A Comparative Analysis of the Camera-like Eyes of Jumping Spiders and Humans

Abstract
1. Introduction
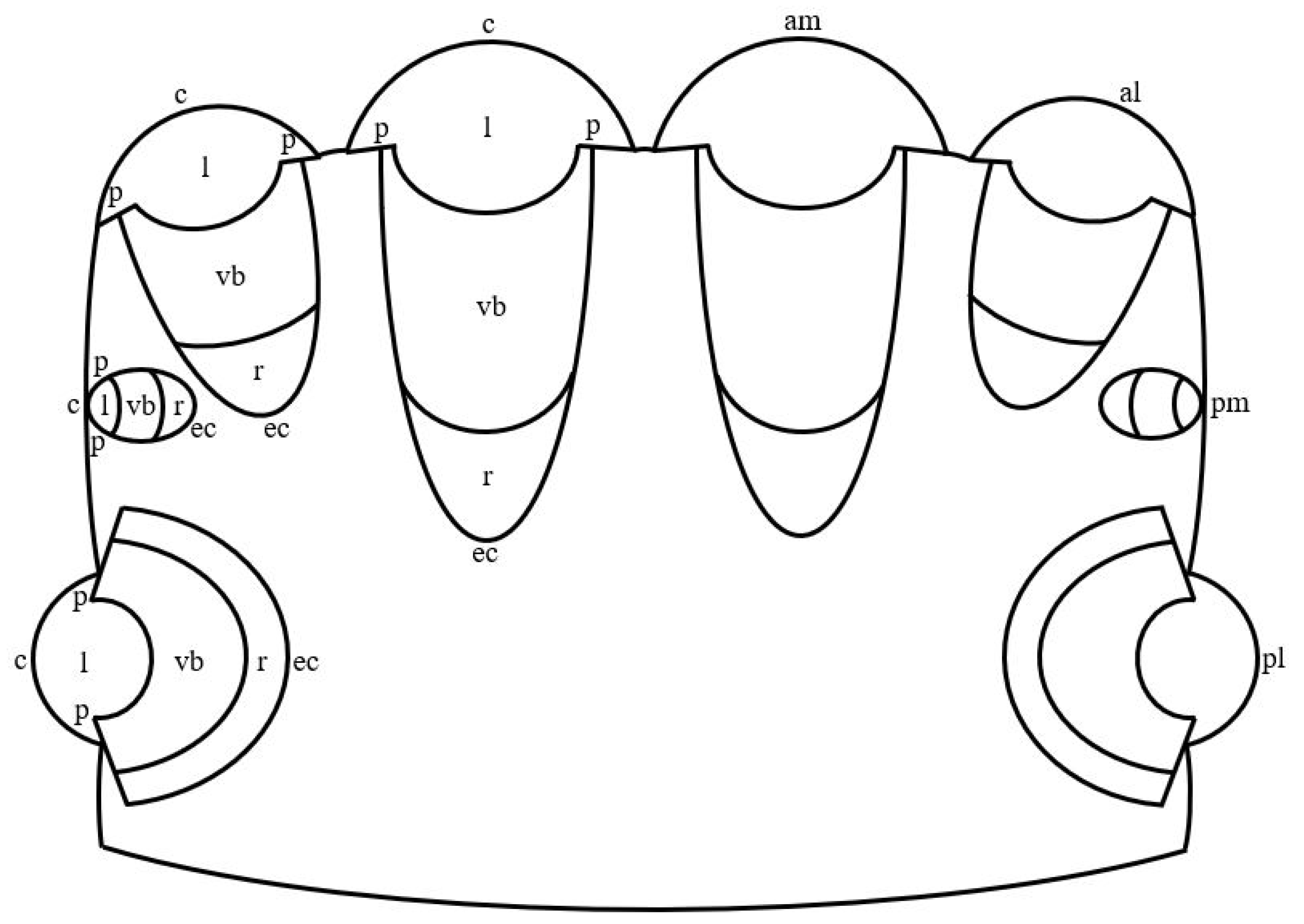
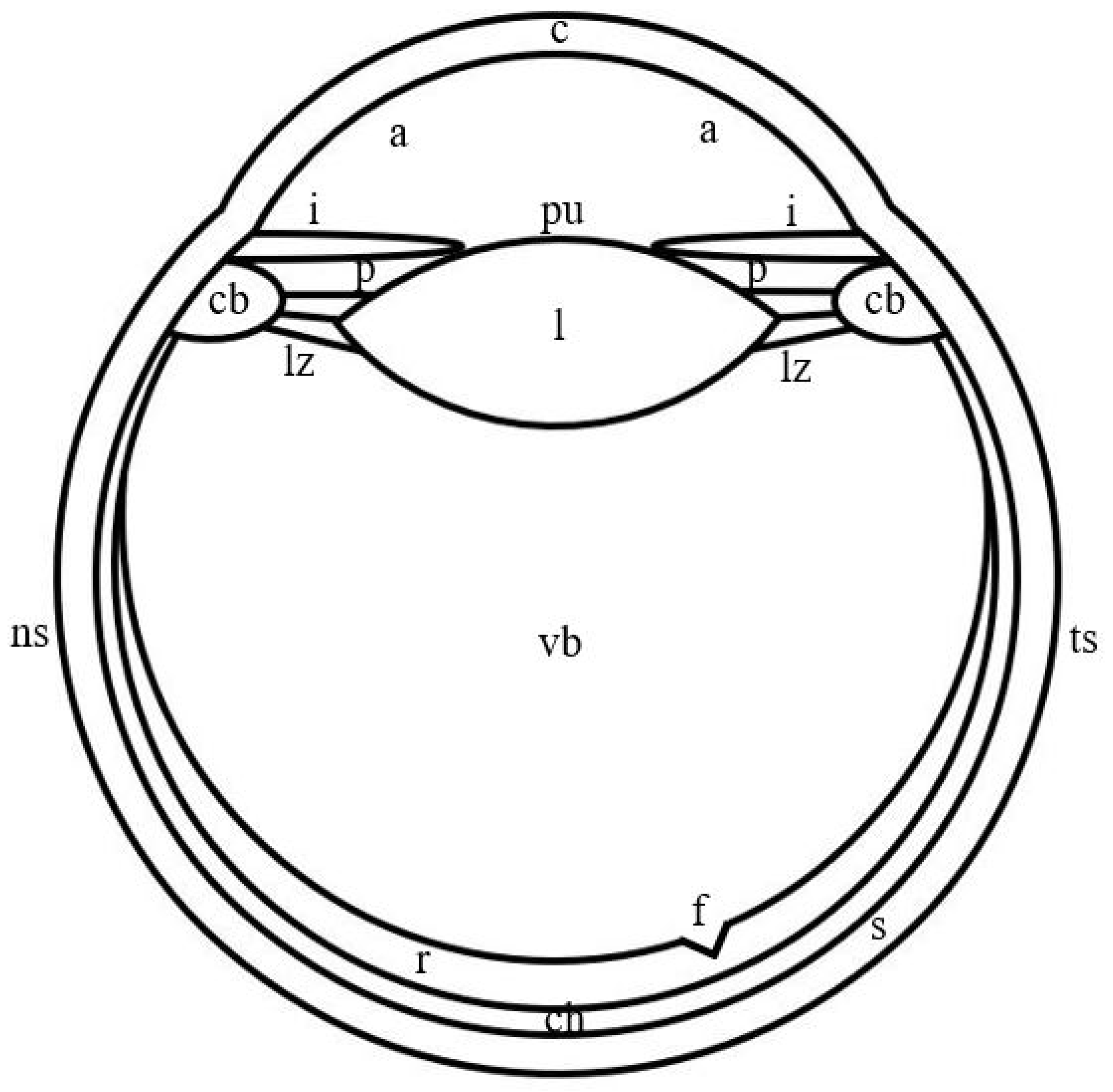
2. General Characteristics of the Camera-like Eyes of Jumping Spiders and Humans
3. The Outer Shell of the Camera-like Eyes of Jumping Spiders and Humans
3.1. Cornea
3.2. Eye Capsule and Sclera
4. The Inner Shell of the Camera-like Eyes of Jumping Spiders and Humans
5. The Pupil of the Camera-like Eyes of Jumping Spiders and Humans
6. The Lens of the Camera-like Eyes of Jumping Spiders and Humans
7. The Vitreous Body of the Camera-like Eyes of Jumping Spiders and Humans
8. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Land, M.F. Structure of the retinae of the principal eyes of jumping spiders (Salticidae: Dendryphantinae) in relation to visual optics. J. Exp. Biol. 1969, 51, 443–470. [Google Scholar] [CrossRef]
- Land, M.F.; Nilsson, D.-E. Animal Eyes, 2nd ed.; Oxford University Press: Oxford, UK, 2012. [Google Scholar]
- Shepeleva, I.P. A Comparative Morphology and Optical Properties of the Eyes of Gastropod Mollusks Stylommatophora (Gastropoda: Pulmonata). Ph.D. Thesis, Lomonosov Moscow State University, Moscow, Russia, 2007. [Google Scholar]
- Shepeleva, I.P. A comparative analysis of the camera-like eyes of gastropod mollusks and humans. Sens. Syst. 2013, 27, 317–326. [Google Scholar]
- Shepeleva, I.P. A comparative analysis of the resolving power of the camera-like eyes of gastropod mollusks and humans. Russ. J. Physiol. 2018, 104, 412–424. [Google Scholar]
- Shepeleva, I.P. Light-refracting, light-sensitive and light-insulating apparatus of the camera-like eyes. J. Gen. Biol. 2021, 82, 229–240. [Google Scholar]
- Shepeleva, I.P. Optical sensitivity of camera-like eyes to with light. Vision 2021, 5, 44. [Google Scholar] [CrossRef] [PubMed]
- Jacob, E.M.; Long, S.M.; Harland, D.P. Lateral eyes direct principle eyes as jumping spiders track objects. Curr. Biol. 2018, 28, 1092–1093. [Google Scholar] [CrossRef]
- World Spider Catalog. Version 22.0. Natural History Museum Bern. 2021. Available online: http://wsc.nmbe.ch (accessed on 26 October 2021).
- Meehan, C.J.; Olson, E.J.; Reudink, M.W.; Kyser, T.K.; Curry, R.L. Herbivory in a spider through exploitation of an ant-plant mutualism. Curr. Biol. 2009, 19, 892–893. [Google Scholar] [CrossRef]
- Foelix, R.F. Biology of Spiders; Oxford University Press: Oxford, UK, 2011. [Google Scholar]
