1. Introduction
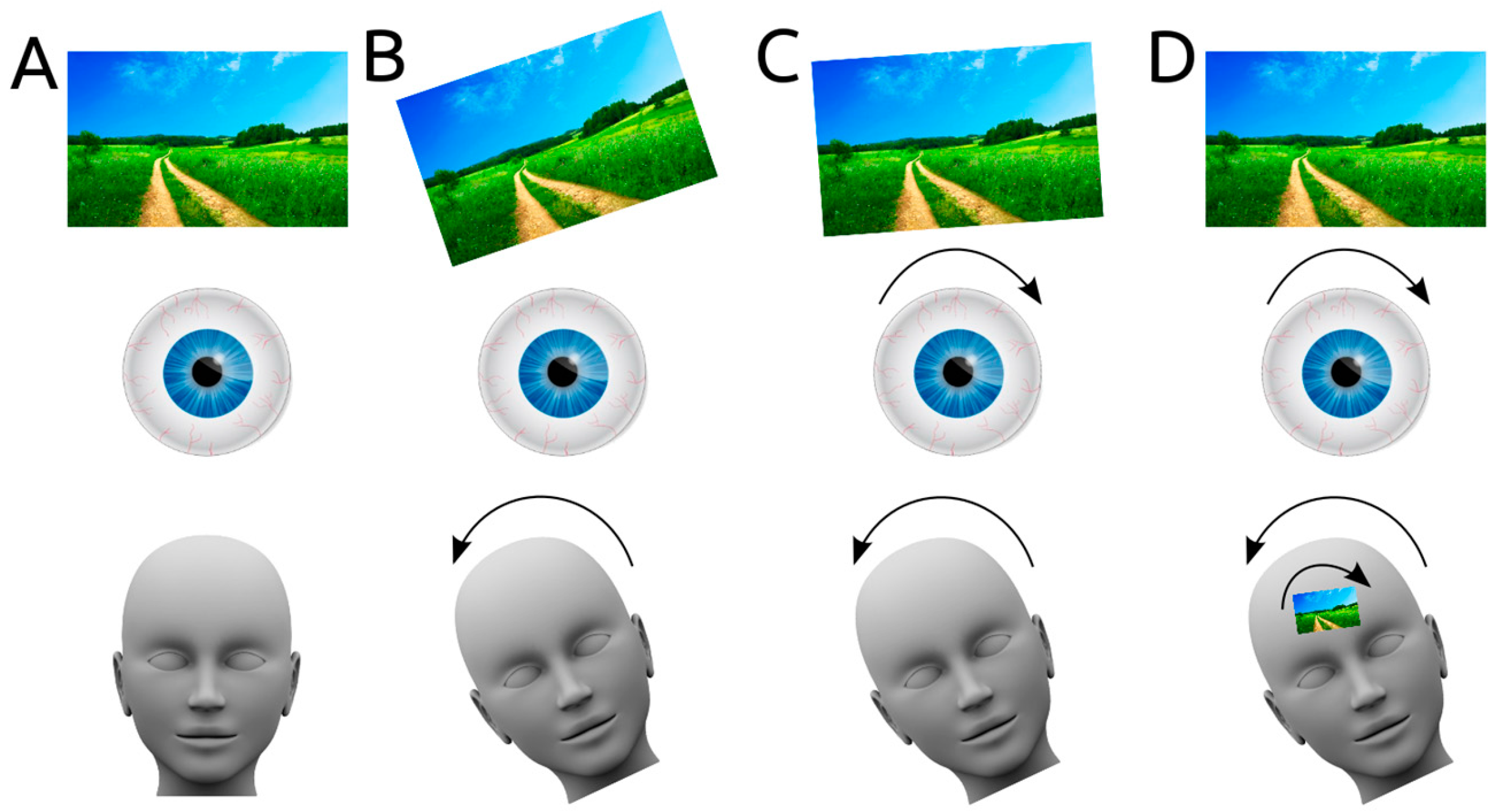
During everyday movements like walking, the human head and eyes continuously move, yet humans have a relatively stable visual perception of the world. This is due to the vestibulo-ocular reflex. When the vestibular system senses a head movement, it signals directly to the eye muscles, and a compensatory eye movement is produced to realign the visual world (
Figure 1A–C). Specifically, when the human head makes a rolling movement, a compensatory torsional eye movement in the opposite direction is generated, thereby keeping the visual field stable.
