
A Review of the Environmental Trigger and Transmission Components for Prediction of Cholera

 , , ,
, , ,
Abstract
:1. Introduction
2. Vibrio cholerae and Its Natural Habitat
3. Trigger and Transmission Components for Prediction of Cholera

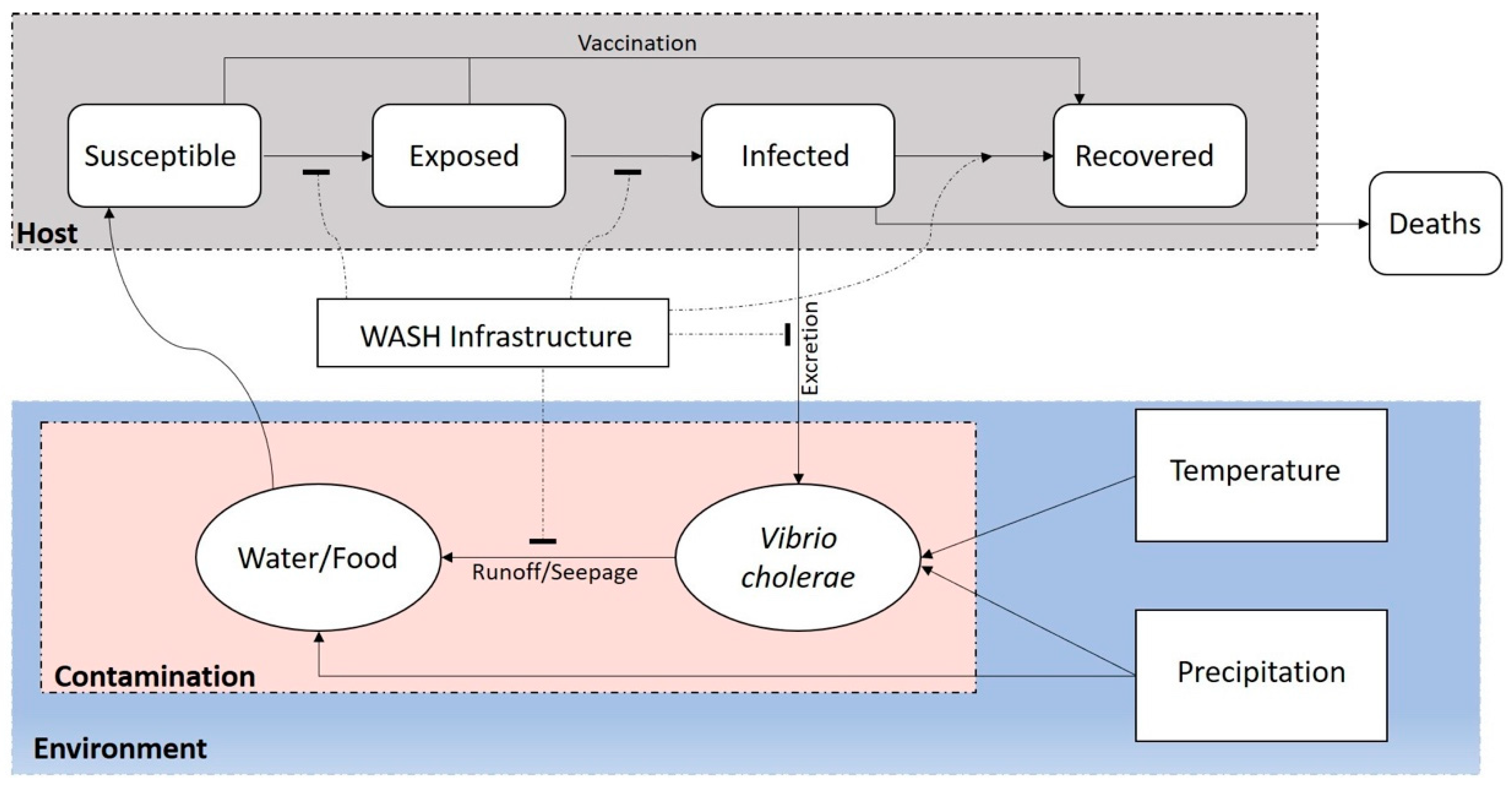{kind=link}
| Author(s) | Study Descriptions/Methodology | Important Findings and Outcomes |
|---|---|---|
| Codeço 2001 [18] | Proposed mathematical model to explain the dynamics of epidemic and endemic cholera. This study is one of the first applications of the SIR model for cholera transmission. |
|
| Wang et al., 2015 [83] | Separated ordinary differential equation (ODE) and reaction-convection-diffusion partial differential equation (PDE) models to examine the homogeneous and heterogeneous environments associated with cholera transmission. |
|
| Meszaros et al., 2020 [84] | Proposed a mathematical model for cholera incorporating transmission within and between households. |
|
| Abrams et al., 2013 [85] | Developed three cholera surveillance models to forecast the expected number of cases in Haiti during the 2010–2011 cholera epidemic. |
|
| Torres et al., 2018 [82] | Proposed and analyzed a SITRV (susceptible-infectious-treated-recovered-vaccinated) type model for cholera. |
|
| Che et al., 2020 [86] | Used a “fitted” demographic equation (i.e., disease-free equation) to capture total population and a fitted low-high risk structured cholera differential equation model to study reported cholera cases in Cameroon 1987–2004. |
|
| Dangbé et al., 2018 [87] | Proposed a model considering climatic factors and human behavior on the spread of cholera |
|
| Baracchini et al., 2017 [56] | Proposed a stochastic, rainfall–temperature driven model to examine the seasonality of cholera in Bangladesh. |
|
| Koepke et al., 2016 [97] | Proposed a predictive ‘susceptible-infected-recovered-susceptible’ (SIRS) type model in the form of continuous-time hidden Markov states to estimate the contribution of water depth and water temperature on the spread of cholera. |
|
| Perez-Saez et al., 2017 [58] | Proposed a probabilistic spatial model to investigate the role human mobility plays in cholera transmission. |
|
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Oprea, M.; Njamkepo, E.; Cristea, D.; Zhukova, A.; Clark, C.G.; Kravetz, A.N.; Monakhova, E.; Ciontea, A.S.; Cojocaru, R.; Rauzier, J.; et al. The seventh pandemic of cholera in Europe revisited by microbial genomics. Nat. Commun. 2020, 11, 5347. [Google Scholar] [CrossRef] [PubMed]
- Colwell, R.R. Global Climate and Infectious Disease: The Cholera Paradigm. Science 1996, 274, 2025–2031. [Google Scholar] [CrossRef] [Green Version]
- Cholera. Available online: https://www.who.int/news-room/fact-sheets/detail/cholera (accessed on 29 June 2021).
- Ali, M.; Nelson, A.R.; Lopez, A.L.; Sack, D.A. Updated global burden of cholera in endemic countries. PLoS Negl. Trop. Dis. 2015, 9, e0003832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jutla, A.S.; Akanda, A.S.; Islam, S. Tracking Cholera in Coastal Regions using Satellite Observations. J. Am. Water Resour. Assoc. 2010, 46, 651–662. [Google Scholar] [CrossRef] [Green Version]
- WHO; UNICEF. Progress on Sanitation and Drinking Water: 2010 Update; WHO: Geneva, Switzerland; UNICEF: New York, NY, USA, 2010; Available online: https://www.who.int/water_sanitation_he (accessed on 5 June 2021).
- Griffith, D.C.; Kelly-Hope, L.A.; Miller, M.A. Review of reported cholera outbreaks worldwide, 1995–2005. Am. J. Trop. Med. Hyg. 2006, 75, 973–977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huq, A.; Colwell, R. Vibrios in the marine and estuarine environment: Tracking Vibrio cholerae. Ecosyst. Health 1996, 2, 198–214. [Google Scholar]
