

Anaerobic Digestion of Cuttings from Grassland in Protected Landscape Areas



Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Area and Fieldwork
2.1.1. Lowland Hay Meadows
2.1.2. Extensively Managed Orchard Meadows
2.2. Laboratory Analysis
2.2.1. Sample Preparation and Chemical Composition Analyses
2.2.2. Biochemical Methane Potential Test
2.2.3. Inoculum
2.2.4. Kinetics of Methane Formation Process
2.3. Statistical Analysis
3. Results and Discussion
3.1. Biomass Yields of Lowland Hay and Orchard Meadows during the Growing Season
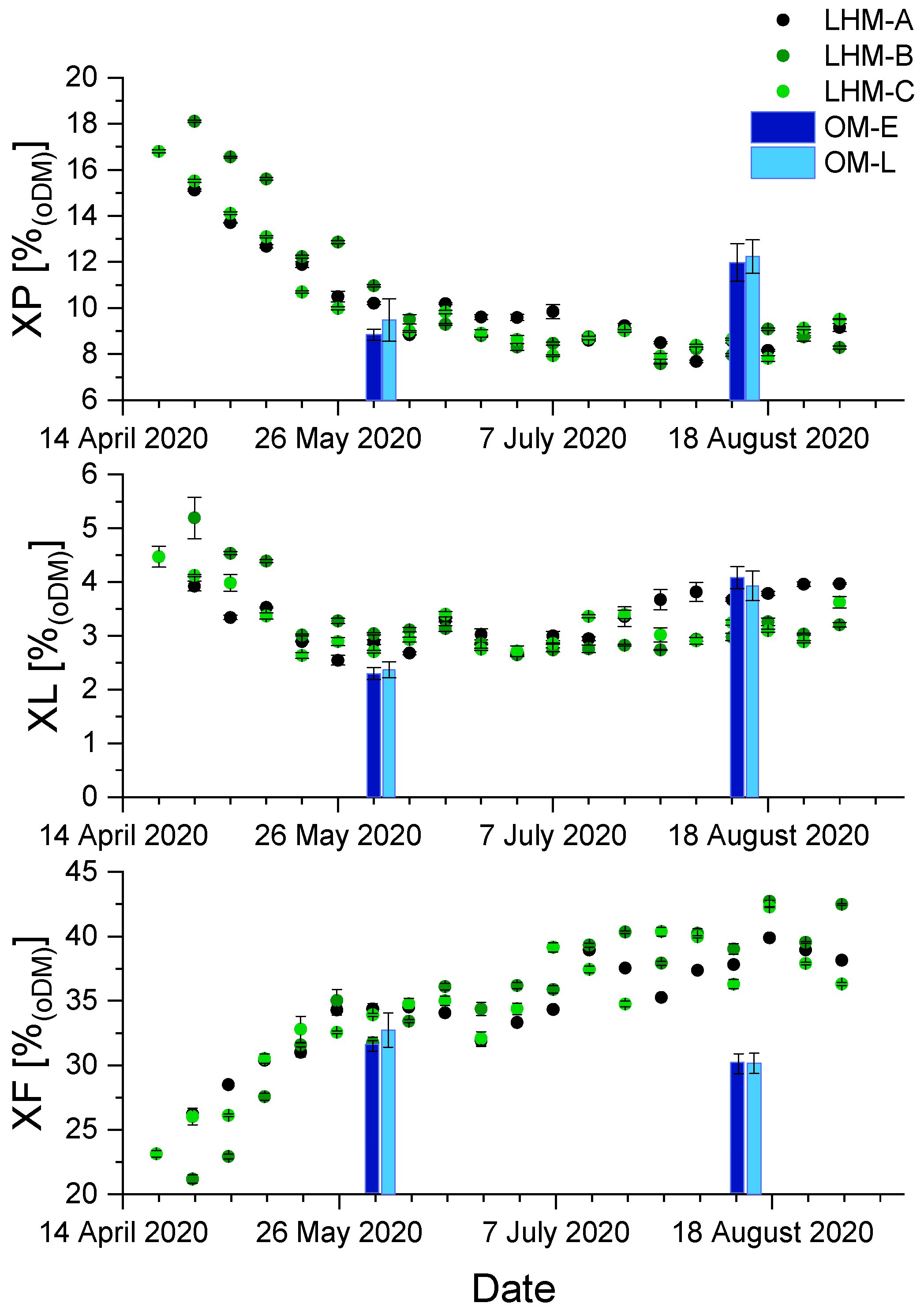
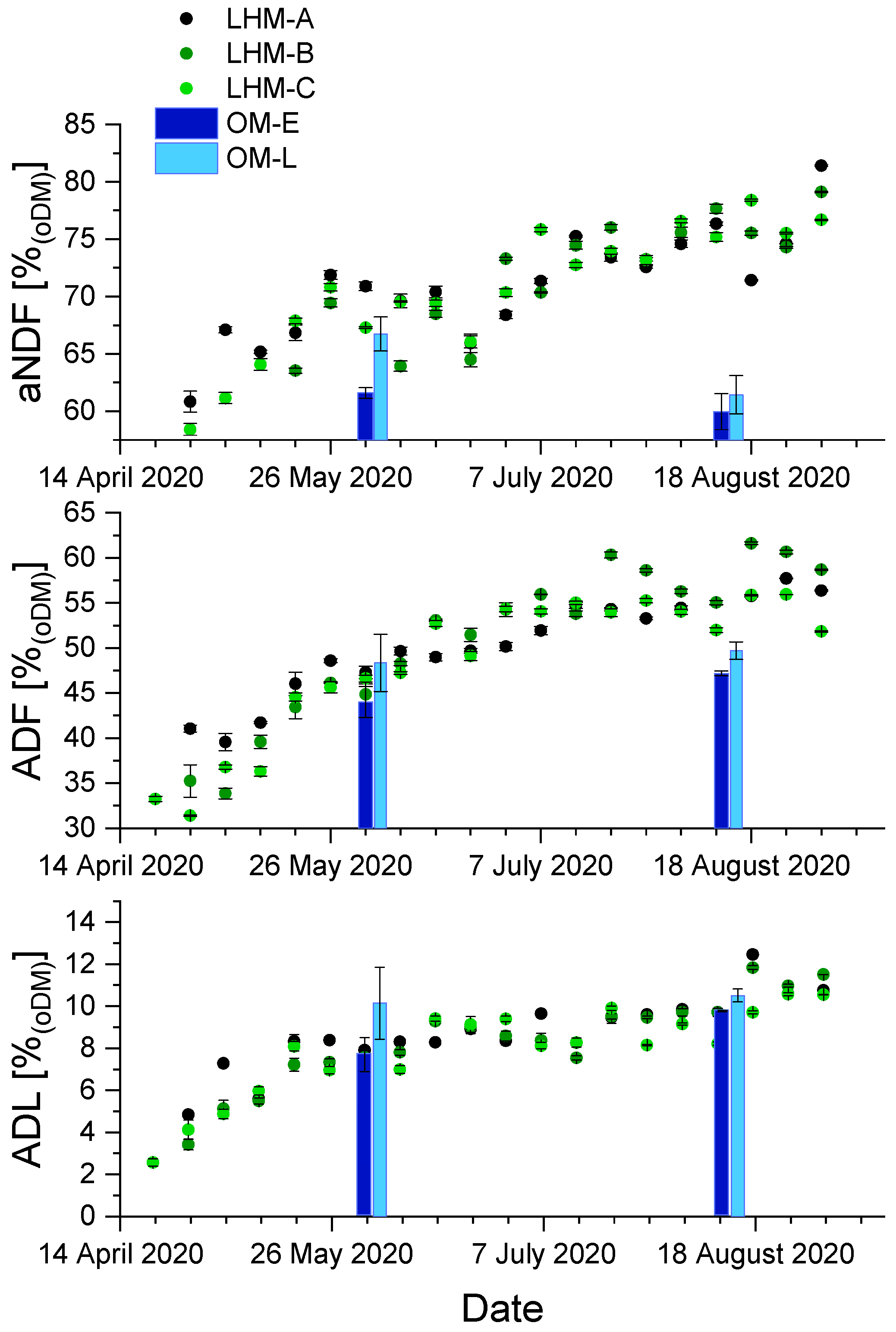
3.2. Chemical Composition of Harvested Biomass during the Vegetation Period
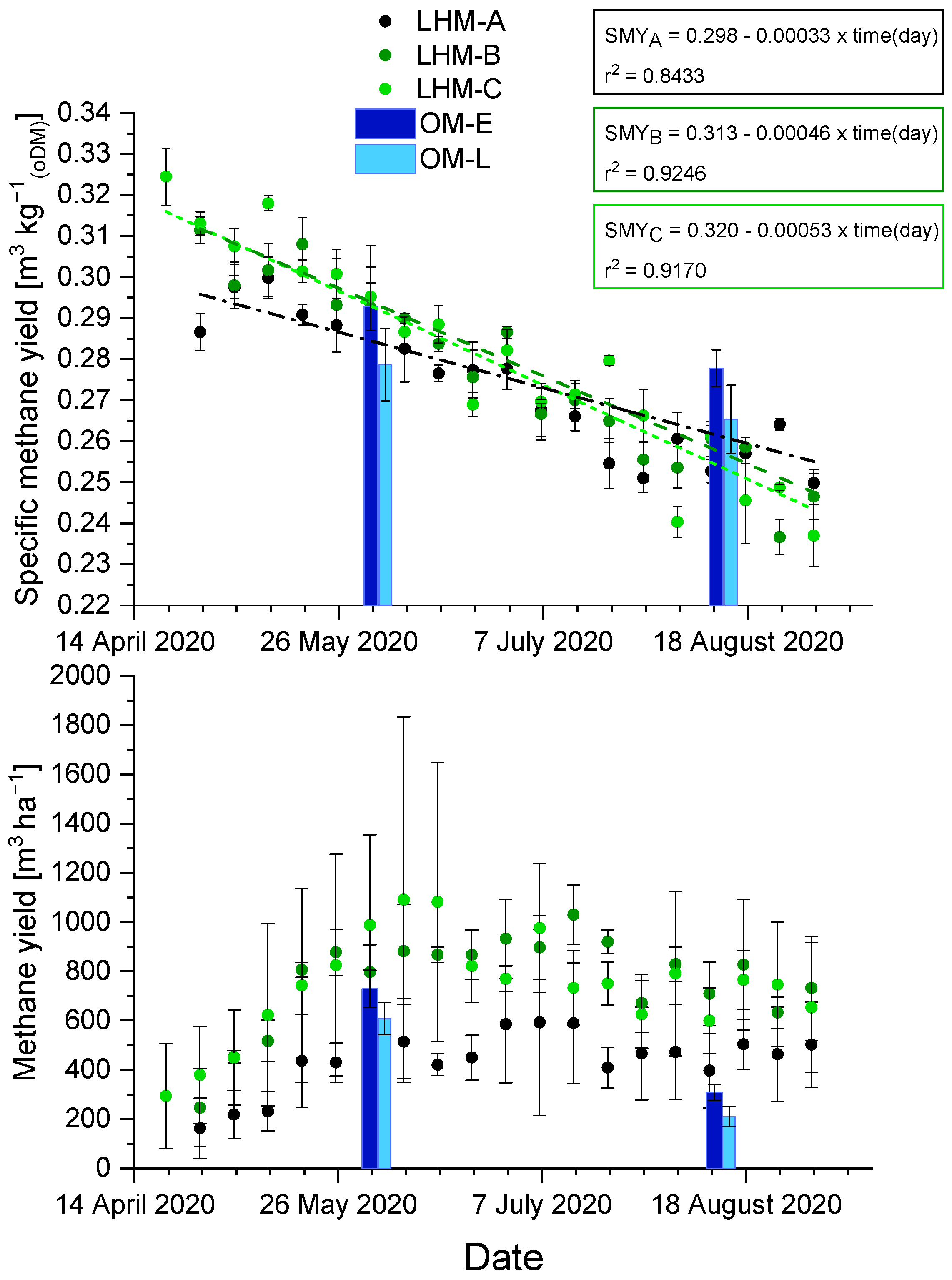
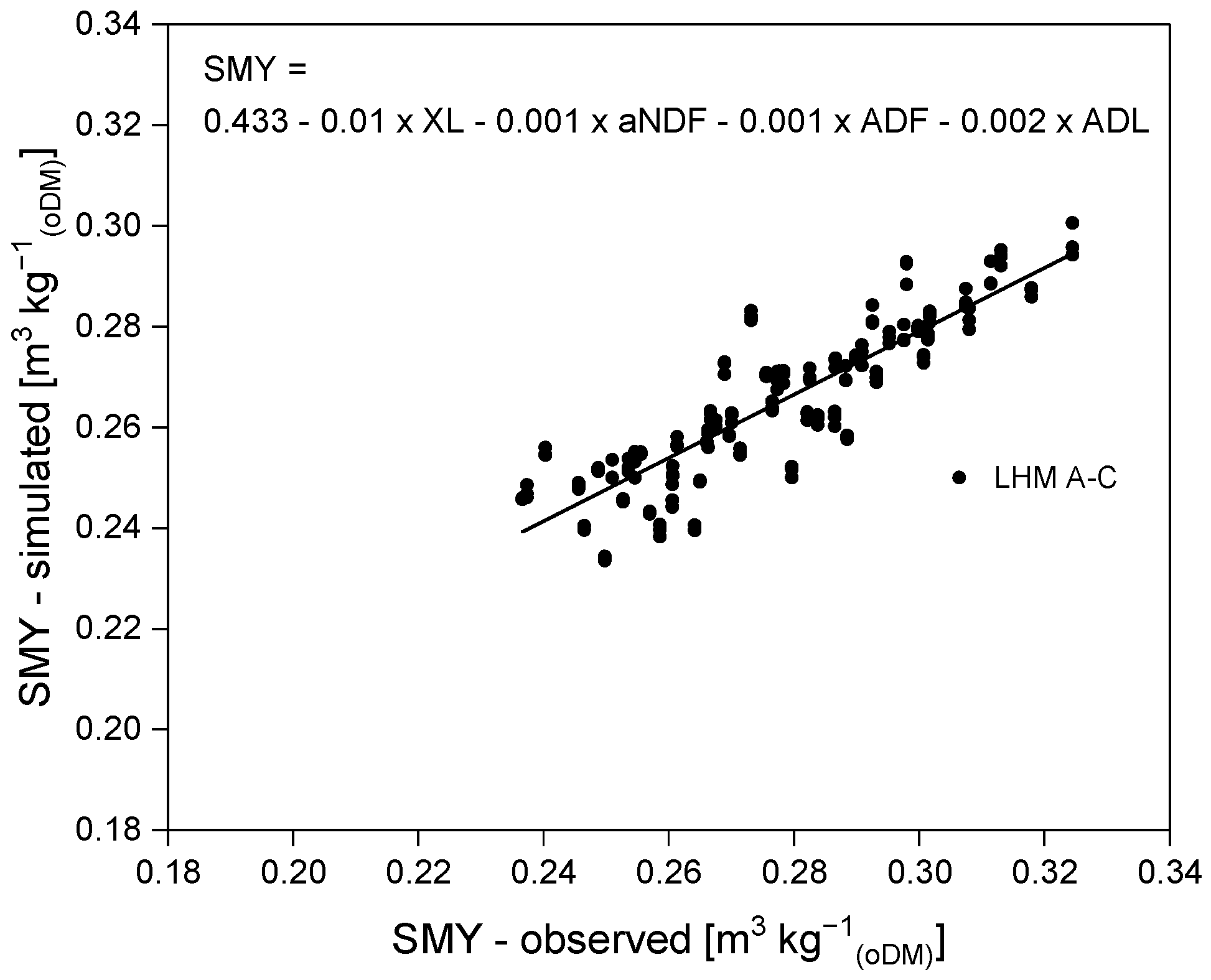
3.3. Methane Yield Potential
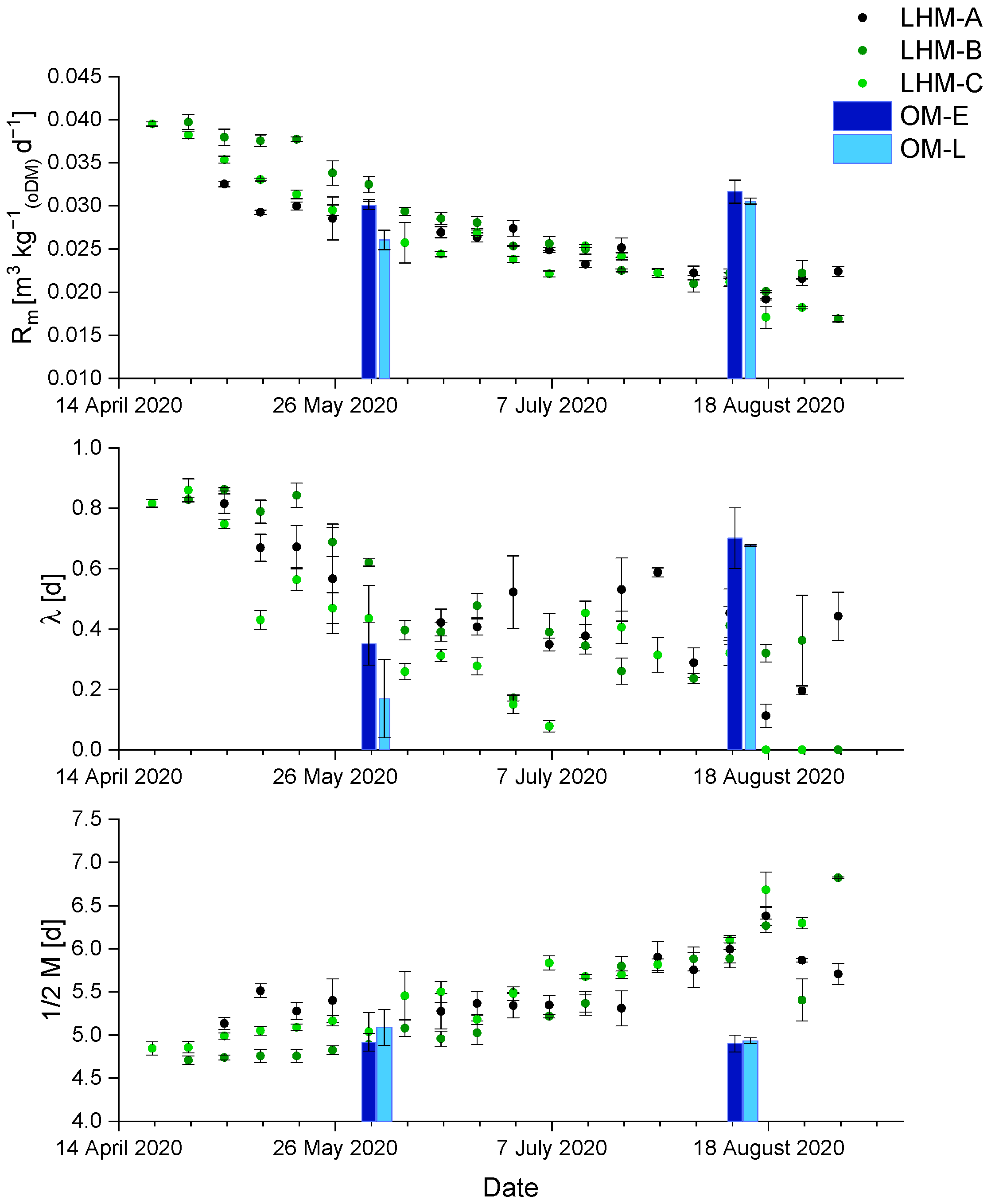
3.4. Kinetics Analysis of Methane Formation (Gompertz)
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Finck, P.; Heinze, S.; Raths, U.; Riecken, U.; Ssymank, A. Rote Liste der Gefährdeten Biotoptypen Deutschlands: Dritte Fortgeschriebene Fassung 2017; Naturschutz und Biologische Vielfalt 156; Bundesamt für Naturschutz: Bonn, Germany, 2017. [Google Scholar] [CrossRef]