However, torsional eye movements induced by the vestibulo-ocular reflex can be prevented by fixating the eyeballs of anaesthetised cats to a metal ring. In these cats, the receptive fields of a proportion of neurons in the visual cortex tilted when the cat’s head was tilted (roll motion), compared to when it was in an upright position [
1,
2,
3]. These results suggest that another mechanism might exist to centrally process vestibular information when the vestibulo-ocular reflex is disrupted (
Figure 1D).
This suggestion is not without controversy: Receptive fields did not tilt as a function of head roll in cats that were not anaesthetised and did not have their eyeballs fixated [
4]. In those cats, receptive field tilt did occur, but it was uncorrelated with head roll. It was thus suggested that the tilting of receptive fields could have been an effect of non-specific arousal (with the head movement as an arousing stimulus), or could be interpreted as “broadening of direction preference”. Therefore, it remains unclear whether central processing of vestibular input influences visual stability beyond its role in the vestibulo-ocular reflex.
Ideally, the hypothesised central processing of vestibular information could be directly tested by comparing the effects of vestibular input on visual tilt and on ocular torsion. If the vestibulo-ocular reflex is sufficient for stabilising the visual world, the ocular torsion induced by vestibular input should be linearly related to the induced visual tilt. Such a study exists: Zink and colleagues reported that both ocular torsion and visual tilt increase with vestibular input [
5].
Human research participants do not normally have their eyes fixated to metal rings while their body is tilted. Instead, Zink and colleagues induced vestibular input by means of galvanic vestibular stimulation. This technique has been known for over a hundred years [
6], with the first reports dating back to around 1900 [
7,
8]. Galvanic vestibular stimulation is an electric current (usually applied via both mastoids) that stimulates the vestibular neuronal afferents and is known to induce spontaneous nystagmus (at intensities over 3 mA) and ocular torsion [
5,
9,
10].
Eye movements induced by galvanic vestibular stimulation occur due to electrical stimulation of semicircular canal afferents [
11]. Although otolith activation occurs at higher stimulation intensity, its contribution to eye movement varies between individuals [
5,