- Cerveira, A.M.; Jackson, R.R.; Nelson, X.J. Dim-light vision in jumping spiders (Araneae, Salticidae): Identification of prey and rivals. J. Exp. Biol. 2019, 222, jeb198069. [Google Scholar] [CrossRef] [PubMed]
- Harland, D.P.; Li, D.; Jackson, R.R. How jumping spiders see the world. In How Animals See the World: Comparative Behavior, Biology, and Evolution of Vision; Lazareva, O.F., Shimizu, T., Wasserman, E.A., Eds.; Oxford University Press: Oxford, UK, 2012; pp. 133–163. [Google Scholar]
- Zurek, D.B.; Nelson, X.J. Saccadic tracking of targets mediating by the anterior-lateral eyes of jumping spiders. J. Comp. Physiol. 2012, 198, 411–417. [Google Scholar] [CrossRef]
- Danilichev, V.F. Modern Ophthalmology; Peter: St. Petersburg, Russia, 2009. [Google Scholar]
- Alekseev, V.N.; Egorov, E.A.; Astakhov, Y.S.; Stavitskaya, T.V. Ophthalmology; GEOTAR-Media: Moscow, Russia, 2010. [Google Scholar]
- Land, M.F. Movements of the retinae of jumping spiders (Salticidae: Dendryphantinae) in response to visual stimuli. J. Exp. Biol. 1969, 51, 471–493. [Google Scholar] [CrossRef]
- Williams, D.S.; McIntyre, P. The principle eyes of a jumping spider have a telephoto component. Nature 1980, 288, 578–580. [Google Scholar] [CrossRef]
- Campbell, F.W.; Gubisch, R.W. Optical quality of the human eye. J. Physiol. 1966, 186, 558–578. [Google Scholar] [CrossRef] [PubMed]
- Curcio, C.A.; Sloan, K.R.; Kalina, R.E.; Hendrickson, A.E. Human photoreceptor topography. J. Comp. Neurol. 1990, 292, 497–523. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, S.; Tateda, H. Spectral sensitivities of jumping spider eyes. J. Comp. Physiol. 1976, 105, 29–41. [Google Scholar] [CrossRef]
- Zurek, D.B.; Cronin, T.W.; Taylor, L.A.; Byrne, K.; Sullivan, M.L.G.; Morehouse, N.I. Spectral filtering enables trichromatic vision in colorful jumping spider. Curr. Biol. 2015, 25, 391–408. [Google Scholar] [CrossRef] [PubMed]
- Morehouse, N.I.; Zurek, D.B.; Taylor, L.A.; Cronin, T. Repeated evolution of color vision underlies rapid diversification of salticid male coloration. In Proceedings of the Abstract of the Annual Meeting of SICB, New Orleans, LA, USA, 4–8 January 2017. [Google Scholar]
- Grüsser, O. Vision and eye movements. In Human Physiology; Schmidt, R.F., Thews, G.M., Eds.; Mir: Moscow, Russia, 1985; pp. 90–105. [Google Scholar]
- Ignatov, I.; Mosin, O. Studying of phototransformation of light signal by photoreceptor pigments—Rhodopsin, iodopsin and bacteriorhodopsin. Eur. J. Mol. Biotechnol. 2014, 5, 104–119. [Google Scholar]