- Deutsche UNESCO-Kommission e.V. Bundesweites Verzeichnis Immaterielles Kulturerbe Nationwide Inventory of Intangible Cultural Heritage in Germany: Jubiläum 20 Jahre Konvention 10 Jahre Verzeichnis. Immaterielles Kulturerbe Wissen. Können. Weitergeben. Available online: https://www.unesco.de/publikationen?page=2#row-9911 (accessed on 24 November 2023).
- Plieninger, T.; Levers, C.; Mantel, M.; Costa, A.; Schaich, H.; Kuemmerle, T. Patterns and drivers of scattered tree loss in agricultural landscapes: Orchard meadows in Germany (1968–2009). PLoS ONE 2015, 10, e0126178. [Google Scholar] [CrossRef]
- Forejt, M.; Syrbe, R.-U. The current status of orchard meadows in Central Europe: Multi-source area estimation in Saxony (Germany) and the Czech Republic. Morav. Geogr. Rep. 2019, 27, 217–228. [Google Scholar] [CrossRef]
- Kilian, S. Streuobst-unverzichtbar für unsere Kulturlandschaft. In Wildtiere in der Agrarlandschaft 14. Kulturlandschaftstag; Bayrische Landesanstalt für Landwirtschaft: Freising, Germany, 2016; pp. 29–39. [Google Scholar]
- Steffan-Dewenter, I.; Leschke, K. Effects of habitat management on vegetation and above-ground nesting bees and wasps of orchard meadows in Central Europe. Biodivers. Conserv. 2003, 12, 1953–1968. [Google Scholar] [CrossRef]