9]. A contemporary model on the contribution of semicircular canal and otolith activation is provided by Day and colleagues [
12], who built on earlier models by the same group [
13] and others [
10].
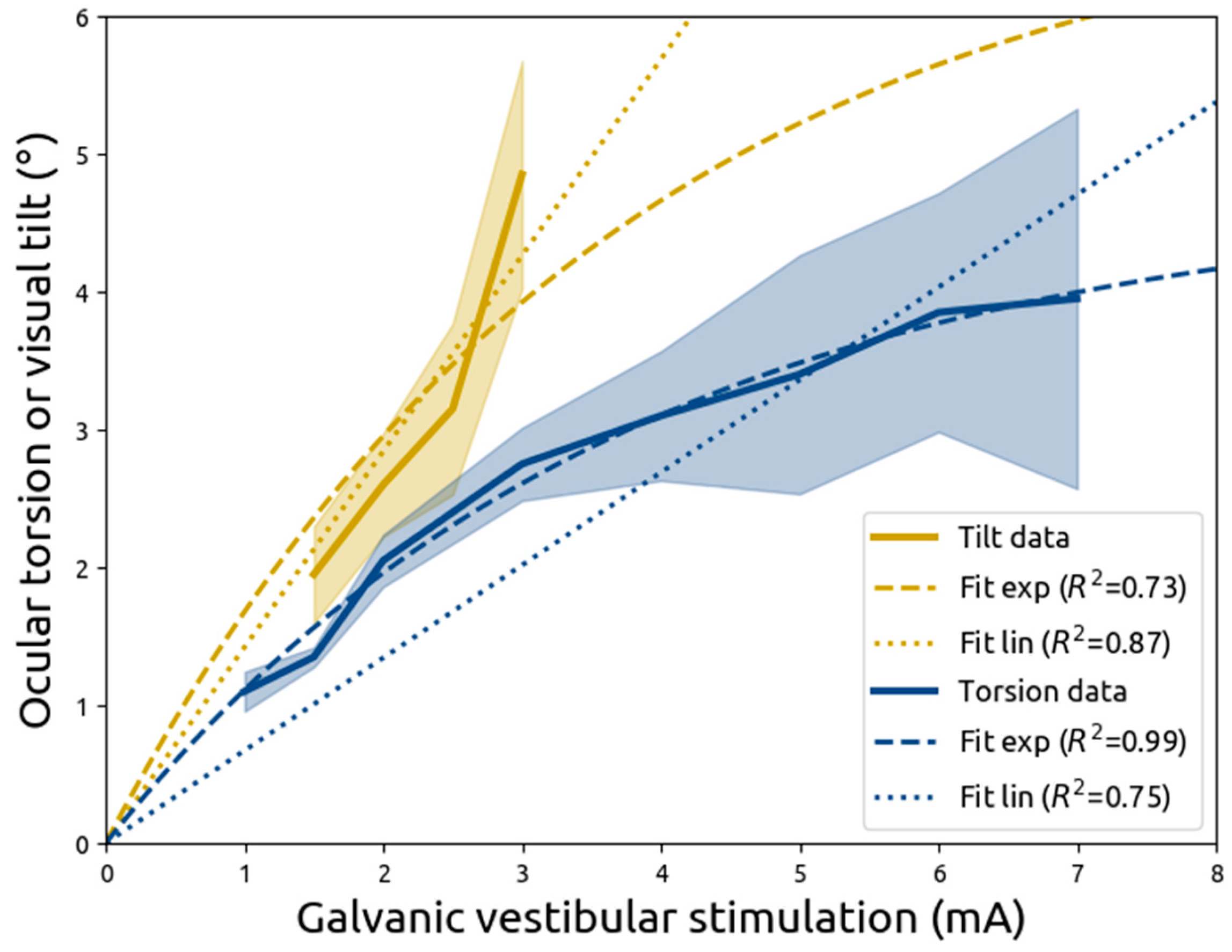
In the aforementioned study by Zink and colleagues, it was reported that both ocular torsion and visual tilt increase with galvanic vestibular stimulation at higher current intensities [
5]. Importantly, induced ocular torsion would have invoked visual tilt merely due to the rotation of both eyeballs. However, from the analysis by Zink and colleagues, it is unclear whether ocular torsion could account for all the induced visual tilt, or whether part of the visual tilt could have been ascribed to the central processing of the vestibular input.
The current study aims to investigate the relationship between ocular torsion and visual tilt through re-analysing the results of Zink and colleagues by modelling the contributions of vestibular stimulation on ocular torsion and visual tilt. Specifically, the models introduced here take into account a hypothesised limit on ocular torsion. If such a limit exists, visual tilt will increasingly depend on central processing when vestibular input increases. Alternatively, if both ocular torsion and visual tilt are linearly related to vestibular input, it would suggest the vestibulo-ocular reflex is sufficient, and no central processing of vestibular input is required to stabilise vision.
2. Materials and Methods
2.1. Zink et al. (1998)
Relevant data was extracted from Zink and colleagues [
5], who conveniently provided it in tables with descriptive statistics of both visual tilt and ocular torsion as a function of vestibular stimulation intensity (reproduced in
Table 1). They used electrodes taped to both mastoids to deliver a unipolar direct current. During a trial, the polarity and intensity of the stimulation was kept constant, but they could be varied between trials. Specifically, Zink and colleagues tested both left-anodal and right-anodal stimulation, and they applied current intensities between 1 and 7 mA. Not all participants received all current intensities, presumably because the stimulation caused discomfort or pain. The numbers of participants in each cell of Zink and colleagues’ design are reported in
Table 1.
Stimulation trials lasted for 5 s, during which static ocular torsion was measured using a laser-scanning opthalmoscope that recorded the fundus in both eyes on video.
