Strain-Dependent Activity of Zika Virus and Exposure History in Serological Diagnostics

 ,
,  , , ,
, , ,
Abstract
1. Introduction
2. Results
2.1. RT-PCR Sensitivity is Strain Dependent
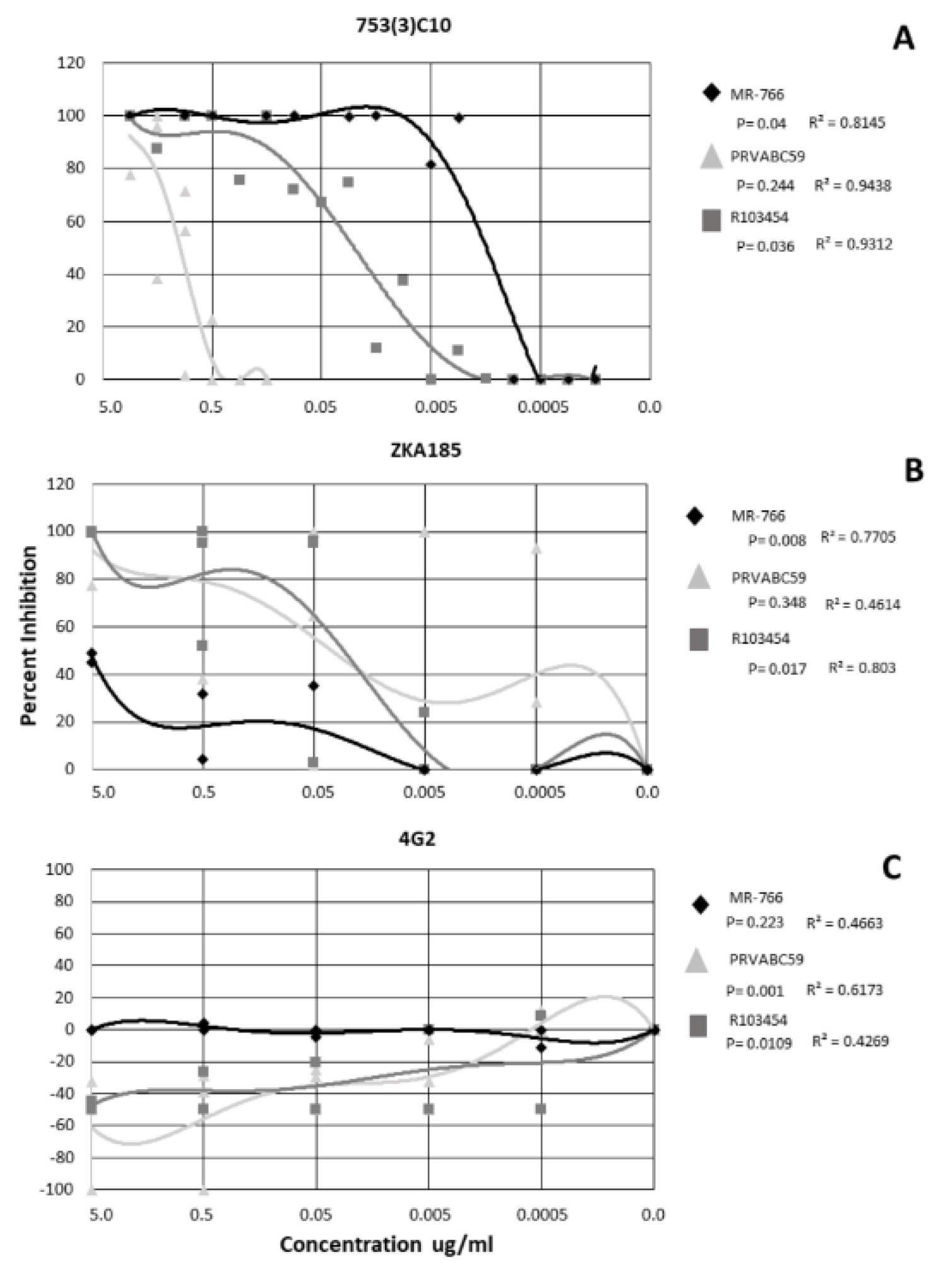
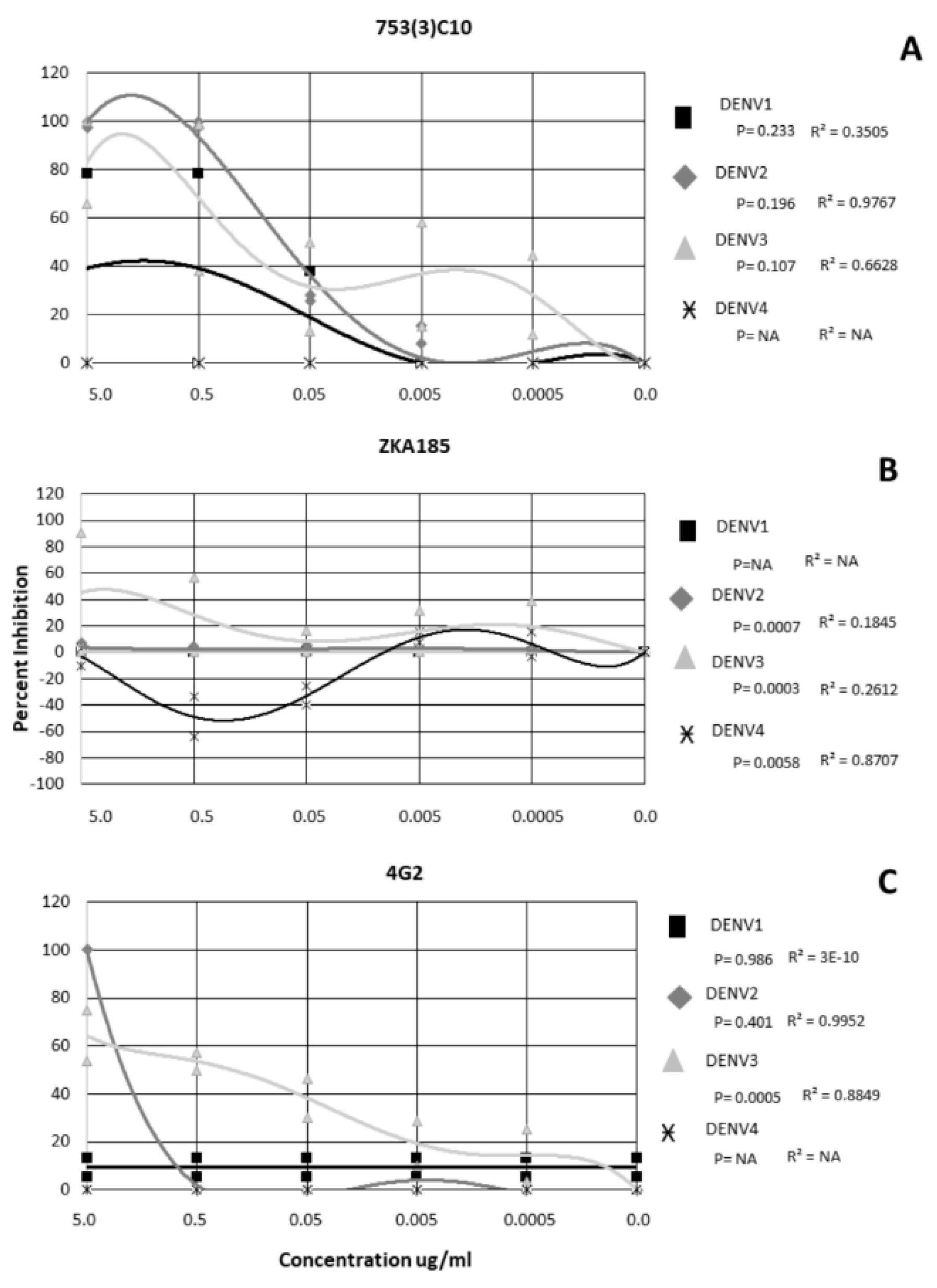
2.2. Monoclonal Antibody Neutralization of ZIKV and DENV is Strain/Serotype Dependent
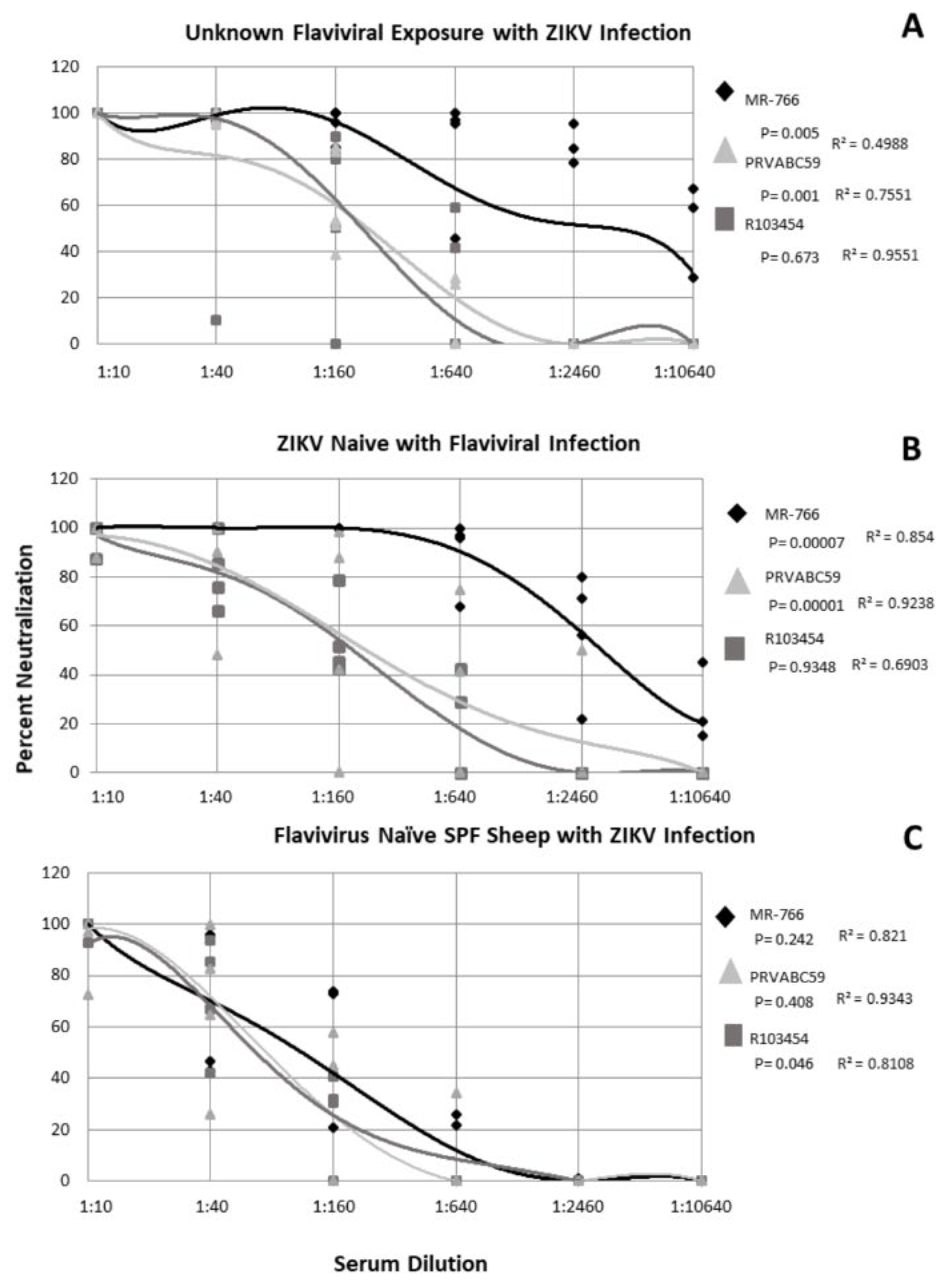
2.3. Subjects with Confirmed ZIKV Exposure with Unknown Flaviviral Exposure History Exhibit Strain-Specific Neutralization of ZIKV
2.4. ZIKV-Naïve Subjects with Flaviviral Exposure History Neutralize ZIKV in a Strain Dependent Manner
2.5. Flavivirus Naïve SPF Sheep with ZIKV Only Exposure Exhibit Similar Neutralization of ZIKV Strains
2.6. Subjects with Confirmed ZIKV Exposure Exhibit Cross-Neutralization of Other Flaviviruses
