
Limited Spectroscopy Data and Machine Learning for Detection of Zika Virus Infection in Aedes aegypti Mosquitoes

 , , , , and
, , , , and
Abstract
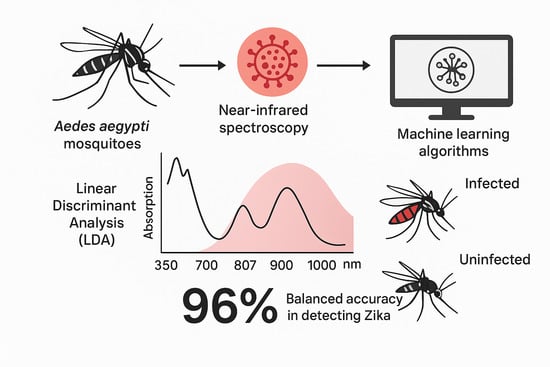
1. Introduction
2. Dataset
- The absorbance measurements at various wavelengths, ranging from 350 to 2500 nm;
- The infection status of the mosquito (infected or uninfected);
- The duration measured in days post infection (DPI).
3. Methodology
3.1. Applying LDA
3.2. SVM for Classification
4. Results
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huang, Y.J.S.; Higgs, S.; Vanlandingham, D.L. Emergence and re-emergence of mosquito-borne arboviruses. Curr. Opin. Virol. 2019, 34, 104–109. [Google Scholar] [CrossRef]
- Weaver, S.C.; Reisen, W.K. Present and future arboviral threats. Antivir. Res. 2010, 85, 328–345. [Google Scholar] [CrossRef]
- Weaver, S.C.; Charlier, C.; Vasilakis, N.; Lecuit, M. Zika, Chikungunya, and Other Emerging Vector-Borne Viral Diseases. Annu. Rev. Med. 2018, 69, 395–408. [Google Scholar] [CrossRef] [PubMed]
- Harrington, L.C.; Scott, T.W.; Lerdthusnee, K.; Coleman, R.C.; Costero, A.; Clark, G.G.; Jones, J.J.; Kitthawee, S.; Kittayapong, P.; Sithiprasasna, R.; et al. Dispersal of the dengue vector Aedes aegypti within and between rural communities. Am. J. Trop. Med. Hyg. 2005, 72, 209–220. [Google Scholar] [CrossRef] [PubMed]
- Maciel-de Freitas, R.; Lourenço-de Oliveira, R. Presumed unconstrained dispersal of Aedes aegypti in the city of Rio de Janeiro. Rev. Saúde Pública 2009, 43, 8–12. [Google Scholar] [CrossRef] [PubMed]
- Maciel-de-Freitas, R.; Codeço, C.T.; de Oliveira, R.L. Daily survival rates and dispersal of Aedes aegypti females in Rio de Janeiro, Brazil. Am. J. Trop. Med. Hyg. 2007, 76, 659–665. [Google Scholar] [CrossRef]
- Scott, T.W.; Chow, E.; Strickman, D.; Kittayapong, P.; Wirtz, R.A.; Lorenz, L.H.; Edman, J.D. Blood-Feeding Patterns of Aedes aegypti (Diptera: Culicidae) Collected in a Rural Thai Village. J. Med. Entomol. 1993, 30, 922–927. [Google Scholar] [CrossRef]
- Scott, T.W.; Amerasinghe, P.H.; Morrison, A.C.; Lorenz, L.H.; Clark, G.G.; Strickman, D.; Kittayapong, P.; Edman, J.D. Longitudinal Studies of Aedes aegypti (Diptera: Culicidae) in Thailand and Puerto Rico: Blood Feeding Frequency. J. Med. Entomol. 2000, 37, 89–101. [Google Scholar] [CrossRef]
- Maciel-de-Freitas, R.; Marques, W.d.A.; Peres, R.C.; Cunha, S.P.d.; Lourenço-de-Oliveira, R. Variation in Aedes aegypti (diptera: Culicidae) container productivity in a slum and a suburban district of rio de janeiro during dry and wet seasons. Memórias Inst. Oswaldo Cruz 2007, 102, 489–496. [Google Scholar] [CrossRef]
- Runge-Ranzinger, S.; McCall, P.J.; Kroeger, A.; Horstick, O. Dengue disease surveillance: An updated systematic literature review. Trop. Med. Int. Health 2014, 19, 1116–1160. [Google Scholar] [CrossRef]