To measure visual tilt, participants were positioned in front of a half-open dome with a diameter of 60 cm that completely covered their visual field and was covered with a pattern of randomly placed coloured dots. The dome prevented participants from using straight lines in the environment as references. Perceived visual tilt was measured using a centrally presented line that participants could adjust to the level of rotation that they perceived during galvanic vestibular stimulation.
Ocular torsion occurred towards the anode (counter-clockwise under left anodal stimulation, clockwise in right anodal stimulation). Visual tilt occurred away from the anode (clockwise in left anodal stimulation and counter-clockwise in right anodal stimulation). The results from Zink and colleagues are reproduced in
Table 1.
For all analyses presented here, the left and right anodal stimulation was averaged within each stimulation intensity. One measurement that was inconsistently reported by Zink and colleagues has been omitted from the current study. Specifically, Zink and colleagues refer to a measurement of visual tilt at a stimulation intensity of 4.5 mA. However, their visual tilt plot includes a point at 3.5 mA, and the point in question does not appear in their visual tilt table.
2.2. Linear Models
Zink and colleagues fit linear models that describe both ocular torsion and visual tilt as a function of stimulation intensity (Equations (1) and (2)). Free variables
a and
b in these equations determine the slope and intercept of the function. It should also be noted that the
a and
b parameters in the ocular torsion equation are independent from those in the visual tilt equation.
in which
Tocular is ocular torsion in degrees,
V is vestibular input in mA,
ao is the slope in degrees per mA, and
bo the intercept in degrees in the relationship between ocular torsion and visual tilt.
Tvisual is visual tilt in degrees,
av is the slope in degrees per mA, and
bv is the intercept in degrees.
2.3. Directly Proportional Linear Models
By accounting for an intercept in Equations (1) and (2), Zink and colleagues allowed the baseline ocular torsion to be different from 0 degrees at 0 mA of stimulation. Because stimulation was applied in two directions, according to Equation (1) the eye is in a different baseline position depending on whether −0 or 0 mA of stimulation is applied. The same is true for Equation (2) and visual tilt. In other words, a non-zero ao parameter in Equation (1) means that the eye is in two different orientations at the same time when no stimulation is applied. The same is true for the av parameter in Equation (2): non-zero values would mean visual perception will be tilted in two directions at the same time when no stimulation is applied.
Evidently, this cannot be true: At 0 mA of stimulation, the eye and visual field should be un-rotated. The best way to account for this is by requiring that vestibular input and ocular torsion (or visual tilt) be directly proportional. This idea was implemented in Equations (3) and (4), which both have only one free variable that determines the slope of the function.
in which
Tocular is ocular torsion in degrees,
V is vestibular input in mA, and
ao is the slope in degrees per mA in the relationship between ocular torsion and visual tilt.
Tvisual is visual tilt in degrees, and
av is the slope in degrees per mA in the relationship between visual tilt and vestibular input.
2.4. Exponential Model of Ocular Torsion and Resulting Visual Tilt
If ocular torsion is indeed limited by an upper bound, neither linear model described above would describe the relationship between galvanic vestibular stimulation and ocular torsion accurately. Instead, as vestibular input increases, the slope of the increase in ocular torsion is expected to decrease and to become 0 when an asymptote is reached. Equation (5) describes such a relationship, in which at high levels of vestibular input, ocular torsion will be no higher than asymptote b.
If perceived visual tilt is solely a product of ocular torsion, its relationship with vestibular input should be the same as that of ocular torsion, as described by Equation (6).
in which
Tocular is the ocular torsion in degrees,
V is the vestibular input in mA,
ao determines the slope of the function (with lower numbers reflecting a steeper slope), and
bo determines the asymptote of the function (preventing
Tocular to ever rise above
bo, regardless of the value of
V).
Tvisual is the visual tilt in degrees, and a
v and
bv have the same purpose as a
o and b
o, but can be of different values.
