
Optimization of Classical Lipase Activity Assays for Fish Digestive Tract Samples

 ,
,  ,
,  , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Reagents and Materials
2.2. Fish Sample Testing
2.3. Test towards the Standardization of Lipase Methods
2.4. p-NP Substrate Method
2.5. p-NP Absorption Spectrum
2.6. p-NP Standard Curve
2.7. Effect of Type of Bile Salt
2.8. Effect of Sodium Taurocholate (NaT) Concentration
2.9. Effect of CaCl2 Concentration
2.10. β-NC Method
2.11. β-N Standard Curve
2.12. Natural Oil pH Stat Method
2.13. Oil In Vitro Digestibility
2.14. Native Lipase Electrophoretic Analysis
2.15. Statistical Analysis
3. Results
3.1. Absorption Spectrum and Standard Curve with p-NP
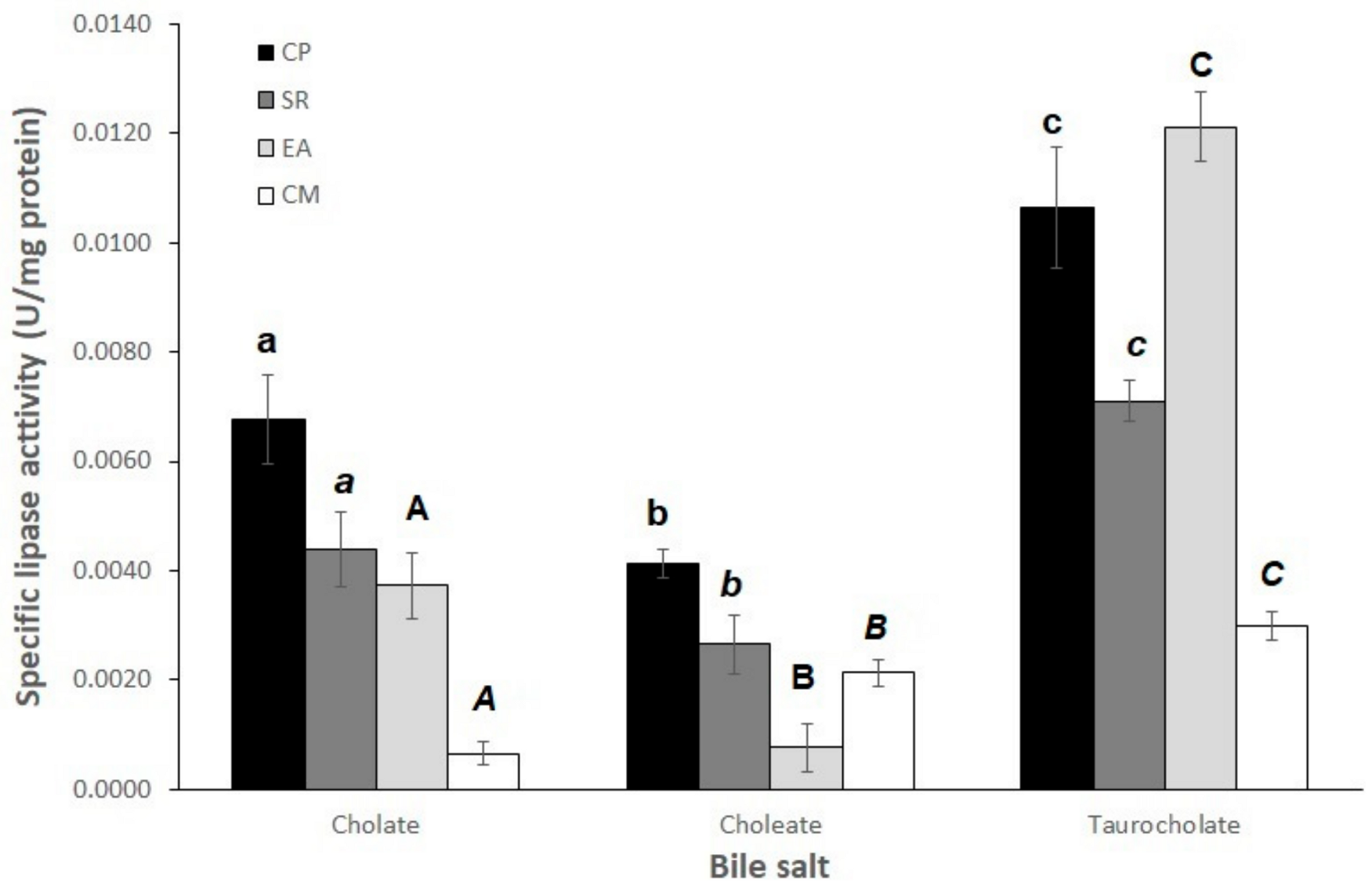
3.2. Effect of Type of Bile Salt on Lipase Activity in p-NP Substrates
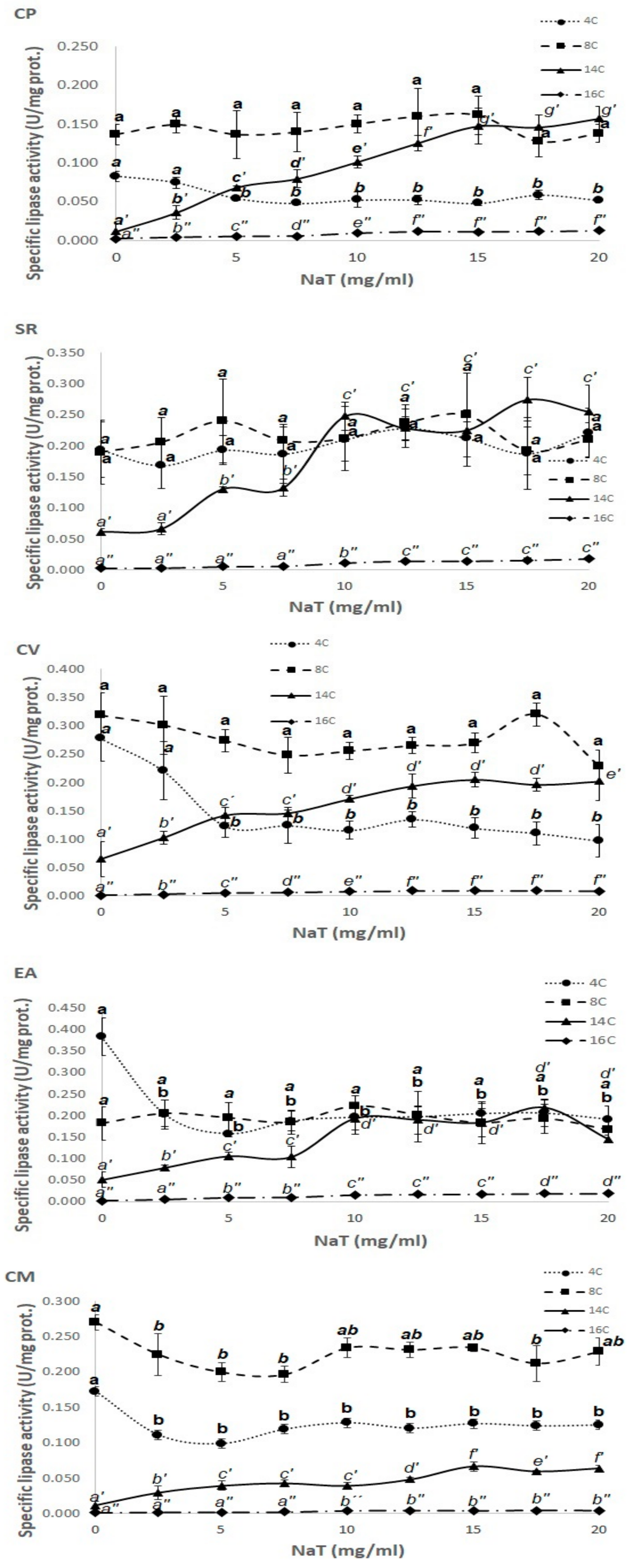
3.3. Effect of NaT Concentration on Lipase Activity in p-NP Substrates
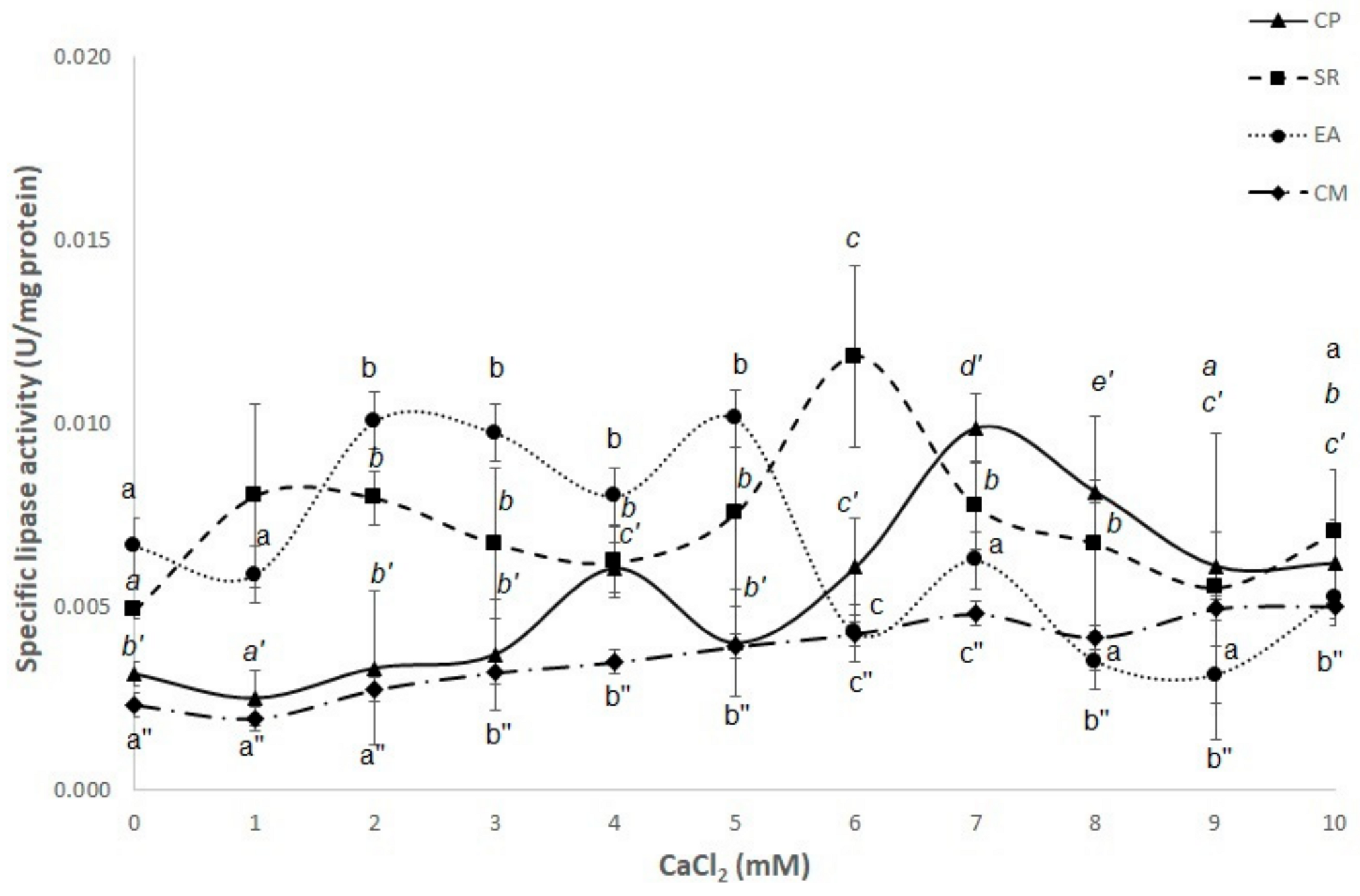
3.4. Effect of CaCl2 Concentration on Lipase Activity in p-NP Substrates
3.5. Absorption Spectrum and Standard Curve with β-N
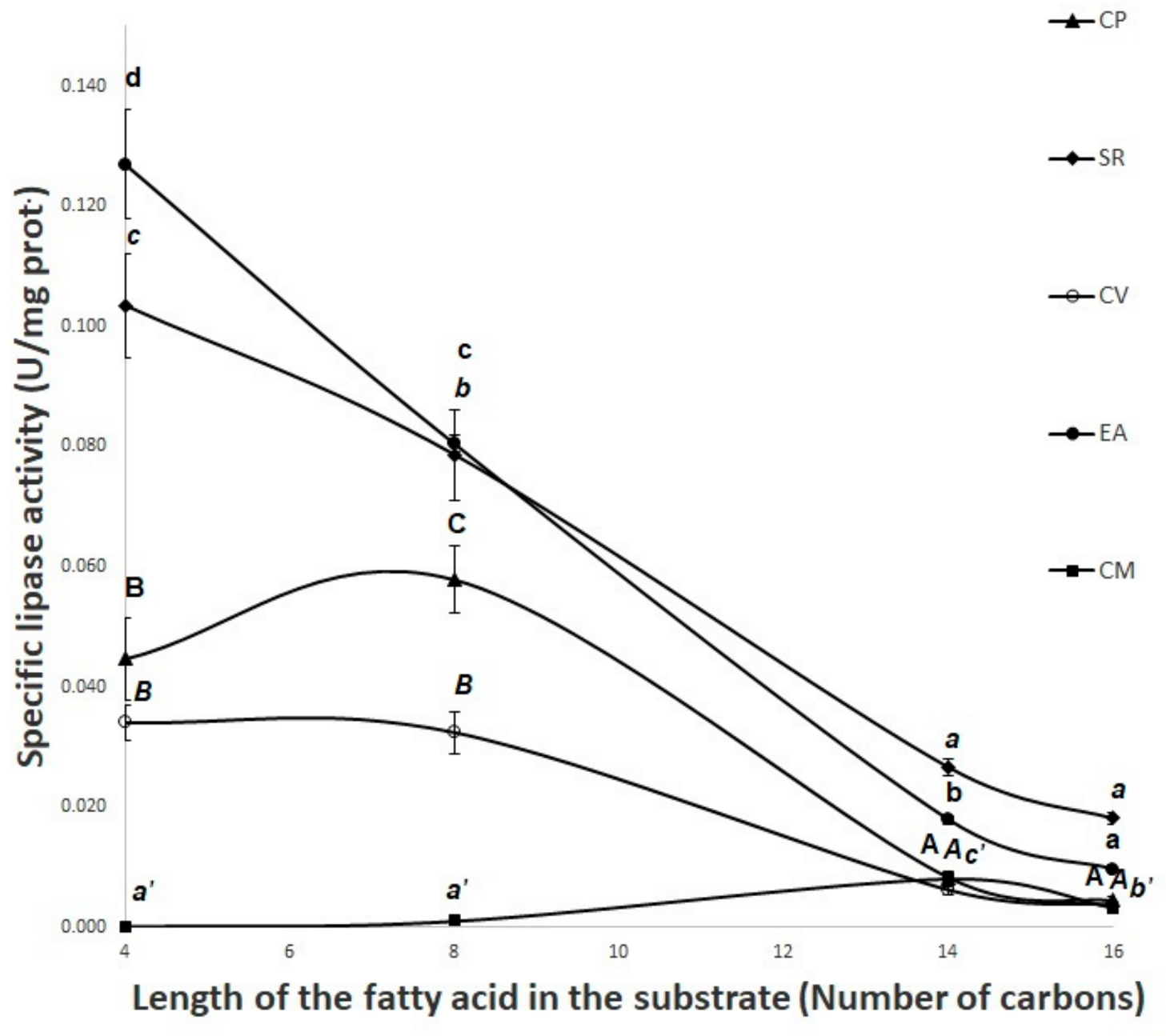
3.6. Lipase Activity Using Olive Oil as Substrate—pH Stat Method
3.7. Native Lipase Isoforms
3.8. Proposed Final Protocols for Standardization of Lipase Activity Measurement Using p-NPM, β-NC, and Natural Oil as Substrates
3.8.1. p-NPM Method
3.8.2. β-NC Method
3.8.3. Olive Oil with pH Stat Method
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Caro, J.D.; Rouimi, P.; Rovery, M. Hydrolysis of p-nitrophenyl acetate by the peptide chain fragment (336–449) of porcine pancreatic lipase. Eur. J. Biochem. 1986, 158, 601–607. [Google Scholar] [CrossRef] [PubMed]
- del Monte, A.; González-Bacerio, J.; Romero, L.; Aragón, C.; Vega, F.; Nolasco, H.; Díaz, J. Extraction systems for isolating esterases having interfacial adsorption. Rev. Colomb. Biotecnol. 2009, 11, 7–16. Available online: http://scielo.org.co/pdf/biote/v11n1/v11n1a02.pdf (accessed on 18 January 2024).
