
Morphometric Analysis of Two Shell Color Strains of the Bay Scallop Argopecten irradians

Abstract
1. Introduction
2. Materials and Methods
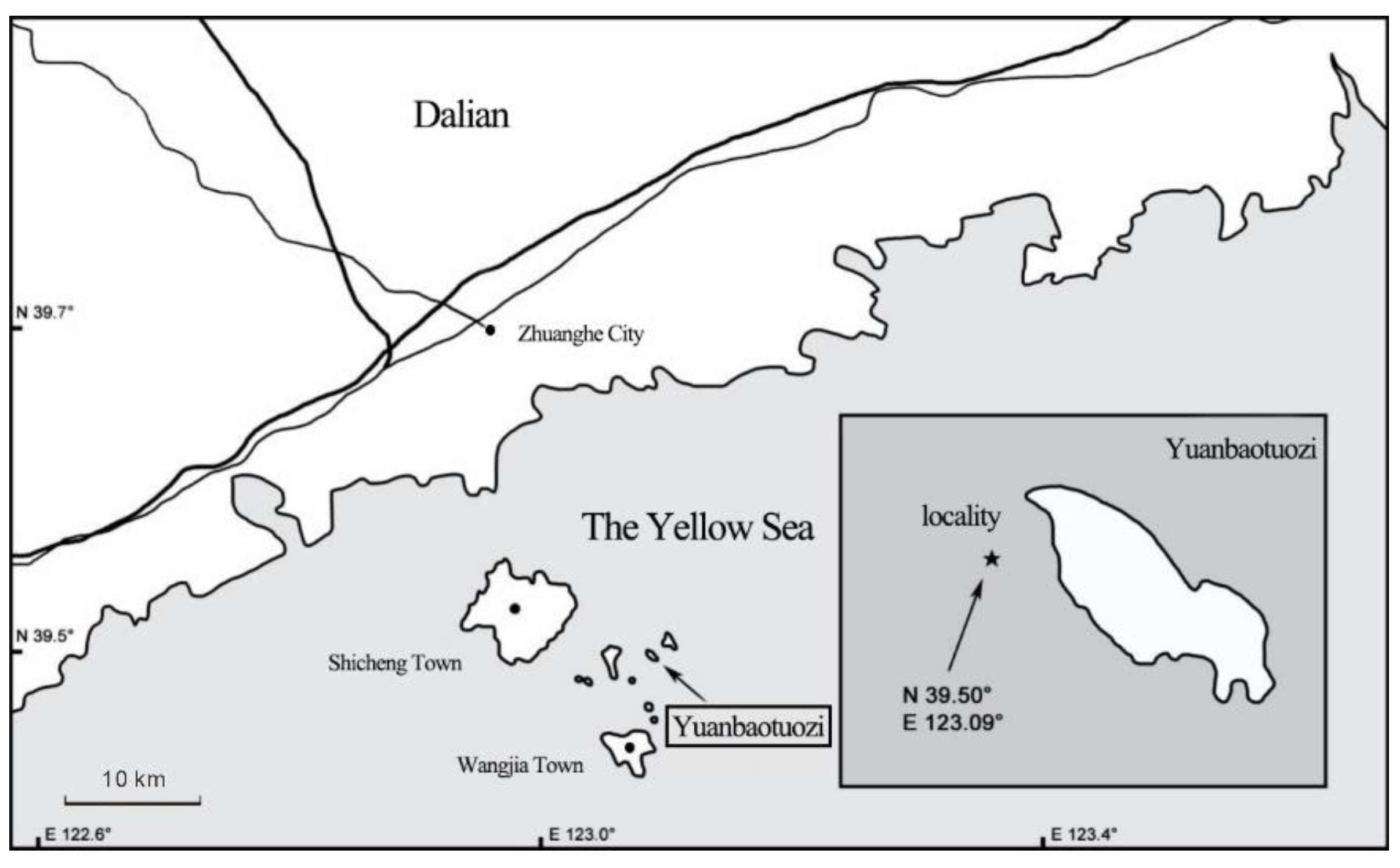
2.1. Source and Material Treatment
2.2. Methods
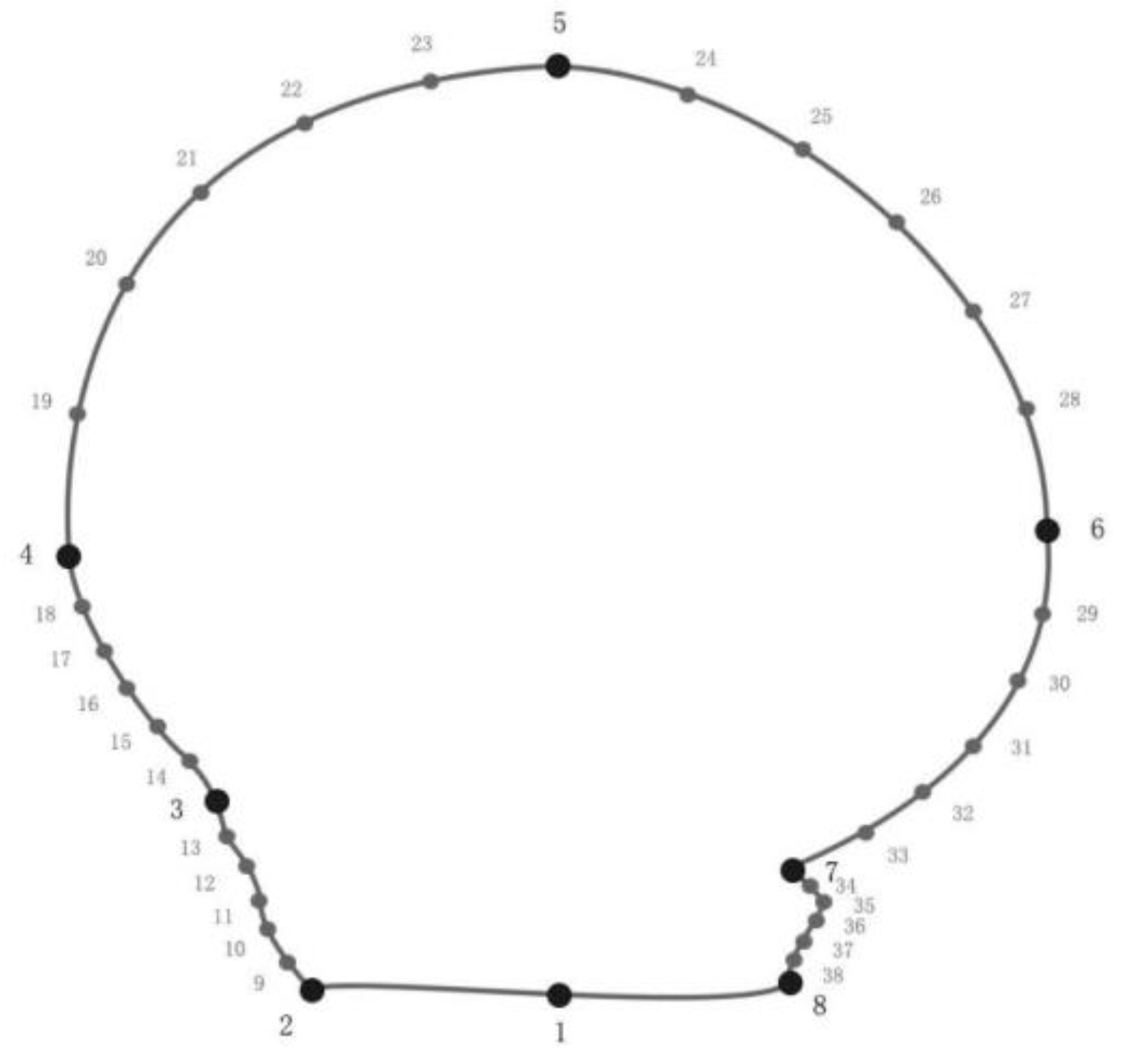
2.2.1. Image Acquisition
2.2.2. Standardized Processing of Data
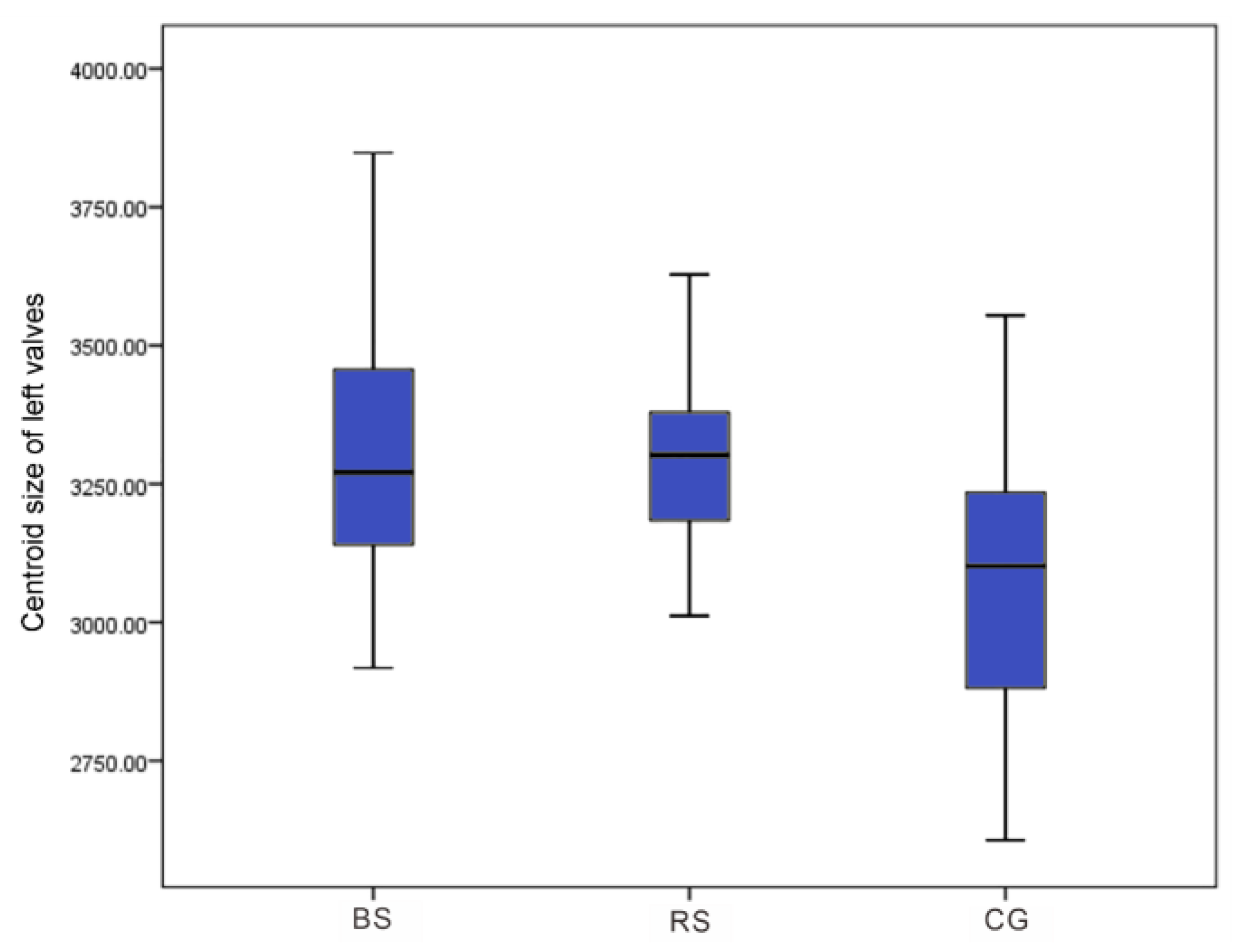
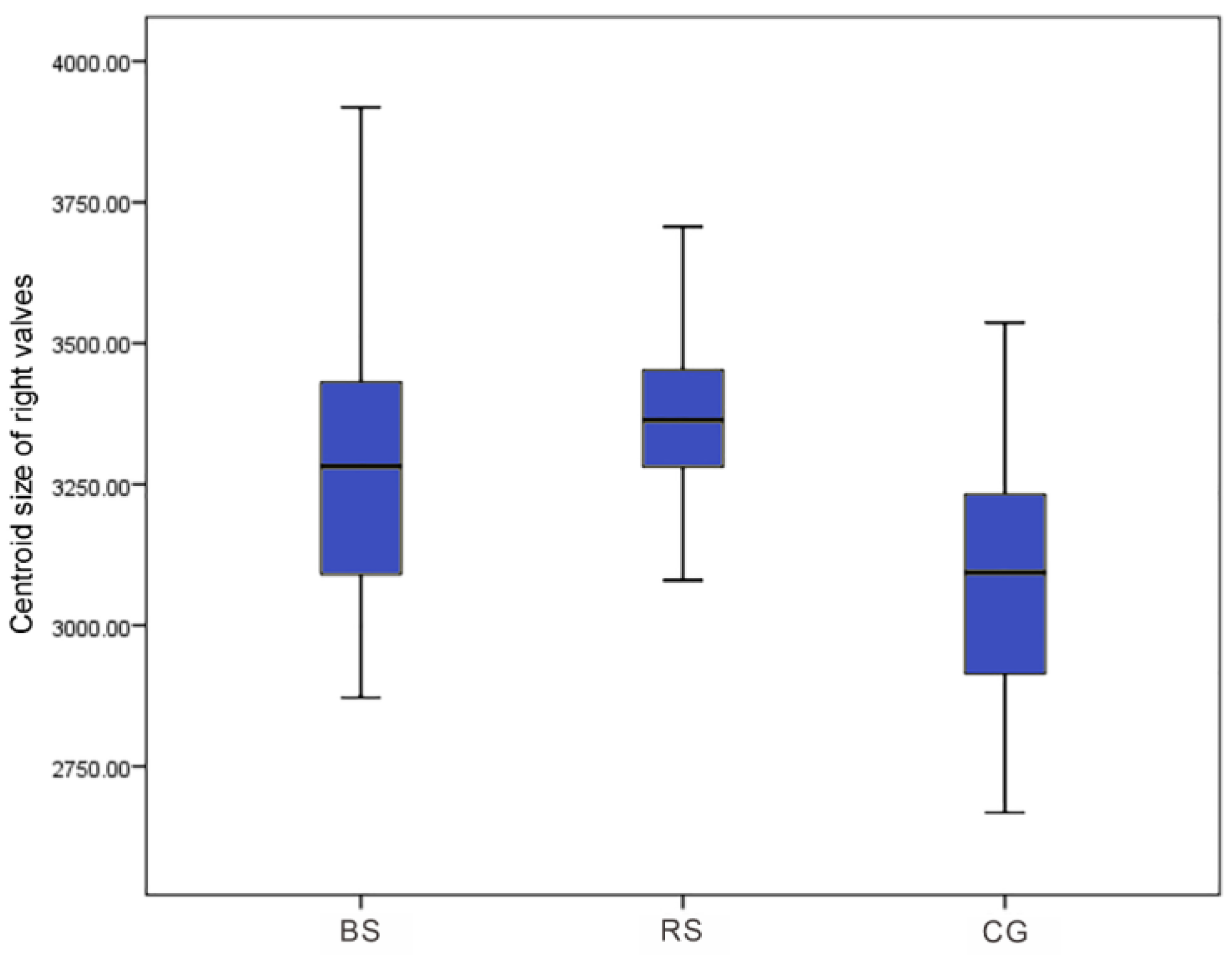
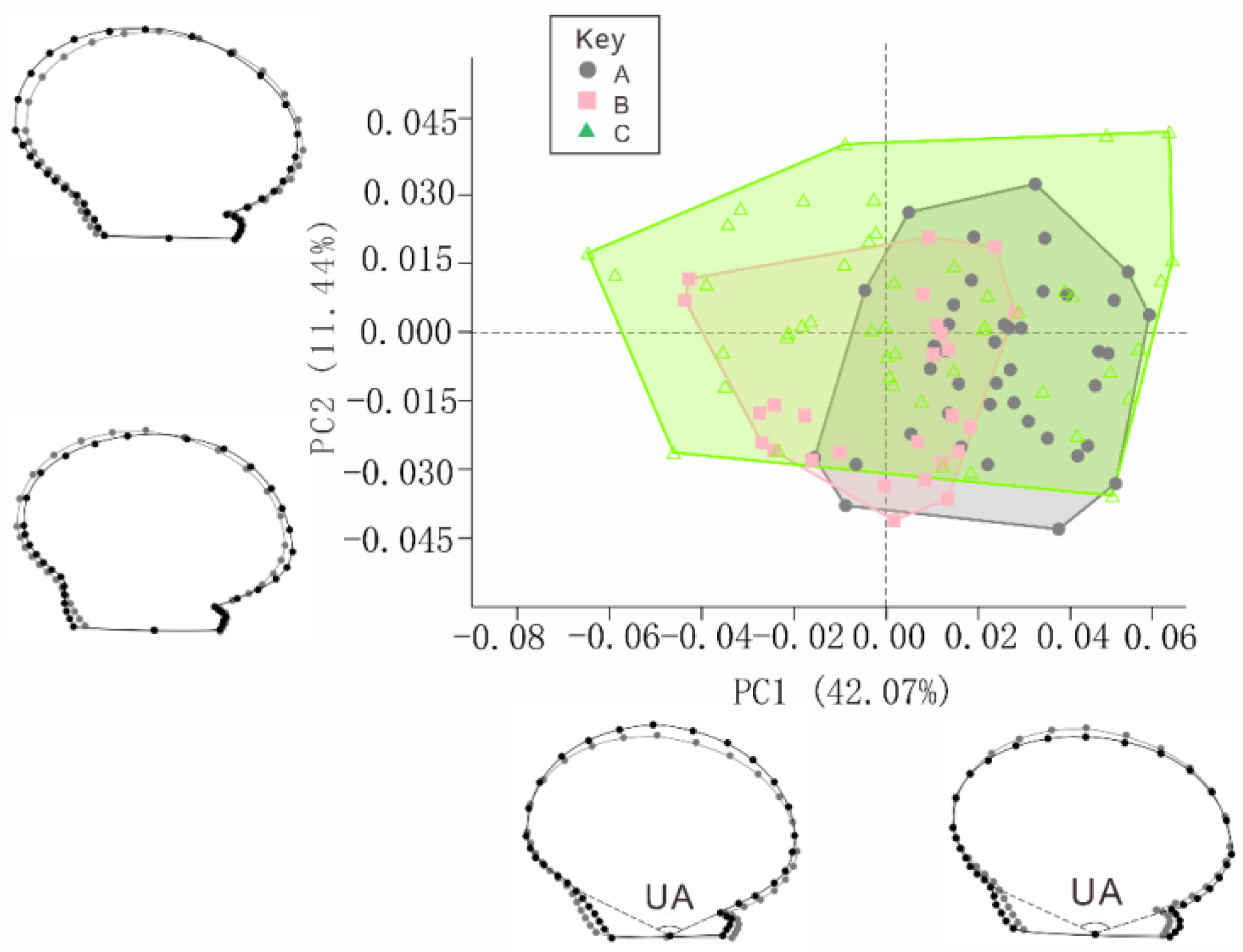
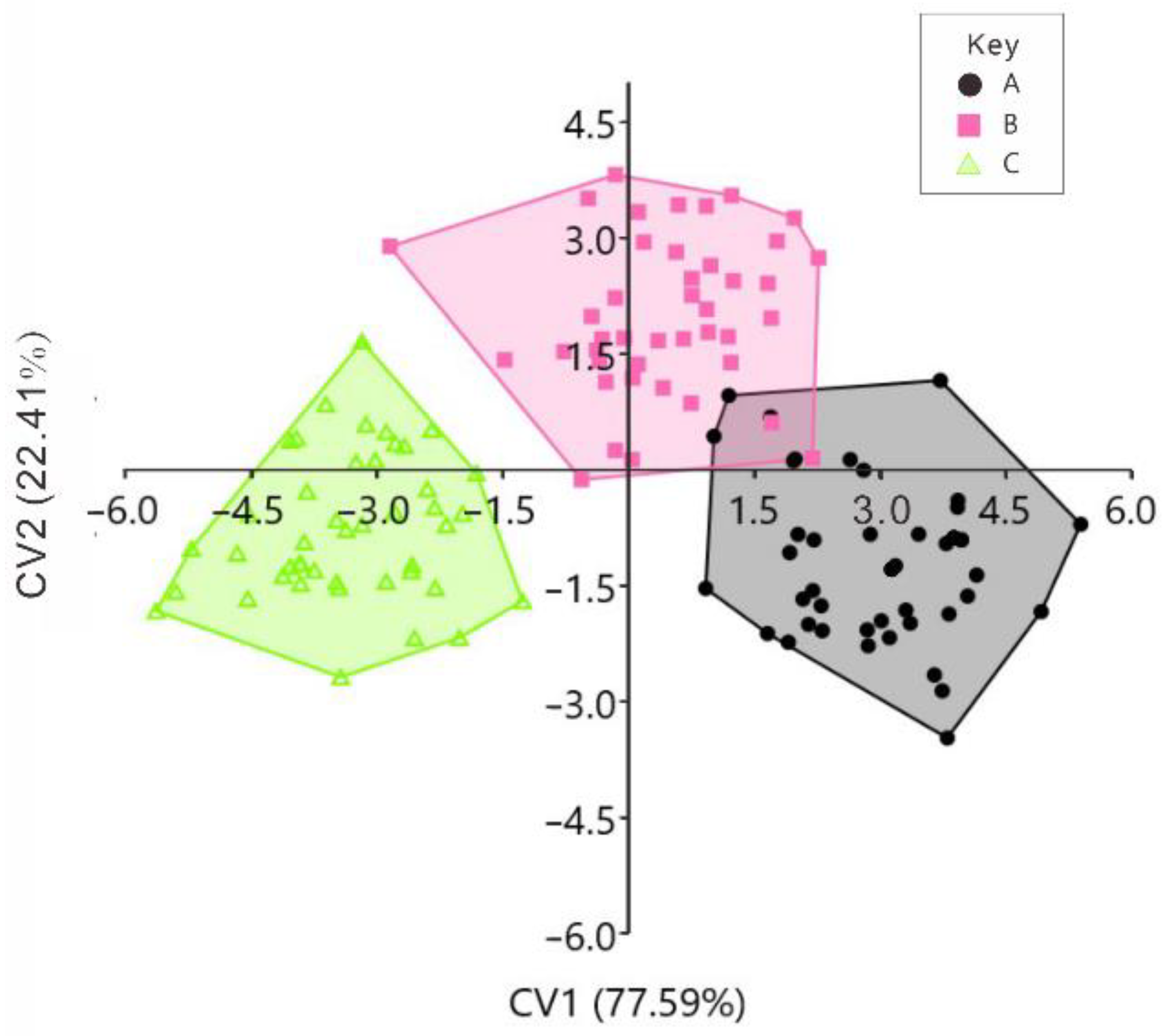
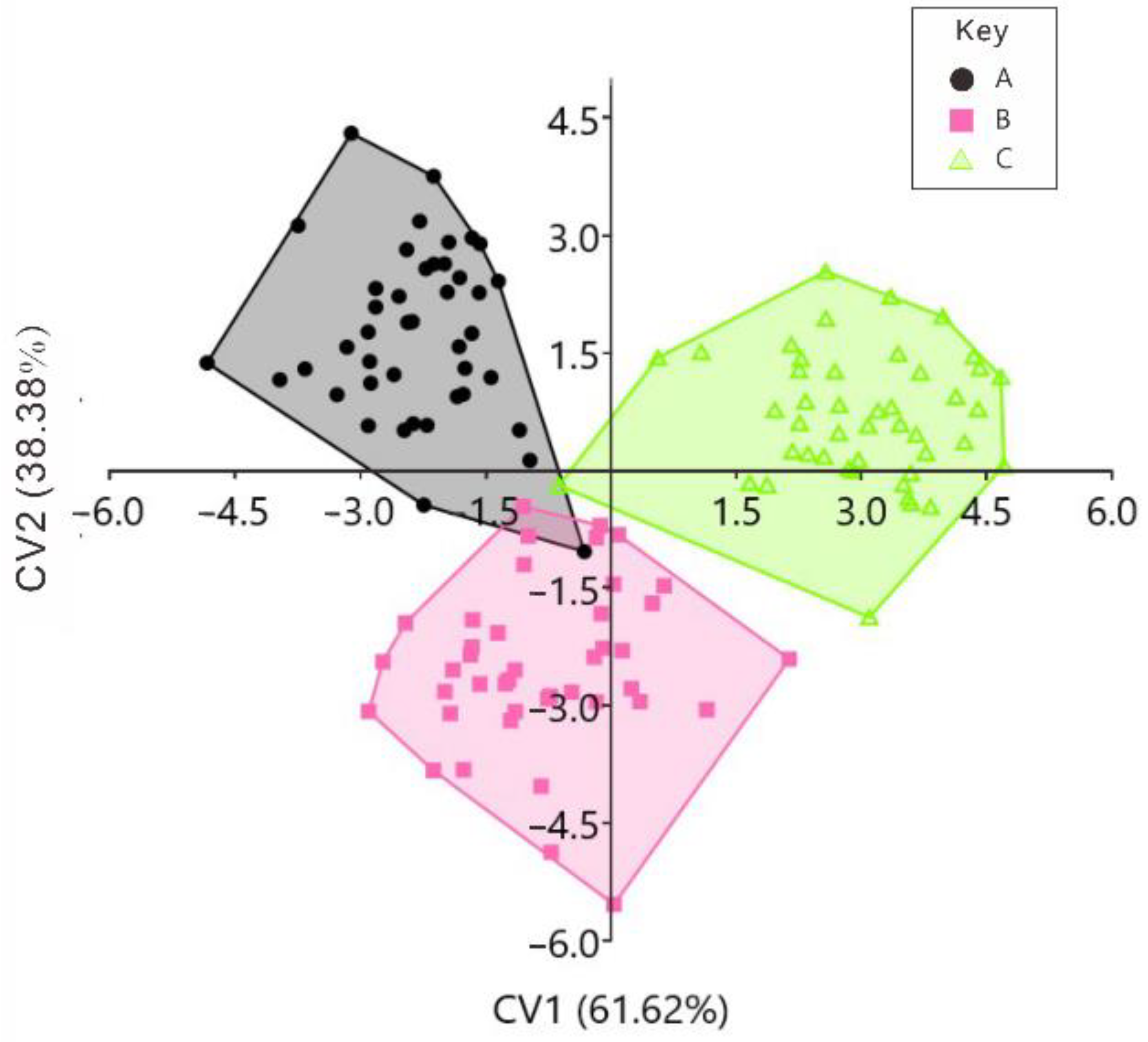
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gaspar, M.B.; Santos, M.N.; Vasconcelos, P.; Monteiro, C.C. Shell morphometric relationships of the most common bivalve species (Mollusca: Bivalvia) of the Algarve coast (southern Portugal). Hydrobiologia 2002, 477, 73–80. [Google Scholar] [CrossRef]
- Manuel, J.L.; Dadswell, M.J. Swimming of juvenile sea scallops, Placopecten magellanicus (Gmelin): A minimum size for effective swimming? J. Exp. Mar. Biol. Ecol. 1993, 174, 137–175. [Google Scholar] [CrossRef]
- Márquez, F.; Robledo, J.; Peñalosa, G.E.; Molen, S.V.D. Use of different geometric morphometrics tools for the discrimination of phenotypic stocks of the striped clam Ameghinomya antiqua (Veneridae) in north Patagonia, Argentina. Fish. Res. 2010, 101, 127–131. [Google Scholar] [CrossRef]
- Márquez, F.; González-José, R.; Bigatti, G. Combined methods to detect pollution effects on shell shape and structure in Neogastropods. Ecol. Indic. 2011, 11, 248–254. [Google Scholar] [CrossRef]
- Serb, J.M.; Alejandrino, A.; Otárola-castillo, E.; Adams, D.C. Morphological convergence of shell shape in distantly related scallop species (Mollusca: Pectinidae). Zool. J. Linn. Soc. 2011, 163, 571–584. [Google Scholar] [CrossRef]
- Stanley, S.M. Relation of shell form to life habitats in the Bivalvia (Mollusca). Mem. Geol. Soc. Am. 1970, 125, 1–296. [Google Scholar] [CrossRef]