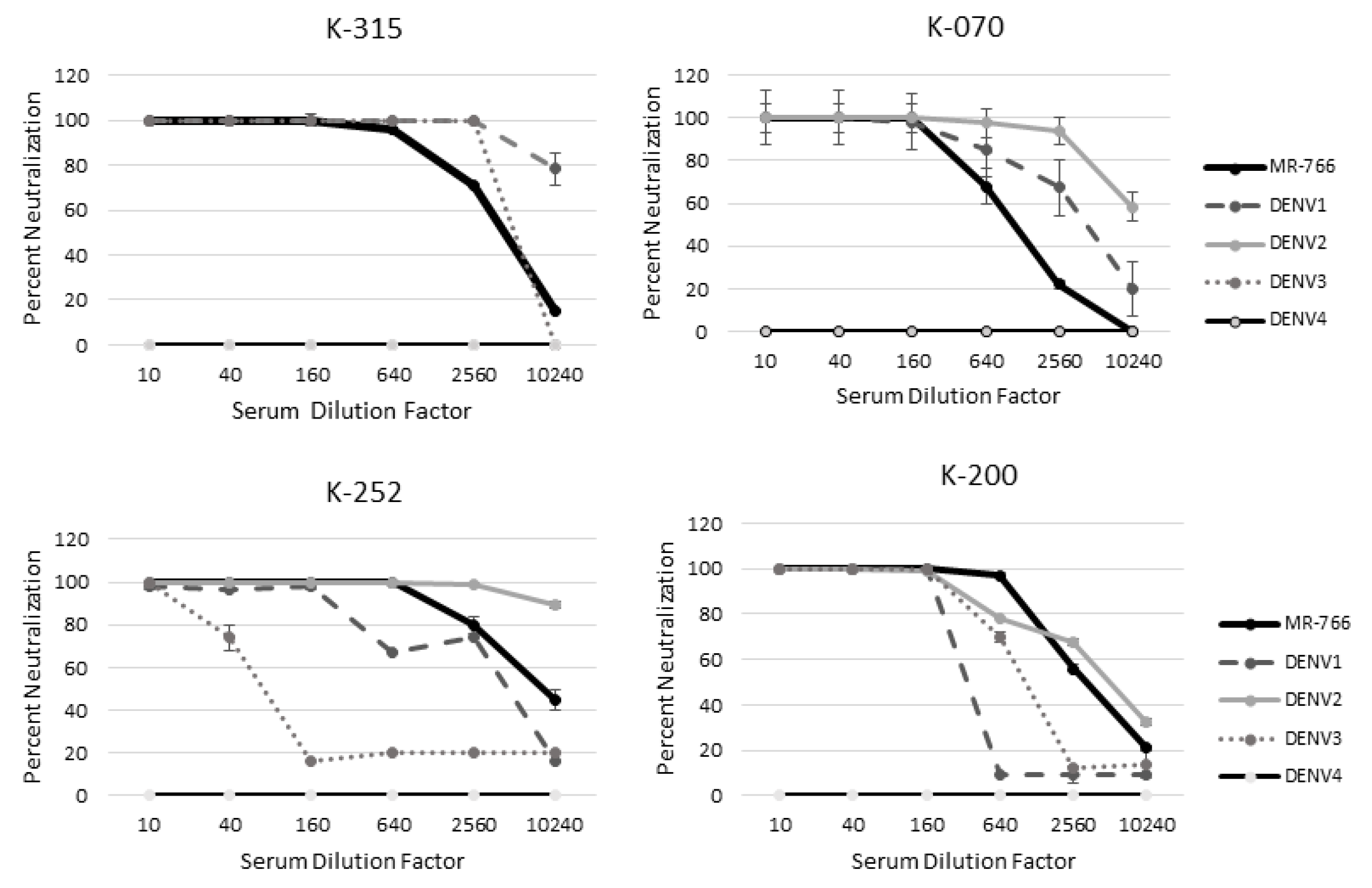
2.7. ZIKV-Naïve Subjects with Previous Flavivirus Exposure Effectively Neutralize ZIKV and Other Flaviviruses
2.8. MR−766 Is Neutralized More Than DENV by ZIKV-Naïve Patient Serum
3. Discussion
4. Methods
4.1. Viruses
4.2. Monoclonal Antibodies
4.3. Subject Specimens
4.4. Serologic and Molecular Assays
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Speer, S.; Pierson, T. Diagnostics for Zika virus on the horizon. Science 2016, 353, 750–751. [Google Scholar] [CrossRef]
- Waggoner, J.J.; Pinsky, B.A. Zika Virus: Diagnostics for an Emerging Pandemic Threat. J. Clin. Microbiol. 2016, 54, 860–867. [Google Scholar] [CrossRef]
- Gorchakov, R.; Berry, R.M.; Patel, S.M.; El Sahly, H.M.; Ronca, S.E.; Murray, K.O. Optimizing PCR Detection of Zika Virus from Various Body Fluids. Am. J. Trop. Med. Hyg. 2019, 100, 427–433. [Google Scholar] [CrossRef]