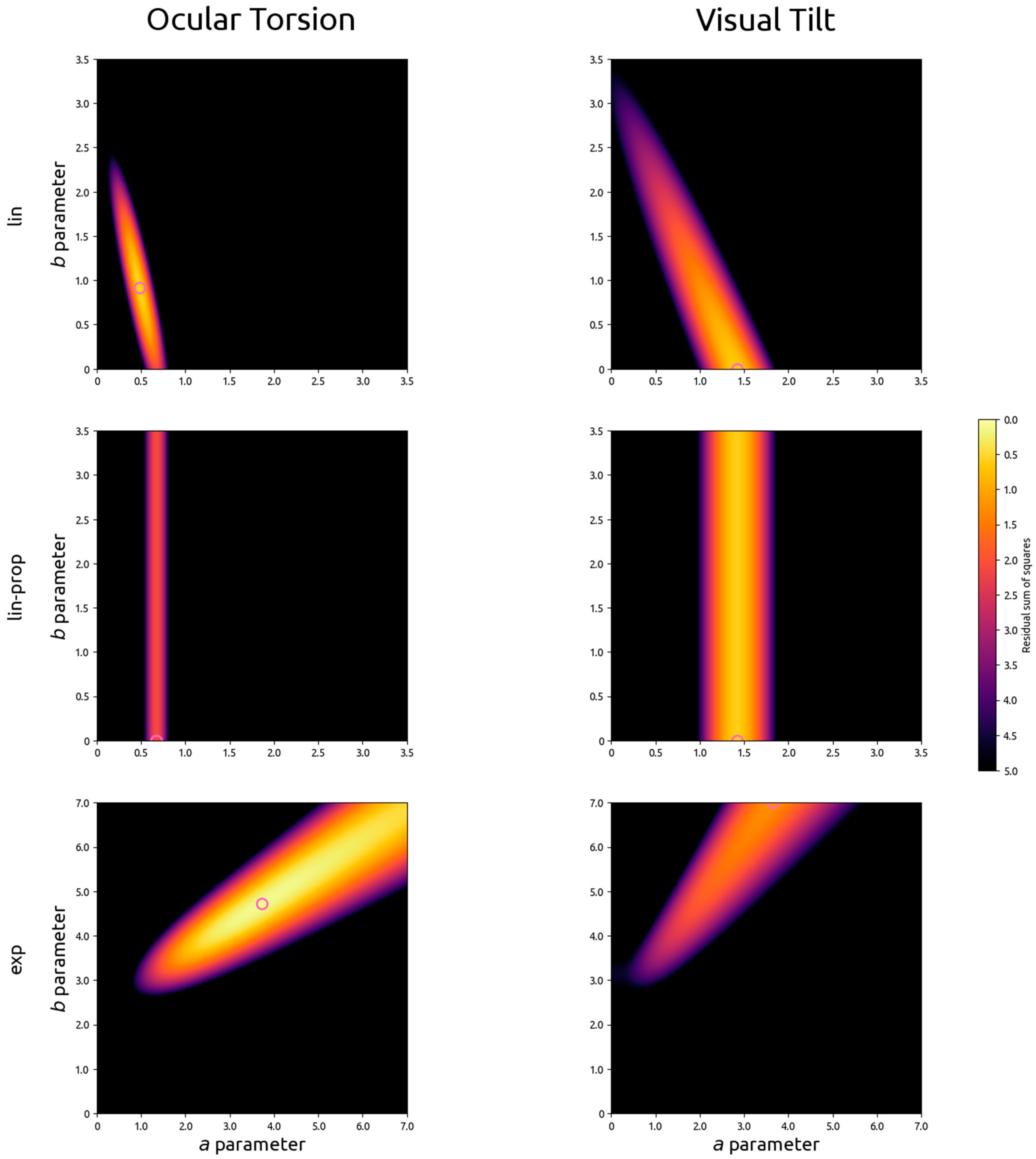
2.5. Curve Fitting
To fit Equations (1)–(6) to the data reported by Zink and colleagues (reproduced in
Table 1), the unsigned results from left-anodal and right-anodal stimulation were first averaged. Then, using least squares estimation in a full exploration of parameter space, the optimal combination of parameter values were assessed for the three types of models outlined above (linear, directly proportional linear, and exponential) for ocular torsion and visual tilt independently.
