Abstract
This study explored whether consistent differences can be found between early-wood and late-wood in terms of elemental content of tree rings. The species to study was Pinus sylvestris L. growing within an even-aged stand planted during the early 1970s in eastern Siberia. The wood specimens were extracted from the north and south sides of trees and subsequently scanned through an X-ray fluorescent facility Itrax Multiscanner. A sequence of relatively wide tree-rings was chosen for the analysis. The scanning data on a number of elements (Al, Si, P, S, Cl, K, Ca, Ti, Mn, Fe, Cu, Zn, Sr, and Hg) were split into early-wood and late-wood data for each year of growth. The early- and late-wood data in the same ring were analyzed for basic statistics against each other as well as against available meteorological data. In the northern direction, the elements Al, Si, P, Cl, Cu, and Zn are always more abundant in the late-wood, while Ca, Fe, and Sr are always more abundant in the early-wood. What is important is how the differences for P, Ca, Fe, Cu, Zn, and Sr were always significant. The calcium content in the early-wood was the most consistently reflective regarding the meteorological data for the early summer (June). In some trees, the late-wood K content was well correlated with the Vysotskii–Ivanov climatic index. In the southern direction, Cu and Zn were always more abundant in the late-wood, while Sr was more abundant in the early-wood. The differences for all three elements were always significant. The cases of consistent relationships, though rare, help to develop a research program in the area of dendrochemistry.
1. Introduction
The studies of tree rings have been a powerful instrument in environmental research due to the tree rings’ ability to provide annual resolution of data and long data sequences. While the tree ring width as well as the cell morphology and wood density served to be primary parameters of the rings in dendrochronology, since the 1970s–1980s, studies of element concentration in trees have been developing by the area of environmental chemistry. A combination of tree ring analysis and elemental analysis have given a new perspective to environmental research [1].
Due to obvious practical importance, the dendrochemical tree ring studies have often concentrated on the emissions of pollutants as a result of industrial activities [2,3,4,5,6,7,8]. This area of inquiry has a clear significance for the practice of legal actions against subjects committing pollution.
Another developing research area deals with relationships of elemental content in tree rings and the factors of physical environment, which includes events from ancient volcano eruptions to actual weather conditions. Hevia et al. [9] have found that a number of elements peaked around the years of known eruptions in tree rings in Pyrenean Mountain pine. Also, the authors related the elemental variations to temperature and precipitation factors. Sometimes, to reveal the relations with weather parameters, one needs to subject the wood samples to additional treatment. Scharnweber et al. [10] suggested to extract the wooden specimen in a number of solvents (water, alcohol, and acetone) to separate the elements tightly bound in wood from volatile substances inducing noise into the environmental signal. Gavrikov et al. [11] used some other chemical agents (alcohol and hydrochloric acid) and found that only after the treatment were correlations of elemental content with meteorological parameters able to be found in some species (esp., in Siberian larch).
A complication—worthy of attention for both practical and theoretical research—is that sufficient variations of inorganics’ content are observed among and within species [12]. Thus, this variety may obscure the relationships sought. A question has been inspected whether the elemental sequences obtained from different sides of the same Scots pine tree correlate with each other [13]. The inference was that the sequences of the majority of expected elements in sequences from different trunk sides did not correlate, with a couple of exclusions which are Ca and K having long trends across tree trunks. Consequently, an apparent lack of consistency may affect the reliability of a research outcome.
It is well known since long ago that the growth of tree rings pursues a definite seasonal pattern in temperate tree species. Typically, the sequences of cells in a tree ring are subdivided into early-wood, with larger lumens and thinner cell walls, and late-wood, with narrower lumens and thicker cell walls. Such a pattern, despite the differences in wood structure, is characteristic of both conifers and dicots among trees [14,15,16]. However, as Vaganov et al. [1] pointed out, the intrinsic factors leading to different rates and durations of the tracheid formation and their specific morphology remain poorly understood. Not only are physical wood properties different within a single tree ring but the early- and late-wood also bear biochemical and chemical differences.
For example, the late-wood in Norway spruce contained significantly more galactomannan and less pectins than the early-wood [17]. In the same species, the relative-to-cellulose content of lignin is significantly higher in the early-wood which was shown with the help of Raman microspectroscopy and infrared spectroscopy [18]. The early-wood vessels were rich in guaiacyl lignin while the vessels of late-wood were rich in syringyl lignin in English oak [19]. Antonova et al. [20] have shown that syringyl and p-hydroxiphenil aromatic residues prevailed in the course of early xylem maturing in Scots pine. At the same time, the maturing xylem of the late-wood contained an increased share of guaiacyl residues.
