Eyes on Memory: Pupillometry in Encoding and Retrieval

{kind=link}
{kind=link}
Abstract
1. Introduction
2. Autonomic and Central Nervous System Control of Pupil Response
3. Novelty Detection and Encoding Mechanisms
3.1. Pupil Response Patterns at Encoding Depend on the Type of Novelty Detected
The Effect of Expectation
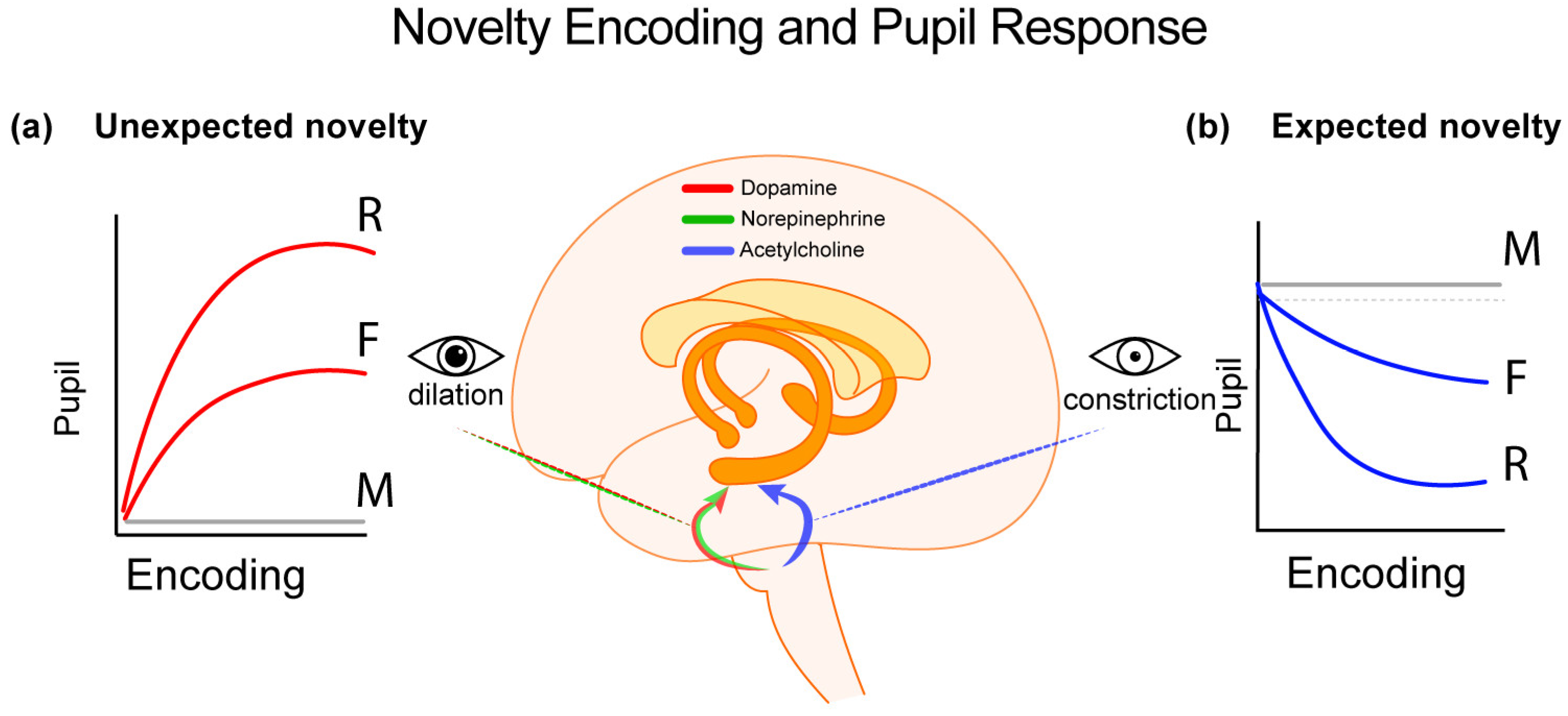
3.2. The Encoding Task as a Confound
4. Pupil Response at Retrieval
4.1. The Pupil Old/New Effect and Implications for the Episodic Memory System Development
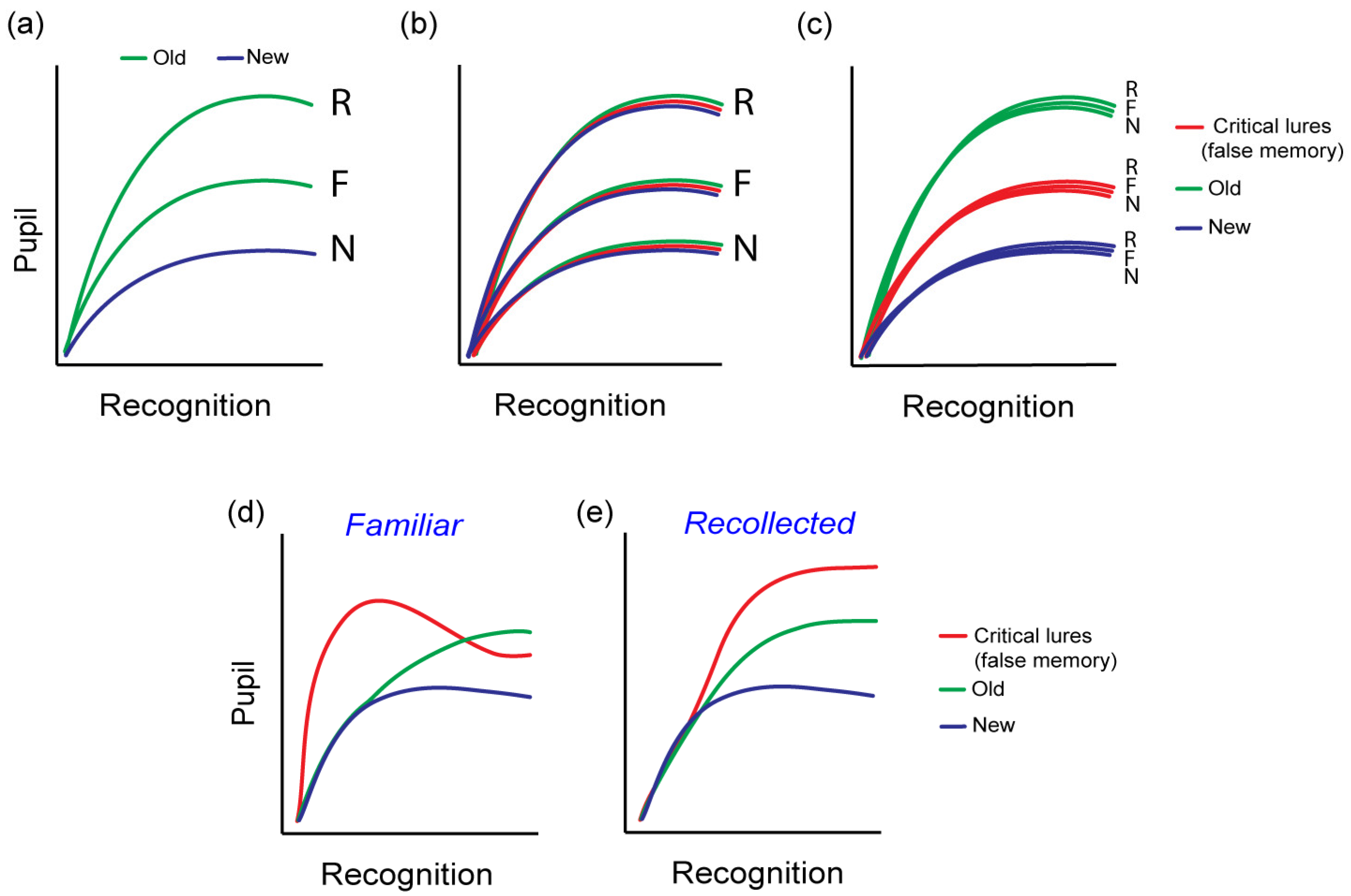
4.2. Pupil Response and the Type of Memory
4.3. The Familiarity/Novelty Conundrum
5. Is the Pupil Old/New Effect a True Memory Signal: The Case of False Memories
Implications for Theories of False Memory
6. Methodological Considerations
- Control of cognitive load and extraneous factors: It is essential to meticulously control for cognitive load and the nature of the task during encoding and retrieval phases. Pupillary responses can be influenced by various cognitive processes that are not directly related to memory, including decision-making, arousal, motivation, and task complexity. Therefore, researchers must strive to isolate memory-specific signals from those related to cognitive effort or emotional responses. This distinction is crucial for ensuring the validity of findings concerning memory processes.