- Petersen, L.R.; Jamieson, D.J.; Powers, A.M.; Honein, M.A. Zika Virus. N. Engl. J. Med. 2016, 374, 1552–1563. [Google Scholar] [CrossRef]
- Musso, D.; Rouault, E.; Teissier, A.; Lanteri, M.C.; Zisou, K.; Broult, J.; Grange, E.; Nhan, T.X.; Aubry, M. Molecular detection of Zika virus in blood and RNA load determination during the French Polynesian outbreak. J. Med. Virol. 2017, 89, 1505–1510. [Google Scholar] [CrossRef] [PubMed]
- Lanciotti, R.S.; Kosoy, O.L.; Laven, J.J.; Velez, J.O.; Lambert, A.J.; Johnson, A.J.; Stanfield, S.M.; Duffy, M.R. Genetic and serologic properties of Zika virus associated with an epidemic, Yap State, Micronesia, 2007. Emerg. Infect. Dis. 2008, 14, 1232–1239. [Google Scholar] [CrossRef] [PubMed]
- Oduyebo, T.; Polen, K.D.; Walke, H.T.; Reagan-Steiner, S.; Lathrop, E.; Rabe, I.B.; Kuhnert-Tallman, W.L.; Martin, S.W.; Walker, A.T.; Gregory, C.J. Update: Interim Guidance for Health Care Providers Caring for Pregnant Women with Possible Zika Virus Exposure—United States (Including U.S. Territories), July 2017. MMWR Morb. Mortal. Wkly. Rep. 2017, 66, 781–793. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. Arboviral Diseases, Neuroinvasive and Non-neuroinvasive 2015 Case Definition Atlanta, Georgia: Centers for Disease Control and Prevention. 2015. Available online: https://wwwn.cdc.gov/nndss/conditions/arboviral-diseases-neuroinvasive-and-non-neuroinvasive/case-definition/2015/ (accessed on 24 February 2020).
- Centers for Disease Control and Prevention. Zika MAC-ELISA: Instructions for Use; Centers for Disease Control and Prevention: Atlanta, GA, USA, 2018.