Regarding the inorganics, the studies of early- and late-wood are rather scarce. Hevia et al. [9] reported on differences in elemental contents between early-wood and late-wood in Mountain pine, including especially how Ca was found to peak in the early-wood after which Ca was decreasing closer to the late-wood.
Our experience shows that a lack of consistency in the chemical effects found in tree rings may be a serious obstacle for progress in this research area. The effects may be obvious in one sample/tree and absent in other samples/trees. Thus, the primary purpose in the field should be the establishment of relationships between elements and the anatomical structures of wood as well as among the elements residing in the xylem.
The goals of this research were as follows: (a) to find, among a number of elements, relationships between early- and late-wood which were consistent enough within tree and among trees; and (b) to find consistent correlations between elemental content in early- and late-wood, on the one hand, and available meteorological parameters, on the other hand.
2. Materials and Methods
2.1. Geographical Location and Species
For this study, an even-aged Scots pine (Pinus sylvestris L.) stand was taken. The trees were planted as 2–3-year-old seedlings in 1971–1972 as a part of an afforestation experiment with an initial planting density of 40,000 saplings per ha [21,22]. The location of the experiment area is shown in Figure 1.
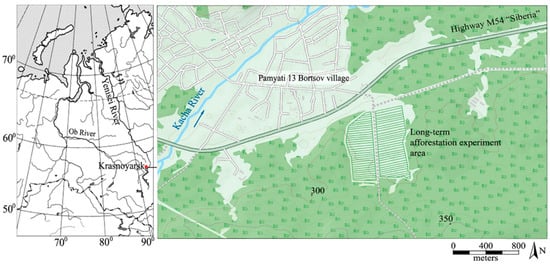
Figure 1.
Geographical location of the study area. The blue arrow is the direction of the river stream.
2.2. Sampling
In autumn 2021, six apparently healthy pines of ca. average size were randomly chosen for the sampling. From each tree, two 12 mm cores were extracted at breast height with the help of a Haglöf borer (Haglöf Sweden AB, Långsele, Sweden), with one from the southern and one from the northern sides of the tree stem. Altogether, 12 cores were sampled and further treated. Under room conditions, the cores were air-dried in a laboratory. With the help of a circular saw, 1.2 mm thick slices were made from the cores perpendicular to the wood grain. The sampling and the subsequent treatment of the cores were performed in observance of basic approaches accepted in dendrochronology [23].
2.3. X-ray Scanning
X-ray facility Itrax Multiscanner (COX Analytical Systems AB, Mölndal, Sweden, 2012) coupled with Multi Scanner Navigator software 6.5.3. was used to scan the slices.
Itrax Multi Scanner excites the elements’ nuclei by a thin X-ray 20 mm by 50 µm beam and detects the energy of the responsive radiation. The radiation is registered by a Si-based detector SDD (Ketek GmbH, München, Germany). The resolution of the detector is 140 eV at 5.9 keV and it can register up to 200,000 photons a second (cps). The spatial resolution of scanning was 100 µm. The analysis of light elements at an acceptable detection limit is performed through an X-ray tube with a Cr anode (1.9 kWt). For Ti and heavier elements, an X-ray tube with Mo anode (3 kWt) is used.
Altogether, the chemical elements monitored in the research included Al, Si, P, S, Cl, K, Ca, Ti, Mn, Fe, Cu, Zn, Sr, and Hg. The reason to choose the elements was that most of the elements are either biologically active or may be biologically active. Also, a parameter physically describing the wood specimen was included which is the registered incoherent radiation, denoted as ‘inc’ in the subsequent tables.
For each element and for each scanning point, the multi-scanner produces a long series of data which are relative units (counts per second). The counts are theoretically related to the number of the element nuclei contained in the point of scanning.
In addition to the counts data, Itrax Multiscanner outputs X-ray images of the wood specimen allowing for sure recognition and dating of the tree rings.
2.4. Time Span of Data
An important detail is the time span included into the analysis. Because of the goals of the research, the widths of both early- and late-wood were crucial for the analysis. Still, the majority of outer tree rings are rather narrow, and so often a single, if any, scanning point falls within the late-wood thus making the measurements unrepresentative. At the same time, the studied rings should not be at the very pith.