- Blest, A.D.; Price, G.D. Retinal mosaics of the principle eyes of some jumping spiders (Salticida: Araneae): Adaptation for high visual acuity. Protoplasma 1984, 120, 172–184. [Google Scholar] [CrossRef]
- Atchison, D.A.; Smith, G. Optics of the Human Eye; Butterworth-Heinemann: Oxford, UK, 2000. [Google Scholar]
- Nagata, T.; Koyanagi, M.; Tsukamoto, H.; Saeki, S.; Isono, K.; Shichida, Y.; Tokunaga, F.; Kinoshita, M.; Arikawa, K.; Terakita, A. Depth perception from image defocus in a jumping spider. Science 2012, 335, 469–471. [Google Scholar] [CrossRef]
- Schiffman, C.R. Sensation and Perception; Peter: St. Petersburg, Russia, 2003. [Google Scholar]
- Nemov, R.S. General Psychology. Sensation and Perception; Yurayt: Moscow, Russia, 2020; Book 1; Volume II. [Google Scholar]
- Homann, H. Beitrfige zur Physiologie der Spinnenaugen. I. Untersuchungsmethoden. II. Das Sechvermorgen der Salticiden. Z. Vergl. Physiol. 1928, 7, 201–268. [Google Scholar] [CrossRef]
- Terakita, A.; Nagata, T. Functional properties of opsins and their contribution to light-sensing physiology. Zool. Sci. 2014, 31, 653–659. [Google Scholar] [CrossRef]
- Forster, L.M. Visual mechanisms of hunting behaviour in Trite planiceps, a jumping spider (Araneae: Salticidae). N. Z. J. Zool. 1979, 6, 79–93. [Google Scholar] [CrossRef]
- Prokopenko, V.T.; Trofimov, V.A.; Sharok, L.P. Psychology of Visual Perception; St. Petersburg State University ITMO: St. Petersburg, Russia, 2006. [Google Scholar]
- Romanenko, V.N.; Cheredova, V.V. Hunting behavior of free-living spiders. Tomsk State Univ. Bull. 2009, 4, 41–46. [Google Scholar]
- Zurek, D.B.; Nelson, X.J. Hyperacute motion detection by the lateral eyes of jumping spiders. Vis. Res. 2012, 66, 26–30. [Google Scholar] [CrossRef]
- Blest, A.D. Ultrastructure of secondary retinae of primitive and advanced jumping spiders (Araneae, Salticidae). Zoomorph. 1983, 102, 125–141. [Google Scholar] [CrossRef]
- Blest, A.D. Retinal mosaics of the principal eyes of jumping spiders (Salticidae) in some neotropical habitats: Optical trade-offs between sizes and habitat illuminances. J. Comp. Physiol. 1985, 157, 391–404. [Google Scholar] [CrossRef]
- Stone, R.A.; Flitcroft, D.I. Ocular shape and myopia. Ann. Acad. Med. Singap. 2004, 33, 7–15. [Google Scholar]
- Eakin, R.M.; Brandenburger, J.L. Fine structure of the eyes of jumping spiders. J. Ultrastruct. Res. 1971, 37, 618–663. [Google Scholar] [CrossRef]
