Risk Modeling of Bat Rabies in the Caribbean Islands

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
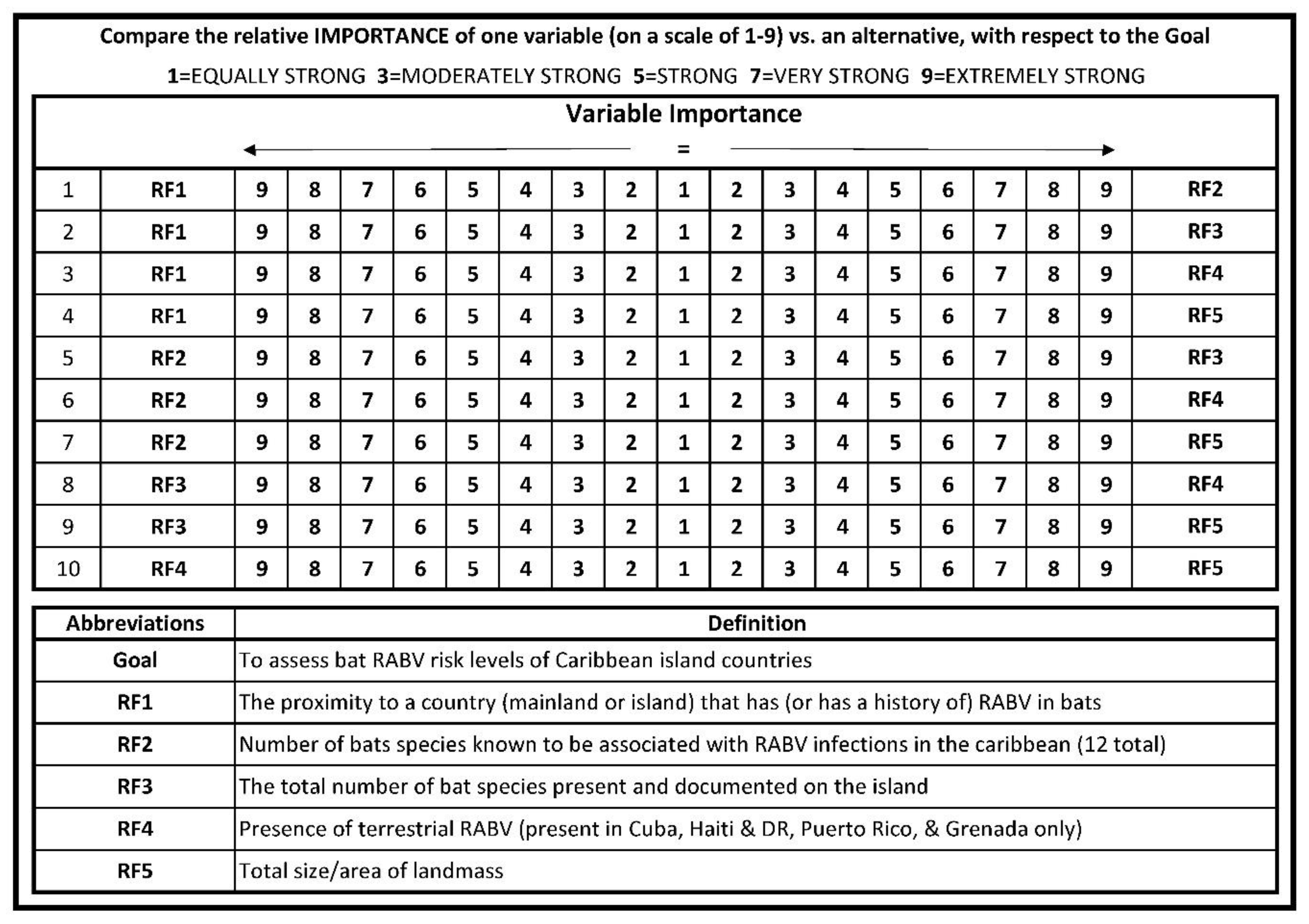
2.1. Risk Factor Selection
2.2. Questionnaire and Subject Matter Expert Selection
2.3. Geospatial Analysis
2.4. Risk Categorization
2.5. Risk Value Calculation
2.6. Model Validation and High-Risk Threshold
3. Results
4. Discussion
4.1. Risk Factors and Weighting
4.2. Evidence for Over-Water Bat Movements
4.3. Bat Rabies Surveillance Efforts
4.4. Island-Specifc Risk
4.5. Study Limitations
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Velasco-Villa, A.; Mauldin, M.R.; Shi, M.; Escobar, L.E.; Gallardo-Romero, N.F.; Damon, I.; Olson, V.A.; Streicker, D.G.; Emerson, G. The history of rabies in the Western Hemisphere. Antivir. Res. 2017, 146, 221–232. [Google Scholar] [CrossRef] [PubMed]
- Belotto, A.; Leanes, L.F.; Schneider, M.C.; Tamayo, H.; Correa, E. Overview of rabies in the Americas. Virus Res. 2005, 111, 5–12. [Google Scholar] [CrossRef] [PubMed]
- Vos, A.; Nunan, C.; Bolles, D.; Müller, T.; Fooks, A.R.; Tordo, N.; Baer, G.M. The occurrence of rabies in pre-Columbian Central America: An historical search. Epidemiol. Infect. 2011, 139, 1445–1452. [Google Scholar] [CrossRef] [PubMed]
- Escobar, L.; Peterson, A.T.; Favi, M.; Yung, V.; Medina-Vogel, G. Bat-borne rabies in Latin America. Rev. Inst. Med. Trop. São Paulo 2015, 57, 63–72. [Google Scholar] [CrossRef]
- Nehaul, B.B.G. Rabies transmitted by bats in British Guiana. Am. J. Trop. Med. Hyg. 1955, 4, 550–553. [Google Scholar] [CrossRef]
- Schneider, M.C.; Romijn, P.C.; Uieda, W.; Tamayo, H.; Silva, D.F.D.; Belotto, A.; Silva, J.B.D.; Leanes, L.F. Rabies transmitted by vampire bats to humans: An emerging zoonotic disease in Latin America? Rev. Panam. Salud Pública 2009, 25, 260–269. [Google Scholar] [CrossRef]
- Ma, X.; Monroe, B.P.; Cleaton, J.M.; Orciari, L.A.; Yager, P.; Li, Y.; Kirby, J.D.; Blanton, J.D.; Petersen, B.W.; Wallace, R.M. Rabies surveillance in the United States during 2016. J. Am. Vet. Med Assoc. 2018, 252, 945–957. [Google Scholar] [CrossRef]