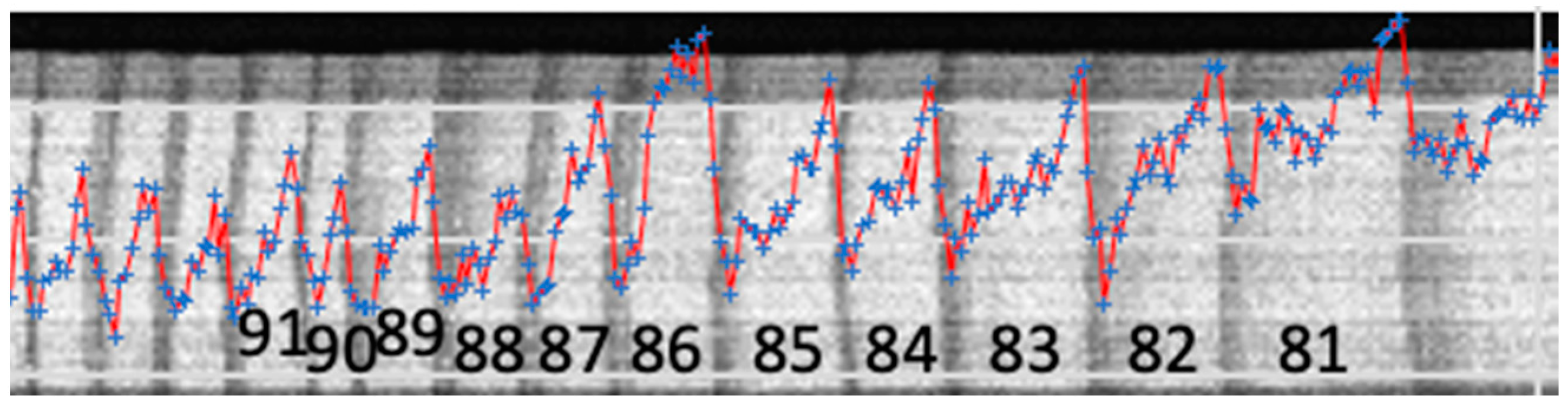
The last growth year was 2021 but only a row of tree rings that are wide enough were taken for the analysis. They are 11 years, from 1981 to 1991. An example image depicting the growth rings under study is shown in Figure 2.
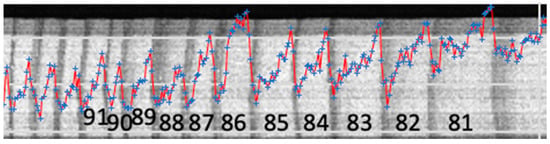
Figure 2.
A fragment of an X-ray image showing the time span (tree rings) included into the analysis. Black numbers from 81 to 91 denote years of the rings’ growth, i.e., from 1981 to 1991. The blue markers denote the levels of Ca count rates, while the red line connects them for visibility.
2.5. Meteorological Data
The meteorological dataset has been used in a previous study [11]. The initial source of the data is the database of the Krasnoyarsk Weather Station, WMO ID = 29570.
Twenty-one various weather parameters were used in the study: (1) mean monthly humidity (from May to August), (2) mean monthly temperature (from May to August), (3) monthly sum of precipitations (from June to August), (4) mean monthly wind speed (from May to August), and (5) a compound humidity index by Vysotskii–Ivanov (from June to August). The latter index has been represented in two forms, ‘as is’ and detrended, and thus is in the form of indices.
The humidity index (HI) is calculated as
where Pr denotes the monthly precipitation while Ep denotes the monthly evaporation.
The precipitation data for May has not been used because the month is transient between winter and summer, and the precipitations may be in solid form.
An overview of the meteorological data is given in the Supplement as ‘Supplementary data Table S1’ file.
2.6. Statistical Treatment
The early- and late-wood within a ring were visually separated on the X-ray images (e.g., as shown in Figure 2). The scanning point closest to the outer border of late-wood stripe was considered to be the end of the ring. The scanning point closest to the inner border of the late-wood stripe was considered to be the end of the early-wood.
Because all the tree rings differ in width a different number of scanning points falls within the early- and late-wood of the rings. To eliminate the impact of the ring width, mean count rates per a scanning point were used throughout the analysis below. That is, for each core and each element, the mean count rates per a scanning point in the early- and late-wood were calculated. Alternatively, for another part of the analysis, the mean count rates were calculated per a scanning point within each tree ring, for the early- and late-wood in that ring combined.
According to the goals of the research, two main statistical methods have been applied. First, this is the standard comparison of means. Because the number of scanning points in the late-wood were 40–80 depending on the tree, the non-parametrical Mann–Whitney U-test has been applied to compare the elements’ count rates in early-wood vs. late-wood. The non-parametrical approach allows one to avoid dealing with normality tests. All the early-wood data of a tree were compared to all the late-wood data of the same tree. Then, the results of all the trees were compared against each other. Second, possible relationships between the elements’ contents and the weather parameters have been estimated through a classical correlation analysis. Due to the small number of data points, a non-parametrical Spearman’s rank correlation has been applied. All the calculations have been performed with the help of R statistical tools [24].