- Ubukata, T. A morphometric study on morphological plasticity of shell form in crevice-dwelling Pterioida (Bivalvia). Biol. J. Linn. Soc. Lond. 2003, 79, 285–297. [Google Scholar] [CrossRef]
- Clarke, B. Natural selection in mixed populations of two polymorphic snails. Heredity 1962, 17, 319–345. [Google Scholar] [CrossRef]
- Murray, J.; Clarke, B. The inheritance of polymorphic shell characters in partula (Gastropoda). Genetics 1966, 54, 1261–1277. [Google Scholar] [CrossRef]
- Cook, L.M.; Freeman, P.M. Heating properties of morphs of the mangrove snail Littoraria pallescens. Biol. J. Linn. Soc. 1986, 29, 295–300. [Google Scholar] [CrossRef]
- Rosin, Z.M.; Kwieciński, Z.; Lesicki, A.; Skórka, P.; Kobak, J.; Szymańska, A.; Osiejuk, T.S.; Kałuski, T.; Jaskulska, M.; Tryjanowski, P. Shell colour, temperature, (micro)habitat structure and predator pressure affect the behaviour of Cepaea nemoralis. Sci. Nat. 2018, 105, 35. [Google Scholar] [CrossRef]
- Savazzi, E. The colour patterns of cypraeid gastropods. Lethaia 2007, 31, 15–27. [Google Scholar] [CrossRef]
- Scheil, A.E.; Hilsmann, S.; Triebskorn, R.; Köhler, H.R. Shell colour polymorphism, injuries and immune defense in three helicid snail species, Cepaea hortensis, Theba pisana and Cornu aspersum maximum. Results Immunol. 2013, 3, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Ekendahl, A. Colour polymorphic prey (Littorina saxatilis Olivi) and predatory effects of a crab population (Carcinus maenas L.). J. Exp. Mar. Biol. Ecol. 1998, 222, 239–246. [Google Scholar] [CrossRef]
- Raffaelli, D. Colour polymorphism in the intertidal snail Littorina rudis Maton. Zool. J. Linn. Soc. 1979, 67, 65–73. [Google Scholar] [CrossRef]