- Khan, R.; Anwar, R.; Akanda, S.; McDonald, M.D.; Huq, A.; Jutla, A.; Colwell, R. Assessment of Risk of Cholera in Haiti following Hurricane Matthew. Am. J. Trop. Med. Hyg. 2017, 97, 896–903. [Google Scholar] [CrossRef] [PubMed]
- Connolly, M.A.; Heymann, D.L. Deadly comrades: War and infectious diseases. Lancet 2002, 360, s23–s24. [Google Scholar] [CrossRef]
- Federspiel, F.; Ali, M. The cholera outbreak in Yemen: Lessons learned and way forward. BMC Public Health 2018, 18, 1338. [Google Scholar] [CrossRef]
- Republic, S.A. WHO Alliance for the Global Elimination of Trachoma by 2020: Progress report on elimination of trachoma, 2014–2016. Relev. Epidemiol. Hebd. 2017, 92, 359–368. [Google Scholar]
- Camacho, A.; Bouhenia, M.; Alyusfi, R.; Alkohlani, A.; Naji, M.A.M.; de Radiguès, X.; Abubakar, A.M.; Almoalmi, A.; Seguin, C.; Sagrado, M.J.; et al. Cholera epidemic in Yemen, 2016–2018: An analysis of surveillance data. Lancet. Glob. Health 2018, 6, e680–e690. [Google Scholar] [CrossRef]
- Jutla, A.; Aldaach, H.; Billian, H.; Akanda, A.; Huq, A.; Colwell, R. Satellite Based Assessment of Hydroclimatic Conditions Related to Cholera in Zimbabwe. PLoS ONE 2015, 10, e0137828. [Google Scholar] [CrossRef] [Green Version]
- Jutla, A.; Whitcombe, E.; Hasan, N.; Haley, B.; Akanda, A.; Huq, A.; Alam, M.; Sack, R.B.; Colwell, R. Environmental factors influencing epidemic cholera. Am. J. Trop. Med. Hyg. 2013, 89, 597–607. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.; Hasan, N.A.; Sadique, A.; Bhuiyan, N.A.; Ahmed, K.U.; Nusrin, S.; Nair, G.B.; Siddique, A.K.; Sack, R.B.; Sack, D.A.; et al. Seasonal cholera caused by Vibrio cholerae serogroups O1 and O139 in the coastal aquatic environment of Bangladesh. Appl. Environ. Microbiol. 2006, 72, 4096–4104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alam, M.; Islam, A.; Bhuiyan, N.A.; Rahim, N.; Hossain, A.; Khan, G.Y.; Ahmed, D.; Watanabe, H.; Izumiya, H.; Faruque, A.S.G.; et al. Clonal transmission, dual peak, and off-season cholera in Bangladesh. Infect. Ecol. Epidemiol. 2011, 1. [Google Scholar] [CrossRef] [Green Version]
- Codeço, C.T. Endemic and epidemic dynamics of cholera: The role of the aquatic reservoir. BMC Infect. Dis. 2001, 1, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huq, A.; Small, E.B.; West, P.A.; Huq, M.I.; Rahman, R.; Colwell, R.R. Ecological relationships between Vibrio cholerae and planktonic crustacean copepods. Appl. Environ. Microbiol. 1983, 45, 275–283. [Google Scholar] [CrossRef] [Green Version]
- Almagro-Moreno, S.; Taylor, R.K. Cholera: Environmental Reservoirs and Impact on Disease Transmission. Microbiol. Spectr. 2013, 1. [Google Scholar] [CrossRef] [Green Version]
- Vezzulli, L.; Grande, C.; Reid, P.C.; Hélaouët, P.; Edwards, M.; Höfle, M.G.; Brettar, I.; Colwell, R.R.; Pruzzo, C. Climate influence on Vibrio and associated human diseases during the past half-century in the coastal North Atlantic. Proc. Natl. Acad. Sci. USA 2016, 113, E5062–E5071. [Google Scholar] [CrossRef] [Green Version]
- Vezzulli, L.; Pezzati, E.; Brettar, I.; Höfle, M.; Pruzzo, C. Effects of Global Warming on Vibrio Ecology. Microbiol. Spectr. 2015, 3. [Google Scholar] [CrossRef] [Green Version]
- Constantin de Magny, G.; Murtugudde, R.; Sapiano, M.R.P.; Nizam, A.; Brown, C.W.; Busalacchi, A.J.; Yunus, M.; Nair, G.B.; Gil, A.I.; Lanata, C.F.; et al. Environmental signatures associated with cholera epidemics. Proc. Natl. Acad. Sci. USA 2008, 105, 17676–17681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pollitzer, R. Cholera studies. 1. History of the disease. Bull. World Health Organ. 1954, 10, 421–461. [Google Scholar] [PubMed]
