X-Ray Absorption Spectroscopy Measurements of Cu-ProIAPP Complexes at Physiological Concentrations

 ,
,  , , and
, , and
Abstract
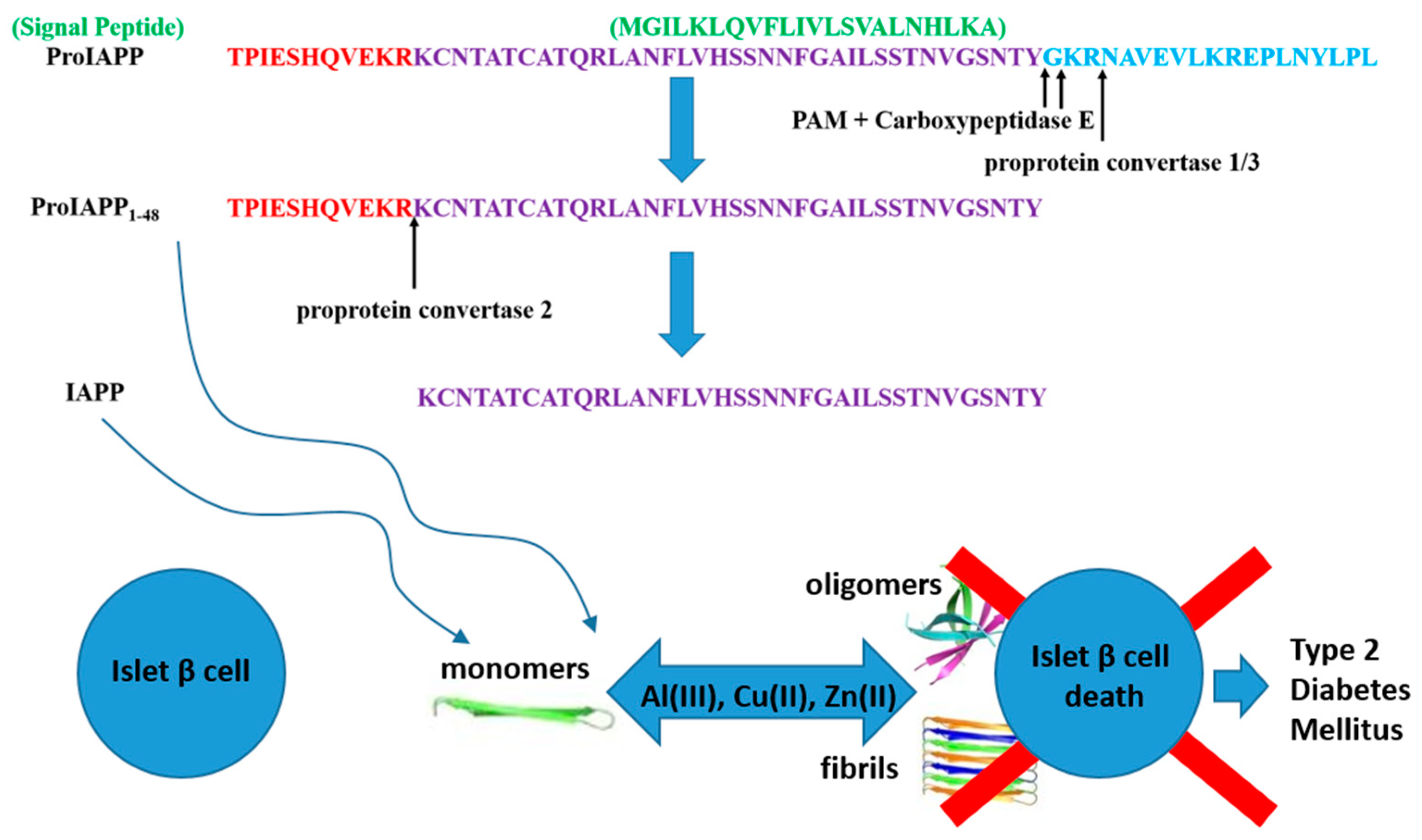
1. Introduction
2. Materials and Methods
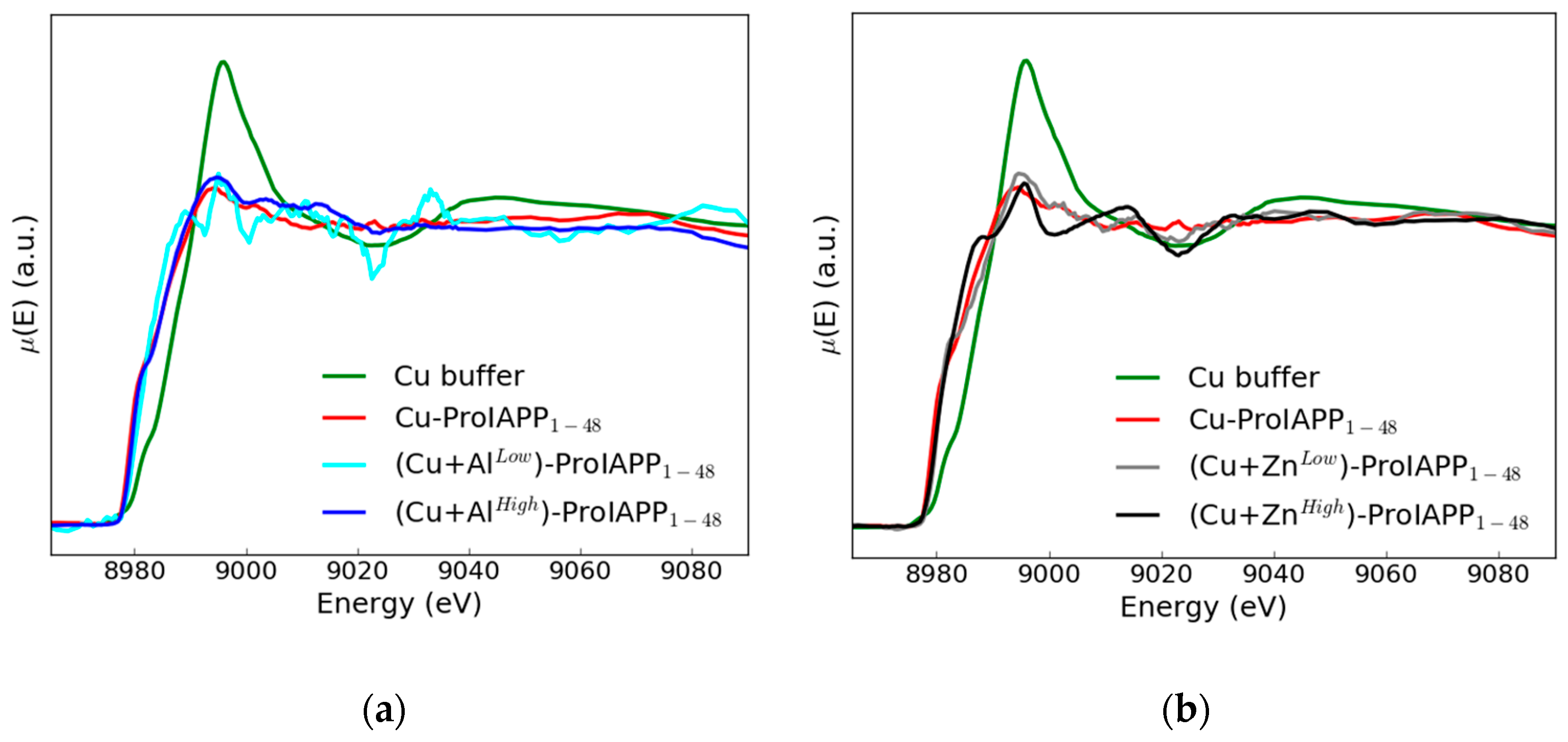
3. Results and Discussion
4. Conclusions
- Though yet at a qualitative level, the existence of differences in the XANES spectral features induced by the presence of the Zn(II) or Al(III) in the Cu(II)-ProIAPP1–48 binding mode sample is clearly established. These differences appear to be dependent (in different way) from the added ion concentration.
- Experiments of the kind we have been able to perform at the ESRF HERFD XAS line demonstrate the general feasibility of XAS measurements on samples where the absorbing atom is present at micromolar concentration. This last fact is of special methodological relevance as it shows that it is possible to perform XAS measurements on very diluted metal-peptide complexes in physiological conditions, when raising metal ions concentration to improve the signal-to-noise ratio is not possible, as this would dramatically alter their physiological coordination mode.
