
Winter Ecology of Agricultural Birds: The Role of Crop Type in Habitat Occupation by the Eurasian Blackcap



Abstract
1. Introduction
2. Material and Methods
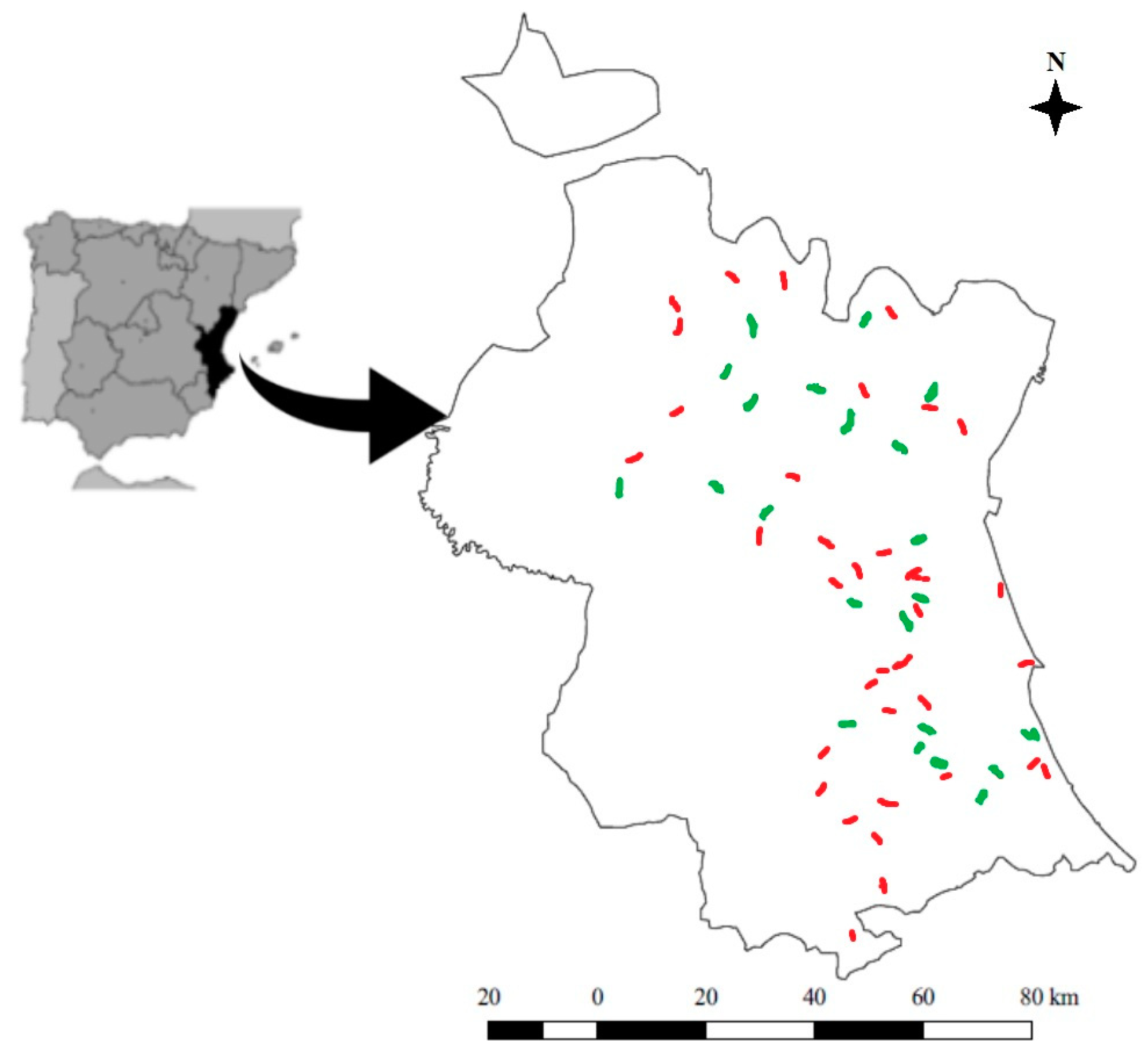
2.1. Study Area
2.2. Bird Sampling
2.3. Data Analysis
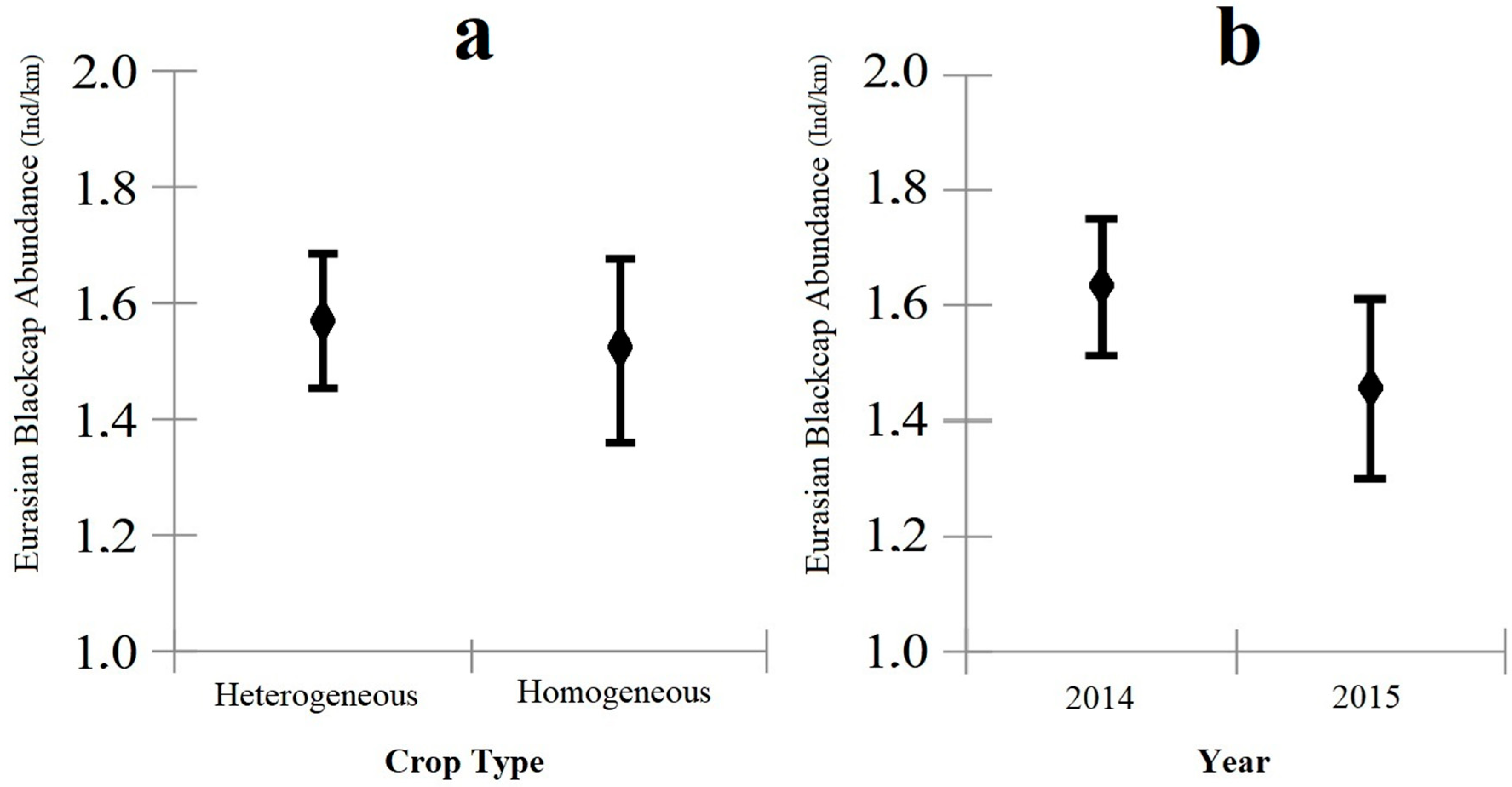
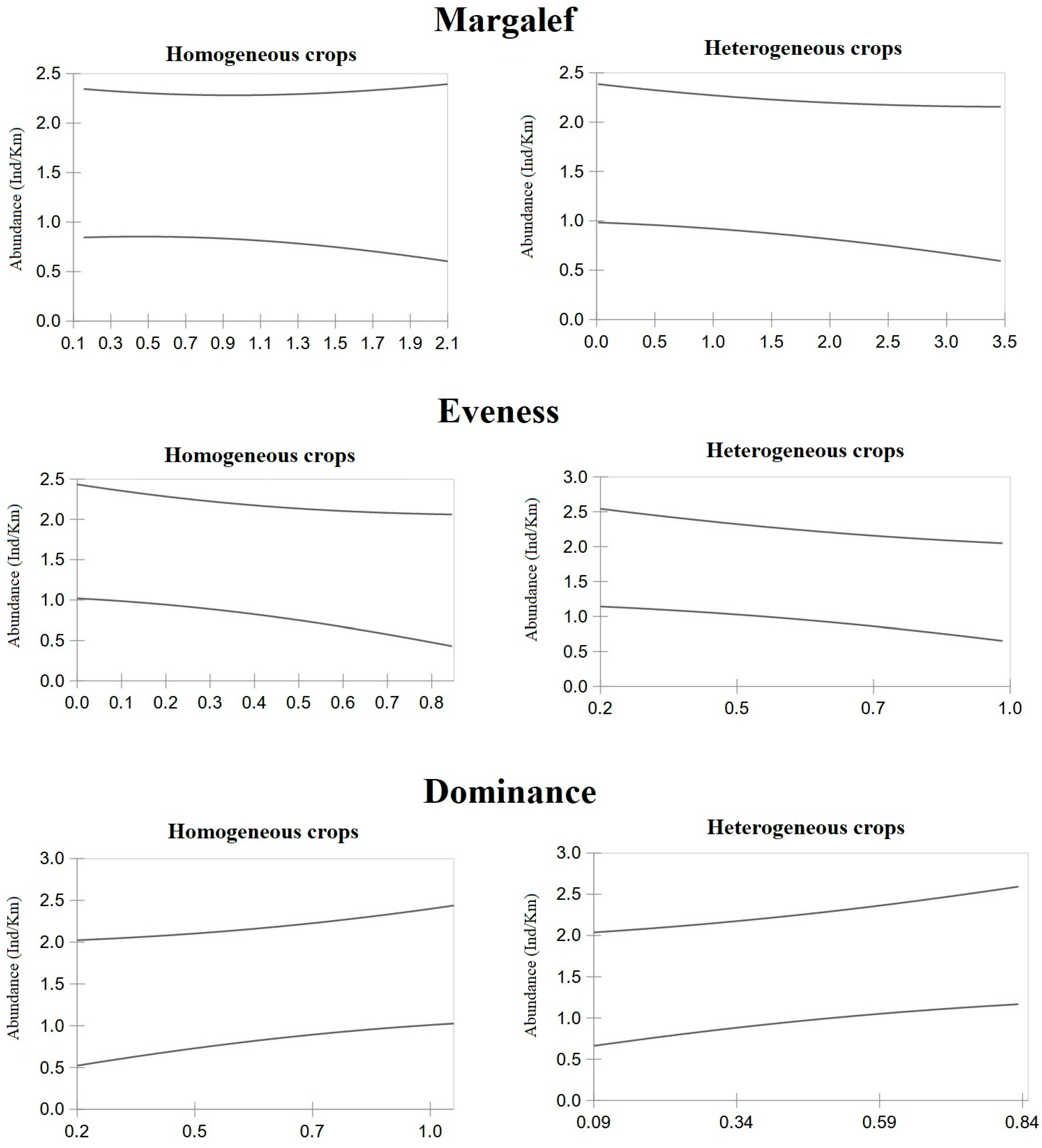
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jokimäki, J.; Clergeau, P.; Kaisanlahti-Jokimäki, M.L. Winter bird communities in urban habitats: A comparative study between central and northern Europe. J. Biogeogr. 2002, 29, 69–79. [Google Scholar] [CrossRef]
- Diaz, M.; Tellería, J. Predicting the effects of agricultural changes in central Spanish croplands on seed-eating overwintering birds. Agric. Ecosyst. Environ. 1994, 49, 289–298. [Google Scholar] [CrossRef]
- Tellería, J.L.; Santos, T. Effects of forest fragmentation on a guild of wintering passerines: The role of habitat selection. Biol. Conserv. 1995, 71, 61–67. [Google Scholar] [CrossRef]
- Senar, J.C.; Borrás, A. Sobrevivir al invierno: Estratégias de las aves invernantes en la Península Ibérica. Ardeola 2004, 51, 133–168. [Google Scholar]
- SEO/BirdLife. Atlas de las aves en invierno en España 2007–2010; Ministerio de Agricultura, Alimentacion y Medio Ambiente-SEO/BirdLife: Madrid, Spain, 2012. [Google Scholar]
- Pérez-Tris, J.; Carbonell, R.; Tellería, J.L. A method for differentiating between sedentary and migratory Blackcaps Sylvia atricapilla in wintering areas of southern Iberia. Bird Study 1999, 46, 299–304. [Google Scholar] [CrossRef]
- Santos, T.; Tellería, J.L. Patrones generales de la distribución invernal de Passeriformes en la Península Ibérica. Ardeola 1985, 32, 17–30. [Google Scholar]
- Carrascal, L.M.; Villén-Pérez, S.; Palomino, D. Preferred temperature and thermal breadth of birds wintering in peninsular Spain: The limited effect of temperature on species distribution. PeerJ 2016, 4, e2156. [Google Scholar] [CrossRef]
- Santos, M.R.D.L.; Cuadrado, M.; Arjona, S. Variation in the abundance of Blackcaps (Sylvia atricapilla) wintering in an olive (Olea europaea) orchard in southern Spain. Bird Study 1986, 33, 81–86. [Google Scholar] [CrossRef]