- Hierarchical and systematic manipulation of experimental factors: Expanding on the previous point, it is ideal to systematically manipulate various factors—including experimental variables such as reward, valence, and anticipation—in separate experiments. This approach allows for meticulous examination of their contributions to memory-related pupil responses.
- Establishing standardized protocols for measuring and interpreting pupil size changes across various research settings is crucial. This involves standardizing the timing of measurements, environmental conditions, and data analysis techniques (see e.g., [71]). Special emphasis should be placed on selecting appropriate baseline conditions to enable the calculation of trial-specific pupillary responses. Baseline conditions should be implemented on a trial-by-trial basis and must effectively control or eliminate luminance effects. Additionally, the sensitivity of pupil responses to psychological factors such as anticipation and surprise should be considered during baseline selection. For instance, using a mask composed of scrambled pixels from a target image could inadvertently increase pupil dilation due to the unexpected nature of the arrangement (unpublished observation shown in [72]). This underscores that not all stimulus types are suitable for pupillometry. Baseline selection should be tailored to the specific stimuli used in an experiment.
- External and internal context: It is imperative that both external and individual-specific internal conditions be controlled with greater rigor than in other experimental methods. The external context encompasses factors such as lighting conditions, stimulus characteristics, and timing. Internal states, including fatigue, the influence of chemical substances (such as caffeine and alcohol), and medication, must also be carefully considered. For additional methodological considerations see [73].
- Integration with other neurophysiological measures: Combining pupillometry with other neurophysiological measures such as EEG or fMRI could provide a more comprehensive understanding of the neural networks underlying memory processes. Furthermore, many models proposed to explain the pupillary response to various cognitive states and memory, as discussed in this review, suggest neurocognitive explanations that necessitate pharmacological, neurophysiological and neuroimaging investigations. Therefore, this represents a critical direction for future research as pupillometry by itself only provides an indirect measure of brain activity and cognitive processing. Such a multimodal approach can help validate findings and elucidate the interplay between autonomic responses and brain activity in the service of memory processes.