- Bekerman, I.; Gottlieb, P.; Vaiman, M. Variations in eyeball diameters of the healthy adults. J. Ophthalmol. 2014, 2014, 503645. [Google Scholar] [CrossRef]
- Fenk, L.M.; Heidlmayr, K.; Linder, P.; Schmid, A. Pupil size in spider eyes is linked to post-ecdusal lens growth. PLoS ONE. 2010, 5, e15838. [Google Scholar] [CrossRef] [PubMed]
- Sebastian, P.A.; Mathew, M.J. Introduction to spiders. In Spiders of India; Sebastian, P.A., Peter, K.A., Eds.; University Press: Hyderabad, India, 2009; pp. 1–40. [Google Scholar]
- Dawson, D.G.; Ubels, J.L.; Edelhauser, H.F. Cornea and sclera. In Adler’s Physiology of the Eye; Levin, L.A., Nilsson, S.F.E., Ver Hoeve, J., Wu, S.M., Eds.; Elsevier Mosby: St. Louis, MI, USA, 2011; pp. 71–130. [Google Scholar]
- Politi, Y.; Priewasser, M.; Pippel, E.; Zaslansky, P.; Hartmann, J.; Siegel, S.; Li, C.; Barth, F.G.; Fratzl, P. A spider’s fang: How to design an injection needle using chitin-based composite material. Adv. Funct. Mater. 2012, 22, 2519–2528. [Google Scholar] [CrossRef]
- Dua, H.S.; Faraj, L.A.; Said, D.G.; Gray, T.; Lowe, J. Human corneal anatomy redefined: A novel pre-Descemet’s layer (Dua’s layer). Ophthalmology 2013, 120, 1778–1785. [Google Scholar] [CrossRef] [PubMed]
- Feneck, E.M.; Lewis, P.N.; Ralphs, J.; Meek, K.M. A comparative study of the elastic fiber system within the mouse and human cornea. Exp. Eye Res. 2018, 177, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Douglas, R.H.; Marshall, N.J. A review of vertebrate and invertebrate ocular filters. In Adaptive Mechanisms in the Ecology of Vision; Archer, S.N., Djamgoz, M.B.A., Loew, E.R., Partridge, J.C., Vallerga, S., Eds.; Springer: Dordrecht, The Netherlands, 1999; pp. 95–162. [Google Scholar]
- Hu, Z.; Liu, F.; Xu, X.; Chen, Z.; Chen, J.; Li, D. Spectral transmission of the principal-eye corneas of jumping spiders: Implications for ultraviolet vision. J. Exp. Biol. 2012, 215, 2853–2859. [Google Scholar] [CrossRef][Green Version]
- Boettner, E.A.; Walter, J.R. Transmission of the ocular media. Investig. Ophthalmol. Vis. Sci. 1962, 1, 776–783. [Google Scholar]
- De Voe, R.D. Ultraviolet and green receptors in principal eyes of jumping spiders. J. Gen. Physiol. 1975, 66, 193–207. [Google Scholar] [CrossRef]
- Blest, A.D.; Hardie, R.C.; McIntyre, P.; Williams, D.S. The spectral sensitivities of identified receptors and the function of retinal tiering in the principle eyes of a jumping spider. J. Comp. Physiol. 1981, 145, 227–239. [Google Scholar] [CrossRef]