3. Results
3.1. The Elemental Content in Early- vs. Late-Wood
Table 1 and Table 2 give an idea of how consistent the relations of elemental content in early- vs. late-wood are in practice. In both tables, ‘yes’ implies ‘always yes’ for all the trees studied. This means that if, for example, one sees ‘yes’ in the row Ca at the intersection with column ‘EW > LW’ (Table 1), then the Ca content is always larger in the early-wood than in the late-wood. By analogy, if ‘yes’ is found in the column ‘Significant’, then it means that the difference between early- and late-wood is always significant at p < 0.05, at least.

Table 1.
The results of a comparison of all the pines regarding the elemental content in early- vs. late-wood. Only northern cores are compared.
Table 2.
The results of a comparison of all the pines regarding the elemental content in early- vs. late-wood. Only southern cores are compared.
In Table 1 and Table 2, empty cells mean ‘not always’. The elemental difference between the early- and late-wood may be inconsistent, that is, larger in the early-wood in one tree and smaller in another. Because it is the consistencies that we would like to highlight, we did not specify the inconsistencies. Also, the elemental differences may be consistent but not always significant at p < 0.05, at least. A single p-value among the trees that is larger than 0.05 places the element case in the category ‘not always’. These latter cases are denoted as empty cells in the column ‘Significant’.
Table 1 and Table 2 show that there is a certain difference between the northern and the southern sides of the trees. This is not to say that North and South as such play any role in the distribution of the elements. In a sense, North and South just denote different sides of the tree trunk. However, the opposite, that is, that North and South as such do not exert a specific influence on the elemental distribution, is also not asserted.
In spite of the differences between the tables, there are still some consistencies. First of all, a number of elements (Cu, Zn, and Sr) show persistent relations between early- vs. late-wood and, importantly, these relations are always significant.
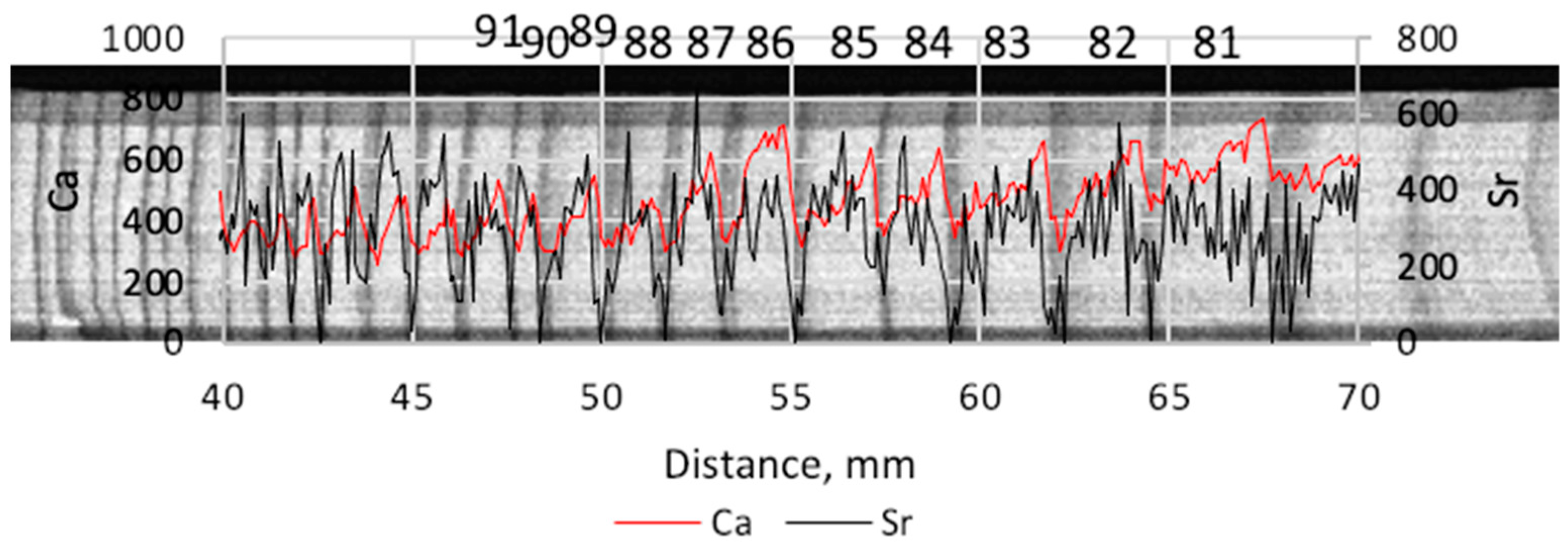
Another group of elements (Al, Si, P, Cl, Ca, and Fe) show persistent relations between the yearly- vs. late-wood but they are not always significant. An example illustrating the variations of Ca and Sr in one of the trees is depicted in Figure 3.