- Cantos, F.J. Análisis de los factores asociados a la invernada de los Sílvidos Presaharianos en la Península Ibérica en base a los resultados del anillamiento científico. Ecología 1996, 10, 429–436. [Google Scholar]
- Carrascal, L.M.; Villén-Pérez, S.; Seoane, J. Thermal, food and vegetation effects on winter bird species richness of Mediterranean oakwoods. Ecol. Res. 2012, 27, 293–302. [Google Scholar] [CrossRef]
- Tellería, J.L.; Fernández-López, J.; Fandos, G. Effect of climate change on mediterranean winter ranges of two migratory passerines. PLoS ONE 2016, 11, e0146958. [Google Scholar] [CrossRef]
- Katuwal, H.B.; Rai, J.; Tomlinson, K.; Rimal, B.; Sharma, H.P.; Baral, H.S.; Quan, R.C. Seasonal variation and crop diversity shape the composition of bird communities in agricultural landscapes in Nepal. Agric. Ecosyst. Environ. 2022, 333, 107973. [Google Scholar] [CrossRef]
- Gonnerman, M.; Shea, S.A.; Sullivan, K.; Kamath, P.; Overturf, K.; Blomberg, E. Dynamic winter weather moderates movement and resource selection of wild turkeys at high-latitude range limits. Ecol. Appl. 2023, 33, e2734. [Google Scholar] [CrossRef] [PubMed]
- Rey, P.J. Preserving frugivorous birds in agro-ecosystems: Lessons from Spanish olive orchards. J. Appl. Ecol. 2011, 48, 228–237. [Google Scholar] [CrossRef]
- Myczko, Ł.; Rosin, Z.M.; Skórka, P.; Wylegała, P.; Tobolka, M.; Fliszkiewicz, M.; Tryjanowski, P. Effects of management intensity and orchard features on bird communities in winter. Ecol. Res. 2013, 28, 503–512. [Google Scholar] [CrossRef]
- Wuczyński, A.; Wuczyński, M. Birds wintering in heterogeneous farmland of Poland: Weather-dependent temporal changes in abundance and habitat associations. Acta Ornithol. 2019, 54, 105–124. [Google Scholar] [CrossRef]
- Moreno-Mosquera, E.A.; Drechsler, R.M.; Monrós, J.S. The effect of vegetation structure on seasonal density of Sylvia warblers in the eastern Iberian Peninsula. Bird Study 2021, 68, 112–121. [Google Scholar] [CrossRef]
- Gainzarain, J.A. Selección de hábitat de la avifauna en una comarca agrícola del Alto Valle del Ebro (Norte de España). Munibe Ciencias Nat. 1996, 48, 3–16. [Google Scholar]