- Jourdain, F.; Samy, A.M.; Hamidi, A.; Bouattour, A.; Alten, B.; Faraj, C.; Roiz, D.; Petrić, D.; Pérez-Ramírez, E.; Velo, E.; et al. Towards harmonisation of entomological surveillance in the Mediterranean area. PLoS Neglected Trop. Dis. 2019, 13, e0007314. [Google Scholar] [CrossRef] [PubMed]
- Codeço, C.T.; Lima, A.W.S.; Araújo, S.C.; Lima, J.B.P.; Maciel-de Freitas, R.; Honório, N.A.; Galardo, A.K.R.; Braga, I.A.; Coelho, G.E.; Valle, D. Surveillance of Aedes aegypti: Comparison of House Index with Four Alternative Traps. PLoS Neglected Trop. Dis. 2015, 9, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Leandro, A.; Maciel-de Freitas, R. Development of an Integrated Surveillance System to Improve Preparedness for Arbovirus Outbreaks in a Dengue Endemic Setting: Descriptive Study. JMIR Public Health Surveill 2024, 10, e62759. [Google Scholar] [CrossRef] [PubMed]
- Leandro, A.S.; Chiba de Castro, W.A.; Garey, M.V.; Maciel-de Freitas, R. Spatial analysis of dengue transmission in an endemic city in Brazil reveals high spatial structuring on local dengue transmission dynamics. Sci. Rep. 2024, 14, 8930. [Google Scholar] [CrossRef]
- Leandro, A.; Chiba de Castro, W.; Defante, R.; Delai, R.; Villela, D.; Maciel-de Freitas, R. Citywide Integrated Aedes aegypti Mosquito Surveillance as Early Warning System for Arbovirus Transmission, Brazil. Emerg. Infect. Dis. 2022, 28, 707. [Google Scholar] [CrossRef]
- Leandro, A.d.S.; Ayala, M.J.C.; Lopes, R.D.; Martins, C.A.; Maciel-de Freitas, R.; Villela, D.A.M. Entomo-Virological Aedes aegypti Surveillance Applied for Prediction of Dengue Transmission: A Spatio-Temporal Modeling Study. Pathogens 2023, 12, 4. [Google Scholar] [CrossRef]
- Runge-Ranzinger, S.; Horstick, O.; Marx, M.; Kroeger, A. What does dengue disease surveillance contribute to predicting and detecting outbreaks and describing trends? Trop. Med. Int. Health 2008, 13, 1022–1041. [Google Scholar] [CrossRef]
- Pepin, K.; Leach, C.; Marques-Toledo, C.; Laass, K.; Paixao, K.; Luis, A.; Hayman, D.; Johnson, N.; Buhnerkempe, M.; Carver, S.; et al. Utility of mosquito surveillance data for spatial prioritization of vector control against dengue viruses in three Brazilian cities. Parasites Vectors 2015, 8, 98. [Google Scholar] [CrossRef]
- Leandro, A.; Oliveira, F.; Defante, R.; Rivas, A.; Martins, C.; Silva, I.; Villela, D.; Teixeira, M.; Xavier, S.; Maciel-de Freitas, R. The fuzzy system ensembles entomological, epidemiological, demographic and environmental data to unravel the dengue transmission risk in an endemic city. BMC Public Health 2024, 24, 2587. [Google Scholar] [CrossRef]
- Gurgel-Gonçalves, R.; Oliveira, W.; Croda, J. The greatest Dengue epidemic in Brazil: Surveillance, Prevention, and Control. Rev. Soc. Bras. Med. Trop. 2024, 57, e002032024. [Google Scholar] [CrossRef]
- Lee, K.S.; Lai, Y.L.; Lo, S.; Barkham, T.; Aw, P.; Ooi, P.; Tai, J.; Hibberd, M.; Johansson, P.; Khoo, S.; et al. Dengue Virus Surveillance for Early Warning, Singapore. Emerg. Infect. Dis. 2010, 16, 847–849. [Google Scholar] [CrossRef]