- Chao, D.Y.; Whitney, M.T.; Davis, B.S.; Medina, F.A.; Munoz, J.L.; Chang, G.J. Comprehensive evaluation of differential serodiagnosis between Zika and dengue viral infection. J. Clin. Microbiol. 2018, 57, e01506–e01518. [Google Scholar] [CrossRef]
- Jaaskelainen, A.J.; Korhonen, E.M.; Huhtamo, E.; Lappalainen, M.; Vapalahti, O.; Kallio-Kokko, H. Validation of serological and molecular methods for diagnosis of zika virus infections. J. Virol. Methods 2019, 263, 68–74. [Google Scholar] [CrossRef]
- Calisher, C.H.; Karabatsos, N.; Dalrymple, J.M.; Shope, R.E.; Porterfield, J.S.; Westaway, E.G.; Brandt, W.E. Antigenic Relationships between Flaviviruses as Determined by Cross-neutralization Tests with Polyclonal Antisera. J. Gen. Virol. 1989, 70, 37–43. [Google Scholar] [CrossRef]
- Tsai, W.Y.; Durbin, A.; Tsai, J.J.; Hsieh, S.C.; Whitehead, S.; Wang, W.K. Complexity of Neutralizing Antibodies against Multiple Dengue Virus Serotypes after Heterotypic Immunization and Secondary Infection Revealed by In-Depth Analysis of Cross-Reactive Antibodies. J. Virol. 2015, 89, 7348–7362. [Google Scholar] [CrossRef] [PubMed]
- Patel, B.; Longo, P.; Miley, M.J.; Montoya, M.; Harris, E.; de Silva, A.M. Dissecting the human serum antibody response to secondary dengue virus infections. PLoS Negl. Trop. Dis. 2017, 11, e0005554. [Google Scholar] [CrossRef] [PubMed]
- Johnson, B.W.; Kosoy, O.; Martin, D.A.; Noga, A.J.; Russell, B.J.; Johnson, A.A.; Petersen, L.R. West Nile virus infection and serologic response among persons previously vaccinated against yellow fever and Japanese encephalitis viruses. Vector Borne Zoonotic Dis. 2005, 5, 137–145. [Google Scholar] [CrossRef]
- Filippone, C.; Marianneau, P.; Murri, S.; Mollard, N.; Avsic-Zupanc, T.; Chinikar, S.; Despres, P.; Caro, V.; Gessain, A.; Berthet, N. Molecular diagnostic and genetic characterization of highly pathogenic viruses: Application during Crimean-Congo haemorrhagic fever virus outbreaks in Eastern Europe and the Middle East. Clin. Microbiol. Infect. 2013, 19, E118–E128. [Google Scholar] [CrossRef] [PubMed]
- Hayes, E.B. Zika virus outside Africa. Emerg. Infect. Dis. 2009, 15, 1347–1350. [Google Scholar] [CrossRef]
- Kuno, G.; Chang, G.J. Full-length sequencing and genomic characterization of Bagaza, Kedougou, and Zika viruses. Arch. Virol. 2007, 152, 687–696. [Google Scholar] [CrossRef]
- Fagbami, A.H. Zika virus infections in Nigeria: Virological and seroepidemiological investigations in Oyo State. J. Hyg. 1979, 83, 213–219. [Google Scholar] [CrossRef]
- Dick, G.W.; Kitchen, S.F.; Haddow, A.J. Zika virus I. Isolations and serological specificity. Trans. R. Soc. Trop. Med. Hyg. 1952, 46, 509–520. [Google Scholar] [CrossRef]
- Lanciotti, R.S.; Lambert, A.J.; Holodniy, M.; Saavedra, S.; Signor, L.C. Phylogeny of Zika Virus in Western Hemisphere, 2015. Emerg. Infect. Dis. 2016, 22, 933–935. [Google Scholar] [CrossRef]
- Bailey, M.J.; Broecker, F.; Freyn, A.W.; Choi, A.; Brown, J.A.; Fedorova, N.; Simon, V.; Lim, J.K.; Evans, M.J.; Garcia-Sastre, A. Human Monoclonal Antibodies Potently Neutralize Zika Virus and Select for Escape Mutations on the Lateral Ridge of the Envelope Protein. J. Virol. 2019, 93. [Google Scholar] [CrossRef]
- Esquivel, R.N.; Patel, A.; Kudchodkar, S.B.; Park, D.H.; Stettler, K.; Beltramello, M.; Allen, J.W.; Mendoza, J.; Ramos, S.; Choi, H. In Vivo Delivery of a DNA-Encoded Monoclonal Antibody Protects Non-human Primates against Zika Virus. Mol. Ther. J. Am. Soc. Gene Ther. 2019, 27, 974–985. [Google Scholar] [CrossRef] [PubMed]
- Frumence, E.; Viranaicken, W.; Bos, S.; Alvarez-Martinez, M.T.; Roche, M.; Arnaud, J.D.; Gadea, G.; Despres, P. A Chimeric Zika Virus between Viral Strains MR766 and BeH819015 Highlights a Role for E-glycan Loop in Antibody-mediated Virus Neutralization. Vaccines 2019, 7, 55. [Google Scholar] [CrossRef] [PubMed]
- Santiago, G.A.; Vazquez, J.; Courtney, S.; Matias, K.Y.; Andersen, L.E.; Colon, C.; Butler, A.E.; Roulo, R.; Bowzard, J.; Villanueva, J.M.; et al. Performance of the Trioplex real-time RT-PCR assay for detection of Zika, dengue, and chikungunya viruses. Nat. Commun. 2018, 9, 1391. [Google Scholar] [CrossRef] [PubMed]
- Diagnostic Test for Zika Virus-Trioplex Real Time RT-PCR Assay Instructions; Centers for Disease Control and Prevention: Atlanta, GA, USA, 2016.
- Emergency Use Authorization-Trioplex Real-time RT-PCR Assay; U.S. Food and Drug Administration: Rockville, Maryland, USA, 2016.