- Winkler, F.M.; Estévez, B.F.; Jollán, L.B.; Garrido, J.P. Inheritance of the general shell color in the scallop Argopecten purpuratus (Bivalvia: Pectinidae). J. Hered. 2001, 92, 521–525. [Google Scholar] [CrossRef] [PubMed]
- Strelkov, P.P.; Gantsevich, M.M.; Basova, L.A. Shell color polymorphism in Macoma balthica L. (Bivalvia, Tellinidae) from the White and Barents Seas. Dokl. Biol. Sci. 2001, 376, 59–61. [Google Scholar] [CrossRef]
- Mitton, J.B. Shell color and pattern variation in Mytilus edulis and its adaptive significance. Chesap. Sci. 1977, 18, 387–390. [Google Scholar] [CrossRef]
- Newkirk, G.F. Genetics of shell color in Mytilus edulis L. and the association of growth rate with shell color. J. Exp. Mar. Biol. Ecol. 1980, 47, 89–94. [Google Scholar] [CrossRef]
- Innes, D.J.; Leslie, E.H. Inheritance of a shell-color polymorphism in the mussel. J. Hered. 1977, 68, 203–204. [Google Scholar] [CrossRef]
- Evans, S.; Camara, M.D.; Langdon, C.G. Heritability of shell pigmentation in the Pacific oyster, Crassostrea gigas. Aquaculture 2009, 286, 211–216. [Google Scholar] [CrossRef]
- Adamkewicz, L.; Castagna, M. Genetics of the shell color and pattern in the bay scallop Argopecten irradians. J. Hered. 1988, 79, 14–17. [Google Scholar] [CrossRef]
- Bai, M.; Yang, X.K.; Li, J.; Wang, W.C. Geometric morphometrics, a super scientific computing tool in morphology comparison. Chin. Sci. Bull. 2014, 59, 887–894. [Google Scholar] [CrossRef]
- Lai, S.Q.; Xie, Y.Y.; Huang, X.P.; Mao, J.X.; Wang, X.B.; Tian, Y.; Chang, Y.Q. Ontogenetic changes during the life span of the scallop Patinopecten yessoensis determined using a geometric morphometric method. J. Shellfish Res. 2023, 42, 259–264. [Google Scholar] [CrossRef]
- Márquez, F.; Amoroso, R.; Sainz, G.M.F.; Molen, S.V.D. Shell morphology changes in the scallop Aequipecten tehuelchus during its life span: A geometric morphometric approach. Aquat. Biol. 2010, 11, 149–155. [Google Scholar] [CrossRef]