- del Monte-Martínez, A.; González-Bacerio, J.; Aragón-Abreu, C.; Palomo-Carmona, J.M.; Guisán-Seijas, J.M.; Díaz-Brito, J. Selective and oriented immobilization of (phospho)lipases from the Caribbean Sea anemone Stichodactyla helianthus (Ellis, 1768) by interfacial adsorption. Rev. CENIC Cienc. Biológicas 2012, 43, 3–8. Available online: https://doaj.org/article/5b82bf6757e54104bb9125eb1e507aae (accessed on 15 March 2024).
- del Monte-Martínez, A.; González-Bacerio, J.; Cutiño-Avila, B.; Ruiz, R.; Avila, R.; Ramos-Leal, M.; Nolasco, H.; Díaz, J.; Guisán, J.M. Esters biotransformation by immobilized interfacial esterases from the Caribbean Sea anemone Stichodactyla helianthus. Biotecnol. Apl. 2015, 32, 3201–3210. Available online: https://doaj.org/article/848e3ddee2bb40d58171eba59562930a (accessed on 15 March 2024).
- del Monte-Martínez, A.; González-Bacerio, J.; Varela, C.M.; Vega-Villasante, F.; Lalana-Rueda, R.; Nolasco, H.; Díaz, J.; Guisán, J.M. Screening and immobilization of interfacial esterases from marine invertebrates as promising biocatalyst derivatives. Appl. Biochem. Biotechnol. 2019, 189, 903–918. [Google Scholar] [CrossRef]
- Rúa, M.L.; Diaz-Maurino, T.; Fernadez, V.M.; Otero, C.; Ballesteros, A. Purification and characterization of two distinct lipases from Candida cylindracea. Biochim. Biophys. Acta 1993, 1156, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Dosanjh, N.S.; Kaur, J. Biochemical analysis of a native and proteolytic fragment of a high-molecular-weight thermostable lipase from a mesophilic Bacillus sp. Prot. Express Purif. 2002, 24, 71–75. [Google Scholar] [CrossRef]
- Yúfera, M.; Moyano, F.J.; Martínez-Rodríguez, G. The digestive function in developing fish larvae and fry. From molecular gene expression to enzymatic activity. In Emerging Issues in Fish Larvae Research; Yúfera, M., Ed.; Springer International Publishing AG: Cham, Switzerland, 2018; p. 296. [Google Scholar] [CrossRef]
- Dandavate, V.; Jinjala, J.; Keharia, H.; Madamwar, D. Production, partial purification and characterization of organic solvent tolerant lipase from Burkholderia multivorans V2 and its application for ester synthesis. Bioresour. Technol. 2009, 100, 3374–3381. [Google Scholar] [CrossRef]
- De Caro, J.D.; Chautan, M.P.; Rouimi, P.; Rovery, M. Acetylation of Lys-373 in porcine pancreatic lipase after reaction of the enzyme or its C-terminal with p-nitrophenyl acetate. Biochimie 1988, 70, 1785–1790. [Google Scholar] [CrossRef]
- Gupta, R.; Rath, P.; Gupta, N.; Bradoo, S. Lipase assays for conventional and molecular screening: An overview. Biotech. Appl. Biochem. 2003, 37, 63–71. [Google Scholar] [CrossRef]
- Lopez-Amaya, C.; Marangoni, A.G. Lipases. In Seafood Enzymes. Utilization and Influence on Postharvest Seafood Quality; Haard, N.F., Simpson, B.K., Eds.; Marcel Dekker: New York, NY, USA, 2000; pp. 121–146. [Google Scholar] [CrossRef]
- Kurtovic, I.; Marshall, S.N.; Zhao, X.; Simpson, B.K. Lipases from mammals and fishes. Rev. Fish. Sci. 2009, 17, 18–40. [Google Scholar] [CrossRef]
- Aloulou, A.; Puccinelli, D.; De Caro, A.M.; Leblond, Y.; Carriere, F. A comparative study on two fungal lipases from Thermomyces lanuginosus and Yarrowia lipolytica shows the combined effects of detergentsand pH on lipase adsorption and activity. Biochim. Biophys. Acta 2007, 1771, 1446–1456. [Google Scholar] [CrossRef] [PubMed]
- Sonesson, A.W.; Elofsson, U.M.; Brismar, H.; Callisen, T.H. Adsorption and mobility of a lipase at a hydrophobic surface in the presence of surfactants. Langmuir 2006, 22, 5810–5817. [Google Scholar] [CrossRef] [PubMed]
- Kimura, H.; Futami, Y.; Tarui, S.; Shinomiya, T. Activation of human pancreatic lipase activity by calcium and bile salts. J. Biochem. 1982, 92, 243–251. [Google Scholar] [CrossRef]
- Larsson, A.; Erlanson-Albertsson, C. The importance of bile salt for the reactivation of pancreatic lipase by colipase. Biochim. Biophys. Acta 1983, 750, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Barnescu, R.; Serban, M.; Rugina, A.; Crisan, I.; Cepisca, C.; Caloianu, M. Biochemical characteristics in heterogeneous catalysis correlated with bioactive effect of the extracted components. Rom. J. Biol. Sci. 1997, 1–2, 84–89. [Google Scholar]