7. Conclusions and Future Directions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Loewenfeld, I.E. The Pupil: Anatomy, Physiology and Clinical Applications; Butterworth-Heinemann: Boston, MA, USA; Oxford, UK; Auckland, New Zealand; Johannesburg, South Africa; Melbourne, Australia; New Delhi, India, 1999; Volume 1. [Google Scholar]
- Steinhauer, S.R.; Siegle, G.J.; Condray, R.; Pless, M. Sympathetic and Parasympathetic Innervation of Pupillary Dilation during Sustained Processing. Int. J. Psychophysiol. 2004, 52, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Gianaros, P.J.; Van der Veen, F.M.; Jennings, J.R. Regional Cerebral Blood Flow Correlates with Heart Period and High-Frequency Heart Period Variability during Working-Memory Tasks: Implications for the Cortical and Subcortical Regulation of Cardiac Autonomic Activity. Psychophysiology 2004, 41, 521–530. [Google Scholar] [CrossRef] [PubMed]
- Reimer, J.; McGinley, M.J.; Liu, Y.; Rodenkirch, C.; Wang, Q.; McCormick, D.A.; Tolias, A.S. Pupil Fluctuations Track Rapid Changes in Adrenergic and Cholinergic Activity in Cortex. Nat. Commun. 2016, 7, 13289. [Google Scholar] [CrossRef] [PubMed]
- Joshi, S.; Gold, J.I. Pupil Size as a Window on Neural Substrates of Cognition. Trends Cogn. Sci. 2020, 24, 466–480. [Google Scholar] [CrossRef] [PubMed]
- Haam, J.; Yakel, J.L. Cholinergic Modulation of the Hippocampal Region and Memory Function. J. Neurochem. 2017, 142, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Hasselmo, M.E. The Role of Acetylcholine in Learning and Memory. Curr. Opin. Neurobiol. 2006, 16, 710–715. [Google Scholar] [CrossRef] [PubMed]
- Ridley, R.M.; Bowes, P.M.; Baker, H.F.; Crow, T.J. An Involvement of Acetylcholine in Object Discrimination Learning and Memory in the Marmoset. Neuropsychologia 1984, 22, 253–263. [Google Scholar] [CrossRef] [PubMed]
- Easton, A.; Ridley, R.M.; Baker, H.F.; Gaffan, D. Unilateral Lesions of the Cholinergic Basal Forebrain and Fornix in One Hemisphere and Inferior Temporal Cortex in the Opposite Hemisphere Produce Severe Learning Impairments in Rhesus Monkeys. Cereb. Cortex 2002, 12, 729–736. [Google Scholar] [CrossRef] [PubMed]
- Schon, K. Scopolamine Reduces Persistent Activity Related to Long-Term Encoding in the Parahippocampal Gyrus during Delayed Matching in Humans. J. Neurosci. 2005, 25, 9112–9123. [Google Scholar] [CrossRef]
- Grothe, M.; Zaborszky, L.; Atienza, M.; Gil-Neciga, E.; Rodriguez-Romero, R.; Teipel, S.J.; Amunts, K.; Suarez-Gonzalez, A.; Cantero, J.L. Reduction of Basal Forebrain Cholinergic System Parallels Cognitive Impairment in Patients at High Risk of Developing Alzheimer’s Disease. Cereb. Cortex 2010, 20, 1685–1695. [Google Scholar] [CrossRef]
- Eldar, E.; Cohen, J.D.; Niv, Y. The Effects of Neural Gain on Attention and Learning. Nat. Neurosci. 2013, 16, 1146–1153. [Google Scholar] [CrossRef] [PubMed]
- Murphy, P.R.; O’Connell, R.G.; O’Sullivan, M.; Robertson, I.H.; Balsters, J.H. Pupil Diameter Covaries with BOLD Activity in Human Locus Coeruleus. Hum. Brain Mapp. 2014, 35, 4140–4154. [Google Scholar] [CrossRef] [PubMed]