- Cain, A.J. The colours of marine bivalve shells with special reference to Macoma balthica. Malacologia 1988, 28, 289–318. [Google Scholar]
- Yonge, C.M. On the habits and adaptations of Aloidis (corbula) gibba. J. Mar. Biol. Associ. UK 1946, 26, 358–376. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.A.S. Polymorphism and selective predation in Donax faba Gmelin (Bivalvia: Tellinacea). J. Exp. Mar. Biol. Ecol. 1975, 17, 89–94. [Google Scholar] [CrossRef]
- Beukema, J.J.; Meehan, B.W. Latitudinal variation in linear growth and other shell characteristics of Macoma balthica. Mar. Bio. 1985, 90, 27–33. [Google Scholar] [CrossRef]
- Wolff, M.; Garrido, J. Comparative study of growth and survival of two color morphs of the Chilean scallop Argopecten purpuratus (Lamarck) in suspended culture. J. Shellfish Res. 1991, 10, 47–53. [Google Scholar]
- Alfonsi, C.; Perez, J.E. Growth and survival in the scallop Nodipecten nodosus as related to self-fertilization and shell colour. Bol. Inst. Oceanogr. Venez. 1998, 37, 69–73. [Google Scholar]
- Zheng, H.P.; Zhang, G.F.; Liu, X.; Que, H.Y. Establishment of different shell color lines of bay scallop Argopecten irradians irradians Lamarck (1819) and their development. Oceanol. Limnol. Sin. 2003, 34, 632–639. [Google Scholar]
- Hou, X.M.; Zhang, F.C.; Mu, Y.T. Characteristics, problems and development strategies of the bay scallop industry in Hebei. Chin. Fish. Econ. 2017, 35, 80–88. (In Chinese) [Google Scholar]
- Xiao, J.; Ford, S.E.; Yang, H.S.; Zhang, G.F.; Zhang, F.S.; Guo, X.M. Studies on mass summer mortality of cultured zhikong scallops (Chlamys farreri Jones et Preston) in China. Aquaculture 2005, 250, 602–615. [Google Scholar] [CrossRef]