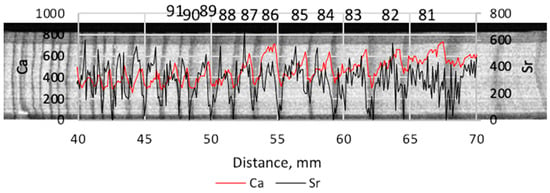
Figure 3.
An example of an X-ray image overlapped with the graph of counts of Ca and Sr in pine #3. The vertical axes give the dimensionless counts for Ca and Sr. The numbers from 81 to 91 denote years of the rings’ growth from 1981 to 1991.
3.2. The Elemental Content vs. Meteorological Data
Altogether, 630 paired combinations ‘early-/late-wood elemental content vs. meteorological parameter’ have been considered, that is, 30 tree ring parameters (elemental contents) by 21 meteorological parameters. As follows from the subsection ‘Time span of data’, the initial data are elemental contents in individual tree rings (separately for early- and late-wood) against meteorological data for the particular years from 1981 to 1991. Consequently, the number of observations in each correlation run was equal to 11.
It is noteworthy that among all the 630 paired combinations, only two elements, Ca and K, were found as well as a few meteorological parameters that more or less met the strict consistency criteria. The criteria are that the correlations in all the trees must be (i) of the same sign and (ii) not less than 0.3 or less than −0.3. The early-wood Ca against mean June humidity and temperature met the criteria.
Table 3 and Table 4 show the Spearman’s rank correlation coefficients for the early-wood Ca vs. the June humidity and temperature, correspondingly. It can be seen from the tables that there is a difference between the northern and southern sides of the tree trunk. While the coefficients in the northern side are rather high, the coefficients in the southern side are on average lower. The p-values are consequently much higher in the southern side while most correlations in the northern side are significant at p < 0.1, at least.
Table 3.
Correlation coefficients for Ca content in the early-wood vs. June humidity.
Table 4.
Correlation coefficients for Ca content in the early-wood vs. June temperature.
Another element showing a correlation with meteorological parameters is K. Table 5 gives the data on the correlations of the late-wood K with the June humidity index by Vysotskii-Ivanov. It can be seen from the table that the Spearman correlation is consistently positive. On the North side, the correlations are significant in half of the trees. On the Southern side, the Spearman coefficients are lower and the p-values are generally higher.
Table 5.
Correlation coefficients for K content in the late-wood vs. index by Vysotskii–Ivanov for June.
The result does not mean that other elements or meteorological parameters do not show sometimes high correlations. However, many of them are inconsistent in the sense that a high correlation in one tree may be coupled with a zero correlation in another. To be certain about that, it is enough to use the initial data supplied in the ‘Supplementary Data Table S1’ file.
4. Discussion
Similar to other scientific areas, the tree ring research is crucially dependent on the consistency factor. Controversial variations of measured values convert an anticipated relationship into a hardly explicable stochastic pattern. It has been found, for example, that there is a lack of correlations of elemental tree ring series among different sides of the tree trunk [13].
Dendrochronology—which has been based on relations between climatic factors and the tree ring width—finds a way out by averaging a large number of tree ring series and obtaining a population response. Another approach may be to collect relationships that are significant in all available specimen, and then to formulate a research program that is founded on this firm basis.
The anatomy of a temperate tree ring has been known since long ago. It typically consists of early- and late-wood stripes the organic chemistry of which has been recently explored from many respects [17,18,19,20]. Regarding the inorganics in early- vs. late-wood, there is still a lack of consistent evidence. Hevia et al. [9] have enumerated a number of results for Ca content in some conifers. Calcium mean content is higher in early-wood in Mountain pine, Siberian spruce, and Siberian fir, and inversely, Ca mean content is higher in late-wood vs. early-wood in bristlecone pine.
In this research, it has been found that, in addition to Ca, a number of important biophile elements may also show a consistent pattern between early- and late-wood. The core of the group is comprised by Cu, Zn, and Sr which always follow the same pattern in both tree trunk directions (North and South), and the differences between early- and late-wood are always significant. Phosphorus, calcium, and iron can also be considered as belonging to the group, although despite that their differences are always significant only in the northern trunk side but not in the southern one (Table 1 and Table 2). As follows from the ‘Time span of data’ subsection, the correlations were calculated with a limited number of data totaling 11 points. As the statistical significance is strongly dependent on the amount of data, the lack of significance may be related to the limited data. A larger amount of data (if they are available) may improve the problem of significance.