- Condori-Condori, R.E.; Streicker, D.G.; Cabezas-Sanchez, C.; Velasco-Villa, A. Enzootic and epizootic rabies associated with vampire bats, Peru. Emerg. Infect. Dis. 2013, 19, 1463. [Google Scholar] [CrossRef]
- Lee, D.N.; Papeş, M.; Van Den Bussche, R.A. Present and potential future distribution of common vampire bats in the Americas and the associated risk to cattle. PLoS ONE 2012, 7, e42466. [Google Scholar] [CrossRef]
- Seetahal, J.; Vokaty, A.; Vigilato, M.; Carrington, C.; Pradel, J.; Louison, B.; Sauers, A.; Roopnarine, R.; Arrebato, J.; Millien, M.; et al. Rabies in the Caribbean: A situational analysis and historic review. Trop. Med. Infect. Dis. 2018, 3, 89. [Google Scholar] [CrossRef]
- Seetahal, J.F.; Greenberg, L.; Satheshkumar, P.S.; Sanchez-Vazquez, M.J.; Legall, G.; Singh, S.; Ramkissoon, V.; Schountz, T.; Munster, V.; Oura, C.A.; et al. The serological prevalence of rabies virus-neutralizing antibodies in the bat population on the Caribbean island of Trinidad. Viruses 2020, 12, 178. [Google Scholar] [CrossRef] [PubMed]
- Nadin-Davis, S.A.; Torres, G.; Ribas, M.D.L.A.; Guzman, M.; De La Paz, R.C.; Morales, M.; Wandeler, A.I. A molecular epidemiological study of rabies in Cuba. Epidemiol. Infect. 2006, 134, 1313–1324. [Google Scholar] [CrossRef] [PubMed]
- Vigilato, M.A.N.; Clavijo, A.; Knobl, T.; Silva, H.M.T.; Cosivi, O.; Schneider, M.C.; Leanes, L.F.; Belotto, A.J.; Espinal, M.A. Progress towards eliminating canine rabies: Policies and perspectives from Latin America and the Caribbean. Philos. Trans. R. Soc. B Biol. Sci. 2013, 368, 20120143. [Google Scholar] [CrossRef] [PubMed]
- Zieger, U.; Cheetham, S.; Santana, S.E.; Leiser-Miller, L.; Matthew-Belmar, V.; Goharriz, H.; Fooks, A.R. Natural exposure of bats in Grenada to rabies virus. Infect. Ecol. Epidemiol. 2017, 7, 1332935. [Google Scholar] [CrossRef] [PubMed]
- Organizacion Panamericana de la Salud (OPS). Republica dominicana: Elimination of Dog Transmitted Rabies in Latin America: Situation Analysis; Organizacion Panamericana de la Salud: Washington, DC, USA, 2004; pp. 58–59. [Google Scholar]