- World Health Organization. Handbook for Integrated Vector Management; World Health Organization: Geneva, Switzerland, 2012. [Google Scholar]
- Beier, J.; Keating, J.; Githure, J.; Macdonald, M.; Impoinvil, D.; Novak, R. Integrated vector management for malaria control. Malar. J. 2008, 7 (Suppl. S1), S4. [Google Scholar] [CrossRef] [PubMed]
- da Silva, L.M.I.; Maciel-de Freitas, R.; Paiva, M.H.S.; da Luz Wallau, G. Horizons of the Future: Preparedness and Response. In Current Topics in Microbiology and Immunology; Springer: Berlin/Heidelberg, Germany, 2025; pp. 1–25. [Google Scholar] [CrossRef]
- Jose, S.A.; Mathew, K.; Hamna Mariyam, K.B.; Jirawattanapanit, A.; Weraprasertsakun, A. Analyzing and forecasting dengue fever incidence in Thailand: A comprehensive study for public health preparedness. Int. J. Biomath. 2024. [CrossRef]
- Jose, S.A.; Raja, R.; Omede, B.I.; Agarwal, R.P.; Alzabut, J.; Cao, J.; Balas, V.E. Mathematical modeling on co-infection: Transmission dynamics of Zika virus and Dengue fever. Nonlinear Dyn. 2023, 111, 4879–4914. [Google Scholar] [CrossRef]
- Joseph, D.; Ramachandran, R.; Alzabut, J.; Jose, S.A.; Khan, H. A Fractional-Order Density-Dependent Mathematical Model to Find the Better Strain of Wolbachia. Symmetry 2023, 15, 845. [Google Scholar] [CrossRef]
- Freitas, A.R.R.; Angerami, R.; Zuben, A.; Donalisio, M. Introduction and transmission of Zika virus in Brazil: New challenges for the Americas. Rev. Inst. Med. Trop. São Paulo 2016, 58, 24. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Barreto, M.; Barral-Netto, M.; Stabeli, R.; Almeida-Filho, N.; Vasconcelos, P.; Teixeira, M.; Buss, P.; Gadelha, P. Zika virus and microcephaly in Brazil: A scientific agenda. Lancet 2016, 387, 919–921. [Google Scholar] [CrossRef]
- Zanluca, C.; De Melo, V.C.A.; Mosimann, A.L.P.; Dos Santos, G.I.V.; dos Santos, C.N.D.; Luz, K. First report of autochthonous transmission of Zika virus in Brazil. Mem. Inst. Oswaldo Cruz 2015, 110, 569–572. [Google Scholar] [CrossRef]
- Faria, N.R.; do Socorro da Silva Azevedo, R.; Kraemer, M.U.G.; Souza, R.; Cunha, M.S.; Hill, S.C.; Thézé, J.; Bonsall, M.B.; Bowden, T.A.; Rissanen, I.; et al. Zika virus in the Americas: Early epidemiological and genetic findings. Science 2016, 352, 345–349. [Google Scholar] [CrossRef] [PubMed]
- Mhamadi, M.; Mencattelli, G.; Gaye, A.; Ndiaye, E.H.; Sow, A.A.; Faye, M.; Ndione, M.H.D.; Diagne, M.M.; Mhamadi, M.; Faye, O.; et al. Rapid On-Site Detection of Arboviruses by a Direct RT-qPCR Assay. Biosensors 2023, 13, 1035. [Google Scholar] [CrossRef]
- Bokobza, L. Near Infrared Spectroscopy. J. Near Infrared Spectrosc. 1998, 6, 3–17. [Google Scholar] [CrossRef]
- Sakudo, A.; Suganuma, Y.; Kobayashi, T.; Onodera, T.; Ikuta, K. Near-infrared spectroscopy: Promising diagnostic tool for viral infections. Biochem. Biophys. Res. Commun. 2006, 341, 279–284. [Google Scholar] [CrossRef]