Author Contributions
Funding
Conflicts of Interest
References
- Cooper, G.J.; Willis, A.C.; Clark, A.; Turner, R.C.; Sim, R.B.; Reid, K.B. Purification and characterization of a peptide from amyloid-rich pancreases of type 2 diabetic patients. Proc. Natl. Acad. Sci. USA 1987, 84, 8628–8632. [Google Scholar] [CrossRef] [PubMed]
- Konarkowska, B.; Aitken, J.F.; Kistler, J.; Zhang, S.G.; Cooper, J. The aggregation potential of human amylin determines its cytotoxicity towards islet beta-cells. FEBS J. 2006, 273, 3614–3624. [Google Scholar] [CrossRef] [PubMed]
- Westermark, P.; Engstrom, U.; Westermark, G.T.; Johnson, K.H.; Permerth, J.; Betsholtz, C. Islet amyloid polypeptide (IAPP) and pro-IAPP immunoreactivity in human islets of Langerhans. Diabetes Res. Clin. Pract. 1989, 7, 219–226. [Google Scholar] [CrossRef]
- Courtade, J.A.; Klimek-Abercrombie, A.M.; Chen, Y.-C.; Patel, N.; Lu, P.Y.T.; Speake, C.; Orban, P.C.; Najafian, B.; Meneilly, G.; Greenbaum, C.J.; et al. Measurement of pro-islet amyloid polypeptide (1–48) in diabetes and islet transplants. J. Clin. Endocrinol. Metab. 2017, 102, 2595–2603. [Google Scholar] [CrossRef] [PubMed]
- Raleigh, D.; Zhang, X.; Hastoy, B.; Clark, A. The β-cell assassin: IAPP cytotoxicity. J. Mol. Endocrinol. 2017, 59, R121–R140. [Google Scholar] [CrossRef] [PubMed]
- Exley, C.; Korchazhkina, O. Promotion of formation of amyloid fibrils by aluminium adenosine triphosphate (AlATP). J. Inorg. Biochem. 2001, 84, 215–224. [Google Scholar] [CrossRef]
- Ward, B.; Walker, K.; Exley, C. Cu(II) inhibits the formation of amylin amyloid in vitro. J. Inorg. Biochem. 2008, 102, 371–375. [Google Scholar] [CrossRef]
- Brender, J.R.; Hartman, K.; Nanga, R.P.R.; Popovych, N.; Bea, R.D.; Vivekanandan, S.; Marsh, E.G.N.; Ramamoorthy, A. Role of zinc in human islet amyloid polypeptide aggregation. J. Am. Chem. Soc. 2010, 132, 8973–8983. [Google Scholar] [CrossRef]
- Ma, L.; Li, X.; Wang, Y.; Zheng, W.; Chen, T. Cu(II) inhibits hIAPP fibrillation and promotes hIAPP-induced beta cell apoptosis through induction of ROS-mediated mitochondrial dysfunction. J. Inorg. Biochem. 2014, 140, 143–152. [Google Scholar] [CrossRef]
- Li, H.; Ha, E.; Donaldson, R.P.; Jeremic, A.M.; Vertes, A. Rapid assessment of human amylin aggregation and its inhibition by copper (II) ions by laser ablation electrospray ionisation mass spectrometry with ion mobility separation. Anal. Chem. 2015, 87, 9829–9837. [Google Scholar] [CrossRef]
- Riba, I.; Barran, P.E.; Cooper, G.J.S.; Unwin, R.D. On the structure of the copper-amylin complex. Int. J. Mass Spectrom. 2015, 391, 47–53. [Google Scholar] [CrossRef]
- Mold, M.; Bunrat, C.; Goswami, P.; Roberts, A.; Roberts, C.; Taylor, N.; Taylor, H.; Wu, L.; Fraser, P.E.; Exley, C. Further insight into the role of metals in amyloid formation by IAPP1–37 and ProIAPP1–48. J. Diabetes Res. Clin. Metab. 2015, 4. [Google Scholar] [CrossRef]
- Sánchez-López, C.; Cortés-Mejía, R.; Miotto, M.C.; Binolfi, A.; Fernández, C.O.; del Campo, J.M.; Quintanar, L. Copper coordination features of human islet amyloid polypeptide: The type 2 diabetes peptide. Inorg. Chem. 2016, 55, 10727–10740. [Google Scholar] [CrossRef]
- Exley, C.; House, E.; Patel, T.; Wu, L.; Fraser, P.E. Human pro-islet amyloid polypeptide (ProIAPP1–48) forms amyloid fibrils and amyloid spherulites in vitro. J. Inorg. Biochem. 2010, 104, 1125–1129. [Google Scholar] [CrossRef] [PubMed]