- Dietzen, C.; Edwards, P.J.; Wolf, C.; Ludwigs, J.D.; Luttik, R. Focal species of birds in European crops for higher tier pesticide risk assessment. Integr. Environ. Assess. Manag. 2013, 10, 247–259. [Google Scholar] [CrossRef]
- Santos, T.; Carbonell, R.; Galarza, A.; Perez-Tris, J.; Ramirez, A.; Tellería, J.L. The importance of northern Spanish farmland for wintering migratory passerines: A quantitative assessment. Bird Conserv. Int. 2014, 24, 1–16. [Google Scholar] [CrossRef]
- Chiatante, G.; Meriggi, A. The Importance of Rotational Crops for Biodiversity Conservation in Mediterranean Areas. PLoS ONE 2016, 11, e0149323. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.B.; Goodale, E. Crop heterogeneity and non-crop vegetation can enhance avian diversity in a tropical agricultural landscape in southern China. Agric. Ecosyst. Environ. 2018, 265, 254–263. [Google Scholar] [CrossRef]
- Tellería, J.L.; Santos, T.; Álvarez, G.; Sáez-Royuela, C. Avifauna de los campos de cereales del interior de España. Monografías. SEO 1988, 2, 173–319. [Google Scholar]
- Castro-Caro, J.C.; Barrio, I.C.; Tortosa, F.S. Is the effect of farming practices on songbird communities landscape dependent? A case study of olive groves in southern Spain. J. Ornithol. 2013, 155, 357–365. [Google Scholar] [CrossRef]
- Morales, M.B.; Guerrero, I.; Oñate, J.J. Efectos de la gestión agraria en las aves de los cultivos cerealistas: Un proceso multiescalar. Rev. Ecosistemas 2013, 22, 25–29. [Google Scholar]
- Peisley, R.K.; Saunders, M.E.; Luck, G.W. A systematic review of the benefits and costs of bird and insect activity in agroecosystems. Springer Sci. Rev. 2015, 3, 113–125. [Google Scholar] [CrossRef]
- Assandri, G.; Bogliani, G.; Pedrini, P.; Brambilla, M. Diversity in the monotony? Habitat traits and management practices shape avian communities in intensive vineyards. Agric. Ecosyst. Environ. 2016, 223, 250–260. [Google Scholar] [CrossRef]
- Martinelli, S.; Battisti, C. A simplified breeding bird community along remnant riparian woodpatches in an agricultural landscape (central Italy). ALULA 2021, 28, 120–123. [Google Scholar]