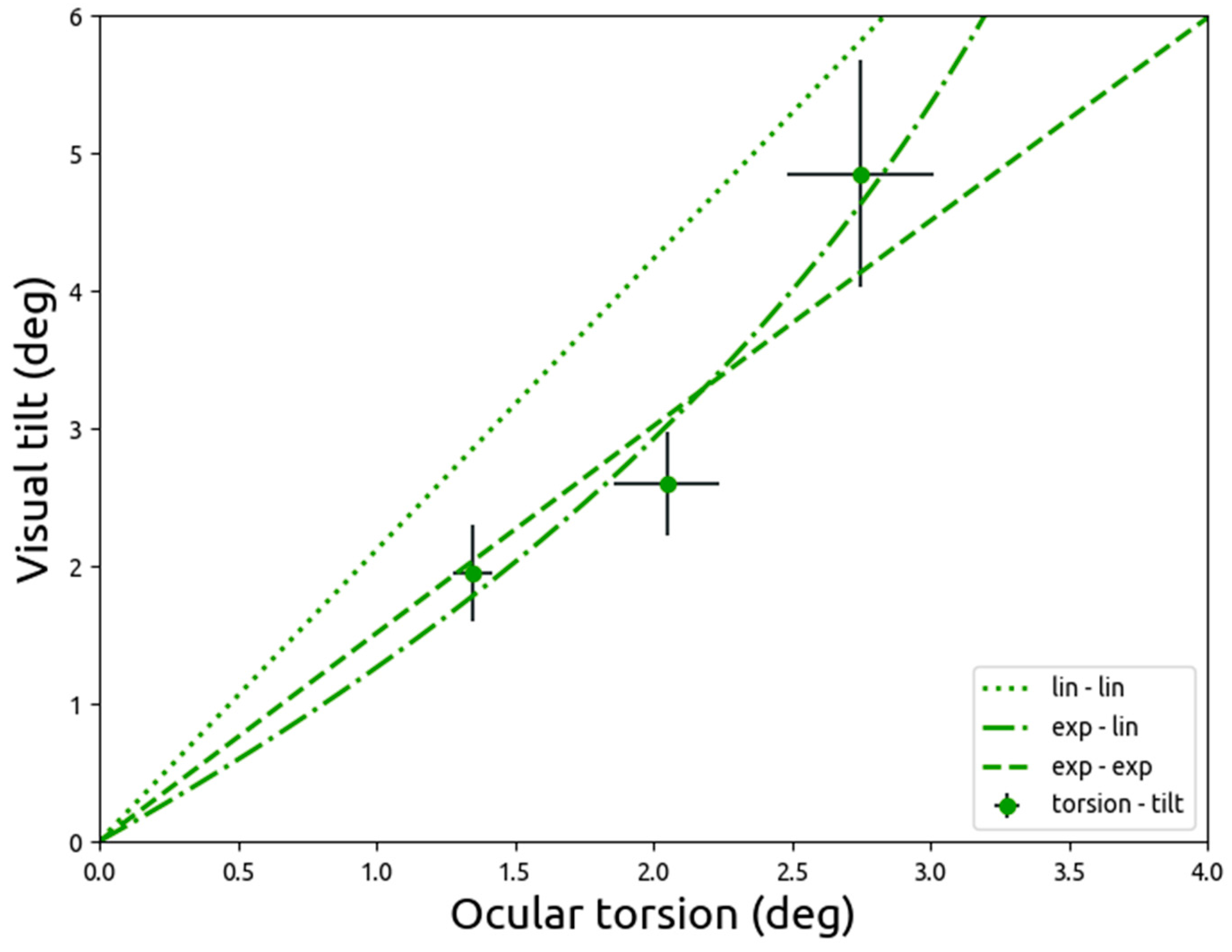
4. Discussion
In the current study, data from Zink and colleagues (1998) was reanalysed using models that more accurately reflected biology, specifically the non-tilted position of the eye at no vestibular input. Zink and colleagues applied galvanic vestibular stimulation, using a unipolar and direct current, and measured either ocular torsion or visual tilt. In the current study, it is demonstrated that a directly proportional linear model fits the data from both measures relatively well. However, ocular torsion is better described by a model with an exponentially decreasing slope that moves towards an asymptote. These results show that with increasing vestibular stimulation, ocular torsion slopes down, whereas visual tilt keeps increasing linearly. The lack of a linear relationship between the vestibular effects on ocular torsion and on visual tilt suggests that ocular torsion is not the sole contributor of visual tilt, but that instead vestibular input could be processed centrally to maintain visual stability.