- Herzog, F. Streuobst: A traditional agroforestry system as a model for agroforestry development in temperate Europe. Agrofor. Syst. 1998, 42, 61–80. [Google Scholar] [CrossRef]
- Ministerium für Ländlichen Raum und Verbraucherschutz. Streuobstkonzeption Baden-Württemberg: Aktiv für Reichtum und Viefalt Unserer Streuobstlandschaften. 2015. Available online: https://mlr.baden-wuerttemberg.de/fileadmin/redaktion/m-mlr/intern/dateien/publikationen/Streuobstkonzeption.pdf (accessed on 9 December 2022).
- Borngraeber, S.; Krismann, A.; Schmieder, A. Ermittlung der Streuobstbestände Baden-Württembergs durch Automatisierte Fernerkundungsverfahren; Naturschutz und Landschaftspflege Baden-Württemberg No. 81. 2023. Available online: https://pudi.lubw.de/detailseite/-/publication/10128 (accessed on 16 November 2023).
- Ministerium für Ländlichen Raum und Verbraucherschutz. Das Rahmenkonzept Band 2: Unsere Strategie: Leitbilder, Entwicklungsziele, Leitprojekte (Kurzfassung). 2012. Available online: https://www.biosphaerengebiet-alb.de/fileadmin/page_images/biosphaerengebiet/rahmenkonzept/Band_2_Unsere_Strategie_24.08.2012.pdf (accessed on 21 November 2023).
