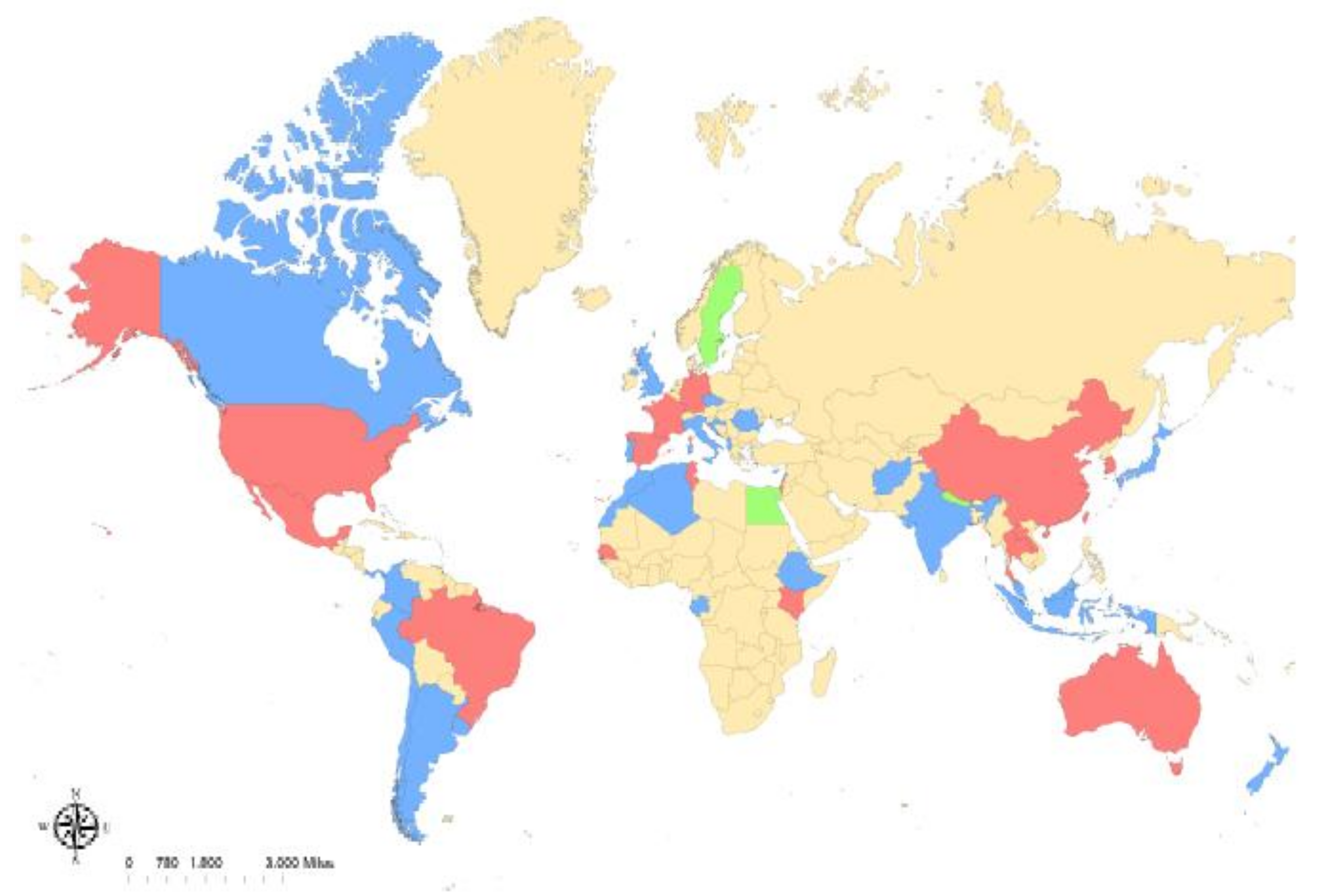
Rickettsia felis: A Review of Transmission Mechanisms of an Emerging Pathogen

{kind=link}
Abstract
:1. Introduction
2. Background
3. Clinical Disease
4. Transmission in Arthropods
5. Transmission to Vertebrates
6. Discussion
Acknowledgments
Conflicts of Interest
References
- Abdad, M.Y.; Stenos, J.; Graves, S. Rickettsia felis, an emerging flea-transmitted human pathogen. Emerg. Health Threat. J. 2011, 4, 7168. [Google Scholar] [CrossRef]
- Schriefer, M.E.; Sacci, J.B.; Taylor, J.P.; Higgins, J.A.; Azad, A.F. Identification of a novel rickettsial infection in a patient diagnosed with murine typhus. J. Clin. Microbiol. 1994, 32, 949–954. [Google Scholar] [PubMed]
- Brown, L.D.; Christofferson, R.C.; Banajee, K.H.; Del Piero, F.; Foil, L.D.; Macaluso, K.R. Cofeeding intra- and interspecific transmission of an emerging insect-borne rickettsial pathogen. Mol. Ecol. 2015, 24, 5475–5489. [Google Scholar] [CrossRef] [PubMed]
- Blanton, L.S.; Walker, D.H. Flea-borne rickettsioses and rickettsiae. Am. J. Trop. Med. Hyg. 2017, 96, 53–56. [Google Scholar] [CrossRef] [PubMed]
- Boostrom, A.; Beier, M.S.; Macaluso, J.A.; Macaluso, K.R.; Sprenger, D.; Hayes, J.; Radulovic, S.; Azad, A.F. Geographic association of Rickettsia felis-infected opossums with human murine typhus, Texas. Emerg. Infect. Dis. 2002, 8, 549–554. [Google Scholar] [CrossRef] [PubMed]
- Maina, A.N.; Fogarty, C.; Krueger, L.; Macaluso, K.R.; Odhiambo, A.; Nguyen, K.; Farris, C.M.; Luce-Fedrow, A.; Bennet, S.; Jiang, J.; et al. Rickettsial infections among Ctenocephalides felis and host animals during a flea-borne rickettsioses outbreak in Orange County, California. PLoS ONE 2016, 11. [Google Scholar] [CrossRef] [PubMed]
- Eremeeva, M.E.; Warashina, W.R.; Sturgeon, M.M.; Buchholz, A.E.; Olmsted, G.K.; Park, S.Y.; Effler, P.V.; Karpathy, S.E. Rickettsia typhi and R. felis in rat fleas (Xenopsylla cheopis), Oahu, Hawaii. Emerg. Infect. Dis. 2008, 14, 1613–1615. [Google Scholar] [CrossRef] [PubMed]
- Odhiambo, A.M.; Maina, A.N.; Taylor, M.L.; Jiang, J.; Richards, A.L. Development and validation of a quantitative real-time polymerase chain reaction assay specific for the detection of Rickettsia felis and not Rickettsia-felis like organisms. Vector Borne Zoonotic Dis. 2014, 14, 476–481. [Google Scholar] [CrossRef] [PubMed]
- Gillespie, J.J.; Beier, M.S.; Rahman, M.S.; Ammerman, N.C.; Shallom, J.M.; Purkayastha, A.; Sobral, B.S.; Azad, A.F. Plasmids and rickettsial evolution: Insight from Rickettsia felis. PLoS ONE 2007, 2. [Google Scholar] [CrossRef] [PubMed]
- Fang, R.; Raoult, D. Antigenic Classification of Rickettsia felis by using monoclonal and polyclonal antibodies. Clin. Vaccine Immunol. 2003, 10, 221–228. [Google Scholar] [CrossRef]