- Hasan, J.A.; Bernstein, D.; Huq, A.; Loomis, L.; Tamplin, M.L.; Colwell, R.R. Cholera DFA: An improved direct fluorescent monoclonal antibody staining kit for rapid detection and enumeration of Vibrio cholerae O1. FEMS Microbiol. Lett. 1994, 120, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Lowenhaupt, E.; Huq, A.; Colwell, R.R.; Adingra, A.; Epstein, P.R. Rapid detection of Vibrio cholerae O1 in west Africa. Lancet 1998, 351, 34. [Google Scholar] [CrossRef]
- Fykse, E.M.; Skogan, G.; Davies, W.; Olsen, J.S.; Blatny, J.M. Detection of Vibrio cholerae by real-time nucleic acid sequence-based amplification. Appl. Environ. Microbiol. 2007, 73, 1457–1466. [Google Scholar] [CrossRef] [Green Version]
- Nandi, B.; Nandy, R.K.; Mukhopadhyay, S.; Nair, G.B.; Shimada, T.; Ghose, A.C. Rapid method for species-specific identification of Vibrio cholerae using primers targeted to the gene of outer membrane protein OmpW. J. Clin. Microbiol. 2000, 38, 4145–4151. [Google Scholar] [CrossRef] [Green Version]
- Bauer, A.; Rørvik, L.M. A novel multiplex PCR for the identification of Vibrio vulnificus, Vibrio cholerae and Vibrio vulnificus. Lett. Appl. Microbiol. 2007, 45, 371–375. [Google Scholar] [CrossRef]
- Hoshino, K.; Yamasaki, S.; Mukhopadhyay, A.K.; Chakraborty, S.; Basu, A.; Bhattacharya, S.K.; Nair, G.B.; Shimada, T.; Takeda, Y. Development and evaluation of a multiplex PCR assay for rapid detection of toxigenic Vibrio cholerae O1 and O139. FEMS Immunol. Med. Microbiol. 1998, 20, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Brumfield, K.D.; Carignan, B.M.; Ray, J.N.; Jumpre, P.E.; Son, M.S. Laboratory Techniques Used to Maintain and Differentiate Biotypes of Vibrio cholerae Clinical and Environmental Isolates. J. Vis. Exp. 2017. [Google Scholar] [CrossRef]
- Thompson, F.L.; Swings, J. Taxonomy of the Vibrios. In The Biology of Vibrios; ASM Press: Washington, DC, USA, 2014; pp. 27–43. [Google Scholar]
- Colwell, R.R.; Grimes, D.J. Nonculturable Microorganisms in the Environment; Springer: Boston, MA, USA, 2000; ISBN 978-1-4757-0273-6. [Google Scholar] [CrossRef]
- Oliver, J.D. Recent findings on the viable but nonculturable state in pathogenic bacteria. FEMS Microbiol. Rev. 2010, 34, 415–425. [Google Scholar] [CrossRef] [Green Version]
- Colwell, R.R. Viable but nonculturable bacteria: A survival strategy. J. Infect. Chemother. 2000, 6, 121–125. [Google Scholar] [CrossRef]
- Colwell, R.R.; Brayton, P.; Herrington, D.; Tall, B.; Huq, A.; Levine, M.M. Viable but non-culturable Vibrio cholerae O1 revert to a cultivable state in the human intestine. World J. Microbiol. Biotechnol. 1996, 12, 28–31. [Google Scholar] [CrossRef]
- Senoh, M.; Ghosh-Banerjee, J.; Ramamurthy, T.; Colwell, R.R.; Miyoshi, S.-I.; Nair, G.B.; Takeda, Y. Conversion of viable but nonculturable enteric bacteria to culturable by co-culture with eukaryotic cells. Microbiol. Immunol. 2012, 56, 342–345. [Google Scholar] [CrossRef]
- Alam, M.; Sultana, M.; Nair, G.B.; Siddique, A.K.; Hasan, N.A.; Sack, R.B.; Sack, D.A.; Ahmed, K.U.; Sadique, A.; Watanabe, H.; et al. Viable but nonculturable Vibrio cholerae O1 in biofilms in the aquatic environment and their role in cholera transmission. Proc. Natl. Acad. Sci. USA 2007, 104, 17801–17806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colwell, R.R.; Spira, W.M. Cholera; Barua, D., Greenough, W.B., Eds.; Springer: Boston, MA, USA, 1992; ISBN 978-1-4757-9690-2. [Google Scholar] [CrossRef]