- Milke, L.M.; Bricelj, V.M.; Parrish, C.C. Comparison of early life history stages of the bay scallop, Argopecten irradians: Effects of microalgal diets on growth and biochemical composition. Aquaculture 2006, 260, 272–289. [Google Scholar] [CrossRef]
- Wilbur, A.E.; Gaffney, P.M. A genetic basis for geographic variation in shell morphology in the bay scallop, Argopecten irradians. Mar. Bio. 1997, 128, 97–105. [Google Scholar] [CrossRef]
- Bao, Z.M.; Huang, X.T.; Xing, Q.; Wang, Y.T. The bay scallop Argopecten irradians “Haiyifeng 12”. Ch. Fish. 2017, 44, 70–73. (In Chinese) [Google Scholar]
- Wang, C.D.; Liu, B.; Ma, B.; Zhao, Y.M.; Zhao, X. The scallop “Bohai red”. Ch. Fish. 2016, 43, 72–77. (In Chinese) [Google Scholar]
- Kraeuter, J.; Adamkewicz, L.; Castagna, M.; Wall, R.; Karney, R. Rib number and shell color in hybridized subspecies of the Atlantic bay scallop, Argopecten irradians. Nautilus 1984, 98, 17–20. [Google Scholar]
- Elek, J.A.; Adamkewicz, S.L. Polymorphism for shell color in the Atlantic bay scallop Argopecten irradians irradians (Lamarck) (Mollusca; bivalvia) on Martha’s Vineyard Island. Am. Malac. Bull. 1990, 7, 117–126. [Google Scholar]
- Zheng, H.P.; Xu, F.; Zhang, G.F.; Liu, X.; Wang, L.S. Relationships between shell colors and quantitative traits in the bay scallop, Argopecten irradians irradians (Lamarck, 1819). Oceanol. Limnol. Sin. 2008, 39, 328–333. (In Chinese) [Google Scholar]
- Wang, X.B.; Ding, S.Q.; Yin, D.H.; Song, J.; Chang, Y.Q. Response to selection for growth in the second generation of two shell color lines of the bay scallop Argopecten irradians. Aquaculture 2020, 528, 735536. [Google Scholar] [CrossRef]
- Shu, Y.; Shi, L.; Lai, S.Q.; Tian, Y.; Chang, Y.Q. Morphometric study and comparison of scallops (Chlamys farreri) cultured in different water depth. Acta. Hydrobiol. Sin. 2021, 45, 132–139. (In Chinese) [Google Scholar]