- Luce-Fedrow, A.; Mullins, K.; Kostik, A.P.; St. John, H.K.; Jiang, J.; Richards, A.L. Strategies for detecting rickettsiae and diagnosing rickettsial diseases. Future Microbiol. 2015, 10, 537–564. [Google Scholar] [CrossRef] [PubMed]
- Azad, A.F.; Sacci, J.B.; Nelson, W.M.; Dasch, G.A.; Schmidtmann, E.T.; Carl, M. Genetic characterization and transovarial transmission of a typhus-like Rickettsia found in cat fleas. Proc. Natl. Acad. Sci. USA 1992, 89, 43–46. [Google Scholar] [CrossRef] [PubMed]
- Parola, P. Rickettsia felis: From a rare disease in the USA to a common cause of fever in sub-Saharan Africa. Clin. Microbiol. Infect. 2011, 17, 996–1000. [Google Scholar] [CrossRef] [PubMed]
- Mediannikov, O.; Socolovschi, C.; Edouard, S.; Fenollar, F.; Mouffok, N.; Bassene, H.; Diatta, G.; Tall, A.; Niangaly, H.; Doumbo, O.; et al. Common epidemiology of Rickettsia felis infection and malaria, Africa. Emerg. Infect. Dis. 2013, 19, 1775–1783. [Google Scholar] [CrossRef] [PubMed]
- Mediannikov, O.; Fenollar, F.; Bassene, H.; Tall, A.; Sokhna, C.; Trape, J.F.; Raoult, D. Description of ‘yaaf’, the vesicular fever caused by acute Rickettsia felis infection in Senegal. J. Infect. 2013, 66, 536–540. [Google Scholar] [CrossRef] [PubMed]
- Angelakis, E.; Mediannikov, O.; Parola, P.; Raoult, D. Rickettsia felis: The complex journey of an emergent human pathogen. Trends Parasitol. 2016, 32, 554–564. [Google Scholar] [CrossRef] [PubMed]
- Socolovschi, C.; Pages, F.; Ndiath, M.O.; Ratmanov, P.; Raoult, D. Rickettsia species in African Anopheles mosquitoes. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Keita, A.K.; Socolovschi, C.; Ahuka-Mundeke, S.; Ratmanov, P.; Butel, C.; Ayouba, A.; Inogwabini, B.I.; Muyembe-Tamfum, J.J.; Mpoudi-Ngole, E.; Delaporte, E.; et al. Molecular evidence for the presence of Rickettsia felis in the feces of wild-living African apes. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Zavala-Velazquz, J.E.; Ruiz-Sosa, J.A.; Sanchez-Elias, R.A.; Becerra-Carmona, G.; Walker, D.H. Rickettsia felis rickettsiosis in Yucatan. Lancet 2000, 356, 1079–1080. [Google Scholar] [CrossRef]
- Brown, L.D.; Macaluso, K.R. Rickettsia felis, an emerging flea-borne rickettsiosis. Curr. Trop. Med. Rep. 2016, 3, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Reif, K.E.; Macaluso, K.R. Ecology of Rickettsia felis: A review. J. Med. Entomol. 2009, 46, 723–736. [Google Scholar] [CrossRef] [PubMed]
- Reif, K.E.; Kearney, M.T.; Foil, L.D.; Macaluso, K.R. Acquisition of Rickettsia felis by cat fleas during feeding. Vector Borne Zoonotic Dis. 2011, 11, 963–968. [Google Scholar] [CrossRef] [PubMed]