- Constantine, D.G. Geographic translocation of bats: Known and potential problems. Emerg. Infect. Dis. 2003, 9, 17. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, S.C.; Kwiecinski, G.G.; Genoways, H.H.; Larsen, R.J.; Larsen, P.A.; Phillips, C.J.; Baker, R.J. Bats of St. Lucia, Lesser Antilles. Special Publications, Museum of Texas Tech University. Acta Chiropterol. 2018, 69, 1–61. [Google Scholar]
- Forde, M.; Morrison, K.; Dewailly, E.; Badrie, N.; Robertson, L. Strengthening integrated research and capacity development within the Caribbean region. BMC Int. Health Hum. Rights 2011, 11, S7. [Google Scholar] [CrossRef]
- Del Rio Vilas, V.J.; Burgeño, A.; Montibeller, G.; Clavijo, A.; Vigilato, M.A.; Cosivi, O. Prioritization of capacities for the elimination of dog-mediated human rabies in the Americas: Building the framework. Pathog. Glob. Health 2013, 107, 340–345. [Google Scholar] [CrossRef]
- Saaty, T.L. Analytic Heirarchy Process. In Wiley StatsRef: Statistics Reference Online; Wiley: Hoboken, NJ, USA, 1980. [Google Scholar]
- Siddayao, G.P.; Valdez, S.E.; Fernandez, P.L. Analytic hierarchy process (AHP) in spatial modeling for floodplain risk assessment. Int. J. Mach. Learn. Comput. 2014, 4, 450. [Google Scholar] [CrossRef]
- Li, X.; Liu, T.; Lin, L.; Song, T.; Du, X.; Lin, H.; Xiao, J.; He, J.; Liu, L.; Zhu, G.; et al. Application of the analytic hierarchy approach to the risk assessment of Zika virus disease transmission in Guangdong Province, China. BMC Infect. Dis. 2017, 17, 65. [Google Scholar] [CrossRef]
- Ohashi, T.; Sugiyama, K.; Koba, Y.; Hasegawa, A.; Yamamoto, T.; Tsutsui, T. Development of a semi-quantitative evaluation system for surveillance of bovine spongiform encephalopathy, using the analytic hierarchy process. Rev. Sci. Tech. 2010, 29, 473. [Google Scholar] [CrossRef] [PubMed]
- Chan, H.K.; Wang, X. Fuzzy extent analysis for food risk assessment. In Fuzzy Hierarchical Model for Risk Assessment; Springer: London, UK, 2013; pp. 89–114. [Google Scholar]
- Marinoni, O. expAhp20—Analytic Hierarchy Process for ArcGIS. 2017. Available online: http://arcscripts.esri.com/details.asp?dbid=13764 (accessed on 10 September 2017).