4.1. Central Processing of Vestibular Information
In multi-sensory research, the most simple explanation for the interaction between two senses (e.g., the vestibular and visual system) is often stochastic resonance [
18]. Stochastic resonance occurs when the general level of activation of multi-sensory neurons is heightened by a stimulus (e.g., galvanic vestibular stimulation or auditory noise), which leads to a higher sensitivity to faint stimuli from another sense (e.g., tactile information) that on its own would lead to sub-threshold activation for detection. An example of such a study is by Ferrè and colleagues [
19], who showed that galvanic vestibular stimulation indeed leads to a higher sensitivity for a faint tactile stimulus. They offer an explanation based in stochastic resonance, hypothesising simultaneous activation of bimodal neurons in parietal opperculum. However, the data from Zink and colleagues re-analysed here cannot be explained by stochastic resonance, as the observed visual tilt was direction-dependent on the stimulation’s current direction.
An alternative explanation is that vestibular information is processed in the visual cortex. When the heads of anaesthetised cats with fixated eyeballs are rolled, the receptive fields of a proportion of neurons in their visual cortex tilt as a result [
1,
2,
3]. This suggests that a mechanism might exist by which head roll that is not otherwise compensated for is compensated for by the rotation of receptive fields.
Contrary to the above, non-anaesthetised cats with non-fixated eyeballs do show tilt in receptive fields in a proportion of neurons in visual cortex, but they are uncorrelated with head roll [
4]. It was suggested that receptive field tilting was a response to general arousal rather than a systematic processing of visual tilt.
4.2. Alternative Explanations for a Non-Linear Relationship between Vestibular Input and Ocular Torsion
Although Zink and colleagues used direct current stimulation, others have employed vestibular stimulation with a sinusoidally alternating current and have instead argued that visual tilt can be completely accounted for by ocular torsion [
20]. In addition, using 100 ms pulses, Aw and colleagues found a linear relationship between current intensity and the velocity of ocular torsion [
21]. In sum, a linear relationship between vestibular input and ocular torsion does exist when pulsed or alternating current galvanic vestibular stimulation is employed.
A direct investigation of the non-linear properties of the torsional response to natural vestibular stimulation (by means of head rotation) is described by Schneider and colleagues, who modelled the gain and intensity of torsional nystagmus [
22]. Specifically, they demonstrate that nystagmus was present during low-frequency (such as Zink and colleagues’ stimulation), but not high-frequency, stimulation, and they argue that it is the contribution of nystagmus that causes the non-linear relationship between vestibular input and ocular torsion. In the mechanism that Schneider and colleagues propose, vestibular input induces nystagmus, which is in turn centrally processed. This process in turn interferes with the vestibularly induced generation of ocular torsion, resulting in a non-linear relationship between vestibular input and ocular torsion.
4.3. Vestibular Effects on Attention
Another potentially interesting role of vestibular input is highlighted by Shuren and colleagues, who rotated participants in a revolving chair before asking them to perform a line bisection task. After leftward rotation, participants showed an increased leftward bisection error, suggesting that vestibular input might induce an attentional bias [
23].
Further evidence for a potential role of vestibular functioning in spatial attention is provided by studies of vestibular stimulation in neglect syndrome. Neglect syndrome occurs primarily after damage to right parietal cortex, and patients display a strong attentional bias towards ipsilesional space. Neglect patients but not control patients (with similar lesions but without neglect) show deviations of their perceived visual vertical during roll head movements [
24], and exhibit a larger deviation of subjective visual vertical and horizontal [
25]. Furthermore, neglect symptoms decrease during [
26] or immediately after caloric vestibular stimulation, but these effects do not linger for more than 15 min [
27].






{kind=link}
{kind=link}
{kind=link}
{kind=link}