- Huq, A.; Xu, B.; Chowdhury, M.A.; Islam, M.S.; Montilla, R.; Colwell, R.R. A simple filtration method to remove plankton-associated Vibrio cholerae in raw water supplies in developing countries. Appl. Environ. Microbiol. 1996, 62, 2508–2512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colwell, R.R.; Huq, A.; Islam, M.S.; Aziz, K.M.A.; Yunus, M.; Khan, N.H.; Mahmud, A.; Sack, R.B.; Nair, G.B.; Chakraborty, J.; et al. Reduction of cholera in Bangladeshi villages by simple filtration. Proc. Natl. Acad. Sci. USA 2003, 100, 1051–1055. [Google Scholar] [CrossRef] [Green Version]
- Cash, R.A.; Music, S.I.; Libonati, J.P.; Snyder, M.J.; Wenzel, R.P.; Hornick, R.B. Response of Man to Infection with Vibrio cholerae. I. Clinical, Serologic, and Bacteriologic Responses to a Known Inoculum. J. Infect. Dis. 1974, 129, 45–52. [Google Scholar] [CrossRef]
- Silva-Valenzuela, C.A.; Camilli, A. Niche adaptation limits bacteriophage predation of Vibrio cholerae in a nutrient-poor aquatic environment. Proc. Natl. Acad. Sci. USA 2019, 116, 1627–1632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faruque, S.M.; Mekalanos, J.J. Phage-bacterial interactions in the evolution of toxigenic Vibrio cholerae. Virulence 2012, 3, 556–565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vezzulli, L.; Guzmán, C.A.; Colwell, R.R.; Pruzzo, C. Dual role colonization factors connecting Vibrio cholerae’s lifestyles in human and aquatic environments open new perspectives for combating infectious diseases. Curr. Opin. Biotechnol. 2008, 19, 254–259. [Google Scholar] [CrossRef] [PubMed]
- McFall-Ngai, M. Divining the Essence of Symbiosis: Insights from the Squid-Vibrio Model. PLoS Biol. 2014, 12, e1001783. [Google Scholar] [CrossRef] [Green Version]
- Martínez, J.L. Bacterial pathogens: From natural ecosystems to human hosts. Environ. Microbiol. 2013, 15, 325–333. [Google Scholar] [CrossRef]
- Mavian, C.; Paisie, T.K.; Alam, M.T.; Browne, C.; de Rochars, V.M.B.; Nembrini, S.; Cash, M.N.; Nelson, E.J.; Azarian, T.; Ali, A.; et al. Toxigenic Vibrio cholerae evolution and establishment of reservoirs in aquatic ecosystems. Proc. Natl. Acad. Sci. USA 2020, 117, 7897–7904. [Google Scholar] [CrossRef] [Green Version]
- Hasan, N.A.; Choi, S.Y.; Eppinger, M.; Clark, P.W.; Chen, A.; Alam, M.; Haley, B.J.; Taviani, E.; Hine, E.; Su, Q.; et al. Genomic diversity of 2010 Haitian cholera outbreak strains. Proc. Natl. Acad. Sci. USA 2012, 109, E2010–E2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conner, J.G.; Teschler, J.K.; Jones, C.J.; Yildiz, F.H. Staying Alive: Vibrio cholerae’s Cycle of Environmental Survival, Transmission, and Dissemination. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef] [Green Version]
- Meibom, K.L.; Blokesch, M.; Dolganov, N.A.; Wu, C.-Y.; Schoolnik, G.K. Chitin induces natural competence in Vibrio cholerae. Science 2005, 310, 1824–1827. [Google Scholar] [CrossRef] [PubMed]
- Ceccarelli, D.; Chen, A.; Hasan, N.A.; Rashed, S.M.; Huq, A.; Colwell, R.R. Non-O1/non-O139 Vibrio cholerae carrying multiple virulence factors and V. cholerae O1 in the Chesapeake Bay, Maryland. Appl. Environ. Microbiol. 2015, 81, 1909–1918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carignan, B.M.; Brumfield, K.D.; Son, M.S. Single Nucleotide Polymorphisms in Regulator-Encoding Genes Have an Additive Effect on Virulence Gene Expression in a Vibrio cholerae Clinical Isolate. Msphere 2016, 1, e00253-16. [Google Scholar] [CrossRef] [Green Version]
- Blokesch, M.; Schoolnik, G.K. Serogroup Conversion of Vibrio cholerae in Aquatic Reservoirs. PLoS Pathog. 2007, 3, e81. [Google Scholar] [CrossRef] [PubMed]