- Walling, S.G.; Brown, R.A.M.; Milway, J.S.; Earle, A.G.; Harley, C.W. Selective Tuning of Hippocampal Oscillations by Phasic Locus Coeruleus Activation in Awake Male Rats. Hippocampus 2011, 21, 1250–1262. [Google Scholar] [CrossRef] [PubMed]
- Szabadi, E. Functional Neuroanatomy of the Central Noradrenergic System. J. Psychopharmacol. 2013, 27, 659–693. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, T.; Duszkiewicz, A.J.; Sonneborn, A.; Spooner, P.A.; Yamasaki, M.; Watanabe, M.; Smith, C.C.; Fernández, G.; Deisseroth, K.; Greene, R.W.; et al. Locus Coeruleus and Dopaminergic Consolidation of Everyday Memory. Nature 2016, 537, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Duszkiewicz, A.J.; McNamara, C.G.; Takeuchi, T.; Genzel, L. Novelty and Dopaminergic Modulation of Memory Persistence: A Tale of Two Systems. Trends Neurosci. 2019, 42, 102–114. [Google Scholar] [CrossRef] [PubMed]
- Wagatsuma, A.; Okuyama, T.; Sun, C.; Smith, L.M.; Abe, K.; Tonegawa, S. Locus Coeruleus Input to Hippocampal CA3 Drives Single-Trial Learning of a Novel Context. Proc. Natl. Acad. Sci. USA 2018, 115, E310–E316. [Google Scholar] [CrossRef] [PubMed]
- Bunzeck, N.; Duzel, E. Absolute Coding of Stimulus Novelty in the Human Substantia Nigra/VTA. Neuron 2006, 51, 369–379. [Google Scholar] [CrossRef] [PubMed]
- Kafkas, A.; Montaldi, D. How Do Memory Systems Detect and Respond to Novelty? Neurosci. Lett. 2018, 680, 60–68. [Google Scholar] [CrossRef]
- Frank, D.; Kafkas, A. Expectation-Driven Novelty Effects in Episodic Memory. Neurobiol. Learn. Mem. 2021, 183, 107466. [Google Scholar] [CrossRef]
- Biel, D.; Bunzeck, N. Novelty Before or After Word Learning Does Not Affect Subsequent Memory Performance. Front. Psychol. 2019, 10, 461358. [Google Scholar] [CrossRef] [PubMed]
- Poppenk, J.; Köhler, S.; Moscovitch, M. Revisiting the Novelty Effect: When Familiarity, Not Novelty, Enhances Memory. J. Exp. Psychol. Learn. Mem. Cogn. 2010, 36, 1321–1330. [Google Scholar] [CrossRef]
- Reggev, N.; Sharoni, R.; Maril, A. Distinctiveness Benefits Novelty (and Not Familiarity), but Only Up to a Limit: The Prior Knowledge Perspective. Cogn. Sci. 2018, 42, 103–128. [Google Scholar] [CrossRef]
- Peinkhofer, C.; Knudsen, G.M.; Moretti, R.; Kondziella, D. Cortical Modulation of Pupillary Function: Systematic Review. PeerJ 2019, 7, e6882. [Google Scholar] [CrossRef]
- Kafkas, A.; Montaldi, D. Recognition Memory Strength Is Predicted by Pupillary Responses at Encoding While Fixation Patterns Distinguish Recollection from Familiarity. Q. J. Exp. Psychol. 2011, 64, 1971–1989. [Google Scholar] [CrossRef] [PubMed]
- Kafkas, A.; Montaldi, D. The Pupillary Response Discriminates between Subjective and Objective Familiarity and Novelty. Psychophysiology 2015, 52, 1305–1316. [Google Scholar] [CrossRef]
- Miller, A.L.; Unsworth, N. Variation in Attention at Encoding: Insights from Pupillometry and Eye Gaze Fixations. J. Exp. Psychol. Learn. Mem. Cogn. 2019, 46, 2277. [Google Scholar] [CrossRef]