- Goodwin, G.G.; Greenhall, A.M. A review of the bats of Trinidad and Tobago: Descriptions, rabies infection, and ecology. Am. Mus. Nat. Hist. Bull. AMNH 1961, 112, 1–301. [Google Scholar]
- Gomes, G.A.; Reid, F.A. Bats of Trinidad and Tobago: A Field Guide and Natural History; Trinibats: Port of Spain, Trinidad, 2015. [Google Scholar]
- Presley, S.J.; Willig, M.R. Bat metacommunity structure on Caribbean islands and the role of endemics. Global Ecol. Biogeogr. 2010, 19, 185–199. [Google Scholar] [CrossRef]
- McCracken, G.F.; Safi, K.; Kunz, T.H.; Dechmann, D.K.; Swartz, S.M.; Wikelski, M. Airplane tracking documents the fastest flight speeds recorded for bats. R. Soc. Open Sci. 2016, 3, 160398. [Google Scholar] [CrossRef]
- Egert-Berg, K.; Hurme, E.R.; Greif, S.; Goldstein, A.; Harten, L.; Flores-Martínez, J.J.; Valdés, A.T.; Johnston, D.S.; Eitan, O.; Borissov, I.; et al. Resource ephemerality drives social foraging in bats. Curr. Biol. 2018, 28, 3667–3673. [Google Scholar] [CrossRef]
- Medellin, R.A.; Rivero, M.; Ibarra, A.; de la Torre, J.A.; Gonzalez-Terrazas, T.P.; Torres-Knoop, L.; Tschapka, M. Follow me: Foraging distances of Leptonycteris yerbabuenae (Chiroptera: Phyllostomidae) in Sonora determined by fluorescent powder. J. Mammal. 2018, 99, 306–311. [Google Scholar] [CrossRef]
- Morrison, D.W. Flight speeds of some tropical forest bats. Am. Midl. Nat. 1980, 1, 189–192. [Google Scholar] [CrossRef]
- Brooks, A.M.; Richmond, C.L.; Warrendorf, D.L.; Woolley, J.; Lacher, T. Flight Speeds of Artibeus Jamaicensis and Monophyllus Plethodon. 2008. Available online: https://trinidad.tamu.edu/wildlife-projects/ (accessed on 9 November 2018).
- Morrison, D.W. Influence of habitat on the foraging distances of the fruit bat, Artibeus jamaicensis. J. Mammal. 1978, 59, 622–624. [Google Scholar] [CrossRef]
- Pedersen, S.C.; Genoways, H.H.; Morton, M.N.; Johnson, J.W.; Courts, S.E. Bats of Nevis, northern Lesser Antilles. Acta Chiropterol. 2003, 5, 251–268. [Google Scholar] [CrossRef][Green Version]
- Holland, R.A. Orientation and navigation in bats: Known unknowns or unknown unknowns? Behav. Ecol. Sociobiol. 2007, 61, 653–660. [Google Scholar] [CrossRef]
- Larsen, P.A.; Hoofer, S.R.; Bozeman, M.C.; Pedersen, S.C.; Genoways, H.H.; Phillips, C.J.; Pumo, D.E.; Baker, R.J. Phylogenetics and phylogeography of the Artibeus jamaicensis complex based on cytochrome-b DNA sequences. J. Mammal. 2007, 88, 712–727. [Google Scholar] [CrossRef]
- Larsen, R.J.; Larsen, P.A.; Genoways, H.H.; Catzeflis, F.M.; Geluso, K.; Kwiecinski, G.G.; Pedersen, S.C.; Simal, F.; Baker, R.J. Evolutionary history of Caribbean species of Myotis, with evidence of a third Lesser Antillean endemic. Mamm. Biol. 2012, 77, 124–134. [Google Scholar] [CrossRef]
- Loureiro, L.O.; Lim, B.K.; Engstrom, M.D. A new species of mastiff bat (Chiroptera, Molossidae, Molossus) from Guyana and Ecuador. Mamm. Biol. 2018, 90, 10–21. [Google Scholar] [CrossRef]