- Pérez, C.; Acebes, P.; Franco, L.; Llusia, D.; Morales, M.B. Olive grove intensification negatively affects wintering bird communities in central Spain. Basic Appl. Ecol. 2023, 70, 27–37. [Google Scholar] [CrossRef]
- Rime, Y.; Luisier, C.; Arlettaz, R.; Jacot, A. Landscape heterogeneity and management practices drive habitat preferences of wintering and breeding birds in intensively-managed fruit-tree plantations. Agric. Ecosyst. Environ. 2020, 295, 106890. [Google Scholar] [CrossRef]
- Busse, P. The European Autumn Migration Pattern of the Blackcap, Sylvia atricapilla–From Faunistic Observations to a Basic Analysis of Orientation Cage Field Data. Ring 2021, 43, 3–39. [Google Scholar] [CrossRef]
- Losada, M.; Salaverri, L.; Docampo, M.; Guitián, J.; Sobral, M. Bird communities after 37 years in a rural area of NW Spain. Nova Acta Científica Compostel. 2023, 30, 1–17. [Google Scholar] [CrossRef]
- Quintero, E.; Arroyo, J.M.; Dirzo, R.; Jordano, P.; Rodríguez-Sánchez, F. Lasting effects of avian-frugivore interactions on seed dispersal and seedling establishment. J. Ecol. 2024, 112, 656–672. [Google Scholar] [CrossRef]
- Berthold, P.; Terrill, S.B. Migratory behaviour and population growth of Blackcaps wintering in Britain and Ireland: Some hypotheses. Rining Migr. 1988, 9, 153–159. [Google Scholar] [CrossRef]
- Caula, S.; Marty, P.; Martin, J.L. Seasonal variation in species composition of an urban bird community in Mediterranean France. Landscape Urban Plan. 2008, 87, 1–9. [Google Scholar] [CrossRef]
- Caula, S.; de Villalobos, A.E.; Marty, P. Seasonal dynamics of bird communities in urban forests of a Mediterranean city (Montpellier, Southern France). Urban Ecosyst. 2014, 17, 11–26. [Google Scholar] [CrossRef]
- Šálek, M.; Kalinová, K.; Reif, J. Conservation potential of semi-natural habitats for birds in intensively-used agricultural landscapes. J. Nat Conserv. 2022, 66, 126124. [Google Scholar] [CrossRef]
- Murgui, E. Seasonal patterns of bird crop use in traditional irrigated land (the Huerta Of Valencia, Spain). Ardeola 2010, 57, 87–102. [Google Scholar]
- Tellería, J.L.; Pérez-Tris, J. Seasonal distribution of a migratory bird: Effects of local and regional resource tracking. J. Biogeogr. 2003, 30, 1583–1591. [Google Scholar] [CrossRef]