- Naber, M.; Frässle, S.; Rutishauser, U.; Einhäuser, W.; Frassle, S.; Rutishauser, U.; Einhauser, W. Pupil Size Signals Novelty and Predicts Later Retrieval Success for Declarative Memories of Natural Scenes. J. Vis. 2013, 13, 11. [Google Scholar] [CrossRef]
- Wetzel, N.; Einhäuser, W.; Widmann, A. Picture-Evoked Changes in Pupil Size Predict Learning Success in Children. J. Exp. Child Psychol. 2020, 192, 104787. [Google Scholar] [CrossRef]
- Papesh, M.H.; Goldinger, S.D.; Hout, M.C. Memory Strength and Specificity Revealed by Pupillometry. Int. J. Psychophysiol. 2012, 83, 56–64. [Google Scholar] [CrossRef]
- Kafkas, A.; Montaldi, D. Striatal and Midbrain Connectivity with the Hippocampus Selectively Boosts Memory for Contextual Novelty. Hippocampus 2015, 25, 1262–1273. [Google Scholar] [CrossRef]
- Bergt, A.; Urai, A.E.; Donner, T.H.; Schwabe, L. Reading Memory Formation from the Eyes. Eur. J. Neurosci. 2018, 47, 1525–1533. [Google Scholar] [CrossRef]
- Kahneman, D. Attention and Effort; Prentice-Hall: Hoboken, NJ, USA, 1973. [Google Scholar]
- Granholm, E.; Asarnow, R.F.; Sarkin, A.J.; Dykes, K.L. Pupillary Responses Index Cognitive Resource Limitations. Psychophysiology 1996, 33, 457–461. [Google Scholar] [CrossRef]
- Cronin, S.L.; Lipp, O.V.; Marinovic, W. Pupil Dilation during Encoding, but Not Type of Auditory Stimulation, Predicts Recognition Success in Face Memory. Biol. Psychol. 2023, 178, 108547. [Google Scholar] [CrossRef]
- Võ, M.L.-H.; Jacobs, A.M.; Kuchinke, L.; Hofmann, M.; Conrad, M.; Schacht, A.; Hutzler, F. The Coupling of Emotion and Cognition in the Eye: Introducing the Pupil Old/New Effect. Psychophysiology 2008, 45, 130–140. [Google Scholar] [CrossRef]
- Kafkas, A.; Montaldi, D. Familiarity and Recollection Produce Distinct Eye Movement, Pupil and Medial Temporal Lobe Responses When Memory Strength Is Matched. Neuropsychologia 2012, 50, 3080–3093. [Google Scholar] [CrossRef]
- Otero, S.C.; Weekes, B.S.; Hutton, S.B. Pupil Size Changes during Recognition Memory. Psychophysiology 2011, 48, 1346–1353. [Google Scholar] [CrossRef]
- Pilarczyk, J.; Sterna, R.; Schwertner, E.; Pacula, B.; Bartoszek, M.; Kuniecki, M. Physiological Reactions at Encoding Selectively Predict Recognition of Emotional Images. Biol. Psychol. 2022, 175, 108429. [Google Scholar] [CrossRef]
- Kafkas, A. Encoding-Linked Pupil Response Is Modulated by Expected and Unexpected Novelty: Implications for Memory Formation and Neurotransmission. Neurobiol. Learn. Mem. 2021, 180, 107412. [Google Scholar] [CrossRef]
- Ranganath, C.; Rainer, G. Neural Mechanisms for Detecting and Remembering Novel Events. Nat. Rev. Neurosci. 2003, 4, 193–202. [Google Scholar] [CrossRef]
- Mandler, G. Recognizing: The Judgment of Previous Occurrence. Psychol. Rev. 1980, 87, 252–271. [Google Scholar] [CrossRef]
- Montaldi, D.; Kafkas, A. Neural Mechanisms of Familiarity. In The Oxford Handbook of Human Memory: Foundations and Applications; Wagner, A.D., Kahana, M.J., Eds.; Oxford University Press: New York, NY, USA, 2024; ISBN 978-0-19-091801-9. [Google Scholar]