Like dendrochronology, dendrochemistry does not deal with alive cells but rather cells where the lumens are empty. Thus, the only place for the chemical elements to reside is the cell walls. It is well known that after cell death the cell walls remain rich in compound organic substances, such as lignins, pectins, pentosans, and others. All the organic molecules may contain active centers (like carboxyl groups –COOH or similar structures) which are able to exchange protons to metal ions flowing with the sap.
Because the organic speciation of early- and late-wood is to some extent dissimilar, diverse ions may be consistently attached to organic molecules in the alternative parts of the tree ring. Two elements, Cu and Zn, show a similar pattern, a persistent higher content in the late-wood vs. early-wood. The origin of such a behavior may be related to their atomic properties, that is, to the fact that the ionic radii of the elements are rather proximate. The ion Cu2+ has the radius 0.073 nm while Zn2+ has the radius 0.074 nm. Hypothetically, the elements may attach to active organic centers of the same kind which are more abundant in the late-wood.
Another remarkable pair of elements are Ca and Sr. They are considered to be geochemical analogs of each other because Sr often replaces Ca in minerals as well as in living tissues. The ion Ca2+ has the radius 0.1 nm while Sr2+ has the radius 0.118 nm which is somewhat larger but more or less close to the former to provide similar properties. Calcium variates consistently between early- and late-wood (Figure 3) growing in the very beginning of the growth season and falling around the start of late-wood formation. This type of behavior has been found in a number of studies [25,26].
The Ca content decrease in the late-wood is moderate. However, Sr drops radically in the late-wood (Figure 3), often down to “zero” values, which is probably the source of its steady significant difference between early- and late-wood. Because we deal with a facility that has its own technical limits of radiation detection, “zero” values are not obligatory real physical zero but a level that the facility percepts as the element absence.
Chemically, the absence may hypothetically mean that either Sr2+ loses out in a competition for active centers to Ca2+ or that Sr2+ just cannot attach to the active centers available in late-wood which may be due to a somewhat larger ionic radius.
The third opportunity is that Sr2+ is outcompeted by some other elements which are close to it in terms of ionic radii. A group of elements have ionic radii larger than Ca2+ but smaller than Sr2+. This group includes Ce3+ (0.101 nm), U3+ and Hg2+ (both 0.102 nm), and La3+ and Bi3+ (both 0.103 nm). However, all these elements are really very scarce in the mineral soil of the site. A preliminary estimation of the elements availability through ICP-AES (OES) atomic/optic emission spectroscopy (unpublished data) has shown that U and Hg are below the detection limit of the method, Bi is contained at 7.2, and Ce and La are contained at 53 and 32 mg/kg, correspondingly. To compare, Ca is found at 9864 and Sr is found at 166 mg/kg. Thus, it is unlikely that the group of scarce elements could influence the variations of Sr in the tree rings.
To summarize the differences in elemental contents in early- and late-wood, the consistent relationships may be a basis for an in-depth inquiry regarding why the elements are distributed in the wood this or that way. It is most probable that methods including extraction and re-scanning techniques [10,11] may help to penetrate into mechanisms of elemental sorption and distribution in tree rings.
Hevia et al. [9] reported on elemental peaks of Ca, Fe, and some other elements’ content in tree rings in the years following known large volcanic eruptions. The eruptions led to abrupt summer cooling in many global areas and, probably, impacted the absorption and the distribution of elements within tree rings. However, well-described inner mechanisms through which such elemental reactions appear are not yet known.
Nevertheless, if some definite relationships steadily replicate within and among trees, then these relationships may be used to do a next step in research. In this study, the pairs ‘early-wood Ca–June humidity’, ‘early-wood Ca–June temperature’, and ‘late-wood K–June humidity index’ showed a rather consistent pattern in the sense that Spearman’s coefficients were high enough for all the sampled trees. Although the p-values are somewhat higher than that accepted in natural sciences, this may be in part attributed to low data availability. There is a hope that the significance levels may increase with a longer series of wide tree rings.
It is worth paying attention to how the correlation of early-wood Ca with June humidity is negative while its correlation with June temperature is positive (Table 3 and Table 4). Generally, high air humidity should suppress transpiration in plants, especially at the time of active growth. If so, then the amount of sap flowing through the tree trunk and carrying the ions should be lower. On the other hand, higher temperatures should lead to increased transpiration and more sap flowing through the tree trunk. If, hypothetically, the amount of deposited Ca is dependent on the amount of sap that has flown through the xylem, then these environmental factors may explain the Ca variations in tree rings.
A note should be made that the positive relation of Ca content to the ambient air temperature seems to contradict to some extent the observations of the Ca peaks after volcanic eruptions [9]. A cooling in the environment should reduce the Ca content (Table 4). However, the results presented here apply (1) only to June mean temperature and (2) to the normal weather conditions without excessive variations. Abrupt cooling for a longer time may be out of normal tree reactions and employ some different mechanisms of elemental distribution in the xylem.