- Ashwood, F. Lowland Calcareous Grassland: Creation and Management in Land Regeneration; The Land Regeneration and Urban Greenspace Research Group: Farnham, UK, 2014. [Google Scholar]
- Karlik, P.; Poschlod, P. History or abiotic filter: Which is more important in determining the species composition of calcareous grasslands? Preslia 2009, 81, 321–340. [Google Scholar]
- LUBW. FFH-Lebensraumtyp 6510: Magere Flachland-Mähwiese. Available online: https://www.lubw.baden-wuerttemberg.de/natur-und-landschaft/6510-magere-flachland-mahwiesen (accessed on 25 November 2023).
- Fischer, M.; Stöcklin, J. Local Extinctions of Plants in Remnants of Extensively Used Calcareous Grasslands 1950–1985. Conserv. Biol. 1997, 11, 727–737. [Google Scholar] [CrossRef]
- Willems, J.H. Phytosociological and geographical survey of Mesohromion communities in Western Europe. Vegetatio 1982, 48, 227–240. [Google Scholar] [CrossRef]
- LfL. Gruber Tabelle zur Fütterung der Milchkühe, Zuchtrinder, Schafe, Ziegen. 2023. Available online: https://www.lfl.bayern.de/publikationen/informationen/040183/index.php (accessed on 18 November 2023).