- Wu, X.Y.; Jaaskelainen, S.; Linko, Y.Y. Purification and partial characterization of Rhizomucor miehei lipase for ester synthesis. Appl. Biochem. Biotech. 1996, 59, 145–158. [Google Scholar] [CrossRef]
- Palomo, J.M.; Fuentes, M.; Fernandez-Lorente, G.; Mateo, C.; Guisan, J.M.; Fernandez-Lafuente, R. General trend of lipase to self-assemble giving bimolecular aggregates greatly modifies the enzyme functionality. Biomacromolecules 2003, 4, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Guisan, J.M.; Fernandez-Lafuente, R.; Bastida, A.; Blanco, R.M.; Soler, G.; Garcia-Junceda, E. Modulation of activity/stability properties of lipase from Pseudomonas flourescens by multipoint covalent immobilization on glyoxyl-supports. In Engineering of/with Lipases; Malcata, F.X., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1996; pp. 243–256. Available online: http://tdl.libra.titech.ac.jp/journaldocs/recordID/article.bib-01/ZR000000015830 (accessed on 3 February 2024).
- Sharma, A.K.; Tiwari, R.P.; Hoondal, G.S. Properties of a thermostable and solvent stable extracellular lipase from a Pseudomonas sp. AG-8. Biotechnol. Bioeng. 2002, 77, 693–703. [Google Scholar] [CrossRef]
- Nolasco-Soria, H.; Moyano-López, F.; Vega-Villasante, F.; Del Monte, A.; Espinoza-Chaurand, D.; Gisbert, E. Lipase and Phospholipase Activity Methods for Marine Organisms. In Lipases and Phospholipases: Methods and Protocols; Methods in Molecular Biology; Sandoval, G., Ed.; Humana Press: New York, NY, USA, 2018; Chapter 7; Volume 1835. [Google Scholar] [CrossRef]
- Nolasco-Soria, H. Fish digestive lipase quantification methods used in aquaculture studies. Front. Aquac. 2023, 2, 21225216. [Google Scholar] [CrossRef]
- Bornscheuer, U.; Reif, O.W.; Lausch, R.; Freitag, R.; Scheper, T.; Kolisis, F.N.; Menge, U. Lipase of Pseudomonas cepacia for biotechnological purposes: Purification, crystallization and characterization. Biochim. Biophys. Acta 1994, 1201, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Zeng, Q.M.; Zong, M.H. Substrate specificity of lipase from Burkholderia cepacia in the synthesis of 3-arylaliphatic acid esters of floxuridine. J. Biotechnol. 2009, 142, 267–270. [Google Scholar] [CrossRef] [PubMed]
- Berner, D.L.; Hammond, E.G. Phylogeny of lipase specificity. Lipids 1970, 5, 558–562. [Google Scholar] [CrossRef] [PubMed]
- Snellman, E.A.; Sullivan, E.R.; Colwell, R.R. Purification and properties of the extracellular lipase, LipA, of Acinetobacter sp. RAG-1. Eur. J. Biochem. 2002, 269, 5771–5779. [Google Scholar] [CrossRef] [PubMed]
- Zouari, N.; Miled, N.; Cherif, S.; Mejdoub, H.; Gargouri, Y. Purification and characterization of a novel lipase from the digestive glands of a primitive animal: The scorpion. Biochim. Biophys. Acta 2005, 1726, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Cygler, M.; Grochulski, P.; Schrag, J.D. Structural determinants defining common stereoselectivity of lipases toward secondary alcohols. Can. J. Microb. 1995, 41, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Bornscheuer, U.T. Methods to increase enantioselectivity of lipases and esterases. Curr. Opin. Biotech. 2002, 13, 543–547. [Google Scholar] [CrossRef]
- Jensen, R.G.; De Jong, F.A.; Lambert-Davis, L.G.; Hamosh, M. Fatty acid and positional selectivities of gastric lipase from premature human infants, in vitro studies. Lipids 1994, 29, 433–435. [Google Scholar] [CrossRef] [PubMed]
- González-Bacerio, J.; Rodríguez, J.; del Monte, A. Lipases: Enzymes having the potential for developing immobilised biocatalysts by interfacial adsorption. Rev. Colomb. Biotecnol. 2010, 12, 124–140. Available online: http://www.scielo.org.co/scielo.php?pid=S0123-34752010000100013&script=sci_abstract#:~:text=Lipases%3A%20enzymes%20having%20the%20potential%20for%20developing%20immobilised,colomb.%20biotecnol%5Bonline%5D.%202010%2C%20vol.12%2C%20n.1%2C%20pp.113-140.%20ISSN%200123-3475 (accessed on 15 March 2024).