- Mann, B.R.; McMullen, A.R.; Swetnam, D.M.; Salvato, V.; Reyna, M.; Guzman, H.; Bueno, R.; Dennett, J.A.; Tesh, R.B.; Barrett, A.D. Continued evolution of West Nile virus, Houston, Texas, USA, 2002–2012. Emerg. Infect. Dis. 2013, 19, 1418–1427. [Google Scholar] [CrossRef] [PubMed]
- Barr, K.L.; Anderson, B.D.; Heil, G.L.; Friary, J.A.; Gray, G.C.; Focks, D.A. Dengue serotypes 1–4 exhibit unique host specificity in vitro. Virus Adapt. Treat. 2012, 4, 65–73. [Google Scholar] [CrossRef][Green Version]
- Brault, A.C.; Langevin, S.A.; Ramey, W.N.; Fang, Y.; Beasley, D.W.C.; Barker, C.M.; Sanders, T.A.; Reisen, W.K.; Barrett, A.D.T.; Bowen, R.A. Reduced Avian Virulence and Viremia of West Nile Virus Isolates from Mexico and Texas 23. Am. J. Trop. Med. Hyg. 2011, 85, 758–767. [Google Scholar] [CrossRef] [PubMed]
- Balmaseda, A.; Stettler, K.; Medialdea-Carrera, R.; Collado, D.; Jin, X.; Zambrana, J.V.; Jaconi, S.; Cameroni, E.; Saborio, S.; Rovida, F. Antibody-based assay discriminates Zika virus infection from other flaviviruses. Proc. Natl. Acad. Sci. USA 2017, 114, 8384–8389. [Google Scholar] [CrossRef]
- Wang, Q.; Yan, J.; Gao, G.F. Monoclonal Antibodies against Zika Virus: Therapeutics and Their Implications for Vaccine Design. J. Virol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Stettler, K.; Beltramello, M.; Espinosa, D.A.; Graham, V.; Cassotta, A.; Bianchi, S.; Vanzetta, F.; Minola, A.; Jaconi, S.; Mele, F. Specificity, cross-reactivity, and function of antibodies elicited by Zika virus infection. Science 2016, 353, 823–826. [Google Scholar] [CrossRef] [PubMed]
- Beltramello, M.; Williams, K.L.; Simmons, C.P.; Macagno, A.; Simonelli, L.; Quyen, N.T.; Sukupolvi-Petty, S.; Navarro-Sanchez, E.; Young, P.R.; de Silva, A.M. The human immune response to Dengue virus is dominated by highly cross-reactive antibodies endowed with neutralizing and enhancing activity. Cell Host Microbe 2010, 8, 271–283. [Google Scholar] [CrossRef] [PubMed]
- Dejnirattisai, W.; Wongwiwat, W.; Supasa, S.; Zhang, X.; Dai, X.; Rouvinski, A.; Jumnainsong, A.; Edwards, C.; Quyen, N.T.H.; Duangchinda, T. A new class of highly potent, broadly neutralizing antibodies isolated from viremic patients infected with dengue virus. Nat. Immunol. 2015, 16, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Henchal, E.A.; Gentry, M.K.; McCown, J.M.; Brandt, W.E. Dengue virus-specific and flavivirus group determinants identified with monoclonal antibodies by indirect immunofluorescence. Am. J. Trop. Med. Hyg. 1982, 31, 830–836. [Google Scholar] [CrossRef] [PubMed]
- Halstead, S.B. Dengue Antibody-Dependent Enhancement: Knowns and Unknowns. Microbiol. Spectr. 2014. [Google Scholar] [CrossRef] [PubMed]
- Takada, A.; Kawaoka, Y. Antibody-dependent enhancement of viral infection: Molecular mechanisms and in vivo implications. Rev. Med. Virol. 2003, 13, 387–398. [Google Scholar] [CrossRef] [PubMed]
- Pierson, T.C. Modeling antibody-enhanced dengue virus infection and disease in mice: Protection or pathogenesis? Cell Host Microbe 2010, 7, 85–86. [Google Scholar] [CrossRef]
- Moureau, G.; Cook, S.; Lemey, P.; Nougairede, A.; Forrester, N.L.; Khasnatinov, M.; Charrel, R.N.; Firth, A.E.; Gould, E.A.; de Lamballerie, X. New Insights into Flavivirus Evolution, Taxonomy and Biogeographic History, Extended by Analysis of Canonical and Alternative Coding Sequences. PLoS ONE 2015, 10, e0117849. [Google Scholar] [CrossRef]
- Brault, A.C. Changing patterns of West Nile virus transmission: Altered vector competence and host susceptibility. Vet. Res. 2009, 40, 1–19. [Google Scholar] [CrossRef]
- Musso, D.; Gubler, D.J. Zika Virus. Clin. Microbiol. Rev. 2016, 29, 487–524. [Google Scholar] [CrossRef]
- Barr, K.L.; Anderson, B.D. Dengue viruses exhibit strain-specific infectivity and entry requirements in vitro. Virus Adapt. Treat. 2013, 5, 1–9. [Google Scholar]