- Jilg, T.; Aulendorf, L.A.Z.B.W. Fütterung von Hochleistungskühen mit FFH-Grünland–Geht das? Landinfo 2011, 6, 33–36. [Google Scholar]
- Bundesministerium für Umwelt, Naturschutz und nukleare Sicherheit; Bundesamt für Naturschutz Die Lage der Natur in Deutschland: Ergebnisse von EU-Vogelschutz- und FFH-Bericht, Berlin, Bonn. 2020. Available online: https://www.bfn.de/sites/default/files/BfN/natura2000/Dokumente/bericht_lage_natur_2020.pdf (accessed on 14 February 2024).
- Scheibe, U. FFH-Mähwiesen (Flachland- und Bergmähwiesen). Landinfo 2015, 5, 20–23. [Google Scholar]
- NABU. Vollzugsdefizite und Verstöße Gegen das Verschlechterungsverbot bei FFH-Lebensraumtypen auf Grünlandstandorten in Deutschland: Beschwerde des NABU an die Europäische Kommission Wegen Nichtbeachtung des Gemeinschaftsrechts; NABU: Berlin, Germany, 2014. [Google Scholar]
- LAZBW. FFH—Mähwiesen: Grundlagen—Bewirtschaftung—Wiederherstellung, Aulendorf. 2018. Available online: https://pudi.lubw.de/detailseite/-/publication/86574 (accessed on 24 November 2023).
- Dalle Fratte, M.; Montagnoli, A.; Anelli, S.; Armiraglio, S.; Beatrice, P.; Ceriani, A.; Lipreri, E.; Miali, A.; Nastasio, P.; Cerabolini, B.E.L. Mulching in lowland hay meadows drives an adaptive convergence of above- and below-ground traits reducing plasticity and improving biomass: A possible tool for enhancing phytoremediation. Front. Plant Sci. 2022, 13, 1062911. [Google Scholar] [CrossRef]
- Baumann, A.; Bergmann, F.; Neub, M.; Opitz, H.; Rupp, J.; Westermann, K. Naturschutzfachliche Bedeutung, Gefährdungen und Schutzmöglichkeiten der Elzniederung östlich der A5 bei Kenzingen und Herbolzheim. Naturschutz Südlichen Oberrh. 2014, 7, 249–254. [Google Scholar]
- Lüker-Jans, N.; Simmering, D.; Otte, A. The impact of biogas plants on regional dynamics of permanent grassland and maize area—The example of Hesse, Germany (2005–2010). Agric. Ecosyst. Environ. 2017, 241, 24–38. [Google Scholar] [CrossRef]
- Plieninger, T.; Bieling, C.; Ohnesorge, B.; Schaich, H.; Schleyer, C.; Wolff, F. Exploring Futures of Ecosystem Services in Cultural Landscapes through Participatory Scenario Development in the Swabian Alb, Germany. Ecol. Soc. 2013, 18, 39. [Google Scholar] [CrossRef]
- Humbert, J.-Y.; Ghazoul, J.; Walter, T. Meadow harvesting techniques and their impacts on field fauna. Agric. Ecosyst. Environ. 2009, 130, 1–8. [Google Scholar] [CrossRef]
- Van de Poel, D.; Zehm, A. Die Wirkung des Mähens auf die Fauna der Wiesen—Eine Literaturauswertung für den Naturschutz. In Handbuch Naturschutz und Landschaftspflege; Konold, W., Böcker, R., Hampicke, U., Eds.; Wiley: Weinheim, Germany, 2014; pp. 1–19. ISBN 9783527678471. [Google Scholar]
- Unterweger, P.A.; Klammer, J.; Unger, M.; Betz, O. Insect hibernation on urban green land: A winter-adapted mowing regime as a management tool for insect conservation. BioRisk 2018, 13, 1–29. [Google Scholar] [CrossRef]
- Diacon, J.; Bürgi, M.; Dalang, T. Systematisches Review zu Bewirtschaftungseinflüssen auf Trockenwiesen und –Weiden (SR-TWW). Schlussbericht, Birmensdorf. 2011. Available online: https://infohabitat.ch/wp-content/uploads/2019/01/Tw_SystemReview_TwwBewirtschaftungseinfluesse_15022011_de.pdf (accessed on 26 November 2023).
- Dieterich, M.; Dorsch, H.; Bahrs, E. Übersicht Möglicher Biodiversitätsmaßnahmen, Kirchheim unter Teck. 2021. Available online: https://mlr.baden-wuerttemberg.de/fileadmin/redaktion/m-mlr/intern/dateien/PDFs/Biodiv/biodiv-ma%C3%9Fnahmenkatalog_demobetriebe.pdf (accessed on 26 November 2023).
- Dandikas, V.; Heuwinkel, H.; Lichti, F.; Drewes, J.E.; Koch, K. Correlation between Biogas Yield and Chemical Composition of Grassland Plant Species. Energy Fuels 2015, 29, 7221–7229. [Google Scholar] [CrossRef]
- Data and Map Service of the LUBW. Erfassungseinheit Nr.: 6510800046030080; Flurstücksnummer: 7429; Gemarkung Eningen unter Achalm (Landkreis Reutlingen, Germany). Available online: https://udo.lubw.baden-wuerttemberg.de/public/api/processingChain?conditionValuesSetHash=C10A56A&selector=ROOT.Natur%20und%20Landschaft.Natura%202000%20-%20FFH-M%C3%A4hwiesen.nais_map%3Anais_map_z_mw_udo_permalink.sel&processings=nais_map%3Anais_map_mw_erhebungsbogen%2Fnais_map_mw_erhebungsbogen_udo.rpt&sourceOrderAsc=false&columns=020789b6-1f17-4820-a4a7-a5823f7e35b8&offset=0&limit=2147483647&executionConfirmed=false (accessed on 28 November 2023).