- Colwell, R.R.; Huq, A.; Chowdhury, M.A.; Brayton, P.R.; Xu, B. Serogroup conversion of Vibrio cholerae. Can. J. Microbiol. 1995, 41, 946–950. [Google Scholar] [CrossRef] [PubMed]
- Baracchini, T.; King, A.A.; Bouma, M.J.; Rodó, X.; Bertuzzo, E.; Pascual, M. Seasonality in cholera dynamics: A rainfall-driven model explains the wide range of patterns in endemic areas. Adv. Water Resour. 2017, 108, 357–366. [Google Scholar] [CrossRef] [Green Version]
- Martinez, P.P.; Reiner, R.C.; Cash, B.A.; Rodó, X.; Shahjahan Mondal, M.; Roy, M.; Yunus, M.; Faruque, A.S.G.; Huq, S.; King, A.A.; et al. Cholera forecast for Dhaka, Bangladesh, with the 2015–2016 El Niño: Lessons learned. PLoS ONE 2017, 12, e0172355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez-Saez, J.; King, A.A.; Rinaldo, A.; Yunus, M.; Faruque, A.S.G.; Pascual, M. Climate-driven endemic cholera is modulated by human mobility in a megacity. Adv. Water Resour. 2017, 108, 367–376. [Google Scholar] [CrossRef]
- Deen, J.; Mengel, M.A.; Clemens, J.D. Epidemiology of cholera. Vaccine 2020, 38 (Suppl. 1), A31–A40. [Google Scholar] [CrossRef]
- Glass, R.I.; Becker, S.; Huq, M.I.; Stoll, B.J.; Khan, M.U.; Merson, M.H.; Lee, J.V.; Black, R.E. Endemic cholera in rural Bangladesh, 1966–1980. Am. J. Epidemiol. 1982, 116, 959–970. [Google Scholar] [CrossRef]
- Shears, P. Recent developments in cholera. Curr. Opin. Infect. Dis. 2001, 14, 553–558. [Google Scholar] [CrossRef] [PubMed]
- Akanda, A.S.; Jutla, A.S.; Islam, S. Dual peak cholera transmission in Bengal Delta: A hydroclimatological explanation. Geophys. Res. Lett. 2009, 36, L19401. [Google Scholar] [CrossRef]
- Lattos, A.; Bitchava, K.; Giantsis, I.A.; Theodorou, J.A.; Batargias, C.; Michaelidis, B. The implication of Vibrio bacteria in the winter mortalities of the critically endangered pinna nobilis. Microorganisms 2021, 9, 922. [Google Scholar] [CrossRef] [PubMed]
- Baker-Austin, C.; Stockley, L.; Rangdale, R.; Martinez-Urtaza, J. Environmental occurrence and clinical impact of Vibrio vulnificus and Vibrio vulnificus: A European perspective. Environ. Microbiol. Rep. 2010, 2, 7–18. [Google Scholar] [CrossRef]
- Ceccarelli, D.; Colwell, R.R. Vibrio ecology, pathogenesis, and evolution. Front. Microbiol. 2014, 5. [Google Scholar] [CrossRef]
- Thompson, J.R.; Polz, M.F. Dynamics of Vibrio Populations and Their Role in Environmental Nutrient Cycling. In The Biology of Vibrios; ASM Press: Washington, DC, USA, 2014; pp. 190–203. [Google Scholar] [CrossRef]
- Lobitz, B.; Beck, L.; Huq, A.; Wood, B.; Fuchs, G.; Faruque, A.S.G.; Colwell, R. Climate and infectious disease: Use of remote sensing for detection of Vibrio cholerae by indirect measurement. Proc. Natl. Acad. Sci. USA 2000, 97, 1438–1443. [Google Scholar] [CrossRef] [Green Version]
- Emch, M.; Feldacker, C.; Yunus, M.; Streatfield, P.K.; DinhThiem, V.; Canh, D.G.; Ali, M. Local environmental predictors of cholera in Bangladesh and Vietnam. Am. J. Trop. Med. Hyg. 2008, 78, 823–832. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Cao, C.X.; Wang, D.C.; Kan, B.; Xu, Y.F.; Ni, X.L.; Zhu, Z.C. Environmental factor analysis of cholera in China using remote sensing and geographical information systems. Epidemiol. Infect. 2016, 144, 940–951. [Google Scholar] [CrossRef] [Green Version]
- Jutla, A.S.; Akanda, A.S.; Islam, S. A framework for predicting endemic cholera using satellite derived environmental determinants. Environ. Model. Softw. 2013, 47, 148–158. [Google Scholar] [CrossRef]