- Blest, A.D. Post-embryonic development of the principle retina of a jumping spider. I. The establishment of receptor tiering by conformational changes. Phil. Trans. R. Soc. Lond. 1988, 320, 489–504. [Google Scholar]
- Schwab, I.R. The evolution of eyes: Major steps. The Keeler lecture 2017: Centenary of Keeler Ltd. Eye 2018, 32, 302–313. [Google Scholar] [CrossRef]
- Cronin, N.W.; Johnsen, S.; Marshall, N.J.; Warrant, E.J. Visual Ecology; Princeton University Press: Princeton, NJ, USA; Oxford, UK, 2014. [Google Scholar]
- Cherkasov, V.G.; Kravchuk, S.Y. Human Anatomy; New Book: Vinnytsia, Ukraine, 2014. [Google Scholar]
- Forrester, J.V.; Dick, A.D.; McMenamin, P.G.; Roberts, F.; Pearlman, E. The Eye. Basic Sciences in Practice; Elsevier: Edinburg, Scotland, 2021. [Google Scholar]
- Read, S.A.; Alonso-Caneiro, D.; Vincent, S.J.; Bremner, A.; Fothergill, A.; Ismail, B.; McGraw, R.; Quirk, C.J.; Wrigley, E. Anterior eye tissue morphology: Scleral and conjunctival thickness in children and young adults. Sci. Rep. 2016, 6, 33796. [Google Scholar] [CrossRef] [PubMed]
- Schubert, H.D. Structure of the neural retina. In Ophthalmology, 4th ed.; Yanoff, M., Duker, J.S., Eds.; Elsevier: Edinburgh, Scotland, 2014; pp. 419–423. [Google Scholar]
- Schalch, W.; Bone, R.A.; Landrum, J.T. The functional role of xanthophylls in the primate retina. In Carotenoids: Physical, Chemical and Biological Functions and Properties; Landrum, J.T., Ed.; CRC Press: Boca Raton, FL, USA, 2010; pp. 261–263. [Google Scholar]
- Abass, A.; Vinciguerra, R.; Lopes, B.T.; Bao, F.; Vinciguerra, P.; Ambrósio, R., Jr.; Elsheikh, A. Positions of ocular geometrical and visual axes in Brazilian, Chinese and Italian populations. Curr. Eye Res. 2018, 43, 1404–1414. [Google Scholar] [CrossRef] [PubMed]
- Hildebrand, G.D.; Fielder, A.R. Anatomy and physiology of the retina. In Pediatric Retina; Olitsky, S., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 39–65. [Google Scholar]
- Hollyfield, J.G. Hyaluronan and the functional organization of the interphotoreceptor matrix. Investig. Ophthalmol. Vis. Sci. 1999, 40, 2767–2769. [Google Scholar]
- Al-Ubaidi, M.R.; Naash, M.I.; Conley, S.M. A perspective on the role of the extracellular matrix in progressive retinal degenerative disorders. Investig. Ophthalmol. Vis. Sci. 2013, 54, 8119–8124. [Google Scholar] [CrossRef] [PubMed]