- Yonelinas, A.P. The Nature of Recollection and Familiarity: A Review of 30 Years of Research. J. Mem. Lang. 2002, 46, 441–517. [Google Scholar] [CrossRef]
- Eichenbaum, H. Hippocampus: Cognitive Processes and Neural Representations That Underlie Declarative Memory. Neuron 2004, 44, 109–120. [Google Scholar] [CrossRef]
- Lisman, J.E.; Grace, A.A. The Hippocampal-VTA Loop: Controlling the Entry of Information into Long-Term Memory. Neuron 2005, 46, 703–713. [Google Scholar] [CrossRef]
- Lloyd, B.; Nieuwenhuis, S. The Effect of Reward-Induced Arousal on the Success and Precision of Episodic Memory Retrieval. Sci. Rep. 2024, 14, 2105. [Google Scholar] [CrossRef]
- Kahneman, D.; Beatty, J. Pupil Diameter and Load on Memory. Science 1966, 154, 1583–1585. [Google Scholar] [CrossRef]
- Gross, M.P.; Dobbins, I.G. Pupil Dilation during Memory Encoding Reflects Time Pressure Rather than Depth of Processing. J. Exp. Psychol. Learn. Mem. Cogn. 2020, 47, 264. [Google Scholar] [CrossRef]
- Franzen, L.; Cabugao, A.; Grohmann, B.; Elalouf, K.; Johnson, A.P. Individual Pupil Size Changes as a Robust Indicator of Cognitive Familiarity Differences. PLoS ONE 2022, 17, e0262753. [Google Scholar] [CrossRef]
- Oliveira, J.; Fernandes, M.; Rosa, P.J.; Gamito, P. Is Pupil Activity Associated with the Strength of Memory Signal for Words in a Continuous Recognition Memory Paradigm? Front. Psychol. 2021, 12, 686183. [Google Scholar] [CrossRef]
- Hellmer, K.; Söderlund, H.; Gredebäck, G. The Eye of the Retriever: Developing Episodic Memory Mechanisms in Preverbal Infants Assessed through Pupil Dilation. Dev. Sci. 2018, 21, e12520. [Google Scholar] [CrossRef]
- Montaldi, D.; Mayes, A.R. The Role of Recollection and Familiarity in the Functional Differentiation of the Medial Temporal Lobes. Hippocampus 2010, 20, 1291–1314. [Google Scholar] [CrossRef]
- Siefert, E.M.; He, M.; Festa, E.K.; Heindel, W.C. Pupil Size Tracks Cue–Trace Interactions during Episodic Memory Retrieval. Psychophysiology 2024, 61, e14409. [Google Scholar] [CrossRef]
- Kucewicz, M.T.; Dolezal, J.; Kremen, V.; Berry, B.M.; Miller, L.R.; Magee, A.L.; Fabian, V.; Worrell, G.A. Pupil Size Reflects Successful Encoding and Recall of Memory in Humans. Sci. Rep. 2018, 8, 4949. [Google Scholar] [CrossRef]
- Kafkas, A.; Brown, T.; Olusola, N.; Guo, C. Pupil Response Patterns Distinguish True from False Memories. Sci. Rep. 2023, 13, 17244. [Google Scholar] [CrossRef]
- Kafkas, A.; Montaldi, D. Two Separate, but Interacting, Neural Systems for Familiarity and Novelty Detection: A Dual-Route Mechanism. Hippocampus 2014, 24, 516–527. [Google Scholar] [CrossRef]
- Montefinese, M.; Ambrosini, E.; Fairfield, B.; Mammarella, N. The “Subjective” Pupil Old/New Effect: Is the Truth Plain to See? Int. J. Psychophysiol. 2013, 89, 48–56. [Google Scholar] [CrossRef]
- Laeng, B.; Waterloo, K.; Johnsen, S.H.; Bakke, S.J.; Lag, T.; Simonsen, S.S.; Hogsaet, J. The Eyes Remember It: Oculography and Pupillometry during Recollection in Three Amnesic Patients. J. Cogn. Neurosci. 2007, 19, 1888–1904. [Google Scholar] [CrossRef]
