J. Clin. Microbiol. 1992, 30, 2219–2224. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.A.; Sandai, D.A.; Musa, S.; Hoe, C.H.; Riadzi, M.; Lau, K.L.; Tang, T.H. Rapid diagnosis of leptospirosis by multiplex PCR. Malays J. Med. Sci. 2012, 19, 9–16. [Google Scholar] [PubMed]
- Brenner, D.J.; McWhorter, A.C.; Knutson, J.K.L.; Steigerwalt, A.G. Escherichia vulneris: A new species of Enterobacteriaceae associated with human wounds. J. Clin. Microbiol. 1982, 15, 1133–1140. [Google Scholar] [CrossRef] [PubMed]
- Tansiri, Y.; Sritrakul, T.; Saparpakorn, P.; Boondamnern, T.; Chimprasit, A.; Sripattanakul, S.; Hannongbua, S.; Prapong, S. New potent epitopes from Leptospira borgpetersenii for the stimulation of humoral and cell-mediated immune responses: Experimental and theoretical studies. Inform. Med. Unlocked 2021, 25, 100649. [Google Scholar] [CrossRef]
- Metz, C.E. Basic principles of ROC analysis. Semin. Nucl. Med. 1978, 8, 283–298. [Google Scholar] [CrossRef]
- Zweig, M.H.; Campbell, G. Receiver-operating characteristic (ROC) plots: A fundamental evaluation tool in clinical medicine. Clin. Chem. 1993, 39, 561–577. [Google Scholar] [CrossRef] [PubMed]
- Baker, M. Reproducibility crisis: Blame it on the antibodies. Nature 2015, 521, 274–276. [Google Scholar] [CrossRef] [PubMed]
- Levett, P.N. Leptospirosis. Clin. Microbiol. Rev. 2001, 14, 296–326. [Google Scholar] [CrossRef]
- Mandrekar, J.N. Receiver Operating Characteristic Curve in Diagnostic Test Assessment. J. Thorac. Oncol. 2010, 5, 1315–1316. [Google Scholar] [CrossRef]
- Power, M.; Fell, G.; Wright, M. Principles for high-quality, high-value testing. Evid. Based Med. 2013, 18, 5–10. [Google Scholar] [CrossRef]
- Lizer, J.; Velineni, S.; Weber, A.; Krecic, M.; Meeus, P. Evaluation of 3 Serological Tests for Early Detection Of Leptospira-specific Antibodies in Experimentally Infected Dogs. J. Vet. Intern. Med. 2018, 32, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Dey, S.; Mohan, C.M.; Kumar, T.M.A.S.; Ramadass, P.; Nainar, A.M.; Nachimuthu, K. Recombinant LipL32 antigen-based single serum dilution ELISA for detection of canine leptospirosis. Vet. Microbiol. 2004, 103, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Sinha, D.; Chaudhury, P.; Shankar, H.; Srivastava, S.K. Comparative studies on seroepidemiology of canine leptospirosis by micro agglutination test (MAT) and recombinant Lip L32 ELISA. Indian J. Anim. Sci. 2009, 79, 1089–1094. [Google Scholar]
- Joseph, S.; Mini, M.; Sriram, V.; Sathiadevan, A.; T V, A.; Kumar, S. Evaluation of real-time PCR, MAT, and recombinant LipL32-based ELISA for the diagnosis of canine leptospirosis in a disease-endemic South Indian state, Kerala. Turk. J. Vet. Anim. Sci. 2018, 42, 191–197. [Google Scholar] [CrossRef]
- Thongdee, M.; Chaiwattanarungruengpaisan, S.; Lekcharoen, P.; Yimchoho, N.; Buathong, R.; Wiriyarat, W. A Novel Genotype of Leptospira interrogans Recovered from Leptospirosis Outbreak Samples from Southern Thailand. Jpn. J. Infect. Dis. 2019, 72, 343–346. [Google Scholar] [CrossRef]
- Brown, P.D.; Gravekamp, C.; Carrington, D.G.; van de Kemp, H.; Hartskeerl, R.A.; Edwards, C.N.; Everard, C.O.; Terpstra, W.J.; Levett, P.N. Evaluation of the polymerase chain reaction for early diagnosis of leptospirosis. J. Med. Microbiol. 1995, 43, 110–114. [Google Scholar] [CrossRef] [PubMed]
- Minke, J.M.; Bey, R.; Tronel, J.P.; Latour, S.; Colombet, G.; Yvorel, J.; Cariou, C.; Guiot, A.L.; Cozette, V.; Guigal, P.M. Onset and duration of protective immunity against clinical disease and renal carriage in dogs provided by a bi-valent inactivated leptospirosis vaccine. Vet. Microbiol. 2009, 137, 137–145. [Google Scholar] [CrossRef] [PubMed]
- IDEXX, L. Diagnosing and Managing Canine Leptospirosis: IDEXX. Available online: https://www.idexx.com/files/canine-leptospirosis-test-dx-update.pdf (accessed on 14 April 2022).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Symptomatic Dog | Healthy Dog | Positive Real-Time PCR | Total | |