- Reichenbach, A.; Franze, K.; Agte, S.; Junek, S.; Wurm, A.; Grosche, J.; Savvinov, A.; Guck, J.; Skatchkov, S.N. Live cell as optic fibers in the vertebrate retina. In Selected Topics on Optic Fiber Technology; Yasin, M., Harun, S.W., Arof, H., Eds.; IntechOpen: London, UK, 2012; pp. 247–270. [Google Scholar]
- Ivanov, A.V. Spiders, Their Structure, Lifestyle and Significance for Humans; LSU: Leningrad, Russia, 1965. [Google Scholar]
- Zurek, D.B.; Taylor, A.J.; Evans, C.S.; Nelson, X.J. The role of the anterior lateral eyes in the vision-based behaviour of jumping spiders. J. Exp. Biol. 2010, 213, 2372–2378. [Google Scholar] [CrossRef] [PubMed]
- Zak, P.P.; Ostrovsky, M.A. The yellowness of the optical media of the eye in the physiology and pathology of human vision. Sens. Syst. 1995, 9, 9–20. [Google Scholar]
- Whitehead, A.J.; Mares, J.A.; Danis, R.P. Macular Pigment. Arch. Ophthalmol. 2006, 124, 1038–1045. [Google Scholar] [CrossRef]
- Zimatkin, S.M.; Matsyuk, Y.R.; Mozheyko, L.A.; Mikhalchuk, E.C. Cytology, Histology and Embryology; Higher School: Minsk, Belarus, 2018. [Google Scholar]
- Wang, X.; Xiong, K.; Lu, L.; Gu, D.; Wang, S.; Chen, J.; Xiao, H.; Zhou, G. Developmental origin of the posterior pigmented epithelium of iris. Cell Biochem. Biophys. 2015, 71, 1067–1076. [Google Scholar] [CrossRef]
- Skalicky, S.E. Ocular and Visual Physiology; Springer: Singapore, 2016. [Google Scholar]
- Wyatt, H.J. The form of the human pupil. Vis. Res. 1995, 35, 2021–2036. [Google Scholar] [CrossRef]
- Flesch, P. Light and Light Sources; Springer: Berlin/Heidelberg, Germany, 2006. [Google Scholar]
- Charlier, J.R.; Behague, M.; Buquet, C. Shift of the pupil center with pupil constriction. Investig. Ophthalmol. Vis. Sci. 1994, 35, 1278. [Google Scholar]
- Hughes, M.O. Anatomy of the anterior eye for ocularists. J. Ophthalmol. Prosthet. 2004, 8, 25–35. [Google Scholar]
- Mathôt, S. Pupillometry: Psychology, physiology, and function. J. Cognition. 2018, 1, 16. [Google Scholar] [CrossRef]
- Weeber, H.A.; Eckert, G.; Pechhold, W.; Rob, G.L.; van der Heijde, R.G. Stiffness gradient in the crystalline lens. Graefe’s Arch. Clin. Exp. Ophthalmol. 2007, 245, 1357–1366. [Google Scholar] [CrossRef] [PubMed]
- Alio, J.L.; Anania, A.; Sagnelli, P. The aging of the human lens. In Age-Related Changes of the Human Eye; Cavallotti, C.A.P., Cerulli, L., Eds.; Humana Press: Totowa, NJ, USA, 2008; pp. 61–131. [Google Scholar]