- Kirpich, A.; Weppelmann, T.A.; Yang, Y.; Ali, A.; Morris, J.G.; Longini, I.M. Cholera Transmission in Ouest Department of Haiti: Dynamic Modeling and the Future of the Epidemic. PLoS Negl. Trop. Dis. 2015, 9, e0004153. [Google Scholar] [CrossRef] [PubMed]
- Eisenberg, M.C.; Kujbida, G.; Tuite, A.R.; Fisman, D.N.; Tien, J.H. Examining rainfall and cholera dynamics in Haiti using statistical and dynamic modeling approaches. Epidemics 2013, 5, 197–207. [Google Scholar] [CrossRef] [Green Version]
- Jutla, A.; Akanda, A.; Unnikrishnan, A.; Huq, A.; Colwell, R. Predictive Time Series Analysis Linking Bengal Cholera with Terrestrial Water Storage Measured from Gravity Recovery and Climate Experiment Sensors. Am. J. Trop. Med. Hyg. 2015, 93, 1179–1186. [Google Scholar] [CrossRef] [PubMed]
- Huq, A.; West, P.A.; Small, E.B.; Huq, M.I.; Colwell, R.R. Influence of water temperature, salinity, and pH on survival and growth of toxigenic Vibrio cholerae serovar 01 associated with live copepods in laboratory microcosms. Appl. Environ. Microbiol. 1984, 48, 420–424. [Google Scholar] [CrossRef] [Green Version]
- Singleton, F.L.; Attwell, R.; Jangi, S.; Colwell, R.R. Effects of temperature and salinity on Vibrio cholerae growth. Appl. Environ. Microbiol. 1982, 44, 1047–1058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, R.; Nguyen, T.H.; Shisler, J.; Lin, L.-S.; Jutla, A.; Colwell, R.R. Evaluation of Risk of Cholera after a Natural Disaster: Lessons Learned from the 2015 Nepal Earthquake. J. Water Resour. Plan. Manag. 2018, 144, 04018044. [Google Scholar] [CrossRef]
- Heesterbeek, H.; Anderson, R.M.; Andreasen, V.; Bansal, S.; De Angelis, D.; Dye, C.; Eames, K.T.D.; Edmunds, W.J.; Frost, S.D.W.; Funk, S.; et al. Modeling infectious disease dynamics in the complex landscape of global health. Science 2015, 347, aaa4339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chowell, G.; Sattenspiel, L.; Bansal, S.; Viboud, C. Mathematical models to characterize early epidemic growth: A review. Phys. Life Rev. 2016, 18, 66–97. [Google Scholar] [CrossRef] [Green Version]
- Andrews, J.R.; Basu, S. Transmission dynamics and control of cholera in Haiti: An epidemic model. Lancet 2011, 377, 1248–1255. [Google Scholar] [CrossRef] [Green Version]
- Grad, Y.H.; Miller, J.C.; Lipsitch, M. Cholera modeling: Challenges to quantitative analysis and predicting the impact of interventions. Epidemiology 2012, 23, 523–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brauer, F. Compartmental Models in Epidemiology BT—Mathematical Epidemiology; Brauer, F., van den Driessche, P., Wu, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 19–79. ISBN 978-3-540-78911-6. [Google Scholar] [CrossRef]
- Lemos-Paião, A.P.; Silva, C.J.; Torres, D.F.M. A cholera mathematical model with vaccination and the biggest outbreak of world’s history. AIMS Math. 2019, 3, 448–463. [Google Scholar] [CrossRef]
- Wang, X.; Gao, D.; Wang, J. Influence of human behavior on cholera dynamics. Math. Biosci. 2015, 267, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Meszaros, V.A.; Miller-Dickson, M.D.; Baffour-Awuah, F.; Almagro-Moreno, S.; Ogbunugafor, C.B. Direct transmission via households informs models of disease and intervention dynamics in cholera. PLoS ONE 2020, 15, e0229837. [Google Scholar] [CrossRef] [Green Version]
- Abrams, J.Y.; Copeland, J.R.; Tauxe, R.V.; Date, K.A.; Belay, E.D.; Mody, R.K.; Mintz, E.D. Real-time modelling used for outbreak management during a cholera epidemic, Haiti, 2010–2011. Epidemiol. Infect. 2013, 141, 1276–1285. [Google Scholar] [CrossRef] [Green Version]