- Wilson, D.E.; Reeder, D.M. Mammal Species of the World: A Taxonomic and Geographic Reference, 3rd ed.; JHU Press: Baltimore, MD, USA, 2005. [Google Scholar]
- Bekker, J.P. The Mammals of Aruba (Mammalia: Chiroptera, Rodentia, Lagomorpha); Vereniging voor Zoogdierkunde en Zoogdierbescherming: Nijmegen, The Netherlands, 1996. [Google Scholar]
- Bell, G.P. A possible case of interspecific transmission of rabies in insectivorous bats. J. Mammal. 1980, 61, 528–530. [Google Scholar] [CrossRef] [PubMed]
- Calisher, C.H.; Childs, J.E.; Field, H.E.; Holmes, K.V.; Schountz, T. Bats: Important reservoir hosts of emerging viruses. Clin. Microbiol. Rev. 2006, 19, 531–545. [Google Scholar] [CrossRef]
- MacArthur, R.H.; Wilson, E.O. An equilibrium theory of insular zoogeography. Evolution 1963, 17, 373–387. [Google Scholar] [CrossRef]
- Hoffman, J.D.; Kadlubar, G.D.; Pedersen, S.C.; Larsen, R.J.; Larsen, P.A.; Phillips, C.J.; Kwiecinski, G.G.; Genoways, H.H. Predictors of bat species richness within the Caribbean basin. Spec. Publ. Mus. Tex. Tech Univ. 2019, 71, 337–377. [Google Scholar]
- Loureiro, L.O.; Engstrom, M.D.; Lim, B.K. Comparative phylogeography of mainland and insular species of Neotropical molossid bats (Molossus). Ecol. Evol. 2020, 10, 389–409. [Google Scholar] [CrossRef]
- Muscarella, R.A.; Murray, K.L.; Ortt, D.; Russell, A.L.; Fleming, T.H. Exploring demographic, physical, and historical explanations for the genetic structure of two lineages of Greater Antillean bats. PLoS ONE 2011, 6, e17704. [Google Scholar] [CrossRef]
- Carstens, B.C.; Sullivan, J.; Davalos, L.M.; Larsen, P.A.; Pedersen, S.C. Exploring population genetic structure in three species of Lesser Antillean bats. Mol. Ecol. 2004, 13, 2557–2566. [Google Scholar] [CrossRef]
- Speer, K.A.; Petronio, B.J.; Simmons, N.B.; Richey, R.; Magrini, K.; Soto-Centeno, J.A.; Reed, D.L. Population structure of a widespread bat (Tadarida brasiliensis) in an island system. Ecol. Evol. 2017, 7, 7585–7598. [Google Scholar] [CrossRef] [PubMed]
- Frank, P.A. First record of Molossus molossus tropidorhynchus Gray from the United States. J. Mammal. 1997, 78, 103–105. [Google Scholar] [CrossRef][Green Version]
- Van Gelder, R.G.; Wingate, D.B. The taxonomy and status of bats in Bermuda. Am. Mus. Nat. Hist. Novit. 1961, 2029, 1–9. [Google Scholar]
- Baagøe, H.J.; Bloch, D. Bats (Chiroptera) in the Faroe Islands. Frödskaparrit 1994, 41, 83–88. [Google Scholar]
- Krauel, J.J.; McCracken, G.F. Recent advances in bat migration research. In Bat Evolution, Ecology, and Conservation; Springer: New York, NY, USA, 2013; pp. 293–313. [Google Scholar]
- Hatch, S.K.; Connelly, E.E.; Divoll, T.J.; Stenhouse, I.J.; Williams, K.A. Offshore observations of eastern red bats (Lasiurus borealis) in the mid-Atlantic United States using multiple survey methods. PLoS ONE 2013, 8, e83803. [Google Scholar] [CrossRef]