- Iijima, N.; Tanaka, S.; Ota, Y. Purification and characterization of bile salt-activated lipase from the hepatopancreas of red sea bream, Pagrus major. Fish. Physiol. Biochem. 1998, 18, 59–69. [Google Scholar] [CrossRef]
- Versaw, W.K.; Cuppett, S.L.; Winters, D.D.; Williams, L.E. An improved colorimetric assay for bacterial lipase in nonfat dry milk. J. Food Sci. 1989, 54, 1557–1558. [Google Scholar] [CrossRef]
- Bier, M. Lipases. In Methods in Enzymology; Colowick, S.P., Kaplan, N.O., Eds.; Academic Press: New York, NY, USA, 1955; Volume 1, pp. 627–642. [Google Scholar]
- Bertolini, M.C.; Schrag, J.D.; Cygler, M.; Ziomek, E.; Thomas, D.Y.; Vernet, T. Expression and Characterization of Geotrichum candidum Lipase I Gene. FEBS J. 1995, 228, 863–869. [Google Scholar] [CrossRef]
- Sastry, K.V.; Gupta, P.K. In vitro inhibition of digestive enzymes by heavy metals and their reversal by chelating agent: Part I. Mercuric chloride intoxication. Bull. Environ. Contam. Toxicol. 1978, 20, 729–735. [Google Scholar] [CrossRef] [PubMed]
- Tyndall, J.D.; Sinchaikul, S.; Fothergill-Gilmore, L.A.; Taylor, P.; Walkinshaw, M.D. Crystal structure of a thermostable lipase from Bacillus stearothermophilus P1. J. Mol. Biol. 2002, 323, 859–869. [Google Scholar] [CrossRef] [PubMed]
- Amada, K.; Kwon, H.J.; Haruki, M.; Morikawa, M.; Kanaya, S. Ca2+-induced folding of a family I.3 lipase with repetitive Ca2+ binding motifs at the C-terminus. FEBS Lett. 2001, 509, 17–21. [Google Scholar] [CrossRef] [PubMed]
- Egmond, M.R. Action of lipases. In Engineering of/with Lipases; Malcata, F.X., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1996; pp. 183–191. Available online: https://api.semanticscholar.org/CorpusID:99992337 (accessed on 20 February 2024).
- Tsai, S.W.; Lu, C.C.; Chang, C.S. Surfactant enhancement of (S)-naproxen ester productivity from racemic naproxen by lipase in isooctane. Biotech. Bioeng. 1996, 51, 148–156. [Google Scholar] [CrossRef]
- Solovyev, M.M.; Izvekova, G.I. Seasonal changes in pH values in the intestine of fish from Lake Chany (West Siberia). Inland Water Biol. 2016, 9, 400–404. [Google Scholar] [CrossRef]
- Gisbert, E.; Nolasco, H.; Solovyev, M. Towards the standardization of brush border purification and intestinal alkaline phosphatase quantification in fish with notes on other digestive enzymes. Aquaculture 2018, 487, 102–108. [Google Scholar] [CrossRef]
- Nolasco-Soria, H. Improving and standardizing protocols for alkaline protease quantification in fish. Rev. Aquac. 2020, 13, 43–65. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Nolasco-Soria, H. Amylase quantification in aquaculture fish studies: A revision of most used procedures and presentation of a new practical protocol for its assessment. Aquaculture 2021, 538, 736536. [Google Scholar] [CrossRef]
- Meier, A.R.; Yehl, J.B.; Eckenroad, K.W.; Manley, G.A.; Strein, T.G.; Rovnyak, D. Stepwise aggregation of cholate and deoxycholate dictates the formation and loss of surface-available chirally selective binding sites. Langmuir 2018, 34, 6489–6501. [Google Scholar] [CrossRef] [PubMed]
- Campanelli, A.R.; Candeloro De Sanctis, F.; D’Archivio, A.A.; Giglio, E.; Scarammuzza, L. Crystal structures of bile salts: Sodium taurocholate. J. Incl. Phenom. Macrocycl. Chem. 1991, 11, 247–256. [Google Scholar] [CrossRef]
- Cherry, I.S.; Crandall, L.A. The specificity of pancreatic lipase: Its appearance in the blood after pancreatic injury. Am. J. Physiol. 1932, 100, 266–273. [Google Scholar] [CrossRef]
- Nolasco, H. Métodos Utilizados por el Centro de Investigaciones Biológicas del Noroeste (CIBNOR) para la Medición de Digestibilidad in vitro para Camarón. In Manual de Metodologías de Digestibilidad In Vivo e In Vitro Para Ingredientes y Dietas Para Camarón; Cruz Suárez, L.E., Villarreal Colmenares, H., Salazar, M.T., Nieto López, M.G., Villarreal Cavazos, D.A., Ricque Marie, D., Eds.; Universidad Autónoma de Nuevo León: Monterrey, Mexico, 2008; pp. 215–225. Available online: https://nutricionacuicola.uanl.mx/public/site/images/admin/manual_metodologias.pdf (accessed on 9 January 2024).