- Chirila, T.V.; Hong, Y. The vitreous humor. In Handbook of Biomaterial Properties; Black, J., Hastings, G., Eds.; Chapman & Hall: London, UK, 1998; pp. 125–131. [Google Scholar]



{kind=link}
{kind=link}
| Properties and Functions of Components | Similarities (+) and Differences (−) |
|---|---|
| Outer Shell | |
| Cornea | |
| Properties | |
| Location in the eyes | + |
| Size relative to the size of the outer shell | + |
| Transparency | + |
| Pigmentation | + |
| Structure | − |
| Shape of refractive surfaces | − |
| Lens type | + |
| Refractive index | − |
| Functions | |
| Shaping | + |
| Supporting | + |
| Protective | + |
| Light-refracting | + |
| Light-guiding | + |
| Light filter for UV and IR radiation | + |
| Reducing spherical aberration | − |
| Eye Capsule/Sclera | |
| Properties | |
| Location in the eyes | + |
| Size relative to the size of the outer shell | + |
| Transparency | + |
| Pigmentation | + |
| Structure | − |
| Refractive index | + |
| Functions | |
| Shaping | + |
| Supporting | + |
| Protective | + |
| Muscle attachment site | + |
| Participation in the outflow of intraocular fluid | − |
| Inner Shell | |
| Retina | |
| Properties | |
| Mobility | − |
| Shape | − |
| Optic and non-optic part | + |
| Location of the optic part | − |
| The shape of the optic part | − |
| Location of the non-optic part | − |
| Fovea | +/− |
| The optic part consists of a fovea | − |
| Central and peripheral region in the optic part | +/− |
| Zones in the central and peripheral region of the optic part | − |
| Fovea shape | − |
| Fovea curvature | − |
| Fovea depth | − |
| Fovea width | − |
| The location of the fovea relative to the optical axis of the eye | − |
| Fovea pigmentation | − |
| The presence of all retinal layers in the fovea | − |
| Inversion | +/− |
| The type of tissue in the optic part | + |
| The number of layers in the optic part | ? |
| Layers in the optic part | ? |
| The number of cell types in the optic part | +/− |
| Types of cells in the optic part | +/− |
| The type of tissue in the non-optic part | + |
| The number of layers in the non-optic part | ? |
| Layers in the non-optic part | ? |
| The number of cell types in the non-optic part | + |
| Types of cells in the non-optic part | + |
| Extracellular matrix of the retina | ? |
| Refractive index | + |
| Functions | |
| Shaping | + |
| Supporting | + |
| Light-guiding | + |
| Central vision | +/− |
| Color perception | +/− |