- Che, E.N.; Kang, Y.; Yakubu, A.-A. Risk structured model of cholera infections in Cameroon. Math. Biosci. 2020, 320, 108303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dangbé, E.; Irépran, D.; Perasso, A.; Békollé, D. Mathematical modelling and numerical simulations of the influence of hygiene and seasons on the spread of cholera. Math. Biosci. 2018, 296, 60–70. [Google Scholar] [CrossRef]
- Glass, R.I.; Svennerholm, A.M.; Khan, M.R.; Huda, S.; Imdadul Huq, M.; Holmgren, J. Seroepidemiological studies of EI tor cholera in bangladesh: Association of serum antibody levels with protection. J. Infect. Dis. 1985, 151, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Chao, D.L.; Longini, I.M.; Morris, J.G. Modeling Cholera Outbreaks. In Cholera Outbreaks; Nair, G.B., Takeda, Y., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 195–209. ISBN 978-3-642-55404-9. [Google Scholar] [CrossRef] [Green Version]
- Ritter, A.S.; Chowdhury, F.; Franke, M.F.; Becker, R.L.; Bhuiyan, T.R.; Khan, A.I.; Saha, N.C.; Ryan, E.T.; Calderwood, S.B.; LaRocque, R.C.; et al. Vibriocidal titer and protection from cholera in children. Open Forum Infect. Dis. 2019, 6. [Google Scholar] [CrossRef]
- Faruque, A.S.G.; Malek, M.A.; Khan, A.I.; Huq, S.; Salam, M.A.; Sack, D.A. Diarrhoea in elderly people: Aetiology, and clinical characteristics. Scand. J. Infect. Dis. 2004, 36, 204–208. [Google Scholar] [CrossRef] [PubMed]
- Cassiers, T.; Kesteloot, C. Socio-spatial Inequalities and Social Cohesion in European Cities. Urban. Stud. 2012, 49, 1909–1924. [Google Scholar] [CrossRef]
- Najib, K. Socio-spatial inequalities and dynamics of rich and poor enclaves in three French cities: A policy of social mixing under test. Popul. Space Place 2020, 26. [Google Scholar] [CrossRef]
- Gerometta, J.; Haussermann, H.; Longo, G. Social Innovation and Civil Society in Urban Governance: Strategies for an Inclusive City. Urban. Stud. 2005, 42, 2007–2021. [Google Scholar] [CrossRef]
- Al-Arydah, M.; Mwasa, A.; Tchuenche, J.M.; Smith, R.J. Modeling cholera disease with education and chlorination. J. Biol. Syst. 2013, 21, 1340007. [Google Scholar] [CrossRef] [Green Version]
- Einarsdóttir, J.; Passa, A.; Gunnlaugsson, G. Health education and cholera in rural Guinea-Bissau. Int. J. Infect. Dis. 2001, 5, 133–138. [Google Scholar] [CrossRef] [Green Version]
- Koepke, A.A.; Longini, I.M., Jr.; Halloran, M.E.; Wakefield, J.; Minin, V.N. Predictive modeling of cholera outbreaks in Bangladesh. Ann. Appl. Stat. 2016, 10. [Google Scholar] [CrossRef] [PubMed]
- Kulinkina, A.V.; Mohan, V.R.; Francis, M.R.; Kattula, D.; Sarkar, R.; Plummer, J.D.; Ward, H.; Kang, G.; Balraj, V.; Naumova, E.N. Seasonality of water quality and diarrheal disease counts in urban and rural settings in south India. Sci. Rep. 2016, 6, 20521. [Google Scholar] [CrossRef] [Green Version]
- Altizer, S.; Dobson, A.; Hosseini, P.; Hudson, P.; Pascual, M.; Rohani, P. Seasonality and the dynamics of infectious diseases. Ecol. Lett. 2006, 9, 467–484. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Moreno, D.; Pascual, M.; Bouma, M.; Dobson, A.; Cash, B. Cholera Seasonality in Madras (1901–1940): Dual Role for Rainfall in Endemic and Epidemic Regions. Ecohealth 2007, 4, 52–62. [Google Scholar] [CrossRef]
- Nishiura, H.; Tsuzuki, S.; Yuan, B.; Yamaguchi, T.; Asai, Y. Transmission dynamics of cholera in Yemen, 2017: A real time forecasting. Theor. Biol. Med. Model. 2017, 14, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Predicting Cholera Risk in Yemen. Available online: https://earthobservatory.nasa.gov/images/147101/predicting-cholera-risk-in-yemen (accessed on 5 June 2021).