- Rasco, B.A.; Hultin, H.O. A comparison of dogfish and porcine pancreatic lipases. Comp. Biochem. Physiol. B 1988, 89, 671–677. [Google Scholar] [CrossRef] [PubMed]
- Løkka, G.; Dhanasiri, A.K.S.; Krogdahl, A.; Kortner, T.M. Bile components affect the functions and transcriptome of the rainbow trout intestinal epithelial cell line RTgutGC. Fish. Shellfish. Immun. 2022, 131, 1144–1156. [Google Scholar] [CrossRef]
- Gao, Y.; Yao, Y.; Huang, J.; Sun, Y.; Wu, Q.; Guo, D.; Wang, S. Effect of dietary bile acids supplementation on growth performance, feed utilization, intestinal digestive enzyme activity and fatty acid transporters gene expression in juvenile leopard coral grouper (Plectropomus leopardus). Front. Mar. Sci. 2023, 10, 1171344. [Google Scholar] [CrossRef]
- Hui, D.Y.; Howles, P.N. Carboxyl ester lipase: Structure-function relationship and physiological role in lipoprotein metabolism and atherosclerosis. J. Lipid Res. 2002, 43, 2017–2030. [Google Scholar] [CrossRef]
- Leger, C.; Bauchart, D.; Flanzy, J. Some properties of pancreatic lipase in Salmo gairdnerii Rich.: Km, effects of bile salts and Ca2+, gel filtrations. Comp. Biochem. Physiol. B 1977, 57, 359–363. [Google Scholar] [CrossRef]
- Ruiz, A.; Andree, K.B.; Furones, D.; Holhorea, P.G.; Calduch-Giner, J.À.; Viñas, M.; Pérez-Sánchez, J.; Gisbert, E. Modulation of gut microbiota and intestinal immunenresponse in gilthead seabream (Sparus aurata) by dietary bile salt supplementation. Front. Microbiol. 2023, 14, 1123716. [Google Scholar] [CrossRef]
- Romano, N.; Kumar, V.; Yang, G.; Kajbaf, K.; Rubio, M.B.; Overturf, K.; Brezas, A.; Hardy, R. Bile acid metabolism in fish: Disturbances caused by fishmeal alternatives and some mitigating effects from dietary bile inclusions. Rev. Aquac. 2020, 12, 1792–1817. [Google Scholar] [CrossRef]
- Olsen, R.E.; Henderson, R.J.; Ringo, E. The digestion and selective absorption of dietary fatty acids in Arctic charr, Salvelinus alpinus. Aquac. Nutr. 1998, 4, 13–21. [Google Scholar] [CrossRef]
- Hu, M.; Li, Y.; Decker, E.A.; McClements, D.J. Role of calcium and calcium-binding agents on the lipase digestibility of emulsified lipids using an in vitro digestion model. Food Hydrocoll. 2010, 24, 719–725. [Google Scholar] [CrossRef]
- Kurtovic, I.; Marshall, S.N.; Zhao, X.; Simpson, B.K. Purification and properties of digestive lipases from Chinook salmon (Oncorhynchus tshawytscha) and New Zealand hoki (Macruronus novaezelandiae). Fish. Physiol. Biochem. 2010, 36, 1041–1060. [Google Scholar] [CrossRef] [PubMed]
- Beisson, F.; Tiss, A.; Riviere, C.; Verger, R. Methods for lipase detection and assay: A critical review. Eur. J. Lipid Sci. Technol. 2000, 102, 133–153. [Google Scholar] [CrossRef]
- Tibbetts, S.M.; Scaife, M.A.; Armenta, R.E. Apparent digestibility of proximate nutrients, energy and fatty acids in nutritionally-balanced diets with partial or complete replacement of dietary fish oil with microbial oil from a novel Schizochytrium sp. (T18) by juvenile Atlantic salmon (Salmo salar L.). Aquaculture 2020, 520, 735003. [Google Scholar] [CrossRef]
- Taniguchi, A.; Takano, K.; Kamoi, I. Purification and properties of lipase from Tilapia intestine-digestive enzyme of Tilapia-VI. Nippon Suisan Gakkaishi 2001, 67, 78–84. [Google Scholar] [CrossRef]
- Rueda-Lopez, S.; Martinez-Montano, E.; Viana, M.T. Biochemical characterization and comparison of pancreatic lipases from the Pacific bluefin tuna, Thunnus orientalis; totoaba, Totoaba macdonaldi; and striped bass, Morone saxatilis. J. World Aquac. Soc. 2017, 48, 156–165. [Google Scholar] [CrossRef]
- Espada, J.; Horobin, R.W.; Stockert, J.C. Fluorescent cytochemistry of acid phosphatase and demonstration of fluid-phase endocytosis using an azo dye method. Histochem. Cell Biol. 1997, 108, 481–487. [Google Scholar] [CrossRef]
- Mukundan, M.K.; Gopakumar, K.; Nair, M.R. Purification of a lipase from the hepatopancreas of oil sardine (Sardinella longiceps Linnaeus) and its characteristics and properties. J. Sci. Food Agric. 1985, 36, 191–203. [Google Scholar] [CrossRef]
- Borlongan, I.G. Studies on the digestive lipases of milkfish, Chanos chanos. Aquaculture 1990, 89, 315–325. [Google Scholar] [CrossRef]
- Gjellesvik, D.R.; Lombardo, D.; Walther, B.T. Pancreatic bile salt dependent lipase from cod (Gadus morhua): Purification and properties. Biochim. Biophys. Acta Lipids Lipid Metab. 1992, 1124, 123–134. [Google Scholar] [CrossRef] [PubMed]