5. Conclusions
The data obtained in this study gave evidence that some elements show a consistent distribution pattern between early- and late-wood in Scots pine tree rings. It has to be admitted, however, that the source of the difference that some elements show between early- vs. late-wood is not really known. A very promising opportunity for an explanation may be a deeper understanding of the organic chemistry of the xylem in the contrasting parts of the tree ring. The differences in early-wood from late-wood may be significant and may not be limited to variations in the cell walls. Early-wood cellulose is likely to be different from that of late-wood because of obvious alterations in the protoplasm functioning between spring and late summer seasons. The consequences, hypothetically, may be the different chemical and physical properties of the xylem. Based on this pattern, in-depth research may be designed and performed that would provide insight into chemical mechanisms of elemental sorption and distribution in trees’ xylems.
The impact of the northern and southern directions contains many uncertainties. The data as such are not linked to northern or southern directions for they are just data averaged over a large geographic area. However, the tree ring data are obtained from the inner rings which were closer to the outer conditions at the time when the pines were young and their tops were strongly exposed to ambient factors. Thus, at this time the orientation North-South might have played a role because the sun heat comes mainly from the southern side. As a result, the additional warming might have variated the organic composition of the tree rings, at least slightly. It is possible that the differences in the elemental contents reflect these variations. From eye sight especially, the northern direction provides more consistent patterns and some higher correlations, although this observation should be supported in further studies.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/sci6010004/s1, Table S1: Average elemental contents in tree rings of six Scots pines.
Author Contributions
Conceptualization, V.L.G. and E.A.V.; methodology, V.L.G. and R.A.S.; software, A.I.F.; validation, R.A.S. and A.I.F.; formal analysis, V.L.G.; investigation, V.L.G.; resources, A.I.F.; data curation, A.I.F.; writing—original draft preparation, V.L.G. and Z.T.; writing—review and editing, V.L.G., E.A.V., and Z.T.; visualization, A.I.F.; supervision, E.A.V.; project administration, V.L.G. and R.A.S.; funding acquisition, R.A.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article and supplementary materials.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Vaganov, E.A.; Grachev, A.M.; Shishov, V.V.; Panyushkina, I.P.; Leavitt, S.W.; Knorre, A.A.; Menyailo, O.V. Elemental composition of tree rings: A new perspective in Biogeochemistry. Dokl. Biol. Sci. 2013, 453, 375–379. [Google Scholar] [CrossRef] [PubMed]
- Baes, C.F.; Ragsdale, H.L. Age-specific lead distribution in xylem rings of three tree genera in Atlanta, Georgia. Environ. Pollut.—Ser. B Chem. Phys. 1981, 2, 21–35. [Google Scholar] [CrossRef]
- Baes, C.F.; McLaughlin, S.B. Trace elements in tree rings: Evidence of recent and historical air pollution. Science 1984, 224, 494–497. [Google Scholar] [CrossRef] [PubMed]
- Balouet, J.C.; Oudijk, G. The use of dendroecological methods to estimate the time frame of environmental releases. Environ. Claims J. 2006, 18, 35–52. [Google Scholar] [CrossRef]
- Abreu, S.N.; Soares, A.M.V.M.; Nogueira, A.J.A.; Morgado, F. Tree rings, Populus nigra L., as mercury data logger in aquatic environments: Case study of an historically contaminated environment. Bull. Environ. Contam. Toxicol. 2008, 80, 294–299. [Google Scholar] [CrossRef] [PubMed]
- Bondietti, E.A.; Baes, C.F., III; McLaughlin, S.B. Radial trends in cation ratios in tree rings as indicators of the impact of atmospheric deposition on forests. Can. J. For. Res. 1989, 19, 586–594. [Google Scholar] [CrossRef]
- Prohaska, T.; Stadlbauer, C.; Wimmer, R.; Stingeder, G.; Latkoczy, C.; Hoffmann, E.; Stephanowitz, H. Investigation of element variability in tree rings of young Norway spruce by laser-ablation-ICPMS. Sci. Total Environ. 1998, 219, 29–39. [Google Scholar] [CrossRef]