- Barba-Spaeth, G.; Dejnirattisai, W.; Rouvinski, A.; Vaney, M.C.; Medits, I.; Sharma, A.; Simon-Lorière, E.; Sakuntabhai, A.; Cao-Lormeau, V.M.; Haouz, A. Structural basis of potent Zika-dengue virus antibody cross-neutralization. Nature 2016, 536, 48–53. [Google Scholar] [CrossRef]
- Austin, S.; Dowd, K.; Shrestha, B.; Nelson, C.; Edeling, M.; Johnson, S.; Pierson, T.; Diamond, M.; Fremont, D. Structural Basis of Differential Neutralization of DENV−1 Genotypes by an Antibody that Recognizes a Cryptic Epitope. PLoS Pathog. 2012, 8, e1002930. [Google Scholar] [CrossRef] [PubMed]
- Dowd, K.A.; DeMaso, C.R.; Pelc, R.S.; Speer, S.D.; Smith, A.R.; Goo, L.; Platt, D.J.; Mascola, J.R.; Graham, B.S.; Mulligan, M.J. Broadly Neutralizing Activity of Zika Virus-Immune Sera Identifies a Single Viral Serotype. Cell Rep. 2016, 16, 1485–1491. [Google Scholar] [CrossRef] [PubMed]
- Sharp, T.M.; Fischer, M.; Muñoz-Jordán, J.L.; Paz-Bailey, G.; Staples, J.E.; Gregory, C.J.; Waterman, S.H. Dengue and Zika Virus Diagnostic Testing for Patients with a Clinically Compatible Illness and Risk for Infection with Both Viruses. MMWR Recomm. Rep. 2019, 68, 10. [Google Scholar] [CrossRef] [PubMed]
- Beck, C.; Leparc-Goffart, I.; Desoutter, D.; Debergé, E.; Bichet, H.; Lowenski, S.; Dumarest, M.; Gonzalez, G.; Migné, C.; Vanhomwegen, J. Serological evidence of infection with dengue and Zika viruses in horses on French Pacific Islands. PLoS Negl. Trop. Dis. 2019, 13, e0007162. [Google Scholar] [CrossRef]
- Venturi, G.; Fortuna, C.; Alves, R.M.; Passos do Prado Paschoal, A.G.; da Silva Junior, P.J.; Remoli, M.E.; Benedetti, E.; Amendola, A.; da Silva Batista, E.; Gama, D.V.N. Epidemiological and clinical suspicion of congenital Zika virus infection: Serological findings in mothers and children from Brazil. J. Med. Virol. 2019, 91, 1577–1583. [Google Scholar] [CrossRef]
- Kisuya, B.; Masika, M.M.; Bahizire, E.; Oyugi, J.O. Seroprevalence of Zika virus in selected regions in Kenya. Trans. R. Soc. Trop. Med. Hyg. 2019, 113, 735–739. [Google Scholar] [CrossRef]
- Montoya, M.; Collins, M.; Dejnirattisai, W.; Katzelnick, L.C.; Puerta-Guardo, H.; Jadi, R.; Puerta-Guardo, H.; Jadi, R.; Schlidhauer, S.; Supasa, P.; et al. Longitudinal Analysis of Antibody Cross-neutralization Following Zika Virus and Dengue Virus Infection in Asia and the Americas. J. Infect. Dis. 2018, 218, 536–545. [Google Scholar]
- Voermans, J.J.; Pas, S.D.; van der Linden, A.; GeurtsvanKessel, C.; Koopmans, M.; van der Eijk, A.; Reusken, C.B.E.M. Whole-Blood Testing for Diagnosis of Acute Zika Virus Infections in Routine Diagnostic Setting. Emerg. Infect. Dis. 2019, 25, 1394. [Google Scholar] [CrossRef]
- Mansuy, J.M.; Mengelle, C.; Pasquier, C.; Chapuy-Regaud, S.; Delobel, P.; Martin-Blondel, G.; Izopet, J. Zika Virus Infection and Prolonged Viremia in Whole-Blood Specimens. Emerg. Infect. Dis. 2017, 23, 863–865. [Google Scholar] [CrossRef]
- Rossini, G.; Gaibani, P.; Vocale, C.; Cagarelli, R.; Landini, M.P. Comparison of Zika virus (ZIKV) RNA detection in plasma, whole blood and urine—Case series of travel-associated ZIKV infection imported to Italy, 2016. J. Infect. 2017, 75, 242–245. [Google Scholar] [CrossRef]
- Baylis, S.A.; Blumel, J. External Quality Assessment for Zika Virus Molecular Diagnostic Testing, Brazil. Emerg. Infect. Dis. 2018, 24, 1966. [Google Scholar] [CrossRef] [PubMed]
- Baylis, S.A.; Hanschmann, K.O.; Schnierle, B.S.; Trosemeier, J.H.; Blumel, J. Harmonization of nucleic acid testing for Zika virus: Development of the 1(st) World Health Organization International Standard. Transfusion 2017, 57, 748–761. [Google Scholar] [CrossRef] [PubMed]
- Henchal, E.A.; McCown, J.M.; Burke, D.S.; Seguin, M.C.; Brandt, W.E. Epitopic analysis of antigenic determinants on the surface of dengue−2 virions using monoclonal antibodies. Am. J. Trop. Med. Hyg. 1985, 34, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Bardelli, M.; Espinosa, D.A.; Pedotti, M.; Ng, T.-S.; Bianchi, S.; Simonelli, L.; Lim, E.X.Y.; Foglierini, M.; Zatta, F. A Human Bi-specific Antibody against Zika Virus with High Therapeutic Potential. Cell 2017, 171, 229–241. [Google Scholar] [CrossRef]