- Exley, C.; Mold, M.; Shardlow, E.; Shuker, B.; Ikpe, B.; Wu, L.; Fraser, P.E. Copper is a potent inhibitor of the propensity for human ProIAPP1–48 to form amyloid fibrils in vitro. J. Diabetes Res. Clin. Metab. 2012, 1, 3. [Google Scholar] [CrossRef]
- Lee, S.J.C.; Choi, T.S.; Lee, J.W.; Lee, H.J.; Mun, D.-G.; Akashi, S.; Lee, S.-W.; Lim, M.H.; Kim, H.I. Structure and assembly mechanisms of toxic human islet amyloid polypeptide oligomers associated with copper. Chem. Sci. 2016, 7, 5398–5406. [Google Scholar] [CrossRef] [PubMed]
- De Santis, E.; Minicozzi, V.; Proux, O.; Rossi, G.C.; Silva, K.I.; Lawless, M.J.; Stellato, F.; Saxena, S.; Morante, S. Cu(II)–Zn(II) Cross-Modulation in Amyloid–Beta Peptide Binding: An X-ray Absorption Spectroscopy Study. J. Phys. Chem. B 2015, 119, 15813–15820. [Google Scholar] [CrossRef]
- Stellato, F.; Spevacek, A.; Proux, O.; Minicozzi, V.; Millhauser, G.; Morante, S. Zinc modulates copper coordination mode in prion protein octa-repeat subdomains. Eur. Biophys. J. 2011, 40, 1259–1270. [Google Scholar] [CrossRef]
- Stellato, F.; Minicozzi, V.; Millhauser, G.L.; Pascucci, M.; Proux, O.; Rossi, G.C.; Morante, S. Copper–zinc cross-modulation in prion protein binding. Eur. Biophys. J. 2014, 43, 631–642. [Google Scholar] [CrossRef]
- Stellato, F.; Fusco, Z.; Chiaraluce, R.; Consalvi, V.; Dinarelli, S.; Placidi, E.; Petrosino, M.; Rossi, G.C.R.; Minicozzi, V.; Morante, S. The effect of β-sheet breaker peptides on metal associated Amyloid-β peptide aggregation process. Biophys. Chem. 2017, 229, 110–114. [Google Scholar] [CrossRef]
- Shardlow, E.; Rao, C.; Sattarov, R.; Wu, L.; Fraser, P.E.; Exley, C. Aggregation of the diabetes-related peptide ProIAPP1–48 measured by dynamic light scattering. J. Trace Elem. Med. Biol. 2019, 51, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Proux, O.; Lahera, E.; Del Net, W.; Kieffer, I.; Rovezzi, M.; Testemale, D.; Irar, M.; Thomas, S.; Aguilar-Tapia, A.; Bazarkina, E.F.; et al. High-energy resolution fluorescence detected X-ray absorption spectroscopy: A powerful new structural tool in environmental biogeochemistry sciences. J. Environ. Qual. 2017, 46, 1146–1157. [Google Scholar] [CrossRef] [PubMed]
- Castillo, R.G.; Banerjee, R.; Allpress, C.J.; Rohde, G.T.; Bill, E.; Que, L., Jr.; Lipscomb, J.D.; DeBeer, S. High-energy-resolution fluorescence-detected X-ray absorption of the Q intermediate of soluble methane monooxygenase. JACS 2017, 139, 18024–18033. [Google Scholar] [CrossRef] [PubMed]
- Bjornsson, R.; Lima, F.A.; Spatzal, T.; Weyhermüller, T.; Glatzel, P.; Bill, E.; Einzle, O.; Neese, F.; DeBeer, S. Identification of a spin-coupled Mo(III) in the nitrogenase iron–molybdenum cofactor. Chem. Sci. 2014, 5, 3096–3103. [Google Scholar] [CrossRef]
- Mebs, S.; Kositzki, R.; Duan, J.; Kertess, L.; Senger, M.; Wittkamp, F.; Apfel, U.P.; Happe, T.; Stripp, S.T.; Winkler, M.; et al. Hydrogen and oxygen trapping at the H-cluster of [FeFe]-hydrogenase revealed by site-selective spectroscopy and QM/MM calculations. Biochim. Biophys. Acta Bioenerg. 2018, 1859, 28–41. [Google Scholar] [CrossRef] [PubMed]
- Vollmers, N.J.; Müller, P.; Hoffmann, A.; Herres-Pawlis, S.; Rohrmuüller, M.; Schmidt, W.G.; Gerstmann, U.; Bauer, M. Experimental and theoretical high-energy-resolution X-ray absorption spectroscopy: Implications for the investigation of the entatic state. Inorg. Chem. 2016, 55, 11694–11706. [Google Scholar] [CrossRef] [PubMed]
- Kowalska, J.K.; Lima, F.A.; Pollock, C.J.; Rees, J.A.; DeBeer, S. A practical guide to high-resolution X-ray spectroscopic measurements and their applications in bioinorganic chemistry. Isr. J. Chem. 2016, 56, 803–815. [Google Scholar] [CrossRef]