- Jacoby, L.L.; Kelley, C.M.; Dywan, J. Memory Attributions. In Varieties of Memory and Consciousness: Essays in Honour of Endel Tulving; Roediger, H.L., III, Craik, F.I.M., Eds.; Erlbaum: Hillsdale, MI, USA, 1989; pp. 391–422. [Google Scholar]
- Whittlesea, B.W.A. False Memory and the Discrepancy-Attribution Hypothesis: The Prototype-Familiarity Illusion. J. Exp. Psychol. Gen. 2002, 131, 96–115. [Google Scholar] [CrossRef]
- Bastin, C.; Besson, G.; Simon, J.; Delhaye, E.; Geurten, M.; Willems, S.; Salmon, E. An Integrative Memory Model of Recollection and Familiarity to Understand Memory Deficits. Behav. Brain Sci. 2019, 42, e281. [Google Scholar] [CrossRef]
- Roediger, H.L.; Watson, J.M.; McDermott, K.B.; Gallo, D.A. Factors That Determine False Recall: A Multiple Regression Analysis. Psychon. Bull. Rev. 2001, 8, 385–407. [Google Scholar] [CrossRef]
- Brainerd, C.J.; Reyna, V.F. Fuzzy-Trace Theory and False Memory. Curr. Dir. Psychol. Sci. 2002, 11, 164–169. [Google Scholar] [CrossRef]
- Jacoby, L.L.; Whitehouse, K. An Illusion of Memory: False Recognition Influenced by Unconscious Perception. J. Exp. Psychol. Gen. 1989, 118, 126–135. [Google Scholar] [CrossRef]
- Rosburg, T.; Mecklinger, A.; Frings, C. When the Brain Decides. Psychol. Sci. 2011, 22, 1527–1534. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.K.; Hashtroudi, S.; Lindsay, D.S. Source Monitoring. Psychol. Bull. 1993, 114, 3–28. [Google Scholar] [CrossRef] [PubMed]
- Brainerd, C.J.; Gomes, C.F.A.; Nakamura, K. Dual Recollection in Episodic Memory. J. Exp. Psychol. Gen. 2015, 144, 816–843. [Google Scholar] [CrossRef] [PubMed]
- Ryals, A.J.; Kelly, M.E.; Cleary, A.M. Increased Pupil Dilation during Tip-of-the-Tongue States. Conscious. Cogn. 2021, 92, 103152. [Google Scholar] [CrossRef] [PubMed]
- Kret, M.E.; Sjak-Shie, E.E. Preprocessing Pupil Size Data: Guidelines and Code. Behav. Res. Methods 2019, 51, 1336–1342. [Google Scholar] [CrossRef] [PubMed]
- Kafkas, A. (University of Manchester, Manchester, UK). Unpublished Work. 2013. [Google Scholar]
- Steinhauer, S.R.; Bradley, M.M.; Siegle, G.J.; Roecklein, K.A.; Dix, A. Publication Guidelines and Recommendations for Pupillary Measurement in Psychophysiological Studies. Psychophysiology 2022, 59, e14035. [Google Scholar] [CrossRef]
- Lech, M.; Czyżewski, A.; Kucewicz, M.T. CyberEye: New Eye-Tracking Interfaces for Assessment and Modulation of Cognitive Functions beyond the Brain. Sensors 2021, 21, 7605. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kafkas, A. Eyes on Memory: Pupillometry in Encoding and Retrieval. Vision 2024, 8, 37. https://doi.org/10.3390/vision8020037
Kafkas A. Eyes on Memory: Pupillometry in Encoding and Retrieval. Vision. 2024; 8(2):37. https://doi.org/10.3390/vision8020037
Chicago/Turabian StyleKafkas, Alex. 2024. "Eyes on Memory: Pupillometry in Encoding and Retrieval" Vision 8, no. 2: 37. https://doi.org/10.3390/vision8020037
APA StyleKafkas, A. (2024). Eyes on Memory: Pupillometry in Encoding and Retrieval. Vision, 8(2), 37. https://doi.org/10.3390/vision8020037