- Bookstein, F.L. Distance measures. In Morphometric Tools for Landmark Data: Geometry and Biology; Cambridge University Press: Cambridge, UK, 1992; pp. 88–124. [Google Scholar]
- Rohlf, F.J.; Marcus, L.F. A revolution morphometrics. Trends Ecol. Evol. 1993, 8, 129–132. [Google Scholar] [CrossRef] [PubMed]
- Hammer, O.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Xu, X.Y.; Hu, L.P.; He, J.B.; Jiang, L.M.; Yang, T.; Shi, W.K.; Zhang, Y.H.; Ll, W. Effects of morphometric traits of Argopecten irradians on its quality traits. Trans. Oceanol. Limnol. 2023, 45, 73–78. (In Chinese) [Google Scholar]
- Ibarra, A.M. Correlated responses at age 5 months and 1 year for a number of growth traits to selection for total weight and shell width in catarina scallop (Argopecten ventricosus). Aquaculture 1999, 175, 243–254. [Google Scholar] [CrossRef]
- Zheng, H.P.; Sun, Z.W.; Zhang, T.; Liu, H.; Li, Y.Y. Correlation and path analysis to quantitative traits of noble scallop Chlamys nobilis Reeve at one-year old. Chin. Agric. Sci. Bull. 2009, 25, 322–326. (In Chinese) [Google Scholar]
- Chang, Y.Q.; Zhang, C.S.; Cao, X.B.; Yang, X.G.; Li, Y.F. Effect of morphometrical traits on weight traits in one-year old yesso scallop Patinopecten yessoensis. J. Dalian Ocean. Univ. 2008, 23, 330–334. (In Chinese) [Google Scholar]
- Watanabe, S.; Katayama, S. Relationships among shell shape, shell growth rate, and nutritional condition in the Manila clam (Ruditapes philippinarum) in Japan. J. Shellfish. Res. 2010, 29, 353–359. [Google Scholar] [CrossRef]
- Alunno-Bruscia, M.; Bourget, E.; Fréchette, M. Shell allometry and length-mass-density relationship for Mytilus edulis in an experimental food-regulated situation. Mar. Ecol. Prog. Ser. 2001, 219, 177–188. [Google Scholar] [CrossRef]