|---|---|---|---|---|
| Stray dog | 2 | 99 | 0 | 101 |
| Family dog | 7 | 96 | 6 | 103 |
| Species | Serogroup | Serovar | Strain |
|---|---|---|---|
| L. interrogans | Australis | Australis | Ballico |
| Bataviae | Bataviae | Swart | |
| Canicola | Canicola | Hond Utrecht lV | |
| Hebdomadis | Hebdomadis | Hebdomadis | |
| Icterohaemorrhagiae | Icterohaemorrhagiae | Ictero l | |
| Pomona | Pomona | Pomona | |
| L. biflexa | Semaranga | Patoc | Patoc l |
| L. borgpetersenii | Mini | Mini | Sari |
| Tarassovi | Tarassovi | Perepelitsin |
| LRR Protein Derived Peptide Name: Amino Acid Sequence | Length | Sequence Position | Antigenicity Score [Vaxijen] |
|---|---|---|---|
| 2012-1T: YKKAGSAAA | 9 | 19–27 | 0.977 |
| 2012-2T: KLSQGTEVR | 9 | 73–81 | 1.893 |
| 2012-3T: EVRFDWLTSL | 10 | 79–88 | 1.355 |
| 2012-4T: KLKELKVPS | 9 | 215–223 | 0.551 |
| 2012-1B: LDEKDSATESN | 11 | 45–55 | 1.672 |
| 2012-2B: KDSATESNIDSLS | 13 | 48–60 | 1.142 |
| 2012-3B: SAPPSDKKLSQGTEVR | 16 | 66–81 | 0.935 |
| 2012-4B: LYPREGKNASK | 11 | 104–114 | 1.316 |
| 2012-5B: GSNNSIKDLS | 10 | 155–164 | 0.836 |
| Peptide | Cut Off | AUC (95% CI) | Sn | Sp | PPV | NPV | Ac | K (95% CI) |
|---|---|---|---|---|---|---|---|---|
| 2012-3T | 1.037 | 0.821 (0.754–0.887) | 0.800 | 0.750 | 0.571 | 0.900 | 0.765 | 0.492 (0.372–0.613) |
| 2012-4B | 1.020 | 0.860 (0.802–0.919) | 0.733 | 0.896 | 0.746 | 0.890 | 0.848 | 0.632 (0.515–0.750) |
| 2012-5B | 1.057 | 0.874 (0.824–0.924) | 0.700 | 0.917 | 0.778 | 0.880 | 0.853 | 0.635 (0.517–0.754) |
| 2012-B | 0.917 | 0.828 (0.762–0.893) | 0.717 | 0.847 | 0.662 | 0.878 | 0.809 | 0.550 (0.426–0.675) |
| Methods | Type of Dogs (No. of Dogs with Positive Results) | ||||
|---|---|---|---|---|---|
| Positive PCR Dogs (6) | Dogs with Positive Results by Peptide-Based and LipL32 ELISAs (5) | Dogs with Negative Results by Peptide-Based and LipL32 ELISAs (5) | Vaccinated Dogs (20) | ||
| MAT (serovar with titer ≥ 1:100) | Autumnalis | 1 | - | - | - |
| Bratislava | - | 1 | - | 8 | |
| Canicola | 4 | 2 | - | 11 | |
| Icterohaemorrhagiae | 3 | 1 | - | 13 * | |
| Pomona | - | - | - | 6 | |
| Ballum | - | 1 | - | - | |
| Sejroe | 5 | 4 | - | - | |
| Tarassovi | 4 | 1 | - | - | |
| Grippotyphosa | - | - | - | 3 | |
| Patoc | 3 | 3 | - | - | |
| Peptide-based ELISAs | 2012-4B | 6 | 5 | 0 | 3 * |
| 2012-5B | 6 | 5 | 0 | 3 * | |
| Protein ELISA | LipL32 | 6 | 5 | 0 | 19 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sripattanakul, S.; Prapong, T.; Kamlangdee, A.; Katzenmeier, G.; Haltrich, D.; Hongprayoon, R.; Prapong, S. Leptospira borgpetersenii Leucine-Rich Repeat Proteins and Derived Peptides in an Indirect ELISA Development for the Diagnosis of Canine Leptospiral Infections. Trop. Med. Infect. Dis. 2022, 7, 311. https://doi.org/10.3390/tropicalmed7100311
Sripattanakul S, Prapong T, Kamlangdee A, Katzenmeier G, Haltrich D, Hongprayoon R, Prapong S. Leptospira borgpetersenii Leucine-Rich Repeat Proteins and Derived Peptides in an Indirect ELISA Development for the Diagnosis of Canine Leptospiral Infections. Tropical Medicine and Infectious Disease. 2022; 7(10):311. https://doi.org/10.3390/tropicalmed7100311
Chicago/Turabian StyleSripattanakul, Sineenat, Teerasak Prapong, Attapon Kamlangdee, Gerd Katzenmeier, Dietmar Haltrich, Ratchanee Hongprayoon, and Siriwan Prapong. 2022. "Leptospira borgpetersenii Leucine-Rich Repeat Proteins and Derived Peptides in an Indirect ELISA Development for the Diagnosis of Canine Leptospiral Infections" Tropical Medicine and Infectious Disease 7, no. 10: 311. https://doi.org/10.3390/tropicalmed7100311
APA StyleSripattanakul, S., Prapong, T., Kamlangdee, A., Katzenmeier, G., Haltrich, D., Hongprayoon, R., & Prapong, S. (2022). Leptospira borgpetersenii Leucine-Rich Repeat Proteins and Derived Peptides in an Indirect ELISA Development for the Diagnosis of Canine Leptospiral Infections. Tropical Medicine and Infectious Disease, 7(10), 311. https://doi.org/10.3390/tropicalmed7100311