- Boshamer, J.P.; Bekker, J.P. Nathusius’ pipistrelles (Pipistrellus nathusii) and other species of bats on offshore platforms in the Dutch sector of the North Sea. Lutra 2008, 51, 17. [Google Scholar]
- Fleming, T.H.; Murray, K.L.; Carstens, B. Phylogeography and genetic structure of three evolutionary lineages of West Indian Phyllostomid bats. In Island Bats: Evolution, Ecology and Conservation; University of Chicago Press: Chicago, IL, USA, 2010; pp. 116–150. [Google Scholar]
- Pedersen, S.C.; Kwiecinski, G.G.; Larsen, P.A.; Morton, M.N.; Adams, R.A.; Genoways, H.H.; Swier, V.J. Bats of Montserrat: Population fluctuation and response to hurricanes and volcanoes, 1978–2005. In Island Bats: Evolution, Ecology and Conservation; University of Chicago Press: Chicago, IL, USA, 2009; pp. 302–340. [Google Scholar]
- Rodríguez-Durán, A. Bat assemblages in the West Indies: The role of caves. In Island Bats: Evolution, Ecology and Conservation; University of Chicago Press: Chicago, IL, USA, 2009; pp. 265–280. [Google Scholar]
- Fleming, T.H.; Murray, K.L. Population and genetic consequences of hurricanes for three species of West Indian Phyllostomid bats. Biotropica 2008, 41, 250–256. [Google Scholar] [CrossRef]
- Chiou, H.Y.; Jeng, C.R.; Wang, H.Y.; Inoue, S.; Chan, F.T.; Liao, J.W.; Chang, M.T.; Pang, V.F. Pathology and molecular detection of rabies virus in ferret badgers associated with a rabies outbreak in Taiwan. J. Wildl. Dis. 2016, 52, 57–69. [Google Scholar] [CrossRef]
- Wu, H.; Chang, S.S.; Tsai, H.J.; Wallace, R.M.; Recuenco, S.E.; Doty, J.B.; Vora, N.M.; Chang, F.Y. Notes from the field: Wildlife rabies on an island free from canine rabies for 52 years—Taiwan, 2013. MMWR Morb. Mortal. Wkly Rep. 2014, 63, 178. [Google Scholar]
- Mélade, J.; McCulloch, S.; Ramasindrazana, B.; Lagadec, E.; Turpin, M.; Pascalis, H.; Goodman, S.M.; Markotter, W.; Dellagi, K. Serological evidence of lyssaviruses among bats on Southwestern Indian Ocean Islands. PLoS ONE 2016, 11, e0160553. [Google Scholar] [CrossRef]
- Price, J.L.; Everard, C.O. Rabies virus and antibody in bats in Grenada and Trinidad. J. Wildl. Dis. 1977, 13, 131–134. [Google Scholar] [CrossRef] [PubMed]
- Arguin, P.M.; Murray-Lillibridge, K.; Miranda, M.E.; Smith, J.S.; Calaor, A.B.; Rupprecht, C.E. Serologic evidence of Lyssavirus infections among bats, the Philippines. Emerg. Infect. Dis. 2002, 8, 258. [Google Scholar] [CrossRef] [PubMed]
- Speer, K.A.; Soto-Centeno, J.A.; Albury, N.A.; Quicksall, Z.; Marte, M.G.; Reed, D.L. Bats of the Bahamas: Natural history and conservation. Fla. Mus. Nat. Hist. Bull. 2015, 5, 45–95. [Google Scholar]
- Brunette, G.W. CDC Yellow Book 2018: Health Information for International Travel; Oxford University Press: Oxford, UK, 2017. [Google Scholar]