- Khan, E.; Farooqi, J.Q.; Barr, K.L.; Prakoso, D.; Nasir, A.; Kanji, A.; Shakoor, S.; Malik, F.R.; Hasan, R.; Lednicky, J.A.; et al. Flaviviruses as a Cause of Undifferentiated Fever in Sindh Province, Pakistan: A Preliminary Report. Front. Public Health 2016, 4, 8. [Google Scholar] [CrossRef]
- Costin, J.M.; Jenwitheesuk, E.; Lok, S.M.; Hunsperger, E.; Conrads, K.A.; Fontaine, K.A.; Rees, C.R.; Rossmann, M.G.; Isern, S.; Samudrala, R. Structural optimization and de novo design of dengue virus entry inhibitory peptides. PLoS Negl. Trop. Dis. 2010, 4, e721. [Google Scholar] [CrossRef]
- Schieffelin, J.S.; Costin, J.M.; Nicholson, C.O.; Orgeron, N.M.; Fontaine, K.A.; Isern, S.; Michael, S.F.; Robinson, J.E. Neutralizing and non-neutralizing monoclonal antibodies against dengue virus E protein derived from a naturally infected patient. Virol. J. 2010, 7, 28. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AVG Ct ± SE | AVG CT ± SE | AVG Fold Change ± SE | Log Fold Change | p-Value | |
|---|---|---|---|---|---|
| MR−766 | 21.6 ± 0.04 | ||||
| PRVABC59 | 25.18 ± 0.02 | 3.57 ± 0.01 | 0.08 ± 0 | −3.5767 | 0.0000014 |
| R103451 | 21.02 ± 0.06 | −0.58 ± 0.03 | 1.5 ± 0.03 | 0.5863 | 0.0035 |
| Patient ID | Virus | ||||||
|---|---|---|---|---|---|---|---|
| DENV 1 | DENV 2 | DENV 3 | DENV 4 | YFV | WNV | JEV | |
| ZIKV Exposure with Unknown Exposure History | |||||||
| 50616 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| 50620 | 0 ± 0 | 0 ± 0 | 99.5 ± 0.7 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 67 ± 0.7 |
| 50622 | 0 ± 0 | 0 ± 0 | 99.5 ± 0.7 | 0 ± 0 | 97 ± 0.7 | 0 ± 0 | 61 ± 2.1 |
| 125 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| 88 | 0 ± 0 | 100 ± 0 | 99.5 ± 0.7 | 0 ± 0 | 92 ± 1.4 | 0 ± 0 | 13 ± 3.5 |
| Flavivirus Naïve SPF Sheep with ZIKV Infection | |||||||
| 4155 | 0 ± 0 | 100 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 79 ± 0.7 |
| 4158 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 97 ± 0.7 | 0 ± 0 | 78 ± 5.6 |
| 4072 | 0 ± 0 | 78 ± 1.4 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 82 ± 0 |
| 4171 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 100 ± 0 | 0 ± 0 | 0 ± 0 |
| ZIKV Naïve with Unknown Flavivirus Exposure | |||||||
| 315 | 100 ± 0 | 0 ± 0 | 100 ± 0 | 0 ± 0 | 86.6 ± 1.4 | 85.3 ± 4.2 | 93.2 ± 2.8 |
| 070 | 100 ± 0 | 100 ± 0 | 0 ± 0 | 0 ± 0 | 96.6 ± 1.4 | 47 ± 9.9 | 0 ± 0 |
| 200 | 100 ± 0 | 100 ± 0 | 100 ± 0 | 100 ± 0 | 55 ± 6.3 | 0 ± 0 | 92.2 |
| 252 | 98.1 ± 2.5 | 100 ± 0 | 100 ± 0 | 100 ± 0 | 0 ± 0 | 58.5 ± 1.4 | 91.2 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barr, K.L.; Schwarz, E.R.; Prakoso, D.; Imtiaz, K.; Pu, R.; Morris, J.G., Jr.; Khan, E.; Long, M.T. Strain-Dependent Activity of Zika Virus and Exposure History in Serological Diagnostics. Trop. Med. Infect. Dis. 2020, 5, 38. https://doi.org/10.3390/tropicalmed5010038
Barr KL, Schwarz ER, Prakoso D, Imtiaz K, Pu R, Morris JG Jr., Khan E, Long MT. Strain-Dependent Activity of Zika Virus and Exposure History in Serological Diagnostics. Tropical Medicine and Infectious Disease. 2020; 5(1):38. https://doi.org/10.3390/tropicalmed5010038
Chicago/Turabian StyleBarr, Kelli L., Erika R. Schwarz, Dhani Prakoso, Kehkashan Imtiaz, Ruiyu Pu, J. Glenn Morris, Jr., Erum Khan, and Maureen T. Long. 2020. "Strain-Dependent Activity of Zika Virus and Exposure History in Serological Diagnostics" Tropical Medicine and Infectious Disease 5, no. 1: 38. https://doi.org/10.3390/tropicalmed5010038
APA StyleBarr, K. L., Schwarz, E. R., Prakoso, D., Imtiaz, K., Pu, R., Morris, J. G., Jr., Khan, E., & Long, M. T. (2020). Strain-Dependent Activity of Zika Virus and Exposure History in Serological Diagnostics. Tropical Medicine and Infectious Disease, 5(1), 38. https://doi.org/10.3390/tropicalmed5010038