- van den Driessche, P. Reproduction numbers of infectious disease models. Infect. Dis. Model. 2017, 2, 288–303. [Google Scholar] [CrossRef] [PubMed]
- Aguilar, E.; Peterson, T.C.; Obando, P.R.; Frutos, R.; Retana, J.A.; Solera, M.; Soley, J.; García, I.G.; Araujo, R.M.; Santos, A.R.; et al. Changes in precipitation and temperature extremes in Central America and northern South America, 1961–2003. J. Geophys. Res. Atmos. 2005, 110, 1–15. [Google Scholar] [CrossRef]
- El-Sayed, A.; Kamel, M. Climatic changes and their role in emergence and re-emergence of diseases. Environ. Sci. Pollut. Res. 2020, 27, 22336–22352. [Google Scholar] [CrossRef]
- AR4 Climate Change 2007: The Physical Science Basis—IPCC. Available online: https://www.ipcc.ch/report/ar4/wg1/ (accessed on 29 June 2021).
- Baker-Austin, C.; Trinanes, J.; Gonzalez-Escalona, N.; Martinez-Urtaza, J. Non-Cholera Vibrios: The Microbial Barometer of Climate Change. Trends Microbiol. 2017, 25, 76–84. [Google Scholar] [CrossRef] [Green Version]
- Baker-Austin, C.; Trinanes, J.A.; Taylor, N.G.H.; Hartnell, R.; Siitonen, A.; Martinez-Urtaza, J. Emerging Vibrio risk at high latitudes in response to ocean warming. Nat. Clim. Chang. 2013, 3, 73–77. [Google Scholar] [CrossRef]
- Vezzulli, L.; Baker-Austin, C.; Kirschner, A.; Pruzzo, C.; Martinez-Urtaza, J. Global emergence of environmental non-O1/O139 Vibrio cholerae infections linked with climate change: A neglected research field? Environ. Microbiol. 2020, 22, 4342–4355. [Google Scholar] [CrossRef]
- Anderson, P.K.; Cunningham, A.A.; Patel, N.G.; Morales, F.J.; Epstein, P.R.; Daszak, P. Emerging infectious diseases of plants: Pathogen pollution, climate change and agrotechnology drivers. Trends Ecol. Evol. 2004, 19, 535–544. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Usmani, M.; Brumfield, K.D.; Jamal, Y.; Huq, A.; Colwell, R.R.; Jutla, A. A Review of the Environmental Trigger and Transmission Components for Prediction of Cholera. Trop. Med. Infect. Dis. 2021, 6, 147. https://doi.org/10.3390/tropicalmed6030147
Usmani M, Brumfield KD, Jamal Y, Huq A, Colwell RR, Jutla A. A Review of the Environmental Trigger and Transmission Components for Prediction of Cholera. Tropical Medicine and Infectious Disease. 2021; 6(3):147. https://doi.org/10.3390/tropicalmed6030147
Chicago/Turabian StyleUsmani, Moiz, Kyle D. Brumfield, Yusuf Jamal, Anwar Huq, Rita R. Colwell, and Antarpreet Jutla. 2021. "A Review of the Environmental Trigger and Transmission Components for Prediction of Cholera" Tropical Medicine and Infectious Disease 6, no. 3: 147. https://doi.org/10.3390/tropicalmed6030147
APA StyleUsmani, M., Brumfield, K. D., Jamal, Y., Huq, A., Colwell, R. R., & Jutla, A. (2021). A Review of the Environmental Trigger and Transmission Components for Prediction of Cholera. Tropical Medicine and Infectious Disease, 6(3), 147. https://doi.org/10.3390/tropicalmed6030147