- Concha-Frías, B.; Gaxiola-Cortes, M.G.; De la Cruz-Alvarado, F.J.; Jimenez Martinez, L.D.; Peña-Marin, E.S.; Oliva-Arriagada, M.A.; Arias-Moscoso, J.L.; Alvarez-González, C.A. Intestinal Lipase Characterization in Common Snook (Centropomus undecimalis) Juveniles. Fishes 2022, 7, 107. [Google Scholar] [CrossRef]
- Kameshwar Sharma, Y.V.R.; Neelima, B.; Prasidhi, T. Isolation, purification and characterization of secondary structure and kinetic study of lipase from Indian major carp, Catla catla (Catla). Enzym. Eng. 2014, 3, 121. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fish | Average Weight (g) | Digestive Tract Somatic Index | CP-I or I Somatic Index | Soluble Protein Concentration (mg/mL) |
|---|---|---|---|---|
| CP | 442 ± 26.7 | 0.058 ± 0.011 | 0.011 ± 0.003 | 5.70 ± 2.0 |
| SR | 857 ± 3.6 | 0.039 ± 0.002 | 0.008 ± 0.000 | 5.28 ± 0.1 |
| CV | 221.3 ± 1.5 | 0.045 ± 0.001 | 0.008 ± 0.000 | 10.04 ± 0.4 |
| EA | 761.0 ± 85.7 | 0.042 ± 0.004 | 0.004 ± 0.001 | 4.77 ± 0.2 |
| CM | 820.7 ± 90.7 | 0.060 ± 0.020 | 0.024 ± 0.007 | 7.79 ± 0.6 |
| Iijima [34] | Optimized Method | Versaw [35] | Optimized Method | Bier [36] | Optimized Method | |
|---|---|---|---|---|---|---|
| Total assay volume (μL) | 1200 | 200 | 5000 | 200 | 5000 | 5000 |
| Assay device | Tube | Microplate well | Tube | Microplate well | 125 mL flask | 100 mL vessel |
| Substrate | p-NPM | p-NPM | β-NC | β-NC | Olive oil | Olive oil |
| Substrate concentration (mM) | 0.53 | 0.50 | 1.9 | 0.5 | 100 | 17.4 |
| Solvent | 2-Methoxyethanol | DMSO | DMSO | DMSO | Polyvinyl alcohol | Water tauracholate |
| Solvent concentration (mM) | 0.25 | 705 | 136.2 | 705 | 10 ** | 23.34 |
| Bile salt | Sodium cholate | Sodium tauracholate | Sodium tauracholate | Sodium tauracholate | Not indicated | Sodium tauracholate |
| Bile salt concentration (mM) | 5 | 18.6 | 19.3 | 2.8 | 0 | 23.34 |
| pH | 9.0 | 8.0 | 7.2 | 8 | Not indicated | 8 |
| CE (μL) | 5 or 10 | 10 | 50 | 10 | 5000 | 100 |
| Incubation temperature (°C) | 30 | 25 | 40 | 25 | 37 | 25 |
| Incubation time (min) | 15 | 5 | 30 | 10 | 240 | 15 |
| Stopper | Acetone: Heptane (5:2 v/v) | Not required (Kinetic method) | 0.72 N TCA | 2% TCA-12% SDS | Ethyl alcohol/ acetone (1:1 v/v) | Not required (Kinetic method) |
| Classification process | Centrifugation at 6080× g for 2 min | Not required | Ethanol (95%)/ethyl acetate (1:1 v/v) | * | Not required | Not required |
| Absorbance zone | Lower aqueous layer | Full reaction mixture | Sample of reaction mixture | Full reaction mixture | Not required | Not required |
| Absorbance wavelength (nm) | 405 | 400 | 540 | 540 | Not required | Not required |
| ε | 16,500 (pH 9) | 16,876 (pH 8) | Not indicated | 24,286 (pH 8.0) | Not required | Not required |
| Unit definition | One μmol of p-NP release per min | One μmol of p-NP release per min | Not indicated | One μmol of β-N release per min | Not indicated | One μmol of fatty acid (proton) release per min |
| Form for unit calculation | Not indicated | Indicated | Not indicated | Indicated | Not indicated | Indicated |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nolasco-Soria, H.; Alvarez-González, C.A.; Tovar-Ramírez, D.; González-Bacerio, J.; del Monte-Martínez, A.; Vega-Villasante, F. Optimization of Classical Lipase Activity Assays for Fish Digestive Tract Samples. Fishes 2024, 9, 261. https://doi.org/10.3390/fishes9070261
Nolasco-Soria H, Alvarez-González CA, Tovar-Ramírez D, González-Bacerio J, del Monte-Martínez A, Vega-Villasante F. Optimization of Classical Lipase Activity Assays for Fish Digestive Tract Samples. Fishes. 2024; 9(7):261. https://doi.org/10.3390/fishes9070261
Chicago/Turabian StyleNolasco-Soria, Héctor, Carlos Alfonso Alvarez-González, Dariel Tovar-Ramírez, Jorge González-Bacerio, Alberto del Monte-Martínez, and Fernando Vega-Villasante. 2024. "Optimization of Classical Lipase Activity Assays for Fish Digestive Tract Samples" Fishes 9, no. 7: 261. https://doi.org/10.3390/fishes9070261
APA StyleNolasco-Soria, H., Alvarez-González, C. A., Tovar-Ramírez, D., González-Bacerio, J., del Monte-Martínez, A., & Vega-Villasante, F. (2024). Optimization of Classical Lipase Activity Assays for Fish Digestive Tract Samples. Fishes, 9(7), 261. https://doi.org/10.3390/fishes9070261