- La Penna, G.; Minicozzi, V.; Morante, S.; Rossi, G.C.; Stellato, F. A first-principle calculation of the XANES spectrum of Cu2+ in water. J. Chem. Phys. 2015, 143, 124508. [Google Scholar] [CrossRef] [PubMed]
- Stellato, F.; Calandra, M.; D’Acapito, F.; De Santis, E.; La Penna, G.; Rossi, G.C.; Morante, S. Multi-scale theoretical approach to X-ray absorption spectra in disordered systems: An application to the study of Zn(II) in water. Phys. Chem. Chem. Phys. 2018, 20, 24775–24782. [Google Scholar] [CrossRef] [PubMed]
- Banci, L.; Bertini, I.; Mangani, S. Integration of XAS and NMR techniques for the structure determination of metalloproteins. Examples from the study of copper transport proteins. J. Synchrotron Radiat. 2005, 12, 94–97. [Google Scholar] [CrossRef]
- Koutmou, K.S.; Casiano-Negroni, A.; Getz, M.M.; Pazicni, S.; Andrews, A.J.; Penner-Hahn, J.E.; Al-Hashimi, H.M.; Fierke, C.A. NMR and XAS reveal an inner-sphere metal binding site in the P4 helix of the metallo-ribozyme ribonuclease P. Proc. Natl. Acad. Sci. USA 2010, 107, 2479–2484. [Google Scholar] [CrossRef] [PubMed]
- Arcovito, A.; Moschetti, T.; D’Angelo, P.; Mancini, G.; Vallone, B.; Brunori, M.; Della Longa, S. An X-ray diffraction and X-ray absorption spectroscopy joint study of neuroglobin. Arch. Biochem. Biophys. 2008, 475, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Frankær, C.G.; Knudsen, M.V.; Norén, K.; Nazarenko, E.; Ståhl, K.; Harris, P. The structures of T6, T3R3 and R6 bovine insulin: Combining X-ray diffraction and absorption spectroscopy. Acta Crystallogr. D 2012, 68, 1259–1271. [Google Scholar] [CrossRef] [PubMed]
- Koebke, K.J.; Ruckthong, L.; Meagher, J.L.; Mathieu, E.; Harland, J.; Deb, A.; Lehnert, N.; Policar, C.; Tard, C.; Penner-Han, J.E.; et al. Clarifying the Copper Coordination Environment in a de Novo Designed Red Copper Protein. Inorg. Chem. 2018, 57, 12291–12302. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Sample(µm) | [Peptide] (µm) | [Cu] (µm) | [Zn] (µm) | [Al] (µm) |
|---|---|---|---|---|
| Cu buffer | 0 | 1600 | 0 | 0 |
| Cu-ProIAPP1–48 | 10 | 9 | 0 | 0 |
| (Cu + ZnLow)-ProIAPP1–48 | 10 | 9 | 50 | 0 |
| (Cu + AlLow)-ProIAPP1–48 | 10 | 9 | 0 | 50 |
| (Cu + ZnHigh)-ProIAPP1–48 | 10 | 9 | 1500 | 0 |
| (Cu + AlHigh)-ProIAPP1–48 | 10 | 9 | 0 | 1500 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Santis, E.; Shardlow, E.; Stellato, F.; Proux, O.; Rossi, G.; Exley, C.; Morante, S. X-Ray Absorption Spectroscopy Measurements of Cu-ProIAPP Complexes at Physiological Concentrations. Condens. Matter 2019, 4, 13. https://doi.org/10.3390/condmat4010013
De Santis E, Shardlow E, Stellato F, Proux O, Rossi G, Exley C, Morante S. X-Ray Absorption Spectroscopy Measurements of Cu-ProIAPP Complexes at Physiological Concentrations. Condensed Matter. 2019; 4(1):13. https://doi.org/10.3390/condmat4010013
Chicago/Turabian StyleDe Santis, Emiliano, Emma Shardlow, Francesco Stellato, Olivier Proux, Giancarlo Rossi, Christopher Exley, and Silvia Morante. 2019. "X-Ray Absorption Spectroscopy Measurements of Cu-ProIAPP Complexes at Physiological Concentrations" Condensed Matter 4, no. 1: 13. https://doi.org/10.3390/condmat4010013
APA StyleDe Santis, E., Shardlow, E., Stellato, F., Proux, O., Rossi, G., Exley, C., & Morante, S. (2019). X-Ray Absorption Spectroscopy Measurements of Cu-ProIAPP Complexes at Physiological Concentrations. Condensed Matter, 4(1), 13. https://doi.org/10.3390/condmat4010013