- Zhang, A.G.; Wang, L.L.; Yang, X.L.; Hu, X.C.; Fu, Y.B.; Li, C.H.; Chen, A.H.; Yuan, X.T. Relationship between shell morphological traits and body weight in two estuarine clams, Meretrix meretrix and Cyclina sinensis in Shuangtaizi Estuary, Bohai Sea in China. J. Shellfish. Res. 2018, 37, 989–996. [Google Scholar] [CrossRef]
- Deng, Y.W.; Fu, S.; Du, X.D.; Wang, Q.H. Realized heritability and genetic gain estimates of larval shell length in the Chinese pearl oyster Pinctada martensii at three different salinities. N. Am. J. Aquacilt. 2009, 71, 302–306. [Google Scholar] [CrossRef]
- Ackerly, S.C. The structure of ontogenetic variation in the shell of Pecten. Palaeontology 1992, 35, 847–867. [Google Scholar]
- Minchin, D. Introductions: Some biological and ecological characteristics of scallops. Aquat. Living Resour. 2003, 16, 521–532. [Google Scholar] [CrossRef]
- Sherratt, E.; Serb, J.M.; Adams, D.C. Rates of morphological evolution, asymmetry and morphological integration of shell shape in scallops. BMC Evol. Biol. 2017, 17, 248. [Google Scholar] [CrossRef]
- Serb, J.M.; Sherratt, E.; Alejandrino, A.; Adams, D.C. Phylogenetic convergence and multiple shell shape optima for gliding scallops (Bivalvia: Pectinidae). J. Evolution. Biol. 2017, 30, 1736–1747. [Google Scholar] [CrossRef]
- Speiser, D.I.; Wilkens, L.A. Neurobiology and behaviour of the scallop. In Developments in Aquaculture and Fisheries Science; Elsevier: Amsterdam, The Netherlands, 2016; pp. 219–251. [Google Scholar] [CrossRef]
- Stanley, S.M. Adaptive morphology of the shell in bivalves and gastropods. In Form and Function; Elsevier: Amsterdam, The Netherlands, 1988; pp. 105–141. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Shell Length (mm) | Shell Height (mm) | Shell Width (g) |
|---|---|---|---|
| RS | 62.18 ± 2.86 | 57.05 ± 3.43 | 25.23 ± 1.66 |
| BS | 64.81 ± 3.08 | 58.37 ± 3.40 | 25.43 ± 1.71 |
| CG | 58.76 ± 3.83 | 53.08 ± 4.46 | 24.82 ± 1.77 |
| Discriminant Method | Samples | Number of Sample Categories | Total | Discrimination Rate (%) | ||