- Liu, Y.; Ta, W.; Bao, T.; Yang, Z.; Song, H.; Liu, N.; An, Z. Trace elements in tree rings and their environmental effects: A case study in Xi’an City. Sci. China Ser. D 2009, 52, 504–510. [Google Scholar] [CrossRef]
- Hevia, A.; Sánchez-Salguero, R.; Camarero, J.J.; Buras, A.; Sangüesa-Barreda, G.; Galván, J.D.; Gutiérrez, E. Towards a better understanding of long-term wood-chemistry variations in old-growth forests: A case study on ancient Pinus uncinata trees from the Pyrenees. Sci. Total Environ. 2018, 625, 220–232. [Google Scholar] [CrossRef]
- Scharnweber, T.; Rocha, E.; González Arrojo, A.; Ahlgrimm, S.; Gunnarson, B.E.; Holzkämper, S.; Wilmking, M. To extract or not to extract? Influence of chemical extraction treatment of wood samples on element concentrations in tree-rings measured by X-ray fluorescence. Front. Environ. Sci. 2023, 11, 1031770. [Google Scholar] [CrossRef]
- Gavrikov, V.; Fertikov, A.; Sharafutdinov, R.; Pyzhev, A.; Vaganov, E. Dendrochemical Challenge in Climate Science: Whether Chemical Elements in Wood Reflect the Fluctuations in Weather Parameters. Plants 2022, 11, 3240. [Google Scholar] [CrossRef] [PubMed]
- Rowell, R.M. Handbook of Wood Chemistry and Wood Composites, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2013; p. 688. [Google Scholar]
- Gavrikov, V.L.; Fertikov, A.I.; Vidus, V.E.; Sharafutdinov, R.A.; Vaganov, E.A. Elemental Variability in Stems of Pinus sylvestris L.: Whether a Single Core Can Represent All the Stem. Diversity 2023, 15, 281. [Google Scholar] [CrossRef]
- Wodzicki, T.J. Mechanism of xylem differentiation in Pinus silvestris L. J. Exp. Bot. 1971, 22, 670–687. [Google Scholar] [CrossRef]
- Deslauriers, A.; Morin, H.; Begin, Y. Cellular phenology of annual ring formation of Abies balsamea in Quebec boreal forest (Canada). Can. J. For. Res. 2003, 33, 190–200. [Google Scholar] [CrossRef]
- Rossi, S.; Deslauriers, A.; Anfodillo, T. Assessment of cambial activity and xylogenesis by microsampling tree species: An example at the alpine timberline. IAWA J. 2006, 27, 383–394. [Google Scholar] [CrossRef]
- Bertaud, F.; Holmbom, B. Chemical composition of earlywood and latewood in Norway spruce heartwood, sapwood and transition zone wood. Wood Sci. 2004, 38, 245–256. [Google Scholar] [CrossRef]
- Fredriksson, M.; Pedersen, N.B.; Thygesen, L.G. The cell wall composition of Norway spruce earlywood and latewood revisited. Int. Wood Prod. J. 2018, 9, 80–85. [Google Scholar] [CrossRef]
- Kim, J.S.; Daniel, G. Variations in cell wall ultrastructure and chemistry in cell types of earlywood and latewood in English oak (Quercus robur). IAWA J. 2016, 37, 383–401. [Google Scholar] [CrossRef]
- Antonova, G.F.; Zheleznichenko, T.V.; Stasova, V.V. Pecularities of lignification in earlywood and laterwood in Scots pine. In Proceedings of the Structure, Properties and Quality of Wood—Materials of VI B.N.Ugolev International Symposium Dedicated to 50th Anniversary of the Regional Coordination Committee on Modern Problems of Wood Science, Krasnoyarsk, Russia, 10–16 September 2018; pp. 46–58. (In Russian). [Google Scholar]
- Menyailo, O.V.; Hungate, B.A.; Zech, W. Tree species mediated soil chemical changes in a Siberian artificial afforestation experiment. Plant Soil 2002, 242, 171–182. [Google Scholar] [CrossRef]
- Schugalei, L.S. The Siberian afforestation experiment: History, methodology, and problems. In Tree Species Effects on Soils: Implications for Global Change; Binkley, D., Menyailo, O., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 257–268. [Google Scholar]
- Schweingruber, F.H. Tree rings. In Basics and Applications in Dendrochronology; D. Reidel Publishing Company: Dordrecht, The Netherlands, 1988; pp. 95–141. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023; Available online: https://www.R-project.org/ (accessed on 9 September 2023).
- Silkin, P.P.; Ekimova, N.V. Relationship of strontium and calcium concentrations with the parameters of cell structure in Siberian spruce and fir tree-rings. Dendrochronologia 2012, 30, 189–194. [Google Scholar] [CrossRef]
- Scharnweber, T.; Hevia, A.; Buras, A.; van der Maaten, E.; Wilmking, M. Common trends in elements? Within- and between-tree variations of wood-chemistry measured by X-ray fluorescence—A dendrochemical study. Sci. Total Environ. 2016, 566, 1245–1253. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).