- Data and Map Service of the LUBW. Erfassungseinheit Nr.: 6510800046029669; Flurstücksnummer: 4931; Gemarkung Eningen unter Achalm (Landkreis Reutlingen, Germany). Available online: https://udo.lubw.baden-wuerttemberg.de/public/api/processingChain?conditionValuesSetHash=C4950AD&selector=ROOT.Natur%20und%20Landschaft.Natura%202000%20-%20FFH-M%C3%A4hwiesen.nais_map%3Anais_map_z_mw_udo_permalink.sel&processings=nais_map%3Anais_map_mw_erhebungsbogen%2Fnais_map_mw_erhebungsbogen_udo.rpt&sourceOrderAsc=false&columns=0bc54797-738d-4545-aea6-5c88c242b380&offset=0&limit=2147483647&executionConfirmed=false (accessed on 28 November 2023).
- Data and Map Service of the LUBW. Erfassungseinheit Nr.: 6510800046030070; Flurstücksnummer: 7430; Gemarkung Eningen unter Achalm (Landkreis Reutlingen, Germany). Available online: https://udo.lubw.baden-wuerttemberg.de/public/api/processingChain?conditionValuesSetHash=50FC495&selector=ROOT.Natur%20und%20Landschaft.Natura%202000%20-%20FFH-M%C3%A4hwiesen.nais_map%3Anais_map_z_mw_udo_permalink.sel&processings=nais_map%3Anais_map_mw_erhebungsbogen%2Fnais_map_mw_erhebungsbogen_udo.rpt&sourceOrderAsc=false&columns=4e79a835-0123-42aa-8d88-775ba81bddfc&offset=0&limit=2147483647&executionConfirmed=false (accessed on 28 November 2023).
- Piepho, H.-P.; Michel, V.; Williams, E. Neighbor balance and evenness of distribution of treatment replications in row-column designs. Biom. J. 2018, 60, 1172–1189. [Google Scholar] [CrossRef]
- Seither, M.; Engel, S.; King, K.; Martin, E. FFH—Mähwiesen: Grundlagen—Bewirtschaftung—Wiederherstellung, Aulendorf. 2018. Available online: https://fortbildung-lazbw.lgl-bw.de/lazbw/webbasys/download/Shop/2018_GL_lazbw_FFH_Maehwiesen_Grundlagen.pdf (accessed on 28 November 2023).
- VDLUFA. Method Book III—The Chemical Analysis for Feedstuffs; VDLUFA: Darmstadt, Germany, 2012. [Google Scholar]
- European Commission. Commission Regulation 2009/152/EC: Laying Down the Methods of Sampling and Analysis for the Official Control of Feed; Official Journal of the European Union: Luxembourg, 2009. [Google Scholar]
- Energie und Umwelttechnik. Fermentation of Organic Materials—Characterization of the Substrate, Sampling, Collection of Material Data, Fermentation Tests: VDI 4630; Beuth Verlag GmbH: Berlin, Germany, 2016. [Google Scholar]
- Hülsemann, B.; Zhou, L.; Merkle, W.; Hassa, J.; Müller, J.; Oechsner, H. Biomethane Potential Test: Influence of Inoculum and the Digestion System. Appl. Sci. 2020, 10, 2589. [Google Scholar] [CrossRef]
- Lindner, J.; Zielonka, S.; Oechsner, H.; Lemmer, A. Effects of mechanical treatment of digestate after anaerobic digestion on the degree of degradation. Bioresour. Technol. 2015, 178, 194–200. [Google Scholar] [CrossRef]
- Tallowin; Jefferson. Hay production from lowland semi-natural grasslands: A review of implications for ruminant livestock systems. Grass Forage Sci. 1999, 54, 99–115. [Google Scholar] [CrossRef]
- Landesamt für Geologie, Rohstoffe und Bergbau. Bodenkunde; GeoLa Boden; Bodenkundliche Einheiten; Pararendzina und Rendzina aus schuttreichen Fließerden und Hangschutt, Kartiereinheit q21. Available online: https://media.lgrb-bw.de/link/bod3200/q21.pdf (accessed on 28 November 2023).
- Landesamt für Geologie, Rohstoffe und Bergbau (LGRB) Wissen. Böden der Trauf- und Talhänge. LGRBwissen. Available online: https://lgrbwissen.lgrb-bw.de/bodenkunde/mittlere-westliche-alb/bodenlandschaften/boeden-trauf-talhaenge (accessed on 29 November 2023).
- Ludewig, K.; Donath, T.W.; Zelle, B.; Eckstein, R.L.; Mosner, E.; Otte, A.; Jensen, K. Effects of reduced summer precipitation on productivity and forage quality of floodplain meadows at the Elbe and the Rhine River. PLoS ONE 2015, 10, e0124140. [Google Scholar] [CrossRef]
- Zhao, X.-Q.; Zhou, X.-M. Ecological Basis of Alpine Meadow Ecosystem Management in Tibet: Haibei Alpine Meadow Ecosystem Research Station. Ambio 1999, 28, 642–647. [Google Scholar]
- Herrmann, C.; Prochnow, A.; Heiermann, M.; Idler, C. Biomass from landscape management of grassland used for biogas production: Effects of harvest date and silage additives on feedstock quality and methane yield. Grass Forage Sci. 2014, 69, 549–566. [Google Scholar] [CrossRef]
- Michaud, A.; Andueza, D.; Picard, F.; Plantureux, S.; Baumont, R. Seasonal dynamics of biomass production and herbage quality of three grasslands with contrasting functional compositions. Grass Forage Sci. 2012, 67, 64–76. [Google Scholar] [CrossRef]
- Grygierzec, B. Fodder value of hay from extensively used Arrhenatheretum elatioris typicum community. Fragm. Agron. 2012, 29, 37–44. [Google Scholar]
- Thumm, U.; Tonn, B.; Schreiber, K.F.; Brauckmann, H.J.; Broll, G.; Krebs, S.; Poschlod, P. Grünlandaufwüchse—Futter oder Bioenergie; Regionalkultur: Ubstadt-Weiher, Germany, 2009. [Google Scholar]
- Dullau, S.; Rydgren, K.; Kirmer, A.; Jäger, U.G.; Meyer, M.H.; Tischew, S. The Dessau Grassland Experiment—Impact of Fertilization on Forage Quality and Species Assembly in a Species-Rich Alluvial Meadow. Agriculture 2021, 11, 339. [Google Scholar] [CrossRef]