- Sikulu-Lord, M.; de Freitas, R.M. Application of Infrared Techniques for Characterisation of Vector-Borne Disease Vectors. In Infrared Spectroscopy; IntechOpen: London, UK, 2022; Chapter 7. [Google Scholar] [CrossRef]
- Fernandes, J.N.; dos Santos, L.M.B.; Chouin-Carneiro, T.; Pavan, M.G.; Garcia, G.A.; David, M.R.; Beier, J.C.; Dowell, F.E.; de Freitas, R.M.; Sikulu-Lord, M.T. Rapid, noninvasive detection of Zika virus in Aedes aegypti mosquitoes by near-infrared spectroscopy. Sci. Adv. Am. Assoc. Adv. Sci. (AAAS) 2018, 4, eaat0496. [Google Scholar] [CrossRef]
- Batistuti Sawazaki, M.R.; Bachour Junior, B.; Mulato, M.; de Oliveira Filho, J.I. Machine Learning-Assisted Infectious Disease Detection in Low-Income Areas: Toward Rapid Triage of Dengue and Zika Virus Using Open-Source Hardware. Adv. Intell. Discov. 2025. [Google Scholar] [CrossRef]
- Abu-Mostafa, Y.; Magdon-Ismail, M.; Lin, H.T. Learning from Data: A Short Course; AMLBook: Pasadena, CA, USA, 2012. [Google Scholar]
- Martinez, A.; Kak, A. PCA versus LDA. IEEE Trans. Pattern Anal. Mach. Intell. 2001, 23, 228–233. [Google Scholar] [CrossRef]
- Teng, C.H.; Hsu, W.H.; Busey, T.; Schneider, B.; Moses, K.; Zheng, W.S.; Lai, J.H.; Yuen, P.; Aleksic, P.; Wagner, M.; et al. LDA (Linear Discriminant Analysis). In Encyclopedia of Biometrics; Springer: Berlin/Heidelberg, Germany, 2009; pp. 1–899. [Google Scholar] [CrossRef]
- Murphy, K.P. Machine Learning: A Probabilistic Perspective; MIT Press: Cambridge, MA, USA, 2013. [Google Scholar] [CrossRef]
- Bishop, C.M. Pattern Recognition and Machine Learning (Information Science and Statistics); Springer: Berlin/Heidelberg, Germany, 2006; pp. 1–738. [Google Scholar] [CrossRef]
- Cristianini, N.; Shawe-Taylor, J. An Introduction to Support Vector Machines and Other Kernel-based Learning Methods; Cambridge University Press: Cambridge, UK, 2000. [Google Scholar]
- Boser, B.E.; Guyon, I.M.; Vapnik, V.N. A Training Algorithm for Optimal Margin Classifiers. In Proceedings of the 5th Annual Workshop on Computational Learning Theory (COLT’92), Pittsburgh, PA, USA, 27–29 July 1992; pp. 144–152. [Google Scholar]
- Takahashi, A.; Flores, E.A.; Maciel-de-Freitas, R.; Kariyawasam, T.; Sikulu-Lord, M.T. Comparative Evaluation of a Portable and Benchtop Near-Infrared Spectrometers for Characterizing the Age and Blood Feeding History of Aedes aegypti Mosquitoes in the Lab. Preprints 2025, 2025081494. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dataset | 4 DPI | 7 DPI | 10 DPI | |||
|---|---|---|---|---|---|---|
| TPR (%) | SPC (%) | TPR (%) | SPC (%) | TPR (%) | SPC (%) | |
| C1TR | 83.3 | 96.8 | 93.5 | 96.4 | - | - |
| C1VS | 100.0 | 94.1 | 100.0 | 100.0 | - | - |
| C2HT | 98.7 | 98.3 | 100.0 | 98.3 | 100.0 | 86.7 |
| C2AB | 98.7 | 85.0 | 96.2 | 80.0 | 97.4 | 68.3 |
| Dataset | TPR (%) | SPC (%) | PRC (%) | ACC (%) | F1S (%) |