| Peripheral vision | +/− |
| Light perception | +/− |
| Perception of the depth of space based on monocular cues | +/− |
| Perception of the depth of space based on binocular cues | +/− |
| Reducing chromatic aberration | − |
| Pupil | |
| Properties | |
| Presence | +/− |
| Location relative to the cornea | + |
| Location relative to the lens | − |
| Location in the shell of the eye | − |
| Shape | − |
| Diameter change | − |
| Functions | |
| Penetration of light into the eye | + |
| Regulation of the amount of light entering the eye | − |
| Buffer when switching from bright light to dark | − |
| Influence on visual parameters through diameter change | − |
| A channel for aqueous humor between the anterior and posterior chambers of the eye | − |
| Lens | |
| Properties | |
| Location relative to the cornea | − |
| Size relative to eye size | − |
| Transparency | + |
| Consistency | + |
| Pigmentation | − |
| Size relative to pupil size | +/− |
| Shape | +/− |
| Shape of refractive surfaces | +/− |
| Structure | − |
| Optical homogeneity | − |
| Refractive index | +/− |
| Functions | |
| Light-refracting | + |
| Light-guiding | + |
| Shaping | + |
| Supporting | + |
| Protective | + |
| Participation in accommodation | − |
| Reducing spherical aberration | +/− |
| Reducing chromatic aberration | − |
| Light filter for UV and IR radiation | + |
| Vitreous body | |
| Properties | |
| Location in the eyes | + |
| Size relative to eye size | + |
| Transparency | + |
| Pigmentation | + |
| Structure | − |
| Refractive index | + |
| Functions | |
| Shaping | + |
| Supporting | + |
| Protective | + |
| Light-guiding | + |
| Participation in the formation of the lens | − |
| Light filter for UV and IR radiation | + |
| Metabolic | − |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shepeleva, I.P. A Comparative Analysis of the Camera-like Eyes of Jumping Spiders and Humans. Vision 2022, 6, 2. https://doi.org/10.3390/vision6010002
Shepeleva IP. A Comparative Analysis of the Camera-like Eyes of Jumping Spiders and Humans. Vision. 2022; 6(1):2. https://doi.org/10.3390/vision6010002
Chicago/Turabian StyleShepeleva, Irina P. 2022. "A Comparative Analysis of the Camera-like Eyes of Jumping Spiders and Humans" Vision 6, no. 1: 2. https://doi.org/10.3390/vision6010002
APA StyleShepeleva, I. P. (2022). A Comparative Analysis of the Camera-like Eyes of Jumping Spiders and Humans. Vision, 6(1), 2. https://doi.org/10.3390/vision6010002