- Bruinenberg, M.H.; Valk, H.; Korevaar, H.; Struik, P.C. Factors affecting digestibility of temperate forages from seminatural grasslands: A review. Grass Forage Sci. 2002, 57, 292–301. [Google Scholar] [CrossRef]
- Meserszmit, M.; Swacha, G.; Pavlů, L.; Pavlů, V.; Trojanowska-Olichwer, A.; Kącki, Z. Species composition of semi-natural mesic grasslands as a factor influencing the methane yield of plant biomass (Central Europe). GCB Bioenergy 2022, 14, 54–64. [Google Scholar] [CrossRef]
- Khan, R.J.; Lau, C.Y.; Guan, J.; Lam, C.H.; Zhao, J.; Ji, Y.; Wang, H.; Xu, J.; Lee, D.-J.; Leu, S.-Y. Recent advances of lignin valorization techniques toward sustainable aromatics and potential benchmarks to fossil refinery products. Bioresour. Technol. 2022, 346, 126419. [Google Scholar] [CrossRef] [PubMed]
- Mankar, A.R.; Modak, A.; Pant, K.K. Recent Advances in the Valorization of Lignin: A Key Focus on Pretreatment, Characterization, and Catalytic Depolymerization Strategies for Future Biorefineries. Adv. Sustain. Syst. 2022, 6, 2100299. [Google Scholar] [CrossRef]
- Agregán, R.; Lorenzo, J.M.; Kumar, M.; Shariati, M.A.; Khan, M.U.; Sarwar, A.; Sultan, M.; Rebezov, M.; Usman, M. Anaerobic Digestion of Lignocellulose Components: Challenges and Novel Approaches. Energies 2022, 15, 8413. [Google Scholar] [CrossRef]
- Li, W.; Khalid, H.; Zhu, Z.; Zhang, R.; Liu, G.; Chen, C.; Thorin, E. Methane production through anaerobic digestion: Participation and digestion characteristics of cellulose, hemicellulose and lignin. Appl. Energy 2018, 226, 1219–1228. [Google Scholar] [CrossRef]
- Van Soest, P.J. Nutritional Ecology of the Ruminant, 2nd ed.; Cornell University Press: Ithaca, NY, USA, 1994; ISBN 978-1-5017-3235-5. [Google Scholar]
- Buxton, D.R. Quality-related characteristics of forages as influenced by plant environment and agronomic factors. Anim. Feed Sci. Technol. 1996, 59, 37–49. [Google Scholar] [CrossRef]
- Dal Prà, A.; Davolio, R.; Immovilli, A.; Burato, A.; Ronga, D. Plant Composition and Feed Value of First Cut Permanent Meadows. Agronomy 2023, 13, 681. [Google Scholar] [CrossRef]
- Rath, J.; Heuwinkel, H.; Herrmann, A. Specific Biogas Yield of Maize Can Be Predicted by the Interaction of Four Biochemical Constituents. BioEnergy Res. 2013, 6, 939–952. [Google Scholar] [CrossRef]
- Lymperatou, A.; Engelsen, T.K.; Skiadas, I.V.; Gavala, H.N. Prediction of methane yield and pretreatment efficiency of lignocellulosic biomass based on composition. Waste Manag. 2023, 155, 302–310. [Google Scholar] [CrossRef]
- Mezule, L.; Strazdina, B.; Dalecka, B.; Skripsts, E.; Juhna, T. Natural Grasslands as Lignocellulosic Biofuel Resources: Factors Affecting Fermentable Sugar Production. Energies 2021, 14, 1312. [Google Scholar] [CrossRef]
- Hoiß, B.; Berg, M.; Krämer, M. Die Herbstzeitlose im extensiven Grünland. Anliegen Nat. 2022, 3, 123–126. [Google Scholar]
- Chizzola, R.; Bassler-Binder, G.; Winter, S.; Zebeli, Q.; Kriechbaum, M. Persistence of alkaloids of typical poisonous plants autumn crocus and marsh ragwort in grass silage. Vet. Med. Austria 2015, 102, 285–292. [Google Scholar]
- Winter, S.; Penker, M.; Kriechbaum, M. Integrating farmers’ knowledge on toxic plants and grassland management: A case study on Colchicum autumnale in Austria. Biodivers. Conserv. 2011, 20, 1763–1787. [Google Scholar] [CrossRef]
- Wiggering, H.; Diekötter, T.; Donath, T.W. Regulation of Jacobaea vulgaris by varied cutting and restoration measures. PLoS ONE 2022, 17, e0248094. [Google Scholar] [CrossRef]
- Heller, R.; Brandhorst, C.; Hülsemann, B.; Lemmer, A.; Oechsner, H. Comparison of Different Mechanical Pretreatment Methods for the Anaerobic Digestion of Landscape Management Grass. Energies 2023, 16, 8091. [Google Scholar] [CrossRef]
- Khan, M.U.; Usman, M.; Ashraf, M.A.; Dutta, N.; Luo, G.; Zhang, S. A review of recent advancements in pretreatment techniques of lignocellulosic materials for biogas production: Opportunities and Limitations. Chem. Eng. J. Adv. 2022, 10, 100263. [Google Scholar] [CrossRef]
- Rodriguez, C.; Alaswad, A.; Benyounis, K.Y.; Olabi, A.G. Pretreatment techniques used in biogas production from grass. Renew. Sustain. Energy Rev. 2017, 68, 1193–1204. [Google Scholar] [CrossRef]
- Meyer, A.; Raju, C.S.; Kucheryavskiy, S.; Holm-Nielsen, J.B. The energy balance of utilising meadow grass in Danish biogas production. Resour. Conserv. Recycl. 2015, 104, 265–275. [Google Scholar] [CrossRef]
- Meserszmit, M.; Chrabąszcz, M.; Chylińska, M.; Szymańska-Chargot, M.; Trojanowska-Olichwer, A.; Kącki, Z. The effect of harvest date and the chemical characteristics of biomass from Molinia meadows on methane yield. Biomass Bioenergy 2019, 130, 105391. [Google Scholar] [CrossRef]
- Amon, T.; Amon, B.; Kryvoruchko, V.; Zollitsch, W.; Mayer, K.; Gruber, L. Biogas production from maize and dairy cattle manure—Influence of biomass composition on the methane yield. Agric. Ecosyst. Environ. 2007, 118, 173–182. [Google Scholar] [CrossRef]