- Ożarowska, A.; Meissner, W. Increasing body condition of autumn migrating Eurasian blackcaps Sylvia atricapilla over four decades. Eur. Zool. J. 2024, 91, 151–161. [Google Scholar] [CrossRef]
- Villalva, P.; Quintero, E.; Isla, J.; Jordano, P. Fleshy-Fruit Consumption in Doñana. Bull. Ecol. Soc. Am. 2025, 106, e02197. [Google Scholar] [CrossRef]
- Rey, P.J. Spatio-Temporal Variation in Fruit and Frugivorous Bird Abundance in Olive Orchards. Ecology 1995, 76, 1625–1635. [Google Scholar] [CrossRef]
- Moreno-Mosquera, E.A.; Drechsler, R.M.; Rivera, S.; Monrós, J.S. Breeding habitat selection of three sympatric species of Mediterranean warblers in eastern Spain. Avian Biol. Res. 2024, 17, 3–11. [Google Scholar] [CrossRef]
- Bibby, C.J.; Burgess, N.D.; Hill, D.A. Bird Census Techniques; Academic Press: London, UK, 1992. [Google Scholar]
- Matteucci, S.D.; Colma, A. Metodología Para El Estudio de la Vegetación; Secretaría General de la Organización de los Estados Americanos. Programa Regional de Desarrollo Científico y Tecnológico: Washington, WA, USA, 1982. [Google Scholar]
- Bonham, C.D. Measurements for Terrestrial Vegetation; Wiley-Interscience: New York, NY, USA, 1989. [Google Scholar]
- Mostacedo, B.; Fredericksen, T. Manual de Métodos Básicos de Muestreo y Análisis en Ecología Vegetal; Proyecto de Manejo Forestal Sostenible (BOLFOR): Santa Cruz, Bolivia, 2000. [Google Scholar]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. PAST:paquete de programas de estadística paleontológica para enseñanza y análisis de datos. Palaeontol Electron 2001, 4, 1. [Google Scholar]
- Papadimitriou, F. Modelling indicators and indices of landscape complexity: An approach using GIS. Ecol. Indic. 2002, 2, 17–25. [Google Scholar] [CrossRef]
- XLSTAT. XLSTAT-Ecology Software, version 5; Copyright Addinsoft 1995–2017. 2017. Available online: https://www.xlstat.com (accessed on 25 June 2018).
- Belda, E.J.; Barba, E.; Monrós, J.S. Resident and transient dynamics, site fidelity and survival in wintering Blackcaps Sylvia atricapilla: Evidence from capture-recapture analyses. Ibis 2007, 149, 396–404. [Google Scholar] [CrossRef]