|---|---|---|---|---|---|---|
| BS | RS | CG | ||||
| Original discrimination | BS | 40 | 3 | 0 | 43 | 93.02 |
| RS | 1 | 41 | 0 | 42 | 97.62 | |
| CG | 0 | 0 | 44 | 44 | 100.00 | |
| Total | 41 | 44 | 44 | 129 | 96.90 | |
| Jack-knifed discrimination | BS | 29 | 13 | 1 | 43 | 67.44 |
| RS | 11 | 20 | 11 | 42 | 47.62 | |
| CG | 1 | 13 | 30 | 44 | 68.18 | |
| Total | 41 | 46 | 42 | 129 | 61.24 | |
| Discriminant Method | Samples | Number of Sample Categories | Total | Discrimination Rate (%) | ||
|---|---|---|---|---|---|---|
| BS | RS | CG | ||||
| Original discrimination | BS | 42 | 1 | 0 | 43 | 97.67 |
| RS | 0 | 42 | 0 | 42 | 100.00 | |
| CG | 0 | 1 | 43 | 44 | 97.73 | |
| Total | 42 | 43 | 44 | 129 | 98.45 | |
| Jack-knifed discrimination | BS | 27 | 12 | 4 | 43 | 62.79 |
| RS | 9 | 21 | 12 | 42 | 50.00 | |
| CG | 6 | 5 | 33 | 44 | 75.00 | |
| Total | 42 | 38 | 49 | 129 | 62.79 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, X.; Hao, Z.; Mao, J.; Wang, L.; Wang, X.; Tian, Y. Morphometric Analysis of Two Shell Color Strains of the Bay Scallop Argopecten irradians. Fishes 2024, 9, 267. https://doi.org/10.3390/fishes9070267
Huang X, Hao Z, Mao J, Wang L, Wang X, Tian Y. Morphometric Analysis of Two Shell Color Strains of the Bay Scallop Argopecten irradians. Fishes. 2024; 9(7):267. https://doi.org/10.3390/fishes9070267
Chicago/Turabian StyleHuang, Xianpeng, Zhenlin Hao, Junxia Mao, Luo Wang, Xubo Wang, and Ying Tian. 2024. "Morphometric Analysis of Two Shell Color Strains of the Bay Scallop Argopecten irradians" Fishes 9, no. 7: 267. https://doi.org/10.3390/fishes9070267
APA StyleHuang, X., Hao, Z., Mao, J., Wang, L., Wang, X., & Tian, Y. (2024). Morphometric Analysis of Two Shell Color Strains of the Bay Scallop Argopecten irradians. Fishes, 9(7), 267. https://doi.org/10.3390/fishes9070267