- World Health Organization. WHO expert consultation on rabies: Third report. In World Health Organization, Technical Report Series 1012; World Health Organization: Geneva, Switzerland, 2018. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Risk Factor Alternatives | |||||
|---|---|---|---|---|---|
| RF1a | RF2a | RF3a | RF4a | RF5a | |
| RF1 | 1 | 0.33 | 4 | 6 | 5 |
| RF2 | 3 | 1 | 5 | 6 | 6 |
| RF3 | 0.25 | 0.2 | 1 | 3 | 4 |
| RF4 | 0.125 | 0.125 | 0.33 | 1 | 2 |
| RF5 | 0.2 | 0.125 | 0.25 | 0.5 | 1 |
| Risk Factor | Variable Weight | |
|---|---|---|
| RF1 | Proximity to a country in which bat rabies has been detected | 0.28 |
| RF2 | Total number of rabies virus-associated bat species | 0.48 |
| RF3 | Total number of bat species present | 0.13 |
| RF4 | Presence of rabies in carnivores | 0.06 |
| RF5 | Island area (km2) | 0.05 |
| Ranking | Island(s) | Model Risk Value | Validation Risk Value |
|---|---|---|---|
| 1 | Cuba | 8.73 | 6.17 |
| 2 | Dominican Republic | 8.64 | 6.08 |
| Haiti | 8.64 | 6.08 | |
| 3 | Trinidad and Tobago | 8.25 | 5.69 |
| 4 | Grenada | 7.87 | 5.31 |
| Ranking | Self-Declared Bat Rabies-Free Island(s) | Model Risk Value |
|---|---|---|
| 1 | St. Vincent and the Grenadines | 7.87 |
| 2 | Jamaica | 6.37 |
| 3 | Puerto Rico | 6.34 |
| 4 | Cayman Islands | 5.92 |
| 5 | Dominica | 5.35 |
| 6 | St. Lucia | 3.75 |
| 7 | Martinique | 3.44 |
| Guadeloupe | 3.44 | |
| Montserrat | 3.44 | |
| 8 | Antigua | 3.18 |
| Barbados | 3.18 | |
| Anguilla | 3.18 | |
| Barbuda | 3.18 | |
| Nevis | 3.18 | |
| Saint-Martin | 3.18 | |
| Sint Maarten | 3.18 | |
| St. Kitts | 3.18 | |
| Saba | 3.18 | |
| Saint Barthelemy | 3.18 | |
| St. Eustatius | 3.18 | |
| 9 | Aruba | 3.16 |
| 10 | British Virgin Islands | 2.91 |
| U.S. Virgin Islands | 2.91 | |
| 11 | Bonaire | 2.40 |
| Curacao | 2.40 | |
| 12 | The Bahamas | 1.57 |
| Turks and Caicos Islands | 1.57 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morgan, C.N.; Wallace, R.M.; Vokaty, A.; Seetahal, J.F.R.; Nakazawa, Y.J. Risk Modeling of Bat Rabies in the Caribbean Islands. Trop. Med. Infect. Dis. 2020, 5, 35. https://doi.org/10.3390/tropicalmed5010035
Morgan CN, Wallace RM, Vokaty A, Seetahal JFR, Nakazawa YJ. Risk Modeling of Bat Rabies in the Caribbean Islands. Tropical Medicine and Infectious Disease. 2020; 5(1):35. https://doi.org/10.3390/tropicalmed5010035
Chicago/Turabian StyleMorgan, Clint N., Ryan M. Wallace, Alexandra Vokaty, Janine F.R. Seetahal, and Yoshinori J. Nakazawa. 2020. "Risk Modeling of Bat Rabies in the Caribbean Islands" Tropical Medicine and Infectious Disease 5, no. 1: 35. https://doi.org/10.3390/tropicalmed5010035
APA StyleMorgan, C. N., Wallace, R. M., Vokaty, A., Seetahal, J. F. R., & Nakazawa, Y. J. (2020). Risk Modeling of Bat Rabies in the Caribbean Islands. Tropical Medicine and Infectious Disease, 5(1), 35. https://doi.org/10.3390/tropicalmed5010035