- Saunders, S.P.; Meehan, T.D.; Michel, N.L.; Bateman, B.L.; DeLuca, W.; Deppe, J.L.; Wilsey, C.B. Unraveling a century of global change impacts on winter bird distributions in the eastern United States. Glob. Change Biol. 2022, 28, 2221–2235. [Google Scholar] [CrossRef]
- Sapir, N.; Abramsky, Z.; Shochat, E.; Izhaki, I. Scale-dependent habitat selection in migratory frugivorous passerines. Naturwissenschaften 2004, 91, 544–547. [Google Scholar] [CrossRef]
- Concepción, E.D.; Díaz, M.; Baquero, R.A. Effects of landscape complexity on the ecological effectiveness of agri-environment schemes. Landsc. Ecol. 2008, 23, 135–148. [Google Scholar] [CrossRef]
- Van Doren, B.M.; Conway, G.J.; Phillips, R.J.; Evans, G.C.; Roberts, G.C.; Liedvogel, M.; Sheldon, B.C. Human activity shapes the wintering ecology of a migratory bird. Glob. Change Biol. 2021, 27, 2715–2727. [Google Scholar] [CrossRef]
- Lehnardt, Y.; Sapir, N. Redistribution of songbirds within a migratory stopover site as a response to sylviid warbler song playback. Ibis 2024, 166, 1184–1198. [Google Scholar] [CrossRef]
- Berthold, P. Microevolution of migratory behaviour illustrated by the Blackcap Sylvia atricapilla: 1993 Witherby Lecture. Bird Study 1995, 42, 89–100. [Google Scholar] [CrossRef]
- Bruggisser, O.T.; Schmidt-Entling, M.H.; Bacher, S. Effects of vineyard management on biodiversity at three trophic levels. Biol. Conserv. 2010, 143, 1521–1528. [Google Scholar] [CrossRef]
- Johnson, M.D.; Sherry, T.W.; Holmes, R.T.; Marra, P.P. Assessing habitat quality for a migratory songbird wintering in natural and agricultural habitats. Conserv. Biol. 2006, 20, 1433–1444. [Google Scholar] [CrossRef] [PubMed]
- Rey, P.J.; Gutiérrez, J.E. Pecking of olives by frugivorous birds: A shift in feeding behaviour to overcome gape limitation. J. Avian Biol. 1996, 27, 327–333. [Google Scholar] [CrossRef]
- Hernandez, A. Cherry removal by seed-dispersing mammals: Mutualism through commensal association with frugivorous birds. Pol. J. Ecol. 2008, 56, 127–138. [Google Scholar]
- Pérez-Tris, J.; Tellería, J.L. Migratory and sedentary blackcaps in sympatric non-breeding grounds: Implications for the evolution of avian migration. J. Anim. Ecol. 2002, 71, 211–224. [Google Scholar] [CrossRef]