- Adams, J.R.; Schmidtmann, E.T.; Azad, A.F. Infection of colonized cat fleas, Ctenocephalides felis (Bouche) with Rickettsia-like microorganisms. Am. J. Trop. Med. Hyg. 1990, 43, 400–409. [Google Scholar] [CrossRef] [PubMed]
- Higgins, J.A.; Sacci, J.B.; Schriefer, M.E.; Endris, R.G.; Azad, A.F. Molecular identification of Rickettsia-like microorganisms associated with colonized cat fleas (Ctenocephalides felis). Insect Mol. Biol. 1994, 3, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Healy, S.P.; Brown, L.D.; Hagstrom, M.R.; Foil, L.D.; Macaluso, K.R. Effect of Rickettsia felis strain variation on infection, transmission, and fitness in the cat flea (Siphonaptera: Pulicidae). J. Med. Entomol. 2017, 54, 1037–1043. [Google Scholar] [CrossRef] [PubMed]
- Hirunkanokpun, S.; Thepparit, C.; Foil, L.D.; Macaluso, K.R. Horizontal transmission of Rickettsia felis between cat fleas, Ctenocephalides felis. Mol. Ecol. 2011, 20, 4577–4586. [Google Scholar] [CrossRef] [PubMed]
- Gillespie, J.J.; Driscoll, T.P.; Verhoeve, V.I.; Utsuki, T.; Husseneder, C.; Chouljenko, V.N.; Azad, A.F.; Macaluso, K.R. Genomic diversification in strains of Rickettsia felis Isolated from different arthropods. Genome Biol. Evol. 2014, 7, 35–56. [Google Scholar] [CrossRef] [PubMed]
- Macaluso, K.R.; Pornwiroon, W.; Popov, V.L.; Foil, L.D. Identification of Rickettsia felis in the salivary glands of cat fleas. Vector Borne Zoonotic Dis. 2008, 8, 391–396. [Google Scholar] [CrossRef] [PubMed]
- Thepparit, C.; Hirunkanokpun, S.; Popov, V.L.; Foil, L.D.; Macaluso, K.R. Dissemination of bloodmeal acquired Rickettsia felis in cat fleas, Ctenocephalides felis. Parasit. Vectors 2013, 6, 149. [Google Scholar] [CrossRef] [PubMed]
- Brown, L.D.; Banajee, K.H.; Foil, L.D.; Macaluso, K.R. Transmission mechanisms of an emerging insect-borne rickettsial pathogen. Parasit. Vectors 2016, 9, 237. [Google Scholar] [CrossRef] [PubMed]
- Harris, E.K.; Verhoeve, V.I.; Banajee, K.H.; Macaluso, J.A.; Azad, A.F.; Macaluso, K.R. Comparative vertical transmission of Rickettsia by Dermacentor variabilis and Ambylomma maculatum. Ticks Tick Borne Dis. 2017, 8, 598–604. [Google Scholar] [CrossRef] [PubMed]
- Dieme, C.; Bechah, Y.; Socolovschi, C.; Audoly, G.; Berenger, J.M.; Faye, O.; Raoult, D.; Parola, P. Transmission potential of Rickettsia felis infection by Anopheles gambiae mosquitoes. Proc. Natl. Acad. Sci. USA 2015, 112, 8088–8093. [Google Scholar] [CrossRef] [PubMed]
- Behar, A.; McCormick, L.J.; Periman, S.J. Rickettsia felis infection in a common household insect pest, Liposcelis bostrychophila (Psocoptera: Liposcelidae). Appl. Environ. Microbiol. 2010, 76, 2280–2285. [Google Scholar] [CrossRef] [PubMed]
- Thepparit, C.; Sunyakumthorn, P.; Guillotte, M.L.; Popov, V.L.; Foil, L.D.; Macaluso, K.R. Isolation of rickettsial pathogen from a non-hematophagous arthropod. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [PubMed]