- Oslaj, M.; Mursec, B.; Vindis, P. Biogas production from maize hybrids. Biomass Bioenergy 2010, 34, 1538–1545. [Google Scholar] [CrossRef]
- Melts, I.; Normak, A.; Nurk, L.; Heinsoo, K. Chemical characteristics of biomass from nature conservation management for methane production. Bioresour. Technol. 2014, 167, 226–231. [Google Scholar] [CrossRef]
- Zheng, Y.; Zhao, J.; Xu, F.; Li, Y. Pretreatment of lignocellulosic biomass for enhanced biogas production. Prog. Energy Combust. Sci. 2014, 42, 35–53. [Google Scholar] [CrossRef]
- Tsapekos, P.; Kougias, P.G.; Egelund, H.; Larsen, U.; Pedersen, J.; Trénel, P.; Angelidaki, I. Mechanical pretreatment at harvesting increases the bioenergy output from marginal land grasses. Renew. Energy 2017, 111, 914–921. [Google Scholar] [CrossRef]
- Dahunsi, S.O. Mechanical pretreatment of lignocelluloses for enhanced biogas production: Methane yield prediction from biomass structural components. Bioresour. Technol. 2019, 280, 18–26. [Google Scholar] [CrossRef]
- Herrmann, C.; Idler, C.; Heiermann, M. Biogas crops grown in energy crop rotations: Linking chemical composition and methane production characteristics. Bioresour. Technol. 2016, 206, 23–35. [Google Scholar] [CrossRef]
- Rani, P.; Bansal, M.; Pathak, V.V. Experimental and kinetic studies for improvement of biogas production from KOH pretreated wheat straw. Curr. Res. Green Sustain. Chem. 2022, 5, 100283. [Google Scholar] [CrossRef]
- Maurus, K.; Ahmed, S.; Getz, W.; Kazda, M. Sugar beet silage as highly flexible feedstock for on demand biogas production. Sugar Ind. 2018, 143, 691–698. [Google Scholar] [CrossRef]
- De Moor, S.; Velghe, F.; Wierinck, I.; Michels, E.; Ryckaert, B.; de Vocht, A.; Verbeke, W.; Meers, E. Feasibility of grass co-digestion in an agricultural digester, influence on process parameters and residue composition. Bioresour. Technol. 2013, 150, 187–194. [Google Scholar] [CrossRef]
- Hidaka, T.; Arai, S.; Okamoto, S.; Uchida, T. Anaerobic co-digestion of sewage sludge with shredded grass from public green spaces. Bioresour. Technol. 2013, 130, 667–672. [Google Scholar] [CrossRef]
- Güsewell, J.; Eltrop, L.; Hufendiek, K. Seasonal flexibilisation: A solution for biogas plants to improve profitability. Adv. Appl. Energy 2021, 2, 100034. [Google Scholar] [CrossRef]
- Thrän, D.; Dotzauer, M.; Lenz, V.; Liebetrau, J.; Ortwein, A. Flexible bioenergy supply for balancing fluctuating renewables in the heat and power sector—A review of technologies and concepts. Energy Sustain. Soc. 2015, 5, 35. [Google Scholar] [CrossRef]
- Boob, M.; Elsaesser, M.; Thumm, U.; Hartung, J.; Lewandowski, I. Different management practices influence growth of small plants in species-rich hay meadows through shading. Appl. Veg. Sci. 2021, 24, e12625. [Google Scholar] [CrossRef]
- Boob, M.; Truckses, B.; Seither, M.; Elsäßer, M.; Thumm, U.; Lewandowski, I. Management effects on botanical composition of species-rich meadows within the Natura 2000 network. Biodivers. Conserv. 2019, 28, 729–750. [Google Scholar] [CrossRef]
- Mönch-Tegeder, M.; Lemmer, A.; Oechsner, H. Enhancement of methane production with horse manure supplement and pretreatment in a full-scale biogas process. Energy 2014, 73, 523–530. [Google Scholar] [CrossRef]
- Al Seadi, T.; Rutz, D.; Prassl, H.; Köttner, M.; Finsterwalder, T.; Volk, S.; Janssen, R. Biogas: Handbook. 2008. Available online: https://www.wip-munich.de/pv-policy-group-european-best-practice-report/annex2-39_wp4_d4.1_master-handbook.pdf (accessed on 23 January 2024).



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| XP | XL | XF | aNDF | ADF | ADL | SMY | ||
|---|---|---|---|---|---|---|---|---|
| SMY | Pearson Correlation | 0.724 ** | 0.252 ** | −0.795 ** | −0.795 ** | −0.841 ** | −0.804 ** | 1 |
| Rm | λ | ½ M | SMY | ||
|---|---|---|---|---|---|
| Rm | Pearson Correlation | 1 | |||
| λ | Pearson Correlation | 0.876 ** | 1 | ||
| ½ M | Pearson Correlation | −0.897 ** | −0.766 ** | 1 | |
| SMY | Pearson Correlation | 0.901 ** | 0.693 ** | −0.777 ** | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brandhorst, C.; Hülsemann, B.; Ohnmacht, B.; Lemmer, A. Anaerobic Digestion of Cuttings from Grassland in Protected Landscape Areas. Inventions 2024, 9, 23. https://doi.org/10.3390/inventions9010023
Brandhorst C, Hülsemann B, Ohnmacht B, Lemmer A. Anaerobic Digestion of Cuttings from Grassland in Protected Landscape Areas. Inventions. 2024; 9(1):23. https://doi.org/10.3390/inventions9010023
Chicago/Turabian StyleBrandhorst, Christina, Benedikt Hülsemann, Benjamin Ohnmacht, and Andreas Lemmer. 2024. "Anaerobic Digestion of Cuttings from Grassland in Protected Landscape Areas" Inventions 9, no. 1: 23. https://doi.org/10.3390/inventions9010023
APA StyleBrandhorst, C., Hülsemann, B., Ohnmacht, B., & Lemmer, A. (2024). Anaerobic Digestion of Cuttings from Grassland in Protected Landscape Areas. Inventions, 9(1), 23. https://doi.org/10.3390/inventions9010023