- Hera, I.D.L.; Pérez-Tris, J.; Tellería, J.L. Habitat distribution of migratory and sedentary blackcaps Sylvia atricapilla wintering in southern Iberia: A morphological and biogeochemical approach. J. Avian Biol. 2012, 43, 333–340. [Google Scholar] [CrossRef]
- Morganti, M.; Assandri, G.; Aguirre, J.I.; Ramirez, Á.; Caffi, M.; Pulido, F. How residents behave: Home range flexibility and dominance over migrants in a Mediterranean passerine. Anim. Behav. 2017, 123, 293–304. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Variables | Occupied (N = 45) | Non-Occupied (N = 15) | t | df | p |
|---|---|---|---|---|---|
| Coverage | 42.8 ± 24.5 | 37.9 ± 19.3 | −0.71 | 58 | 0.48 |
| Richness | 0.98 ± 0.01 | 1.01 ± 0.04 | 0.19 | 57 | 0.85 |
| Dominance | 0.48 ± 0.00 | 0.47 ± 0.01 | 0.13 | 57 | 0.89 |
| Evenness | 0.85 ± 0.00 | 0.91 ± 0.01 | 0.7 | 54 | 0.48 |
| U | p | ||||
| Height | 2.28 ± 0.87 | 1.93 ± 0.79 | 265 | 0.22 | |
| Altitude | 181 ± 3.86 | 438 ± 19.9 | 499.5 | 0.001 |
| Homogeneous | Heterogeneous | t | df | p | |
|---|---|---|---|---|---|
| Margalef | 0.74 ± 0.03 | 1.11 ± 0.02 | 1.74 | 43 | 0.09 |
| Evenness | 0.57 ± 0.02 | 1.04 ± 0.01 | 3.24 | 43 | <0.01 |
| Dominance | 0.67 ± 0.01 | 0.42 ± 0.00 | −4.51 | 43 | <0.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moreno-Mosquera, E.A.; Drechsler, R.M.; Monrós, J.S. Winter Ecology of Agricultural Birds: The Role of Crop Type in Habitat Occupation by the Eurasian Blackcap. Sci 2025, 7, 57. https://doi.org/10.3390/sci7020057
Moreno-Mosquera EA, Drechsler RM, Monrós JS. Winter Ecology of Agricultural Birds: The Role of Crop Type in Habitat Occupation by the Eurasian Blackcap. Sci. 2025; 7(2):57. https://doi.org/10.3390/sci7020057
Chicago/Turabian StyleMoreno-Mosquera, Eyda Annier, Robby M. Drechsler, and Juan S. Monrós. 2025. "Winter Ecology of Agricultural Birds: The Role of Crop Type in Habitat Occupation by the Eurasian Blackcap" Sci 7, no. 2: 57. https://doi.org/10.3390/sci7020057
APA StyleMoreno-Mosquera, E. A., Drechsler, R. M., & Monrós, J. S. (2025). Winter Ecology of Agricultural Birds: The Role of Crop Type in Habitat Occupation by the Eurasian Blackcap. Sci, 7(2), 57. https://doi.org/10.3390/sci7020057