|---|---|---|---|---|---|
| 4 DPI | |||||
| C1TR | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 |
| C1VS | 100.0 ± 0.0 | 93.5 ± 10.3 | 96.5 ± 6.3 | 97.6 ± 4.2 | 98.1 ± 3.5 |
| C2HT | 100.0 | 86.4 | 90.5 | 94.1 | 95.0 |
| C2AB | 100.0 | 95.0 | 96.2 | 97.8 | 98.1 |
| 7 DPI | |||||
| C1TR | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 |
| C1VS | 100.0 ± 0.0 | 93.7 ± 9.6 | 94.8 ± 7.9 | 96.9 ± 4.8 | 97.2 ± 4.3 |
| C2HT | 100 | 93.2 | 95.1 | 97.1 | 97.5 |
| C2AB | 94.8 | 100 | 100 | 97.1 | 97.3 |
| 10 DPI | |||||
| C1TR | - | - | - | - | - |
| C1VS | - | - | - | - | - |
| C2HT | 98.7 | 98.3 | 98.7 | 98.5 | 98.7 |
| C2AB | 94.8 | 100 | 100 | 97.1 | 97.3 |
| No DPI Stratification (%) | |||||
|---|---|---|---|---|---|
| Dataset | TPR (%) | SPC (%) | PRC (%) | ACC (%) | F1S (%) |
| C1TR | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 |
| C1VS | 100.0 ± 0.0 | 91.8 ± 7.2 | 95.3 ± 4.3 | 96.9 ± 2.8 | 97.5 ± 2.2 |
| C2HT | 99.6 | 92.7 | 94.6 | 96.6 | 97.0 |
| C2AB | 98.3 | 96.7 | 97.4 | 97.6 | 97.8 |
| Range (nm) | TPR (%) | SPC (%) | PRC (%) | ACC (%) | F1S (%) |
|---|---|---|---|---|---|
| Cohort 2 Head/Thorax | |||||
| 350–1000 | 99.6 | 92.7 | 94.6 | 96.6 | 97.0 |
| 1001–1800 | 90.4 | 41.6 | 66.7 | 69.1 | 76.8 |
| 1801–2500 | 98.3 | 36.0 | 66.5 | 71.1 | 79.3 |
| 400–750 (VIS) | 93.5 | 88.2 | 91.1 | 91.2 | 92.3 |
| Cohort 2 Abdomen | |||||
| 350–1000 | 98.3 | 96.7 | 97.4 | 97.6 | 97.8 |
| 1001–1800 | 84.3 | 37.8 | 63.4 | 63.9 | 72.4 |
| 1801–2500 | 97.4 | 52.2 | 72.3 | 77.6 | 83.0 |
| 400–750 (VIS) | 97.4 | 75.6 | 83.6 | 87.8 | 90.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reigoto, L.; Maciel-de-Freitas, R.; Sikulu-Lord, M.T.; Garcia, G.A.; Araujo, G.; Lima, A. Limited Spectroscopy Data and Machine Learning for Detection of Zika Virus Infection in Aedes aegypti Mosquitoes. Trop. Med. Infect. Dis. 2025, 10, 308. https://doi.org/10.3390/tropicalmed10110308
Reigoto L, Maciel-de-Freitas R, Sikulu-Lord MT, Garcia GA, Araujo G, Lima A. Limited Spectroscopy Data and Machine Learning for Detection of Zika Virus Infection in Aedes aegypti Mosquitoes. Tropical Medicine and Infectious Disease. 2025; 10(11):308. https://doi.org/10.3390/tropicalmed10110308
Chicago/Turabian StyleReigoto, Leonardo, Rafael Maciel-de-Freitas, Maggy T. Sikulu-Lord, Gabriela A. Garcia, Gabriel Araujo, and Amaro Lima. 2025. "Limited Spectroscopy Data and Machine Learning for Detection of Zika Virus Infection in Aedes aegypti Mosquitoes" Tropical Medicine and Infectious Disease 10, no. 11: 308. https://doi.org/10.3390/tropicalmed10110308
APA StyleReigoto, L., Maciel-de-Freitas, R., Sikulu-Lord, M. T., Garcia, G. A., Araujo, G., & Lima, A. (2025). Limited Spectroscopy Data and Machine Learning for Detection of Zika Virus Infection in Aedes aegypti Mosquitoes. Tropical Medicine and Infectious Disease, 10(11), 308. https://doi.org/10.3390/tropicalmed10110308