- Yusuf, M.; Turner, B. Characterization of Wolbachia-like bacteria isolated from the parthenogenic stored-product pest psocid Liposcelis bostrychophila (Badonnel) (Psocoptera). J. Stored Prod. Res. 2004, 40, 207–225. [Google Scholar] [CrossRef]
- Wedincamp, J.; Foil, L.D. Infection and seroconversion of cats exposed to cat fleas (Ctenocephalides felis Bouche) infected with Rickettsia felis. J. Vector Ecol. 2000, 25, 123–126. [Google Scholar] [PubMed]
- Lappin, M.R.; Hawley, J. Presence of Bartonella species and Rickettsia species DNA in the blood, oral cavity, skin and claw beds of cats in the United States. Vet. Dermatol. 2009, 20, 509–514. [Google Scholar] [CrossRef] [PubMed]
- Azad, A.F.; Beard, C.B. Rickettsial pathogens and their arthropods. Emerg. Infect. Dis. 1998, 4, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Azad, A.F.; Traub, R. Transmission of murine typhus rickettsiae by Xenopsylla cheopis with notes on experimental infection and effects of temperature. Am. J. Trop. Med. Hyg. 1985, 34, 555–563. [Google Scholar] [CrossRef]
- Bechah, Y.; Capo, C.; Mege, J.L.; Raoult, D. Epidemic typhus. Lancet Infect. Dis. 2008, 8, 417–426. [Google Scholar] [CrossRef]
- Foil, L.D.; Andress, E.; Freeland, R.L.; Roy, A.F.; Rutledge, R.; Triche, P.C.; O’Reily, K.L. Experimental infection of domestic cats with Bartonella henselae by inoculation of Ctenocephalides felis (Siphonaptera: Pulicidae) feces. J. Med. Entomol. 1998, 35, 625–628. [Google Scholar] [CrossRef] [PubMed]
- Grasperge, B.J.; Reif, K.E.; Morgan, T.D.; Sunyakumthorn, P.; Bynog, J.; Paddock, C.D.; Macaluso, K.R. Susceptibility of inbred mice to Rickettsia parkeri. Infect. Immun. 2012, 80, 1846–1852. [Google Scholar] [CrossRef] [PubMed]
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Legendre, K.P.; Macaluso, K.R. Rickettsia felis: A Review of Transmission Mechanisms of an Emerging Pathogen. Trop. Med. Infect. Dis. 2017, 2, 64. https://doi.org/10.3390/tropicalmed2040064
Legendre KP, Macaluso KR. Rickettsia felis: A Review of Transmission Mechanisms of an Emerging Pathogen. Tropical Medicine and Infectious Disease. 2017; 2(4):64. https://doi.org/10.3390/tropicalmed2040064
Chicago/Turabian StyleLegendre, Kelsey P., and Kevin R. Macaluso. 2017. "Rickettsia felis: A Review of Transmission Mechanisms of an Emerging Pathogen" Tropical Medicine and Infectious Disease 2, no. 4: 64. https://doi.org/10.3390/tropicalmed2040064
APA StyleLegendre, K. P., & Macaluso, K. R. (2017). Rickettsia felis: A Review of Transmission Mechanisms of an Emerging Pathogen. Tropical Medicine and Infectious Disease, 2(4), 64. https://doi.org/10.3390/tropicalmed2040064